Фотосинтез


Фотосинтез ( / ˌ foʊ t ə ˈ s sis ɪ n θ ə s ɪ s / FOH -tə- SINTH -ə- ) [ 1 ] — это система биологических процессов , с помощью которых фотосинтезирующие организмы , такие как большинство растений, водорослей и цианобактерий , преобразуют световую энергию , обычно получаемую от солнечного света, в химическую энергию, необходимую для поддержания их метаболизма .
Фотосинтез обычно относится к оксигенному фотосинтезу , процессу, в ходе которого образуется кислород.
Фотосинтезирующие организмы хранят вырабатываемую таким образом химическую энергию во внутриклеточных органических соединениях (соединениях, содержащих углерод), таких как сахара, гликоген , целлюлоза и крахмалы .
Чтобы использовать эту накопленную химическую энергию, клетки организма метаболизируют органические соединения посредством клеточного дыхания . Фотосинтез играет решающую роль в производстве и поддержании содержания кислорода в атмосфере Земли и обеспечивает большую часть биологической энергии, необходимой для сложной жизни на Земле. [ 2 ]
Некоторые бактерии также осуществляют аноксигенный фотосинтез , при котором бактериохлорофилл используется для расщепления сероводорода в качестве восстановителя вместо воды, производя серу вместо кислорода. Археи , такие как Halobacterium, также осуществляют тип аноксигенного фотосинтеза, не связывающего углерод , при котором более простой фотопигмент ретиналь и его родопсина микробные производные используются для поглощения зеленого света и приводят в действие протонные насосы для прямого синтеза аденозинтрифосфата (АТФ), «энергетической валюты». "клеток. Такой архейный фотосинтез мог быть самой ранней формой фотосинтеза, развившейся на Земле еще в палеоархее , предшествовавшей фотосинтезу цианобактерий (см. Гипотезу Пурпурной Земли ).
Хотя детали могут различаться у разных видов , процесс всегда начинается, когда световая энергия поглощается реакционными центрами , белками, которые содержат фотосинтетические пигменты или хромофоры . У растений эти белки представляют собой хлорофиллы ( производное порфирина , которое поглощает красный и синий спектры света, отражая тем самым зеленый цвет), содержащиеся внутри хлоропластов и изобилующих клетками листьев . У бактерий они встроены в плазматическую мембрану . В этих светозависимых реакциях некоторая энергия используется для отрыва электронов от подходящих веществ, таких как вода, с образованием газообразного кислорода. Водород , высвобождаемый при расщеплении воды, используется для создания двух важных молекул, участвующих в энергетических процессах: восстановленного никотинамидадениндинуклеотидфосфата (НАДФН) и АТФ.
В растениях, водорослях и цианобактериях сахара синтезируются посредством последующей последовательности светонезависимых реакций, называемых циклом Кальвина . В этом процессе углекислый газ из атмосферы включается в уже существующие органические соединения, такие как рибулозобисфосфат (RuBP). [ 3 ] Используя АТФ и НАДФН, образующиеся в результате светозависимых реакций, полученные соединения затем восстанавливаются и удаляются с образованием дополнительных углеводов, таких как глюкоза . У других бактерий другие механизмы, такие как обратный цикл Кребса для достижения той же цели используются .
Первые фотосинтезирующие организмы, вероятно, возникли на ранних этапах эволюционной истории жизни, используя в качестве источников электронов восстановители , такие как водород или сероводород, а не воду. [ 4 ] Цианобактерии появились позже; избыток кислорода, который они производили, непосредственно способствовал насыщению Земли кислородом , [ 5 ] что сделало возможной эволюцию сложной жизни. Средняя мощность энергии, улавливаемой глобальным фотосинтезом, составляет примерно 130 тераватт . [ 6 ] [ 7 ] [ 8 ] что примерно в восемь раз превышает общее энергопотребление человеческой цивилизации . [ 9 ] Фотосинтезирующие организмы также преобразуют около 100–115 миллиардов тонн Пг (91–104 петаграмм , или миллиард метрических тонн) углерода в биомассу в год. [ 10 ] [ 11 ] Фотосинтез был открыт в 1779 году Яном Ингенхаузом . Он показал, что растениям нужен свет, а не только воздух, почва и вода.
Фотосинтез жизненно важен для климатических процессов, поскольку он улавливает углекислый газ из воздуха и связывает его с растениями, собранными продуктами и почвой. По оценкам, только зерновые связывают 3825 Тг или 3,825 Пг углекислого газа, т.е. 3,825 миллиарда метрических тонн. ежегодно [ 12 ]
Обзор

Большинство фотосинтезирующих организмов являются фотоавтотрофами , что означает, что они способны синтезировать пищу непосредственно из углекислого газа и воды, используя энергию света. Однако не все организмы используют углекислый газ в качестве источника атомов углерода для осуществления фотосинтеза; фотогетеротрофы используют в качестве источника углерода органические соединения, а не углекислый газ. [ 2 ]
У растений, водорослей и цианобактерий фотосинтез выделяет кислород. Этот кислородный фотосинтез на сегодняшний день является наиболее распространенным типом фотосинтеза, используемым живыми организмами. Некоторые тенелюбивые растения (сциофиты) при фотосинтезе производят настолько мало кислорода, что используют весь его сами, вместо того, чтобы выделять его в атмосферу. [ 13 ]
Хотя существуют некоторые различия между кислородным фотосинтезом у растений , водорослей и цианобактерий , общий процесс у этих организмов весьма схож. Существует также множество разновидностей аноксигенного фотосинтеза , используемого в основном бактериями, которые потребляют углекислый газ, но не выделяют кислород. [ 14 ] [ 15 ]
Углекислый газ превращается в сахара в процессе, называемом фиксацией углерода ; фотосинтез улавливает энергию солнечного света для преобразования углекислого газа в углеводы . Фиксация углерода представляет собой эндотермическую окислительно-восстановительную реакцию. В общих чертах фотосинтез является противоположностью клеточного дыхания : в то время как фотосинтез представляет собой процесс восстановления углекислого газа до углеводов, клеточное дыхание представляет собой окисление углеводов или других питательных веществ до углекислого газа. Питательные вещества, используемые в клеточном дыхании, включают углеводы, аминокислоты и жирные кислоты. организма Эти питательные вещества окисляются с образованием углекислого газа и воды, а также с выделением химической энергии, необходимой для метаболизма .
Фотосинтез и клеточное дыхание — это разные процессы, поскольку они происходят посредством разных последовательностей химических реакций и в разных клеточных компартментах (клеточное дыхание в митохондриях ). [ 16 ] [ 17 ]
Общее уравнение фотосинтеза, впервые предложенное Корнелисом ван Нилем, выглядит так: [ 18 ]
- + + → + +
Поскольку вода используется в качестве донора электронов при оксигенном фотосинтезе, уравнение этого процесса имеет вид:
- + + → + +
Это уравнение подчеркивает, что вода является одновременно реагентом в светозависимой реакции и продуктом светонезависимой реакции , но сокращение n молекул воды с каждой стороны дает чистое уравнение:
- + + → +
Другие процессы заменяют воду другими соединениями (такими как арсенит ) в качестве источника электронов; например, некоторые микробы используют солнечный свет для окисления арсенита в арсенат : [ 19 ] Уравнение этой реакции:
- + + → + (используется для создания других соединений в последующих реакциях) [ 20 ]
Фотосинтез происходит в два этапа. На первом этапе светозависимые реакции или световые реакции улавливают энергию света и используют ее для создания переносчика водорода НАДФН и молекулы-хранилища энергии АТФ . На втором этапе светонезависимые реакции используют эти продукты для улавливания и восстановления углекислого газа.
Большинство организмов, использующих кислородный фотосинтез, используют видимый свет для светозависимых реакций, хотя по крайней мере три используют коротковолновое инфракрасное или, точнее, дальнее красное излучение. [ 21 ]
Некоторые организмы используют еще более радикальные варианты фотосинтеза. Некоторые археи используют более простой метод, в котором используется пигмент, аналогичный тем, которые используются для зрения у животных. Бактериородопсин меняет свою конфигурацию в ответ на солнечный свет, действуя как протонный насос. Это создает более непосредственный градиент протонов, который затем преобразуется в химическую энергию. Этот процесс не предполагает фиксации углекислого газа и не выделяет кислород и, по-видимому, развился отдельно от более распространенных типов фотосинтеза. [ 22 ]
Фотосинтетические мембраны и органеллы

- внешняя мембрана
- межмембранное пространство
- внутренняя мембрана (1+2+3: оболочка)
- строма (водянистая жидкость)
- просвет тилакоида (внутри тилакоида)
- тилакоидная мембрана
- гранум (стопка тилакоидов)
- тилакоид (ламелла)
- крахмал
- рибосома
- пластидная ДНК
- пластоглобула (капля липидов)
У фотосинтезирующих бактерий белки, собирающие свет для фотосинтеза, встроены в клеточные мембраны . В простейшей форме это касается мембраны, окружающей саму клетку. [ 23 ] Однако мембрана может быть плотно свернута в цилиндрические листы, называемые тилакоидами . [ 24 ] или собраны в круглые пузырьки, называемые интрацитоплазматическими мембранами . [ 25 ] Эти структуры могут заполнить большую часть внутренней части клетки, придавая мембране очень большую площадь поверхности и, следовательно, увеличивая количество света, которое могут поглощать бактерии. [ 24 ]
У растений и водорослей фотосинтез происходит в органеллах, называемых хлоропластами . Типичная растительная клетка содержит от 10 до 100 хлоропластов. Хлоропласт окружен мембраной. Эта мембрана состоит из фосфолипидной внутренней мембраны, фосфолипидной внешней мембраны и межмембранного пространства. Оболочкой окружена водная жидкость, называемая стромой. В строму встроены стопки тилакоидов (гран), которые являются местом фотосинтеза. Тилакоиды выглядят как уплощенные диски. Сам тилакоид окружен тилакоидной мембраной, а внутри замкнутого объема имеется просвет или тилакоидное пространство. В тилакоидную мембрану встроены интегральные и периферические мембранные белковые комплексы фотосинтетической системы.
Растения поглощают свет преимущественно с помощью пигмента хлорофилла . Зеленая часть спектра света не поглощается, а отражается, поэтому большинство растений имеют зеленый цвет. Помимо хлорофилла, растения также используют пигменты, такие как каротины и ксантофиллы . [ 26 ] Водоросли также используют хлорофилл, но присутствуют и другие пигменты, такие как фикоцианин , каротины и ксантофиллы в зеленых водорослях , фикоэритрин в красных водорослях (родофитах) и фукоксантин в бурых водорослях и диатомовых водорослях , что приводит к широкому разнообразию цветов.
Эти пигменты встроены в растения и водоросли в комплексы, называемые антенными белками. В таких белках пигменты работают вместе. Такое сочетание белков еще называют светособирающим комплексом . [ 27 ]
Хотя все клетки зеленых частей растения имеют хлоропласты, большинство из них находится в специально приспособленных структурах, называемых листьями . Некоторые виды, адаптированные к условиям сильного солнечного света и засушливости , например, многие виды молочайов и кактусов , имеют основные фотосинтетические органы в стеблях. Клетки внутренних тканей листа, называемые мезофиллом , могут содержать от 450 000 до 800 000 хлоропластов на каждый квадратный миллиметр листа. Поверхность листа покрыта водостойкой восковой кутикулой , которая защищает лист от чрезмерного испарения воды и уменьшает поглощение ультрафиолетового или синего света , чтобы свести к минимуму нагревание . Прозрачный слой эпидермиса позволяет свету проходить к клеткам палисадного мезофилла, где происходит большая часть фотосинтеза.
Светозависимые реакции
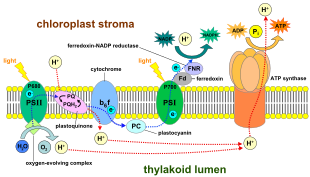
В светозависимых реакциях одна молекула пигмента хлорофилла поглощает один фотон и теряет один электрон . Этот электрон захватывается модифицированной формой хлорофилла, называемой феофитином , которая передает электрон молекуле хинона поток электронов по цепи переноса электронов , что приводит к окончательному восстановлению НАДФ , запуская до НАДФН . Кроме того, это создает протонный градиент (градиент энергии) на мембране хлоропласта , который используется АТФ-синтазой в синтезе АТФ . Молекула хлорофилла в конечном итоге восстанавливает электрон, который она потеряла, когда молекула воды расщепляется в процессе, называемом фотолизом , в результате которого выделяется кислород .
Общее уравнение светозависимых реакций в условиях нециклического потока электронов в зеленых растениях имеет вид: [ 28 ]
Не все длины волн могут света поддерживать фотосинтез. Спектр фотосинтетического действия зависит от типа присутствующих дополнительных пигментов . Например, у зеленых растений спектр действия напоминает спектр поглощения хлорофиллов . и каротиноидов с пиками поглощения в фиолетово-синем и красном свете водорослям использовать Спектр действия красных водорослей представляет собой сине-зеленый свет, что позволяет этим синий конец спектра для роста в более глубоких водах, которые отфильтровывают более длинные волны (красный свет), используемые надземными зелеными растениями. Непоглощенная часть светового спектра придает организмам цвет фотосинтезирующим (например, зеленым растениям, красным водорослям, пурпурным бактериям ) и наименее эффективна для фотосинтеза в соответствующих организмах .
Z-схема
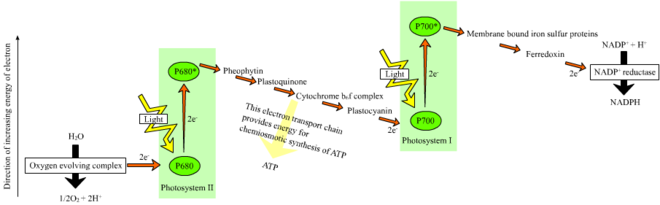
У растений , светозависимые реакции в тилакоидных мембранах хлоропластов НАДФН где они управляют синтезом АТФ и происходят . Светозависимые реакции имеют две формы: циклическую и нециклическую .
В нециклической реакции фотоны захватываются светособирающими антенными комплексами фотосистемы II хлорофиллом (см . и другими вспомогательными пигментами схему справа). Поглощение фотона антенным комплексом ослабляет электрон посредством процесса, называемого фотоиндуцированным разделением зарядов . Антенная система лежит в основе молекулы хлорофилла реакционного центра фотосистемы II. Этот освобожденный электрон поглощается первичной -акцептором электронов молекулой феофитином . Когда электроны перемещаются по цепи переноса электронов (так называемая Z-схема, показанная на диаграмме), хемиосмотический потенциал генерируется за счет перекачки протонных катионов ( H + ) через мембрану в тилакоидное пространство . АТФ-синтаза Фермент использует этот хемиосмотический потенциал для образования АТФ во время фотофосфорилирования , тогда как НАДФН является продуктом терминальной окислительно-восстановительной реакции в Z-схеме . Электрон входит в молекулу хлорофилла в фотосистеме I. Там он далее возбуждается светом, поглощаемым этой фотосистемой . Затем электрон проходит по цепочке акцепторов электронов , которым он передает часть своей энергии . Энергия, доставляемая акцепторам электронов, используется для перемещения ионов водорода через мембрану тилакоида в просвет . Электрон в конечном итоге используется для восстановления кофермента НАДФ с помощью H. + НАДФН (который участвует в светонезависимой реакции); в этот момент путь этого электрона заканчивается.
Циклическая реакция аналогична реакции нециклической, но отличается тем, что генерируется только АТФ и восстановленный НАДФ (НАДФН) не образуется. Циклическая реакция происходит только в фотосистеме I. Как только электрон вытесняется из фотосистемы, электрон передается вниз по молекулам-акцепторам электронов и возвращается в фотосистему I, откуда он был испущен, отсюда и название циклической реакции .
Фотолиз воды
Линейный транспорт электронов через фотосистему оставляет реакционный центр этой фотосистемы окисленным . Подъем другого электрона сначала потребует повторного восстановления реакционного центра. Возбужденные электроны, потерянные из реакционного центра ( Р700 ) фотосистемы I, замещаются переносом от пластоцианина , электроны которого поступают в результате транспорта электронов через фотосистему II . Фотосистема II, как первая ступень Z-схемы , требует внешнего источника электронов для восстановления окисленного хлорофилла в реакционном центре. Источником электронов для фотосинтеза у зеленых растений и цианобактерий является вода. Две молекулы воды окисляются за счет энергии четырех последовательных реакций разделения зарядов фотосистемы II с образованием молекулы двухатомного кислорода и четырех ионов водорода . Образовавшиеся электроны передаются на редокс-активный остаток тирозина , который окисляется за счет энергии P680. + . Это сбрасывает способность P680 поглощать другой фотон и высвобождать еще один фотодиссоциированный электрон. Окисление воды катализируется в фотосистеме II окислительно-восстановительной структурой, содержащей четыре иона марганца и ион кальция ; этот комплекс, выделяющий кислород, связывает две молекулы воды и содержит четыре окислительных эквивалента, которые используются для запуска реакции окисления воды (диаграммы S-состояний Кока). Ионы водорода высвобождаются в просвете тилакоида и, следовательно, способствуют трансмембранному хемиосмотическому потенциалу, который приводит к синтезу АТФ . Кислород — побочный продукт светозависимых реакций, но большинство организмов на Земле используют кислород и его энергию для клеточного дыхания , включая фотосинтезирующие организмы . [ 29 ] [ 30 ]
Светонезависимые реакции
Цикл Кальвина
В светонезависимых (или «темновых») реакциях фермент RuBisCO захватывает CO 2 из атмосферы и в процессе , называемом циклом Кальвина , использует вновь образовавшийся НАДФН и высвобождает трехуглеродные сахара , которые позже соединяются с образованием сахароза и крахмал . Общее уравнение светонезависимых реакций в зеленых растениях имеет вид [ 28 ] : 128

Фиксация углерода приводит к образованию трехуглеродного промежуточного продукта сахара , который затем преобразуется в конечные углеводные продукты. Простые углеродные сахара, образующиеся в результате фотосинтеза, затем используются для образования других органических соединений , таких как строительный материал целлюлоза , предшественники биосинтеза липидов и аминокислот или в качестве топлива для клеточного дыхания . Последнее происходит не только у растений , но и у животных , когда углерод и энергия растений передаются по пищевой цепи .
Фиксация глицерат - или восстановление углекислого газа — это процесс, в котором углекислый газ соединяется с пятиуглеродным сахаром , рибулозо-1,5-бисфосфатом , с образованием двух молекул трехуглеродного соединения, 3-фосфата , также известного как 3-. фосфоглицерат. Глицерат-3-фосфат в присутствии АТФ и НАДФН, образующихся на светозависимых стадиях, восстанавливается до глицеральдегид-3-фосфата . Этот продукт также называют 3-фосфоглицеральдегидом (PGAL) или, в более общем смысле, триозофосфатом . Большая часть (пять из шести молекул) производимого глицеральдегид-3-фосфата используется для регенерации рибулозо-1,5-бисфосфата, чтобы процесс мог продолжаться. Триозофосфаты, не «переработанные» таким образом, часто конденсируются с образованием гексозофосфатов , которые в конечном итоге дают сахарозу , крахмал и целлюлозу , а также глюкозу и фруктозу . Сахара , образующиеся в ходе углеродного метаболизма, образуют углеродные скелеты , которые можно использовать для других метаболических реакций, таких как производство аминокислот и липидов .
Механизмы концентрации углерода
На суше

В жарких и засушливых условиях растения закрывают устьица , чтобы предотвратить потерю воды. В этих условиях содержание CO 2 будет уменьшаться, а газообразного кислорода , образующегося в результате световых реакций фотосинтеза, увеличиваться, вызывая усиление фотодыхания за счет оксигеназной активности рибулозо-1,5-бисфосфаткарбоксилазы/оксигеназы (RuBisCO) и снижение содержания углерода. фиксация. Некоторые растения выработали механизмы увеличения концентрации CO 2 в листьях в этих условиях. [ 31 ]
Растения, которые используют C 4 процесс фиксации углерода химически фиксируют углекислый газ в клетках мезофилла , , добавляя его к трехуглеродной молекуле фосфоенолпирувата (PEP), реакция, под названием катализируемая ферментом PEP карбоксилаза , создавая четырехуглеродную органическую кислоту. щавелевоуксусная кислота . Щавелоуксусная кислота или малат , синтезированные этим процессом, затем транслоцируются в специализированные клетки оболочки пучка фермент RuBisCO , где расположены и другие ферменты цикла Кальвина, и где CO 2, высвобождаемый в результате декарбоксилирования четырехуглеродных кислот, затем фиксируется активностью RuBisCO на трех -углерод- 3-фосфоглицериновые кислоты . Физическое отделение RuBisCO от световых реакций, генерирующих кислород, снижает фотодыхание и увеличивает фиксацию CO и , таким образом, фотосинтетическую способность листа 2 . [ 32 ] С4 Растения растения могут производить больше сахара, чем в условиях С3, высокой освещенности и температуры . Многие важные сельскохозяйственные растения являются растениями C 4 , включая кукурузу , сорго , сахарный тростник и просо . Растения, которые не используют PEP-карбоксилазу для фиксации углерода, называются C 3 растениями, поскольку первичная реакция карбоксилирования , катализируемая RuBisCO, производит трехуглеродные 3-фосфоглицериновые кислоты непосредственно в цикле Кальвина-Бенсона . Более 90% растений используют C 3 фиксацию углерода C 4 ; по сравнению с 3%, которые используют фиксацию углерода [ 33 ] однако эволюция C 4 в более чем шестидесяти линиях растений делает его ярким примером конвергентной эволюции . [ 31 ] C 2 Фотосинтез , который включает концентрацию углерода путем избирательного расщепления фотодыхательного глицина, является одновременно эволюционным предшественником C 4 и полезным механизмом концентрации углерода сам по себе. [ 34 ]
Ксерофиты , такие как кактусы и большинство суккулентов , также используют PEP-карбоксилазу для улавливания углекислого газа в процессе, называемом метаболизмом крассуловой кислоты (CAM). В отличие от метаболизма С4 , который пространственно отделяет фиксацию СО2 к ПЭП от цикла Кальвина, САМ во времени разделяет эти два процесса отличную . Растения CAM имеют анатомию листьев, от растений C 3 , и фиксируют CO 2 ночью, когда их устьица открыты. Растения CAM сохраняют CO 2 в основном в форме яблочной кислоты путем карбоксилирования фосфоенолпирувата до оксалоацетата , который затем восстанавливается до малата. Декарбоксилирование малата в течение дня приводит к высвобождению CO 2 внутри листьев, что позволяет RuBisCO фиксировать углерод до 3-фосфоглицерата. CAM используется 16 000 видами растений. [ 35 ]
Растения, накапливающие оксалаты кальция , такие как Amaranthus Hybridus и Colobanthus quitnsis , демонстрируют разновидность фотосинтеза, при которой кристаллы оксалата кальция функционируют как динамические пулы углерода , поставляя углекислый газ (CO 2 ) в фотосинтезирующие клетки, когда устьица частично или полностью закрыты. Этот процесс получил название тревожного фотосинтеза . В условиях стресса (например, при дефиците воды ) оксалат, высвобождаемый из кристаллов оксалата кальция, превращается в CO 2 под действием фермента оксалатоксидазы , и образующийся CO 2 может поддерживать реакции цикла Кальвина . Реактивная перекись водорода (H 2 O 2 ), побочный продукт оксалатоксидазной реакции, может нейтрализована каталазой быть . Сигнальный фотосинтез представляет собой вариант фотосинтеза, который следует добавить к хорошо известным путям C4 и CAM. Однако тревожный фотосинтез, в отличие от этих путей, действует как биохимический насос, собирающий углерод из внутренней части органа (или из почвы ) , а не из атмосферы. [ 36 ] [ 37 ]
В воде
Цианобактерии обладают карбоксисомами , которые увеличивают концентрацию CO 2 вокруг RuBisCO, увеличивая скорость фотосинтеза. Фермент карбоангидраза , расположенный внутри карбоксисомы, высвобождает CO 2 из растворенных ионов гидрокарбоната (HCO −
3 ). Прежде чем CO 2 сможет диффундировать, RuBisCO, сконцентрированный внутри карбоксисомы, быстро поглощает его. ОЗС −
Ионы 3 образуются из CO 2 вне клетки под действием другой карбоангидразы и активно закачиваются в клетку мембранным белком. Они не могут пересечь мембрану, поскольку заряжены, и внутри цитозоля очень медленно превращаются обратно в CO 2 без помощи карбоангидразы. Это приводит к тому, что HCO −
3 иона накапливаются внутри клетки, откуда они диффундируют в карбоксисомы. [ 38 ] Пиреноиды в водорослях и роголистниках также способствуют концентрации CO 2 вокруг RuBisCO. [ 39 ]
Порядок и кинетика
Общий процесс фотосинтеза проходит в четыре этапа: [ 11 ]
| Этап | Событие | Сайт | Шкала времени |
|---|---|---|---|
| 1 | Перенос энергии в хлорофилле антенны | Тилакоидные мембраны в хлоропластах | Фемтосекунда в Пикосекунда |
| 2 | Перенос электронов в фотохимических реакциях | Пикосекунда в наносекунда | |
| 3 | Электронтранспортная цепь и синтез АТФ | Микросекунда в миллисекунда | |
| 4 | Фиксация углерода и экспорт стабильной продукции | Строма хлоропластов и цитозоль клетки | Миллисекунда в секунду |
Эффективность
Растения обычно преобразуют свет в химическую энергию с эффективностью фотосинтеза 3–6%. [ 40 ] [ 41 ] Поглощенный свет, который не преобразуется, рассеивается в основном в виде тепла , при этом небольшая часть (1–2%) переизлучается в виде флуоресценции хлорофилла на более длинных (более красных) длинах волн . Это обстоятельство позволяет измерять световую реакцию фотосинтеза с помощью хлорофилловых флуориметров . [ 42 ]
Фактическая эффективность фотосинтеза растений зависит от света частоты преобразуемого , интенсивности света , температуры и доли углекислого газа в атмосфере и может варьироваться от 0,1% до 8%. [ 43 ] Для сравнения: солнечные панели преобразуют свет в электрическую энергию с эффективностью примерно 6–20% для панелей массового производства и более 40% в лабораторных устройствах. Ученые изучают фотосинтез в надежде получить растения с повышенной урожайностью . [ 41 ]
Эффективность как световых, так и темновых реакций можно измерить, но взаимосвязь между ними может быть сложной. Например, световая реакция создает АТФ и НАДФН энергетические молекулы , которые C 3 растения могут использовать для фиксации углерода или фотодыхания . [ 44 ] Электроны также могут перетекать к другим стокам электронов. [ 45 ] [ 46 ] [ 47 ] нередко По этой причине авторы проводят различие между работой, выполняемой в нефотореспираторных условиях и в фотодыхательных условиях . [ 48 ] [ 49 ] [ 50 ]
Флуоресценция хлорофилла фотосистемы II может измерять световую реакцию, а инфракрасные газоанализаторы могут измерять темновую реакцию . [ 51 ] Интегрированный флуорометр хлорофилла и система газообмена могут исследовать как световые, так и темновые реакции, когда исследователи используют две отдельные системы вместе. [ 52 ] Инфракрасные газоанализаторы и некоторые датчики влажности достаточно чувствительны, чтобы измерять ассимиляцию CO фотосинтетическую 2 и Δ H 2 O надежными методами . CO 2 обычно измеряется в мкмоль /( м 2 / с ), частей на миллион или объема на миллион; а H 2 O обычно измеряется в ммоль /(м 2 /с) или в мбарах . Измеряя CO 2 ассимиляцию , ΔH 2 O, температуру листа, барометрическое давление , площадь листа и фотосинтетически активную радиацию (ФАР), становится возможным оценить «А» или ассимиляцию углерода «Е» или транспирацию «гс» или устьичная проводимость и «Ci» или внутриклеточный CO 2 . [ 53 ] Тем не менее, там, где это возможно, чаще используют флуоресценцию хлорофилла для измерения стресса растений , поскольку наиболее часто используемые параметры FV/FM и Y(II) или F/FM' можно измерить за несколько секунд, что позволяет исследовать более крупные Популяции растений. [ 50 ]
Системы газообмена , которые обеспечивают контроль уровня CO 2 выше и ниже уровня окружающей среды , позволяют использовать общепринятую практику измерения кривых A/Ci при различных уровнях CO 2 для характеристики фотосинтетической реакции растения. [ 53 ]
Встроенный флуорометр хлорофилла — системы газообмена позволяют более точно измерить реакцию и механизмы фотосинтеза. [ 51 ] [ 52 ] В то время как стандартные системы газообменного фотосинтеза могут измерять Ci или уровни CO 2 в субустьице , добавление интегрированных измерений флуоресценции хлорофилла позволяет более точно измерить CC , оценку концентрации CO 2 в месте карбоксилирования в хлоропластах, чтобы заменить Ci . [ 52 ] [ 54 ] Концентрацию CO 2 в хлоропластах становится возможным оценить путем измерения проводимости мезофилла или г м с помощью интегрированной системы. [ 51 ] [ 52 ] [ 55 ]
Системы измерения фотосинтеза не предназначены для прямого измерения количества света, поглощаемого листом, но анализ флуоресценции хлорофилла , поглощения P700 и P515, а также измерения газообмена позволяют получить подробную информацию, например, о фотосистемах , квантовой эффективности и CO 2 . темпы ассимиляции. даже зависимость эффективности фотосинтеза от длины волны С помощью некоторых инструментов можно анализировать . [ 56 ]
Явление , известное как квантовое блуждание, значительно повышает эффективность переноса энергии света. В фотосинтетической клетке водоросли , бактерии или растения есть светочувствительные молекулы, называемые хромофорами, расположенные в форме антенны, называемой фотокомплексом. Когда фотон поглощается хромофором, он превращается в квазичастицу , называемую экситоном , которая перепрыгивает от хромофора к хромофору к реакционному центру фотокомплекса, совокупности молекул, которые улавливают его энергию в химической форме, доступной для клеточный метаболизм. Волновые свойства экситона позволяют ему покрывать более широкую территорию и опробовать несколько возможных путей одновременно, позволяя мгновенно «выбрать» наиболее эффективный маршрут, по которому он будет иметь наибольшую вероятность прибытия к месту назначения за минимально возможное время.
Поскольку это квантовое блуждание происходит при температурах, намного более высоких, чем обычно происходят квантовые явления, оно возможно только на очень коротких расстояниях. Препятствия в виде деструктивной интерференции заставляют частицу на мгновение терять свои волновые свойства, прежде чем она вновь обретет их после освобождения из заблокированного положения посредством классического «прыжка». Таким образом, движение электрона к фотоцентру рассматривается как серия обычных прыжков и квантовых блужданий. [ 57 ] [ 58 ] [ 59 ]
Эволюция
−4500 — – — – −4000 — – — – −3500 — – — – −3000 — – — – −2500 — – — – −2000 — – — – −1500 — – — – −1000 — – — – −500 — – — – 0 — |
| |||||||||||||||||||||||||||||||||||||||||||||
Возраст окаменелостей того, что считается нитчатыми фотосинтезирующими организмами , составляет 3,4 миллиарда лет. [ 60 ] [ 61 ] Более поздние исследования также предполагают, что фотосинтез мог начаться около 3,4 миллиарда лет назад. [ 62 ] [ 63 ] хотя первые прямые доказательства фотосинтеза получены из тилакоидных мембран, возрастом 1,75 миллиарда лет сохранившихся в кремнях . [ 64 ]
Кислородный фотосинтез — основной источник кислорода в атмосфере Земли , и его самое раннее появление иногда называют кислородной катастрофой . Геологические данные свидетельствуют о том, что кислородный фотосинтез, например, у цианобактерий , стал важным в палеопротерозойскую эпоху около двух миллиардов лет назад. Современный фотосинтез у растений и большинства фотосинтезирующих прокариот является оксигенным, с использованием воды в качестве донора электронов , которая окисляется до молекулярного кислорода в фотосинтетическом реакционном центре .
Симбиоз и происхождение хлоропластов

Некоторые группы животных установили симбиотические отношения с фотосинтезирующими водорослями . Они наиболее распространены в кораллах , губках и морских анемонах . Учёные предполагают, что это связано с особенно простым строением тела и большими поверхностями этих животных по сравнению с объёмами . [ 65 ] Кроме того, некоторые морские моллюски , такие как Elysia viridis и Elysia chromotica , также поддерживают симбиотические отношения с хлоропластами, которые они захватывают из водорослей в своем рационе и затем сохраняют в своем теле (см. Клептопластика ). Это позволяет моллюскам выживать исключительно за счет фотосинтеза в течение нескольких месяцев. [ 66 ] [ 67 ] Некоторые гены растительной из ядра клетки даже были перенесены слизнякам , чтобы хлоропласты могли снабжаться белками, необходимыми для выживания. [ 68 ]
Еще более тесная форма симбиоза может объяснить происхождение хлоропластов. Хлоропласты имеют много общего с фотосинтезирующими бактериями , включая кольцевую хромосому прокариотического типа , рибосому и аналогичные белки в фотосинтетическом реакционном центре . [ 69 ] [ 70 ] Эндосимбиотическая теория предполагает, что фотосинтезирующие бактерии были приобретены (путем эндоцитоза ) ранними эукариотическими клетками и образовали первые растительные клетки. Следовательно, хлоропласты могут быть фотосинтезирующими бактериями, приспособившимися к жизни внутри растительных клеток. Подобно митохондриям , хлоропласты обладают собственной ДНК , отдельной от ядерной ДНК растительных клеток-хозяев, и гены в этой ДНК хлоропластов напоминают гены, обнаруженные у цианобактерий . [ 71 ] ДНК в хлоропластах кодирует окислительно-восстановительные белки, подобные тем, которые обнаруживаются в реакционных центрах фотосинтеза. Гипотеза CoRR предполагает, что такое совместное расположение генов с их генными продуктами необходимо для окислительно-восстановительной регуляции экспрессии генов и объясняет устойчивость ДНК в биоэнергетических органеллах . [ 72 ]
Фотосинтетические эукариотические линии
Исключены симбиотические и клептопластические организмы:
- Глаукофиты многоклеточные , красные и зеленые водоросли — клада Archaeplastida ( одно- и . )
- Криптофиты . — клада Cryptista (одноклеточные)
- Гаптофиты . — клада Haptista (одноклеточные)
- Динофлагелляты Alveolata и хромриды в надтипе Myzozoa и Pseudoblepharisma в типе Ciliophora — кладе ( одноклеточные).
- Охрофиты . — клада Stramenopila (одно- и многоклеточные)
- Хлорарахниофиты . и три вида Paulinella ( типа Cercozoa — клада Rhizaria одноклеточные)
- Эвглениды — клада Excavata (одноклеточные).
За исключением эвгленид, которые встречаются в пределах Excavata , все они относятся к Diaphoretickes . Археопластида и фотосинтетическая Паулинелла получили свои пластиды, окруженные двумя мембранами, в результате первичного эндосимбиоза в двух отдельных событиях, поглотив цианобактерию. Пластиды во всех других группах имеют происхождение из красных или зеленых водорослей и называются «красными линиями» и «зелеными линиями». Единственным известным исключением является инфузория Pseudoblepharisma tenue , которая помимо пластид, произошедших от зеленых водорослей, также имеет в качестве симбионта пурпурную серную бактерию . У динофлагеллят и эвгленид пластиды окружены тремя мембранами, а у остальных линий — четырьмя. Нуклеоморф — остатки исходного ядра водоросли, расположенные между внутренней и внешней мембранами пластиды, имеется у криптофитов (из красной водоросли) и хлорарахниофитов (из зеленой водоросли). [ 73 ] Некоторые динофлагелляты, утратившие свою фотосинтетическую способность, позже вновь восстановили ее в результате новых эндосимбиотических событий с различными водорослями. Многие из этих групп эукариот способны осуществлять фотосинтез, но являются миксотрофами и гетеротрофны в различной степени .
Фотосинтетические прокариотические линии
Считается, что ранние фотосинтетические системы, такие как у зеленых и пурпурных серных и зеленых и пурпурных несерных бактерий , были аноксигенными и использовали в качестве доноров электронов различные другие молекулы, кроме воды . Считается, что зеленые и пурпурные серные бактерии использовали водород и серу в качестве доноров электронов. Зеленые несерные бактерии использовали различные аминокислоты и другие органические кислоты в качестве доноров электронов . Пурпурные несерные бактерии использовали множество неспецифических органических молекул. Использование этих молекул согласуется с геологическими данными о том, что ранняя атмосфера Земли сильно восстанавливалась в то время . [ 74 ]
За возможным исключением Heimdallarchaeota не обнаружен , фотосинтез у архей . [ 75 ] Галоархеи фототрофны и могут поглощать энергию Солнца, но не собирают углерод из атмосферы и, следовательно, не фотосинтезируют. [ 76 ] Вместо хлорофилла они используют родопсины, которые преобразуют энергию света в ионные градиенты, но не могут участвовать в реакциях переноса электронов. [ 77 ] [ 78 ]
У бактерий в настоящее время известны восемь фотосинтетических линий: [ 79 ] [ 80 ] [ 81 ] [ 82 ]
- Цианобактерии — единственные прокариоты, осуществляющие оксигенный фотосинтез, и единственные прокариоты, содержащие два типа фотосистем (тип I (RCI), также известный как тип Fe-S, и тип II (RCII), также известный как хинонный тип). Семь оставшихся прокариот обладают аноксигенным фотосинтезом и используют версии либо типа I, либо типа II.
- Хлороби (зеленые серобактерии) Тип I
- Гелиобактерии типа I
- Хлорацидобактерия типа I
- Протеобактерии (фиолетовые серобактерии и пурпурные несерные бактерии) Тип II
- Хлорофлексота (зеленые несерные бактерии) Тип II
- Гемматимонадота Тип II
- Эремиобактерия типа II
Цианобактерии и эволюция фотосинтеза
Биохимическая способность использовать воду в качестве источника электронов при фотосинтезе однажды развилась у общего предка современных цианобактерий (ранее называвшихся сине-зелеными водорослями). Геологические данные указывают на то, что это преобразующее событие произошло в начале истории Земли, по крайней мере 2450–2320 миллионов лет назад (млн лет назад), и, как предполагается, намного раньше. [ 83 ] [ 84 ] Поскольку во время предполагаемого развития фотосинтеза атмосфера Земли почти не содержала кислорода, считается, что первые фотосинтезирующие цианобактерии не генерировали кислород. [ 85 ] Имеющиеся данные геобиологических исследований архейских (> 2500 млн лет назад) осадочных пород указывают на то, что жизнь существовала 3500 млн лет назад, но вопрос о том, когда возник кислородный фотосинтез, до сих пор остается без ответа. Четкое палеонтологическое окно в эволюцию цианобактерий открылось около 2000 млн лет назад, открыв уже разнообразную биоту цианобактерий. Цианобактерии оставались основными первичными производителями кислорода на протяжении всего протерозойского периода (2500–543 млн лет назад), отчасти потому, что окислительно-восстановительная структура океанов благоприятствовала фотоавтотрофам, способным к фиксации азота . [ 86 ] [ 87 ] Зеленые водоросли присоединились к цианобактериям в качестве основных основных производителей кислорода на континентальных шельфах ближе к концу протерозоя , но только с мезозойской (251–66 млн лет назад) радиацией динофлагеллят, кокколитофорид и диатомей произошло первичное производство кислорода в морских шельфовых водах. принять современную форму. Цианобактерии по-прежнему имеют решающее значение для морских экосистем как основные производители кислорода в океанических круговоротах, как агенты биологической фиксации азота и, в модифицированной форме, как пластиды морских водорослей. [ 88 ]
Экспериментальная история
Открытие
Хотя некоторые этапы фотосинтеза до сих пор до конца не изучены, общее уравнение фотосинтеза известно с 19 века.

Ян ван Гельмонт начал исследование этого процесса в середине 17 века, когда он тщательно измерил массу почвы , которую , использовало растение и массу растения по мере его роста. Заметив, что масса почвы изменилась очень мало, он выдвинул гипотезу , что масса растущего растения должна происходить из воды , единственного вещества, которое он добавлял в горшечное растение. Его гипотеза оказалась частично верной — большая часть набранной массы происходит за счет углекислого газа, а также воды. растения Однако это стало сигналом к идее о том, что основная часть биомассы образуется в результате фотосинтеза, а не из самой почвы.
Джозеф Пристли , химик и священник , обнаружил, что когда он изолировал объем воздуха под перевернутой банкой и зажигал в ней свечу (которая выделяла CO 2 ), свеча сгорала очень быстро, задолго до того, как в ней заканчивался воск. . Далее он обнаружил, что мышь может аналогичным образом «повредить» воздух. Затем он показал, что растение может восстановить воздух, который «повредили» свеча и мышь. [ 89 ]
В 1779 году Ян Ингенхауз Пристли повторил эксперименты . Он обнаружил, что именно влияние солнечного света на растение может заставить его оживить мышь за считанные часы. [ 89 ] [ 90 ]
В 1796 году Жан Сенебье , швейцарский пастор , ботаник и натуралист , продемонстрировал , что зеленые растения потребляют углекислый газ и выделяют кислород под воздействием света . Вскоре после этого Николя-Теодор де Соссюр показал, что увеличение массы растения по мере его роста может происходить не только за счет поглощения CO 2 , но и за счет включения воды. Таким образом, была намечена основная реакция , посредством которой организмы используют фотосинтез для производства пищи (например, глюкозы ). [ 91 ]
Уточнения
Корнелис Ван Ниль сделал ключевые открытия, объясняющие химию фотосинтеза. Изучая пурпурные серобактерии и зеленые бактерии , он первым продемонстрировал, что фотосинтез представляет собой светозависимую окислительно-восстановительную реакцию , в которой водород восстанавливает (отдает свои атомы в виде электронов и протонов ) углекислого газа.
Роберт Эмерсон обнаружил две световые реакции, проверяя продуктивность растений с использованием света разной длины волны. Только красный цвет подавлял световые реакции. Когда синий и красный были объединены, результат был гораздо более существенным. Таким образом, существовало две фотосистемы: одна поглощала длины волн до 600 нм, другая — до 700 нм. Первый известен как PSII, второй — PSI. PSI содержит только хлорофилл «а», PSII содержит преимущественно хлорофилл «а» с большей частью доступного хлорофилла «b», среди других пигментов. К ним относятся фикобилины, являющиеся красными и синими пигментами красных и синих водорослей соответственно, и фукоксантол для бурых водорослей и диатомовых водорослей. Процесс наиболее продуктивен, когда поглощение квантов одинаково как в PSII, так и в PSI, гарантируя, что входная энергия от антенного комплекса распределяется между системами PSI и PSII, что, в свою очередь, приводит в действие фотохимию. [ 11 ]
Роберт Хилл считал, что комплекс реакций состоит из промежуточного соединения с цитохромом b 6 (ныне пластохиноном), а другой - с цитохрома f на этап механизмов генерации углеводов. Они связаны пластохиноном, которому требуется энергия для восстановления цитохрома f. Дальнейшие эксперименты, доказывающие, что кислород, образующийся при фотосинтезе зеленых растений, поступает из воды, были проведены Хиллом в 1937 и 1939 годах. Он показал, что изолированные хлоропласты выделяют кислород в присутствии неприродных восстановителей, таких как железа оксалат , феррицианид или бензохинон, после воздействия. осветить. В реакции Хилла: [ 92 ]
- 2 H 2 O + 2 A + (свет, хлоропласты) → 2 AH 2 + O 2
А – акцептор электронов. Следовательно, на свету акцептор электронов восстанавливается и выделяется кислород. Сэмюэль Рубен и Мартин Кеймен использовали радиоактивные изотопы , чтобы определить, что кислород, выделяемый при фотосинтезе, поступает из воды.

Мелвин Кэлвин и Эндрю Бенсон вместе с Джеймсом Башемом выяснили путь ассимиляции углерода (цикл фотосинтетического восстановления углерода) в растениях. Цикл сокращения выбросов углерода известен как цикл Кальвина , но многие ученые называют его циклом Кальвина-Бенсона, Бенсона-Кельвина или даже циклом Кальвина-Бенсона-Башама (или CBB).
Лауреат Нобелевской премии ученый Рудольф А. Маркус позже смог открыть функцию и значение цепи переноса электронов.
Отто Генрих Варбург и Дин Берк открыли I-квантовую реакцию фотосинтеза, которая расщепляет CO 2 , активируемую дыханием. [ 93 ]
первые экспериментальные доказательства существования фотофосфорилирования in vivo, представил В 1950 году Отто Кандлер используя интактные клетки хлореллы и интерпретируя свои открытия как светозависимое образование АТФ . [ 94 ] В 1954 году Дэниел И. Арнон и др. обнаружил фотофосфорилирование in vitro в изолированных хлоропластах с помощью P 32 . [ 95 ] [ 96 ]
Луи Н. М. Дуйсенс и Ян Амес обнаружили, что хлорофилл «а» поглощает один свет и окисляет цитохром f, в то время как хлорофилл «а» (и другие пигменты) поглощают другой свет, но восстанавливают тот же самый окисленный цитохром, заявив, что две световые реакции происходят в ряд.
Разработка концепции
В 1893 году американский ботаник Чарльз Рид Барнс предложил два термина — фотосинтаксис и фотосинтез — для биологического процесса синтеза сложных соединений углерода из угольной кислоты в присутствии хлорофилла под воздействием света . Термин «фотосинтез» происходит от греческих слов phos ( φῶς — сияние) и синтез ( σύνθεσις — соединение вместе). [ 97 ] [ 98 ] [ 99 ] в то время как другое слово, которое он обозначил, было фотосинтаксис , от синтаксиса ( σύνταξις , конфигурация). Со временем термин фотосинтез вошел в обиход. Позднее открытие аноксигенных фотосинтезирующих бактерий и фотофосфорилирования потребовало переопределения этого термина. [ 100 ]
C3: исследование фотосинтеза C4
В конце 1940-х годов в Калифорнийском университете в Беркли детали фотосинтетического метаболизма углерода разбирались химиками Мелвином Кэлвином , Эндрю Бенсоном, Джеймсом Бэшемом и множеством студентов и исследователей, использующих методы изотопа углерода-14 и бумажной хроматографии. [ 101 ] Путь фиксации CO 2 водорослями хлореллой за долю секунды на свету привел к образованию трехуглеродной молекулы, называемой фосфоглицериновой кислотой (ФГК). За эту оригинальную и новаторскую работу Мелвину Кэлвину в 1961 году была присуждена Нобелевская премия по химии. Параллельно физиологи растений изучали газообмен листьев, используя новый метод инфракрасного газового анализа и листовую камеру, где чистая скорость фотосинтеза варьировалась от от 10 до 13 мкмоль CO 2 ·м −2 ·с −1 , с выводом, что все наземные растения обладают одинаковыми фотосинтетическими способностями и насыщаются светом при менее чем 50% солнечного света. [ 102 ] [ 103 ]
Позже, в 1958–1963 годах, в Корнелльском университете выращенная в полевых условиях, кукуруза, сообщалось, что имела гораздо более высокую скорость фотосинтеза листьев - 40 мкмоль CO 2 ·м. −2 ·с −1 и не насыщаться при почти полном солнечном свете. [ 104 ] [ 105 ] Этот более высокий показатель у кукурузы был почти вдвое выше, чем у других видов, таких как пшеница и соя, что указывает на большие различия в фотосинтезе среди высших растений. В Университете Аризоны подробное исследование газообмена у более чем 15 видов однодольных и двудольных растений впервые выявило, что различия в анатомии листьев являются решающими факторами в дифференциации фотосинтетических способностей между видами. [ 106 ] [ 107 ] У тропических трав, включая кукурузу, сорго, сахарный тростник, бермудскую траву и двудольный амарант, скорость фотосинтеза листьев составляла около 38–40 мкмоль CO 2 ·м. −2 ·с −1 , а листья имеют два типа зеленых клеток, т. е. внешний слой клеток мезофилла, окружающий плотно упакованные клетки хлорофилловой оболочки сосудистого пучка. Этот тип анатомии был назван анатомией Кранца в 19 веке ботаником Готлибом Хаберландтом во время изучения анатомии листьев сахарного тростника. [ 108 ] Виды растений с наибольшей скоростью фотосинтеза и анатомией Кранца не показали явного фотодыхания, очень низкую точку компенсации CO 2 , высокую оптимальную температуру, высокую устойчивость устьиц и более низкую устойчивость мезофилла к диффузии газа и показатели никогда не насыщаются при полном солнечном свете. [ 109 ] Исследование в Аризоне было удостоено награды Citation Classic в 1986 году. [ 107 ] Эти виды позже были названы растениями C4, поскольку первое стабильное соединение, фиксирующее CO 2 на свету, содержит четыре атома углерода в виде малата и аспартата. [ 110 ] [ 111 ] [ 112 ] Другие виды, у которых отсутствует анатомия Кранца, были названы типом C3, например хлопок и подсолнечник, поскольку первым стабильным углеродным соединением является трехуглеродный PGA. При концентрации CO 2 1000 ppm в воздухе растения C3 и C4 имели одинаковую скорость фотосинтеза листьев около 60 мкмоль CO 2 ·м. −2 ·с −1 что указывает на подавление фотодыхания у С3-растений. [ 106 ] [ 107 ]
Факторы

Существует четыре основных фактора, влияющих на фотосинтез, и несколько сопутствующих факторов. Четыре основных: [ 113 ]
- Световое излучение и длина волны
- Водопоглощение
- углекислого газа Концентрация
- Температура .
Общий фотосинтез ограничен рядом факторов окружающей среды. К ним относятся количество доступного света, площадь листьев , которые растение должно улавливать свет (затенение другими растениями является основным ограничением фотосинтеза), скорость, с которой углекислый газ может поступать в хлоропласты для поддержки фотосинтеза, доступность воды и наличие подходящей температуры для проведения фотосинтеза. [ 114 ]
Интенсивность света (облученность), длина волны и температура

Процесс фотосинтеза обеспечивает основной вклад свободной энергии в биосферу и является одним из четырех основных способов, которыми радиация важна для жизни растений. [ 115 ]
Радиационный климат внутри растительных сообществ чрезвычайно изменчив как во времени, так и в пространстве.
В начале 20 века Фредерик Блэкман и Габриэль Маттеи исследовали влияние интенсивности света ( облучения ) и температуры на скорость ассимиляции углерода.
- При постоянной температуре скорость ассимиляции углерода варьируется в зависимости от освещенности, увеличиваясь по мере увеличения освещенности, но достигая плато при более высокой освещенности.
- При низкой освещенности повышение температуры мало влияет на скорость ассимиляции углерода. При постоянной высокой освещенности скорость ассимиляции углерода увеличивается с повышением температуры.
Эти два эксперимента иллюстрируют несколько важных моментов: во-первых, известно, что на фотохимические не влияет реакции, как правило, температура . Однако эти эксперименты ясно показывают, что температура влияет на скорость ассимиляции углерода, поэтому в полном процессе ассимиляции углерода должно происходить два набора реакций. Это светозависимая «фотохимическая» независимая от температуры стадия и светонезависимая, зависящая от температуры стадия. Во-вторых, эксперименты Блэкмана иллюстрируют концепцию ограничивающих факторов . Еще одним ограничивающим фактором является длина волны света. Цианобактерии, обитающие на глубине нескольких метров под водой, не могут получать волны правильной длины, необходимые для фотоиндуцированного разделения зарядов в обычных фотосинтетических пигментах. Для борьбы с этой проблемой у цианобактерий есть светособирающий комплекс, называемый фикобилисомой . [ 116 ] Этот комплекс состоит из ряда белков с различными пигментами, окружающих реакционный центр.
Уровень углекислого газа и фотодыхание

По мере роста концентрации углекислого газа скорость образования сахаров в результате светонезависимых реакций увеличивается, пока не ограничивается другими факторами. RuBisCO , фермент, который захватывает углекислый газ в светонезависимых реакциях, имеет сродство связывания как с углекислым газом, так и с кислородом. Когда концентрация углекислого газа высока, RuBisCO фиксирует углекислый газ. Однако если концентрация углекислого газа низкая, RuBisCO будет связывать кислород вместо углекислого газа. Этот процесс, называемый фотодыханием , использует энергию, но не производит сахара.
Активность оксигеназы RuBisCO невыгодна для растений по нескольким причинам:
- Одним из продуктов оксигеназной активности является фосфогликолят (2 углерода) вместо 3-фосфоглицерата (3 углерода). Фосфогликолят не может метаболизироваться в цикле Кальвина-Бенсона и представляет собой потерю углерода в ходе цикла. Таким образом, высокая оксигеназная активность истощает сахара, необходимые для переработки рибулозо-5-бисфосфата и продолжения цикла Кальвина-Бенсона .
- Фосфогликолят быстро метаболизируется до гликолата, который в высоких концентрациях токсичен для растений; он подавляет фотосинтез.
- Утилизация гликолата — это энергетически дорогостоящий процесс, в котором используется гликолатный путь, и только 75% углерода возвращается в цикл Кальвина-Бенсона в виде 3-фосфоглицерата. В результате реакций также образуется аммиак (NH 3 ), который способен диффундировать за пределы растения, что приводит к потере азота.
- Сильно упрощенное резюме таково:
- 2 гликолат + АТФ → 3-фосфоглицерат + углекислый газ + АДФ + NH 3
Путь утилизации продуктов оксигеназной активности RuBisCO более известен как фотодыхание, поскольку он характеризуется светозависимым потреблением кислорода и выделением углекислого газа.
См. также
- Ян Андерсон (ученый)
- Искусственный фотосинтез
- Цикл Кальвина-Бенсона
- Фиксация углерода
- Клеточное дыхание
- Хемосинтез
- Интеграл дневного света
- Реакция Хилла
- Встроенный флуорометр
- Светозависимая реакция
- Органическая реакция
- Фотобиология
- Фотоингибирование
- Фотосинтетический реакционный центр
- Фотосинтетически активная радиация
- Фотосистема
- Фотосистема I
- Фотосистема II
- квантовый
- Квантовая биология
- Радиосинтез
- Красный край
- Витамин Д
Ссылки
- ^ «Фотосинтез» . lexico.com ( Английский словарь Lexico UK). Издательство Оксфордского университета. Архивировано из оригинала 11 августа 2022 г. Проверено 15 июля 2023 г.
- ^ Перейти обратно: а б Брайант, Дональд А.; Фригаард, Нильс-Ульрик (ноябрь 2006 г.). «Освещенный фотосинтез и фототрофия прокариот» . Тенденции в микробиологии . 14 (11): 488–496. дои : 10.1016/j.tim.2006.09.001 . ПМИД 16997562 .
- ^ Рис, Джейн Б.; Урри, Лиза А.; Каин, Майкл Л.; Вассерман, Стивен А.; Минорский, Петр Васильевич; Джексон, Роберт Б.; Кэмпбел, Нил А. (2011). Биология (международное изд.). Река Аппер-Сэддл, Нью-Джерси: Pearson Education . стр. 235, 244 . ISBN 978-0-321-73975-9 .
Это первоначальное включение углерода в органические соединения известно как фиксация углерода.
- ^ Олсон Дж. М. (май 2006 г.). «Фотосинтез в архейскую эпоху». Исследования фотосинтеза . 88 (2): 109–117. Бибкод : 2006PhoRe..88..109O . дои : 10.1007/s11120-006-9040-5 . ПМИД 16453059 . S2CID 20364747 .
- ^ Бьюик Р (август 2008 г.). «Когда появился кислородный фотосинтез?» . Философские труды Лондонского королевского общества, серия B. 363 (1504): 2731–2743. дои : 10.1098/rstb.2008.0041 . ПМК 2606769 . ПМИД 18468984 .
- ^ Нилсон К.Х., Конрад П.Г. (декабрь 1999 г.). «Жизнь: прошлое, настоящее и будущее» . Философские труды Лондонского королевского общества, серия B. 354 (1392): 1923–1939. дои : 10.1098/rstb.1999.0532 . ПМЦ 1692713 . ПМИД 10670014 .
- ^ Уитмарш, Джон; Говинджи (1999). «Глава 2: Процесс фотосинтеза» . В Сингхале GS; Ренгер Г.; Сопоры СК; Иррганг К.Д.; Говинджи (ред.). Понятия фотобиологии: фотосинтез и фотоморфогенез . Бостон: Kluwer Academic Publishers . стр. 11–51. ISBN 978-0-7923-5519-9 . Архивировано из оригинала 14 августа 2010 г. Проверено 7 июля 2012 г.
Подсчитано, что фотосинтезирующие организмы удаляют 100 × 10 15 грамм углерода/год, фиксируемый фотосинтезирующими организмами. Это эквивалентно 4 × 10 18 кДж/год свободной энергии, хранящейся в восстановленном углероде. (в части 8: «Глобальный фотосинтез и атмосфера»)
- ^ Стегер У., Ахтерберг В., Блок К., Боде Х., Френц В., Гатер С., Ханекамп Г., Имбоден Д., Янке М., Кост М., Курц Р., Нуцингер Х.Г., Циземер Т. (2005). Устойчивое развитие и инновации в энергетическом секторе . Берлин: Шпрингер . п. 32. ISBN 978-3-540-23103-5 . Архивировано из оригинала 2 сентября 2016 г. Проверено 21 февраля 2016 г.
Средняя глобальная скорость фотосинтеза составляет 130 ТВт.
- ^ «Мировое потребление первичной энергии по типам энергии и отдельным группам стран, 1980–2004 гг.» . Управление энергетической информации . 31 июля 2006 года. Архивировано из оригинала (XLS) 9 ноября 2006 года . Проверено 20 января 2007 г.
- ^ Филд CB, Беренфельд М.Дж., Рандерсон Дж.Т., Фальковски П. (июль 1998 г.). «Первичная продукция биосферы: интеграция наземных и океанических компонентов» . Наука . 281 (5374): 237–240. Бибкод : 1998Sci...281..237F . дои : 10.1126/science.281.5374.237 . ПМИД 9657713 . Архивировано из оригинала 25 сентября 2018 г. Проверено 20 апреля 2018 г.
- ^ Перейти обратно: а б с «Фотосинтез». Энциклопедия науки и технологий Макгроу-Хилла . Том. 13. Нью-Йорк: МакГроу-Хилл . 2007. ISBN 978-0-07-144143-8 .
- ^ Франкелиус П. (июль – август 2020 г.). «Предложение переосмыслить сельское хозяйство в климатических расчетах» . Агрономический журнал . 112 (4): 3216–3221. Бибкод : 2020AgrJ..112.3216F . дои : 10.1002/agj2.20286 . S2CID 219423329 .
- ^ Растения: разнообразие и эволюция.
- ^ Георгий, Дришья М.; Винсент, Аннет С.; Макки, Хэмиш Р. (2020). «Обзор аноксигенных фототрофных бактерий и их применения в экологической биотехнологии для устойчивого восстановления ресурсов» . Отчеты по биотехнологии (Амстердам, Нидерланды) . 28 : e00563. дои : 10.1016/j.btre.2020.e00563 . ISSN 2215-017X . ПМЦ 7714679 . ПМИД 33304839 .
- ^ Фукс, Георг (1987). «Восстановление углекислого газа анаэробными бактериями» . В Аресте, М.; Форти, Г. (ред.). Углекислый газ как источник углерода: биохимическое и химическое использование . Дордрехт: Springer Нидерланды. стр. 263–273. дои : 10.1007/978-94-009-3923-3_14 . ISBN 978-94-009-3923-3 . Проверено 10 июня 2024 г.
- ^ Стефано, Джордж Б.; Снайдер, Кристофер; Крим, Ричард М. (17 июля 2015 г.). «Митохондрии, хлоропласты в клетках животных и растений: значение конформационного соответствия» . Medical Science Monitor: Международный медицинский журнал экспериментальных и клинических исследований . 21 : 2073–2078. дои : 10.12659/MSM.894758 . ISSN 1643-3750 . ПМЦ 4517925 . ПМИД 26184462 .
- ^ Симакава, Гинга; Мацуда, Юсуке; Бурлакот, Адриан (2024). «Взаимосвязь между фотосинтезом и дыханием у микробов» . Журнал биологических наук . 49 (2): 45. дои : 10.1007/s12038-023-00417-4 . ISSN 0973-7138 . ПМИД 38516912 .
- ^ Уитмарш и Говинджи 1999 , с. 13.
- ^ Анаэробный фотосинтез , Новости химии и техники , 86 , 33, 18 августа 2008 г., стр. 36
- ^ Кулп Т.Р., Хоефт С.Э., Асао М., Мэдиган М.Т., Холлибо Дж.Т., Фишер Дж.К., Штольц Дж.Ф., Калбертсон К.В., Миллер Л.Г., Оремланд Р.С. (август 2008 г.). «Мышьяк(III) питает аноксигенный фотосинтез в биопленках горячих источников из озера Моно, Калифорния» . Наука . 321 (5891): 967–970. Бибкод : 2008Sci...321..967K . дои : 10.1126/science.1160799 . ПМИД 18703741 . S2CID 39479754 . Архивировано из оригинала 28 июля 2020 г. Проверено 17 января 2020 г.
- ^ «Ученые обнаружили уникальный микроб в самом большом озере Калифорнии» . bio-medicine.org . Январь 2005 г. Архивировано из оригинала 12 июля 2009 г. Проверено 20 июля 2009 г.
- ^ Ингруй М., Эдди Б. (17 августа 2006 г.). Растения: разнообразие и эволюция . Издательство Кембриджского университета. стр. 13–14. ISBN 978-1-139-45546-6 .
- ^ Тавано CL, Донохью Т.Дж. (декабрь 2006 г.). «Развитие фотосинтетического аппарата бактерий» . Современное мнение в микробиологии . 9 (6): 625–631. дои : 10.1016/j.mib.2006.10.005 . ПМК 2765710 . ПМИД 17055774 .
- ^ Перейти обратно: а б Муллино CW (1999). «Тилакоидные мембраны цианобактерий: строение, динамика и функции». Австралийский журнал физиологии растений . 26 (7): 671–677. дои : 10.1071/PP99027 .
- ^ Сенер М.К., Олсен Дж.Д., Хантер К.Н., Шультен К. (октябрь 2007 г.). «Структурно-функциональная модель бактериального фотосинтетического мембранного пузырька на атомном уровне» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (40): 15723–15728. Бибкод : 2007PNAS..10415723S . дои : 10.1073/pnas.0706861104 . ПМК 2000399 . ПМИД 17895378 .
- ^ Кэмпбелл Н.А., Уильямсон Б., Хейден Р.Дж. (2006). Биология, изучающая жизнь . Река Аппер-Сэддл, Нью-Джерси: Прентис-Холл . ISBN 978-0-13-250882-7 . Архивировано из оригинала 2 ноября 2014 г. Проверено 3 февраля 2009 г.
- ^ Цихе Д., Дюншеде Б., Шунеманн Д. (декабрь 2018 г.). «Молекулярный механизм SRP-зависимого транспорта светособирающих белков к мембране тилакоидов у растений» . Исследования фотосинтеза . 138 (3): 303–313. Бибкод : 2018PhoRe.138..303Z . дои : 10.1007/s11120-018-0544-6 . ПМК 6244792 . ПМИД 29956039 .
- ^ Перейти обратно: а б Рэйвен П.Х., Эверт Р.Ф., Эйххорн С.Е. (2005). Биология растений (7-е изд.). Нью-Йорк: WH Freeman and Company . стр. 124–127 . ISBN 978-0-7167-1007-3 .
- ^ «Ячандра/Яно Групп» . Национальная лаборатория Лоуренса Беркли. Архивировано из оригинала 22 июля 2019 г. Проверено 22 июля 2019 г.
- ^ Пушкарь Ю., Яно Дж., Зауэр К., Буссак А., Ячандра В.К. (февраль 2008 г.). «Структурные изменения кластера Mn4Ca и механизм фотосинтетического расщепления воды» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (6): 1879–1884. Бибкод : 2008PNAS..105.1879P . дои : 10.1073/pnas.0707092105 . ПМЦ 2542863 . ПМИД 18250316 .
- ^ Перейти обратно: а б Уильямс Б.П., Джонстон И.Г., Ковшофф С., Хибберд Дж.М. (сентябрь 2013 г.). «Фенотипический ландшафтный вывод раскрывает множество эволюционных путей фотосинтеза C4» . электронная жизнь . 2 : е00961. doi : 10.7554/eLife.00961 . ПМЦ 3786385 . ПМИД 24082995 .
- ^ Таиз Л., Гейгер Э. (2006). Физиология растений (4-е изд.). Синауэр Ассошиэйтс . ISBN 978-0-87893-856-8 .
- ^ Монсон Р.К., Сейдж РФ (1999). «Таксономическое распределение C
4 Фотосинтез» . C 4 Биология растений . Бостон: Academic Press . стр. 551–580. ISBN 978-0-12-614440-6 . Архивировано из оригинала 19 января 2023 г. Проверено 17 апреля 2019 г. - ^ Лундгрен М.Р. (декабрь 2020 г.). «Фотосинтез C 2: многообещающий путь к улучшению урожая?» . Новый фитолог . 228 (6): 1734–1740. дои : 10.1111/nph.16494 . ПМИД 32080851 .
- ^ Додд А.Н., Борланд А.М., Хаслам Р.П., Гриффитс Х., Максвелл К. (апрель 2002 г.). «Метаболизм крассулацовых кислот: пластичный, фантастический» . Журнал экспериментальной ботаники . 53 (369): 569–580. дои : 10.1093/jexbot/53.369.569 . ПМИД 11886877 .
- ^ Тулаку Г., Яннопулос А., Николопулос Д., Бреста П., Доцика Е., Оркула М.Г. и др. (август 2016 г.). «Тревожный фотосинтез: кристаллы оксалата кальция как внутренний источник CO2 в растениях» . Физиология растений . 171 (4): 2577–2585. дои : 10.1104/стр.16.00111 . ПМЦ 4972262 . ПМИД 27261065 .
- ^ Гомес-Эспиноса О, Гонсалес-Рамирес Д, Бреста П, Карабурниотис Г, Браво Л.А. (октябрь 2020 г.). «Разложение кристаллов оксалата кальция в Colobanthus quitnsis в условиях ограничения CO 2 » . Растения . 9 (10): 1307. doi : 10.3390/plants9101307 . ПМК 7600318 . ПМИД 33023238 .
- ^ Бэджер М.Р., Прайс Г.Д. (февраль 2003 г.). «Механизмы концентрации CO2 у цианобактерий: молекулярные компоненты, их разнообразие и эволюция» . Журнал экспериментальной ботаники . 54 (383): 609–622. дои : 10.1093/jxb/erg076 . ПМИД 12554704 .
- ^ Бэджер М.Р., Эндрюс Дж.Т., Уитни С.М., Людвиг М., Йеллоулис, округ Колумбия, Леггат В., Прайс Г.Д. (1998). «Разнообразие и коэволюция Рубиско, пластид, пиреноидов и механизмов концентрации CO 2 на основе хлоропластов в водорослях». Канадский журнал ботаники . 76 (6): 1052–1071. дои : 10.1139/b98-074 .
- ^ Миямото К. «Глава 1 – Производство биологической энергии» . Возобновляемые биологические системы для альтернативного устойчивого производства энергии (Бюллетень сельскохозяйственных услуг ФАО – 128) . Продовольственная и сельскохозяйственная организация Объединенных Наций. Архивировано из оригинала 7 сентября 2013 года . Проверено 4 января 2009 г.
- ^ Перейти обратно: а б Эренберг Р. (15 декабря 2017 г.). «Исправление фотосинтеза» . Познаваемый журнал . Ежегодные обзоры. дои : 10.1146/knowable-121917-115502 . Архивировано из оригинала 7 апреля 2022 г. Проверено 03 апреля 2018 г.
- ^ Максвелл К., Джонсон Дж. Н. (апрель 2000 г.). «Флуоресценция хлорофилла – практическое руководство» . Журнал экспериментальной ботаники . 51 (345): 659–668. дои : 10.1093/jexbot/51.345.659 . ПМИД 10938857 .
- ^ Говинджи, Раджни. «Что такое фотосинтез?» . Биология в Иллинойсе. Архивировано из оригинала 27 мая 2014 года . Проверено 17 апреля 2014 г.
- ^ Розенквист Э., ван Кутен О (2006). «Глава 2: Флуоресценция хлорофилла: общее описание и номенклатура» . В DeEll JA, Toivonen PM (ред.). Практическое применение флуоресценции хлорофилла в биологии растений . Дордрехт, Нидерланды: Kluwer Academic Publishers. стр. 39–78. ISBN 9781461504153 . Архивировано из оригинала 19 января 2023 г. Проверено 17 апреля 2019 г.
- ^ Бейкер Н.Р., Оксборо К. (2004). «Глава 3: Флуоресценция хлорофилла как показатель продуктивности фотосинтеза» . В Папакеоргиу Г., Говинджи (ред.). Флуоресценция хлорофиллы – признак фотосинтеза . Дордрехт, Нидерланды: Springer. стр. 66–79. Архивировано из оригинала 19 января 2023 г. Проверено 17 апреля 2019 г.
- ^ Флексас Дж., Эскальнона Дж.М., Медрано Х. (январь 1999 г.). «Водный стресс вызывает разные уровни фотосинтеза и регуляцию скорости транспорта электронов у виноградных лоз» . Растение, клетка и окружающая среда . 22 (1): 39–48. дои : 10.1046/j.1365-3040.1999.00371.x .
- ^ Фрайер М.Дж., Эндрюс-младший, Оксборо К., Блоуерс Д.А., Бейкер Н.Р. (1998). «Взаимосвязь между ассимиляцией CO 2 , фотосинтетическим транспортом электронов и активным метаболизмом O 2 в листьях кукурузы в поле в периоды низких температур» . Физиология растений . 116 (2): 571–580. дои : 10.1104/стр.116.2.571 . ПМЦ 35114 . ПМИД 9490760 .
- ^ Эрл Х., Саид Эннали С. (2004). «Оценка фотосинтетического транспорта электронов с помощью флуорометрии хлорофилла без светового насыщения Фотосистемы II». Исследования фотосинтеза . 82 (2): 177–186. Бибкод : 2004PhoRe..82..177E . дои : 10.1007/s11120-004-1454-3 . ПМИД 16151873 . S2CID 291238 .
- ^ Дженти Б., Бриантаис Дж., Бейкер Н.Р. (1989). «Взаимосвязь между квантовым выходом фотосинтетического электронного транспорта и тушением флуоресценции хлорофилла». Biochimica et Biophysical Acta (BBA) – Общие предметы . 990 (1): 87–92. дои : 10.1016/s0304-4165(89)80016-9 .
- ^ Перейти обратно: а б Бейкер Н.Р. (2008). «Флуоресценция хлорофилла: исследование фотосинтеза in vivo ». Ежегодный обзор биологии растений . 59 : 89–113. doi : 10.1146/annurev.arplant.59.032607.092759 . ПМИД 18444897 . S2CID 31451852 .
- ^ Перейти обратно: а б с Бернакки С.Дж., Портис А.Р., Накано Х., фон Кеммерер С., Лонг С.П. (2002). «Температурная реакция проводимости мезофилла. Значение для определения кинетики фермента Рубиско и ограничений фотосинтеза in vivo» . Физиология растений . 130 (4): 1992–1998. дои : 10.1104/стр.008250 . ПМК 166710 . ПМИД 12481082 .
- ^ Перейти обратно: а б с д Рибас-Карбо М., Флексас Дж., Робинсон С.А., Черкез Г.Г. (2010). « in vivo Измерение дыхания растений ». Онлайн-исследования Университета Вуллонгонга .
- ^ Перейти обратно: а б Лонг СП, Бернакки CJ (2003). «Измерения газообмена: что они могут сказать нам об основных ограничениях фотосинтеза? Процедуры и источники ошибок» . Журнал экспериментальной ботаники . 54 (392): 2393–2401. дои : 10.1093/jxb/erg262 . ПМИД 14512377 .
- ^ Бернакки С.Дж., Портис А.Р., Накано Х., фон Кеммерер С., Лонг С.П. (2002). «Температурная реакция проводимости незофилла. Значение для определения кинетики фермента Рубиско и ограничений фотосинтеза in vivo » . Физиология растений . 130 (4): 1992–1998. дои : 10.1104/стр.008250 . ПМК 166710 . ПМИД 12481082 .
- ^ Инь Икс, Струик ПК (2009). «Теоретический пересмотр при оценке проводимости мезофилла к диффузии CO 2 в листьях растений C3 путем анализа комбинированных измерений газообмена и флуоресценции хлорофилла» . Растение, клетка и окружающая среда . 32 (11): 1513–1524 [1524]. дои : 10.1111/j.1365-3040.2009.02016.x . ПМИД 19558403 .
- ^ Шрайбер У., Клугхаммер С., Колбовски Дж. (2012). «Оценка зависимых от длины волны параметров фотосинтетического электронного транспорта с помощью нового типа многоцветного хлорофиллового флуориметра ПАМ» . Исследования фотосинтеза . 113 (1–3): 127–144. Бибкод : 2012PhoRe.113..127S . дои : 10.1007/s11120-012-9758-1 . ПМЦ 3430841 . ПМИД 22729479 .
- ^ Палмер Дж. (21 июня 2013 г.). «Растения «занимаются квантовой физикой» » . Новости Би-би-си . Архивировано из оригинала 3 октября 2018 года . Проверено 21 июня 2018 г.
- ^ Ллойд С. (10 марта 2014 г.). «Квантовая биология: лучше жить благодаря квантовой механике» . Природа реальности. Нова: PBS Online; WGBH Бостон. Архивировано из оригинала 3 июля 2017 года . Проверено 8 сентября 2017 г.
- ^ Хилднер Р., Бринкс Д., Нидер Дж.Б., Когделл Р.Дж., ван Хульст Н.Ф. (июнь 2013 г.). «Квантовая когерентная передача энергии по различным путям в отдельных светособирающих комплексах». Наука . 340 (6139): 1448–1451. Бибкод : 2013Sci...340.1448H . дои : 10.1126/science.1235820 . ПМИД 23788794 . S2CID 25760719 .
- ^ Дэвис К. (2 октября 2004 г.). «Фотосинтез начался очень рано» . Новый учёный . Архивировано из оригинала 1 мая 2015 года . Проверено 8 сентября 2017 г.
- ^ Хупер Р. (19 августа 2006 г.). «Открытие зарождения фотосинтеза» . Новый учёный . Архивировано из оригинала 24 мая 2015 года . Проверено 8 сентября 2017 г.
- ^ Кардона Т (март 2018 г.). «Раннеархейское происхождение гетеродимерной фотосистемы I» . Гелион . 4 (3): e00548. Бибкод : 2018Heliy...400548C . doi : 10.1016/j.heliyon.2018.e00548 . ПМЦ 5857716 . ПМИД 29560463 .
- ^ Ховард V (7 марта 2018 г.). «Фотосинтез возник на миллиард лет раньше, чем мы думали, как показывают исследования» . Журнал астробиологии . Архивировано из оригинала 1 октября 2020 года . Проверено 23 марта 2018 г.
- ^ Демулен, Катрин Ф.; Лара, Янник Дж.; Ламбион, Александр; Яво, Эммануэль Ж. (2024). «Старейшие тилакоиды в ископаемых клетках прямо свидетельствуют о кислородном фотосинтезе» . Природа . 625 (7995): 529–534. Бибкод : 2024Natur.625..529D . дои : 10.1038/s41586-023-06896-7 . ПМИД 38172638 . S2CID 266752333 .
- ^ Венн А.А., Лорам Дж.Э., Дуглас А.Э. (2008). «Фотосинтетические симбиозы у животных» . Журнал экспериментальной ботаники . 59 (5): 1069–1080. дои : 10.1093/jxb/erm328 . ПМИД 18267943 .
- ^ Румфо М.Э., Саммер Э.Дж., Манхарт-младший (май 2000 г.). «Морские слизни, работающие на солнечной энергии. Симбиоз моллюсков и хлоропластов водорослей» . Физиология растений . 123 (1): 29–38. дои : 10.1104/стр.123.1.29 . ПМЦ 1539252 . ПМИД 10806222 .
- ^ Мускатин Л., Грин Р.В. (1973). «Хлоропласты и водоросли как симбионты у моллюсков». Международный обзор цитологии . 36 : 137–169. дои : 10.1016/S0074-7696(08)60217-X . ISBN 978-0-12-364336-0 . ПМИД 4587388 .
- ^ Румфо М.Э., Уорфул Дж.М., Ли Дж., Каннан К., Тайлер М.С., Бхаттачарья Д., Мустафа А., Манхарт Дж.Р. (ноябрь 2008 г.). «Горизонтальный перенос гена ядерного гена водоросли psbO фотосинтезирующему морскому слизню Elysia chromotica» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (46): 17867–17871. Бибкод : 2008PNAS..10517867R . дои : 10.1073/pnas.0804968105 . ПМЦ 2584685 . ПМИД 19004808 .
- ^ Дуглас С.Е. (декабрь 1998 г.). «Эволюция пластид: происхождение, разнообразие, тенденции». Текущее мнение в области генетики и развития . 8 (6): 655–661. дои : 10.1016/S0959-437X(98)80033-6 . ПМИД 9914199 .
- ^ Рейес-Прието А., Вебер А.П., Бхаттачарья Д. (2007). «Происхождение и формирование пластид у водорослей и растений». Ежегодный обзор генетики . 41 : 147–168. дои : 10.1146/annurev.genet.41.110306.130134 . ПМИД 17600460 . S2CID 8966320 . [ постоянная мертвая ссылка ]
- ^ Рэйвен Дж.А., Аллен Дж.Ф. (2003). «Геномика и эволюция хлоропластов: что цианобактерии сделали для растений?» . Геномная биология . 4 (3): 209. doi : 10.1186/gb-2003-4-3-209 . ПМК 153454 . ПМИД 12620099 .
- ^ Аллен Дж. Ф. (декабрь 2017 г.). «Гипотеза CoRR для генов в органеллах» . Журнал теоретической биологии . 434 : 50–57. Бибкод : 2017JThBi.434...50A . дои : 10.1016/j.jtbi.2017.04.008 . ПМИД 28408315 .
- ^ Килинг П.Дж. (март 2010 г.). «Эндсимбиотическое происхождение, диверсификация и судьба пластид» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 365 (1541): 729–748. дои : 10.1098/rstb.2009.0103 . ПМК 2817223 . ПМИД 20124341 .
- ^ Гейл Дж (2009). Астробиология Земли: возникновение, эволюция и будущее жизни на планете, находящейся в смятении . Издательство Оксфордского университета. стр. 112–113. ISBN 978-0-19-154835-2 . Архивировано из оригинала 19 января 2023 г. Проверено 17 апреля 2019 г.
- ^ Лю Р., Цай Р., Чжан Дж., Сунь С. (февраль 2020 г.). «Heimdallarchaeota использует энергию света посредством фотосинтеза». биоRxiv . дои : 10.1101/2020.02.20.957134 . S2CID 213816522 .
- ^ ДасСарма С., Швитерман Е.В. (июнь 2021 г.). «Ранняя эволюция фиолетовых пигментов сетчатки на Земле и значение биосигнатур экзопланет» . Международный журнал астробиологии . 20 (3): 241–250. arXiv : 1810.05150 . Бибкод : 2021IJAsB..20..241D . дои : 10.1017/S1473550418000423 . S2CID 119341330 . Разместить резюме в: «Царство фиолетового цвета: возможно, когда-то в жизни на Земле доминировали фиолетовые микроорганизмы» . CBC/Радио Канады . 26 октября 2018 г.
- ^ Гамильтон ТЛ (август 2019 г.). «Проблема с кислородом: экофизиология современных фототрофов и последствия для эволюции оксигенного фотосинтеза» . Свободно-радикальная биология и медицина . 140 : 233–249. doi : 10.1016/j.freeradbiomed.2019.05.003 . ПМИД 31078729 . S2CID 153285864 .
- ^ Шарма А.К., Уолш Д.А., Баптесте Э., Родригес-Валера Ф., Форд Дулиттл В., Папке Р.Т. (май 2007 г.). «Эволюция ионных насосов родопсина у галоархей» . Эволюционная биология BMC . 7 (1): 79. Бибкод : 2007BMCEE...7...79S . дои : 10.1186/1471-2148-7-79 . ПМЦ 1885257 . ПМИД 17511874 .
- ^ Сюн Дж (2006). «Фотосинтез: какой цвет возник?» . Геномная биология . 7 (12): 245. doi : 10.1186/gb-2006-7-12-245 . ПМК 1794423 . ПМИД 17210067 .
- ^ Паоли Л., Рушевей Х.Дж., Форнерис CC, Хубрич Ф., Каутсар С., Бхушан А. и др. (июль 2022 г.). «Биосинтетический потенциал микробиома глобального океана» . Природа . 607 (7917): 111–118. дои : 10.1038/s43705-022-00201-9 . ПМЦ 9758169 . ПМИД 35732736 .
- ^ Хе З., Ферлез Б., Курашов В., Танк М., Гольбек Дж. Х., Брайант Д. А. (октябрь 2019 г.). «Реакционные центры термофильного микроаэрофила Chloracidobacterium thermophilum (Acidobacteria) I: биохимическая и биофизическая характеристика» . Исследования фотосинтеза . 142 (1): 87–103. Бибкод : 2019PhoRe.142...87H . дои : 10.1007/s11120-019-00650-9 . ПМИД 31161318 . S2CID 254941681 .
- ^ Цзэн Ю, Фэн Ф, Медова Х, Дин Дж, Коближек М (май 2014 г.). «Функциональные фотосинтетические реакционные центры 2-го типа, обнаруженные у редкого бактериального типа Gemmatimonadetes» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (21): 7795–7800. Бибкод : 2014PNAS..111.7795Z . дои : 10.1073/pnas.1400295111 . ПМК 4040607 . ПМИД 24821787 .
- ^ Томитани А., Нолл А.Х., Кавано С.М., Оно Т. (апрель 2006 г.). «Эволюционное разнообразие цианобактерий: молекулярно-филогенетические и палеонтологические перспективы» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (14): 5442–5447. Бибкод : 2006PNAS..103.5442T . дои : 10.1073/pnas.0600999103 . ПМЦ 1459374 . ПМИД 16569695 .
- ^ «Цианобактерии: летопись окаменелостей» . ucmp.berkeley.edu . Архивировано из оригинала 24 августа 2010 г. Проверено 26 августа 2010 г.
- ^ Смит А (2010). Биология растений . Нью-Йорк: Garland Science. п. 5. ISBN 978-0-8153-4025-6 . Архивировано из оригинала 19 января 2023 г. Проверено 17 апреля 2019 г.
- ^ Олсон, Стефани Л.; Рейнхард, Кристофер Т.; Лайонс, Тимоти В. (2016). «Цианобактериальная диазотрофия и замедленная оксигенация Земли» . Границы микробиологии . 7 : 1526. дои : 10.3389/fmicb.2016.01526 . ISSN 1664-302X . ПМК 5033965 . ПМИД 27721813 .
- ^ Санчес-Баракальдо, Патрисия; Бьянкини, Джорджо; Уилсон, Джейми Д.; Нолл, Эндрю Х. (2022). «Цианобактерии и биогеохимические циклы в истории Земли» . Тенденции в микробиологии . 30 (2): 143–157. дои : 10.1016/j.tim.2021.05.008 . ISSN 1878-4380 . ПМИД 34229911 .
- ^ Эрреро А., Флорес Э. (2008). Цианобактерии: молекулярная биология, геномика и эволюция (1-е изд.). Кайстер Академик Пресс. ISBN 978-1-904455-15-8 . Архивировано из оригинала 19 января 2023 г. Проверено 17 апреля 2019 г.
- ^ Перейти обратно: а б Мартин Д., Томпсон А., Стюарт И., Гилберт Э., Хоуп К., Каваи Дж., Гриффитс А. (сентябрь 2012 г.). «Парадигма хрупкой Земли в колпаке Пристли» . Экстремальная физиология и медицина . 1 (1): 4. дои : 10.1186/2046-7648-1-4 . ПМК 3707099 . ПМИД 23849304 .
- ^ Гест Х (2000). «Двухсотлетний юбилей доктора Яна Инген-Хоуза, доктора медицины (1730-1799), пионера исследований фотосинтеза» . Исследования фотосинтеза . 63 (2): 183–190. дои : 10.1023/А:1006460024843 . ПМИД 16228428 . S2CID 22970505 .
- ^ Рабинович Е.И. (1945). Фотосинтез и связанные с ним процессы . Том. 1. Архивировано из оригинала 6 августа 2020 г. Получено 14 декабря 2019 г. - из Библиотеки наследия биоразнообразия .
- ^ Уокер Д.А. (2002). « И чье яркое присутствие» – оценка Роберта Хилла и его реакции» (PDF) . Исследования фотосинтеза . 73 (1–3): 51–54. дои : 10.1023/А:1020479620680 . ПМИД 16245102 . S2CID 21567780 . Архивировано из оригинала (PDF) 9 марта 2008 г. Проверено 27 августа 2015 г.
- ↑ Отто Варбург – Биография , заархивированная 15 декабря 2010 г. в Wayback Machine . Nobelprize.org (1 августа 1970 г.). Проверено 3 ноября 2011 г.
- ^ Кандлер О (1950). «О связи между фосфатным обменом и фотосинтезом I. Изменения уровня фосфатов у Chlorellapyrenoidosa как следствие изменений света и темноты] (PDF) . Журнал естественных исследований . 5б (8): 423–437. дои : 10.1515/znb-1950-0806 . S2CID 97588826 . Архивировано (PDF) из оригинала 24 июня 2018 г. Проверено 26 июня 2018 г.
- ^ Арнон Д.И. , Уотли Ф.Р., Аллен М.Б. (1954). «Фотосинтез изолированными хлоропластами. II. Фотофосфорилирование, преобразование света в энергию фосфатной связи». Журнал Американского химического общества . 76 (24): 6324–6329. дои : 10.1021/ja01653a025 .
- ^ Арнон Д.И. (1956). «Фосфорный обмен и фотосинтез». Ежегодный обзор физиологии растений . 7 : 325–354. дои : 10.1146/annurev.pp.07.060156.001545 .
- ^ «Фотосинтез» . Интернет-словарь этимологии . Архивировано из оригинала 7 марта 2013 г. Проверено 23 мая 2013 г.
- ^ Лидделл Х.Г. , Скотт Р. «φῶς» . Греко-английский лексикон . Проект Персей .
- ^ Лидделл Х.Г. , Скотт Р. «σύνθεσις» . Греко-английский лексикон . Проект Персей .
- ^ Гест Х (2002). «История слова фотосинтез и эволюция его определения». Исследования фотосинтеза . 73 (1–3): 7–10. дои : 10.1023/А:1020419417954 . ПМИД 16245098 . S2CID 11265932 .
- ^ Кэлвин М. (июль 1989 г.). «Сорок лет фотосинтеза и связанной с ним деятельности». Исследования фотосинтеза . 21 (1): 3–16. Бибкод : 1989PhoRe..21....3C . дои : 10.1007/BF00047170 . ПМИД 24424488 . S2CID 40443000 .
- ^ Верден Ж (1953). «Таблица скоростей фотосинтеза в оптимальных, близких к естественным условиях». Являюсь. Дж. Бот . 40 (9): 675–679. дои : 10.1002/j.1537-2197.1953.tb06540.x . JSTOR 2439681 .
- ^ Вердуин Дж., Уитвер Э.Э., Коуэлл, Британская Колумбия (июль 1959 г.). «Максимальная скорость фотосинтеза в природе». Наука . 130 (3370): 268–269. Бибкод : 1959Sci...130..268В . дои : 10.1126/science.130.3370.268 . ПМИД 13668557 . S2CID 34122342 .
- ^ Хескет Дж. Д., Масгрейв Р. (1962). «Фотосинтез в полевых условиях. IV. Световые исследования с отдельными листьями кукурузы». Наука о растениеводстве . 2 (4): 311–315. дои : 10.2135/cropsci1962.0011183x000200040011x . S2CID 83706567 .
- ^ Хескет Дж. Д., Мосс Д. Н. (1963). «Изменение реакции фотосинтеза на свет». Наука о растениеводстве . 3 (2): 107–110. дои : 10.2135/cropsci1963.0011183X000300020002x .
- ^ Перейти обратно: а б Эль-Шаркави, Массачусетс, Хескет Дж.Д. (1965). «Фотосинтез видов в зависимости от особенностей анатомии листьев и устойчивости к диффузии CO 2 ». Наука о растениеводстве . 5 (6): 517–521. дои : 10.2135/cropsci1965.0011183x000500060010x .
- ^ Перейти обратно: а б с Эль-Шаркави М.А., Хескет Дж.Д. (1986). «Цитирование Classic-Фотосинтез среди видов в связи с особенностями анатомии листьев и CO 2 сопротивлением диффузии » (PDF) . Курс. Продолжение/Агр.Биол.Экология . 27 : 14. Архивировано из оригинала (PDF) 29 ноября 2023 г. Проверено 6 декабря 2023 г.
- ^ Хаберландт Г (1904). Физиологическая анатомия растений . Лейпциг: Энгельманн. Архивировано из оригинала 19 января 2023 г. Проверено 17 апреля 2019 г.
- ^ Эль-Шаркави М.А. (1965). Факторы, ограничивающие скорость фотосинтеза разных видов растений (кандидатская диссертация). Университет Аризоны, Тусон.
- ^ Карпилов Ю.С. (1960). «Распределение радиоактивности углерода-14 среди продуктов фотосинтеза кукурузы». Учеб. Казанский агропромышленный комплекс. Инст . 14 :15–24.
- ^ Корчак Х.П., Харт CE, Берр ГО (1965). «Фиксация углекислого газа в листьях сахарного тростника» . Физиол растений . 40 (2): 209–213. дои : 10.1104/стр.40.2.209 . ПМК 550268 . ПМИД 16656075 .
- ^ Хэтч, доктор медицинских наук, Slack CR (1966). «Фотосинтез листьев сахарного тростника. Новая реакция карбоксилирования и путь образования сахара» . Биохим. Дж . 101 (1): 103–111. дои : 10.1042/bj1010103 . ПМК 1270070 . ПМИД 5971771 .
- ^ Стирбет А., Лазар Д., Го Й., Говинджи Г. (сентябрь 2020 г.). «Фотосинтез: основы, история и моделирование» . Анналы ботаники . 126 (4): 511–537. дои : 10.1093/aob/mcz171 . ПМЦ 7489092 . ПМИД 31641747 . Проверено 9 февраля 2023 г.
- ^ Чапин Ф.С., Мэтсон П.А., Муни Х.А. (2002). Принципы экологии наземных экосистем . Нью-Йорк: Спрингер. стр. 97–104. ISBN 978-0-387-95443-1 . Архивировано из оригинала 19 января 2023 г. Проверено 17 апреля 2019 г.
- ^ Джонс Х.Г. (2014). Растения и микроклимат: количественный подход к физиологии растений в окружающей среде (Третье изд.). Кембридж: Издательство Кембриджского университета. ISBN 978-0-521-27959-8 . Архивировано из оригинала 19 января 2023 г. Проверено 17 апреля 2019 г.
- ^ Адир Н., Бар-Цви С., Харрис Д. (апрель 2020 г.). «Удивительная фикобилисома» . Biochimica et Biophysica Acta (BBA) — Биоэнергетика . Легкий сбор урожая. 1861 (4): 148047. doi : 10.1016/j.bbabio.2019.07.002 . ПМИД 31306623 . S2CID 196810874 .
Дальнейшее чтение
Книги
- Бидлак Дж. Э., Стерн К. Р., Янски С. (2003). Введение в биологию растений . Нью-Йорк: МакГроу-Хилл. ISBN 978-0-07-290941-8 .
- Бланкеншип RE (2014). Молекулярные механизмы фотосинтеза (2-е изд.). Джон Уайли и сыновья . ISBN 978-1-4051-8975-0 . Архивировано из оригинала 19 января 2023 г. Проверено 17 апреля 2019 г.
- Говинджи, Битти Дж. Т., Гест Х., Аллен Дж. Ф. (2006). Открытия в области фотосинтеза . Достижения в области фотосинтеза и дыхания. Том. 20. Берлин: Шпрингер. ISBN 978-1-4020-3323-0 . Архивировано из оригинала 19 января 2023 г. Проверено 17 апреля 2019 г.
- Рис Дж.Б. и др. (2013). Кэмпбелл Биология . Бенджамин Каммингс . ISBN 978-0-321-77565-8 .
Статьи
- Гупта Р.С., Мухтар Т., Сингх Б. (июнь 1999 г.). «Эволюционные взаимоотношения между фотосинтезирующими прокариотами ( Heliobacterium chromum , Chloroflexus aurantiacus , цианобактериями, Chlorobium tepidum и протеобактериями): последствия, касающиеся происхождения фотосинтеза». Молекулярная микробиология . 32 (5): 893–906. дои : 10.1046/j.1365-2958.1999.01417.x . ПМИД 10361294 . S2CID 33477550 .
- Резерфорд А.В., Фаллер П. (январь 2003 г.). «Фотосистема II: эволюционные перспективы» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 358 (1429): 245–253. дои : 10.1098/rstb.2002.1186 . ПМК 1693113 . ПМИД 12594932 .
Внешние ссылки
- Сборник страниц по фотосинтезу для всех уровней от известного эксперта (Говинджи).
- Углубленное, современное лечение фотосинтеза, также от Говинджи.
- Научное пособие: Фотосинтез. Статья, подходящая для изучения естественных наук в средней школе.
- Метаболизм, клеточное дыхание и фотосинтез - Виртуальная библиотека биохимии и клеточной биологии
- Общий экзамен по фотосинтезу на среднем уровне.
- Общая энергетика фотосинтеза
- Источник кислорода, вырабатываемого фотосинтезом. Интерактивная анимация, учебное пособие.
- Маршалл Дж. (29 марта 2011 г.). «Дебют первого практического искусственного листа» . Новости Дискавери. Архивировано из оригинала 22 марта 2012 г. Проверено 29 марта 2011 г.
- Фотосинтез - светозависимые и светонезависимые стадии. Архивировано 10 сентября 2011 г. на Wayback Machine.
- Академия Хана, видео-знакомство
| Субдисциплины | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| растений Группы | |||||||||||
| Анатомия растений |
| ||||||||||
| Физиология растений Материалы | |||||||||||
| Рост растений и привычка | |||||||||||
| Воспроизведение | |||||||||||
| Таксономия растений | |||||||||||
| Упражняться | |||||||||||
| |||||||||||
метаболизма Карта |
|---|
