Многоклеточный организм

Многоклеточный организм — это организм , состоящий из более чем одной клетки , в отличие от одноклеточных организмов . [1] Все виды животных , наземных растений и большинство грибов , как и многие водоросли , являются многоклеточными, тогда как некоторые организмы частично одно- и частично многоклеточные, например, слизевики и социальные амебы, такие как род Dictyostelium . [2] [3]
Многоклеточные организмы возникают различными путями, например, путем деления клеток или путем агрегации множества одиночных клеток. [4] [3] Колониальные организмы возникают в результате объединения множества одинаковых особей в колонию . Однако часто бывает трудно отличить колониальные протисты от настоящих многоклеточных организмов, поскольку эти две концепции не различны; Колониальные протисты получили название «многоклеточных», а не «многоклеточных». [5] [6] Существуют также макроскопические многоядерные организмы , хотя технически одноклеточные, такие как Xenophyophorea, длина которых может достигать 20 см.
Эволюционная история
[ редактировать ]возникновение
[ редактировать ]Многоклеточность развивалась независимо у эукариот по крайней мере 25 раз . [7] [8] а также в некоторых прокариотах , таких как цианобактерии , миксобактерии , актиномицеты , Magnetoglobus multicellis или Methanosarcina . [3] Однако сложные многоклеточные организмы развились только в шести группах эукариот: животные , симбиомикотановые грибы , бурые водоросли , красные водоросли , зеленые водоросли и наземные растения . [9] Он неоднократно эволюционировал у хлоропластид (зеленых водорослей и наземных растений), один раз у животных, один раз у бурых водорослей, трижды у грибов ( хитриды , аскомицеты и базидиомицеты ). [10] и, возможно, несколько раз для слизевиков и красных водорослей. [11] Первые свидетельства многоклеточной организации, когда одноклеточные организмы координируют поведение и могут быть эволюционным предшественником истинной многоклеточности, получены от цианобактериоподобных организмов, которые жили 3,0–3,5 миллиарда лет назад. [7] Для размножения настоящие многоклеточные организмы должны решить проблему регенерации целого организма из зародышевых клеток (т. е. сперматозоидов и яйцеклеток ), проблему, которая изучается в эволюционной биологии развития . развилось значительное разнообразие типов клеток (100–150 различных типов клеток) по сравнению с 10–20 у растений и грибов. У животных в многоклеточном организме [12]
Потеря многоклеточности
[ редактировать ]В некоторых группах произошла потеря многоклеточности. [13] Грибы преимущественно многоклеточные, хотя ранние дивергентные линии в основном одноклеточные (например, Microsporidia ), и у грибов наблюдались многочисленные возвраты к одноклеточности (например, Saccharomycotina , Cryptococcus и другие дрожжи ). [14] [15] Это также могло произойти с некоторыми красными водорослями (например, Porphyridium ), но они могут быть примитивно одноклеточными. [16] Утрата многоклеточности также считается вероятной у некоторых зеленых водорослей (например, Chlorella vulgaris и некоторых Ulvophyceae ). [17] [18] В других группах, обычно паразитах, произошло снижение многоклеточности, количества или типов клеток (например, миксозойные организмы , многоклеточные организмы, которые ранее считались одноклеточными, вероятно, являются чрезвычайно редуцированными книдариями ). [19]
Рак
[ редактировать ]Многоклеточные организмы, особенно долгоживущие животные, сталкиваются с проблемой рака , который возникает, когда клетки не могут регулировать свой рост в рамках нормальной программы развития. Во время этого процесса можно наблюдать изменения в морфологии тканей. Рак у животных ( многоклеточных животных ) часто описывается как потеря многоклеточности и атавистический возврат к одноклеточному состоянию. [20] Многие гены, ответственные за установление многоклеточности, возникшие при появлении многоклеточных животных, в раковых клетках дерегулированы, включая гены, которые контролируют дифференцировку клеток , адгезию и межклеточную связь . [21] [22] Идет дискуссия о возможности существования рака у других многоклеточных организмов. [23] [24] или даже у простейших . [25] Например, галлы растений характеризуются как опухоли . [26] но некоторые авторы утверждают, что растения не заболевают раком. [27]
Разделение соматических и половых клеток
[ редактировать ]В некоторых многоклеточных группах, которые называются вейсманистами , возникло разделение на стерильную линию соматических клеток и линию зародышевых клеток . Однако вейсманистское развитие сравнительно редко (например, у позвоночных, членистоногих, вольвоксов ), так как большая часть видов обладает способностью к соматическому эмбриогенезу (например, наземные растения, большинство водорослей, многие беспозвоночные). [28] [10]
Гипотезы происхождения
[ редактировать ]Одна из гипотез происхождения многоклеточности заключается в том, что группа клеток со специфическими функциями объединилась в слизнеобразную массу, называемую грексом , которая двигалась как многоклеточная единица. По сути, это то, что делают слизевики . Другая гипотеза состоит в том, что примитивная клетка подверглась делению ядра, став, таким образом, ценоцитом . Затем вокруг каждого ядра (а также клеточного пространства и органелл, занимающих это пространство) образуется мембрана, в результате чего в одном организме образуется группа связанных клеток (этот механизм наблюдается у дрозофилы ). Третья гипотеза заключается в том, что при разделении одноклеточного организма дочерние клетки не смогли разделиться, что привело к скоплению идентичных клеток в одном организме, из которого впоследствии могли развиться специализированные ткани. Это то, что делают эмбрионы растений и животных , а также колониальные хоанофлагелляты . [29] [30]
Поскольку первые многоклеточные организмы были простыми, мягкими организмами, лишенными костей, панциря или других твердых частей тела, они плохо сохранились в летописи окаменелостей. [31] Единственным исключением может быть демоспонж , который, возможно, оставил химический след в древних камнях. Самые ранние окаменелости многоклеточных организмов включают спорную Grypania спиральную и окаменелости черных сланцев палеопротерозойской формации франвильской группы ископаемых B в Габоне ( Gabonionta ). [32] В формации Доушантуо обнаружены микроокаменелости возрастом 600 миллионов лет с признаками многоклеточных свойств. [33]
До недавнего времени филогенетическая реконструкция осуществлялась на основе анатомического (особенно эмбриологического ) сходства. Это неточно, поскольку живые многоклеточные организмы, такие как животные и растения, произошли от своих одноклеточных предков более чем на 500 миллионов лет назад. Такой ход времени позволяет как дивергентному, так и конвергентному времени эволюции имитировать сходства и накапливать различия между группами современных и вымерших предковых видов. Современная филогенетика использует сложные методы, такие как аллоферменты , сателлитная ДНК и другие молекулярные маркеры, для описания признаков, общих для отдаленно родственных линий. [ нужна ссылка ]
Эволюция многоклеточности могла происходить несколькими путями, некоторые из которых описаны ниже:
Симбиотическая теория
[ редактировать ]Эта теория предполагает, что первые многоклеточные организмы возникли в результате симбиоза (сотрудничества) разных видов одноклеточных организмов, каждый из которых выполнял разные роли. Со временем эти организмы станут настолько зависимыми друг от друга, что не смогут выжить независимо, что в конечном итоге приведет к объединению их геномов в один многоклеточный организм. [34] Каждый соответствующий организм станет отдельной линией дифференцированных клеток внутри вновь созданного вида. [ нужна ссылка ]
Этот вид строго взаимозависимого симбиоза можно часто наблюдать, например, во взаимоотношениях между рыбами-клоунами и морскими анемонами Ритерри . В этих случаях крайне сомнительно, что какой-либо вид выживет очень долго, если другой вымрет. Однако проблема этой теории заключается в том, что до сих пор неизвестно, как ДНК каждого организма может быть включена в один геном, чтобы составить их как единый вид. Хотя теоретически такой симбиоз имел место (например, митохондрии и хлоропласты в животных и растительных клетках — эндосимбиоз ), он происходил лишь крайне редко, и даже тогда геномы эндосимбионтов сохраняли элемент различия, отдельно реплицируя свою ДНК. во время митоза вида-хозяина. Например, два или три симбиотических организма, образующих сложный лишайник , хотя выживание и зависят друг от друга, должны размножаться по отдельности, а затем переформироваться, чтобы снова создать один индивидуальный организм. [ нужна ссылка ]
Теория клеточности (синцитиальная)
[ редактировать ]Эта теория утверждает, что один одноклеточный организм с несколькими ядрами мог иметь внутренние мембранные перегородки вокруг каждого из своих ядер. [35] Многие протисты, такие как инфузории или слизевики, могут иметь несколько ядер, что подтверждает эту гипотезу . Однако простого наличия нескольких ядер недостаточно для подтверждения теории. Множественные ядра инфузорий неодинаковы и имеют четко дифференцированные функции. Макроядро . микроядро служит потребностям организма, а используется для полового размножения с обменом генетического материала слизевиков Синцитии образуются из отдельных амебоидных клеток, как и синцитиальные ткани некоторых многоклеточных организмов, а не наоборот. Чтобы считаться обоснованной, эта теория нуждается в наглядном примере и механизме образования многоклеточного организма из ранее существовавшего синцития. [ нужна ссылка ]
Колониальная теория
[ редактировать ]Колониальная теория Геккеля , 1874, предполагает, что симбиоз многих организмов одного вида (в отличие от симбиотической теории , предполагающей симбиоз разных видов) привел к образованию многоклеточного организма. По крайней мере, у некоторых - предположительно, возникших на суше - многоклеточность возникает в результате разделения клеток, а затем воссоединения (например, клеточные слизевики ), тогда как для большинства многоклеточных типов (тех, которые развились в водной среде) многоклеточность возникает как следствие отказа клеток. выделить следующее подразделение. [36] Механизм формирования последней колонии может быть таким же простым, как неполный цитокинез , хотя обычно считается, что многоклеточность включает клеточную дифференцировку . [37]
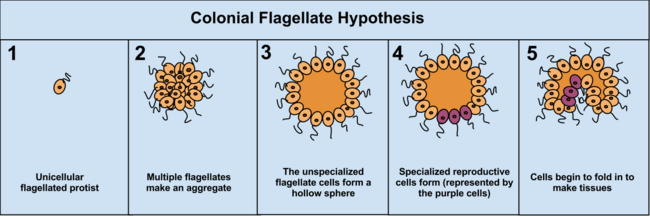
Преимущество гипотезы колониальной теории состоит в том, что она встречается независимо в 16 различных типах протоктистана. Например, во время нехватки пищи амеба Dictyostelium группируется в колонию, которая как одна перемещается на новое место. Некоторые из этих амеб затем слегка дифференцируются друг от друга. Другими примерами колониальной организации у protista являются Volvocaceae , такие как Eudorina и Volvox , последний из которых состоит из до 500–50 000 клеток (в зависимости от вида), лишь часть из которых размножается. [38] Например, у одного вида размножаются 25–35 клеток, 8 – бесполым и около 15–25 – половым. Однако часто бывает трудно отличить колониальные простейшие от настоящих многоклеточных организмов, поскольку эти две концепции не различны; Колониальные протисты получили название «многоклеточных», а не «многоклеточных». [5]
Теория синзооспор
[ редактировать ]что возникновение многоклеточности, по крайней мере у Metazoa, произошло вследствие перехода от временной к пространственной дифференцировке клеток , а не путем постепенной эволюции клеточной дифференцировки, как утверждается в Геккеля Некоторые авторы предполагают , теории гастреи . [39]
ГК-ПИД
[ редактировать ]Около 800 миллионов лет назад [40] незначительное генетическое изменение в одной молекуле, называемой доменом взаимодействия с гуанилаткиназой белка (GK-PID), возможно, позволило организмам перейти от одноклеточного организма к одной из многих клеток. [41]
Роль вирусов
[ редактировать ]Недавно было установлено, что гены, заимствованные у вирусов и мобильных генетических элементов (МГЭ), играют решающую роль в дифференцировке многоклеточных тканей и органов и даже в половом размножении, в слиянии яйцеклеток и сперматозоидов. [42] [43] Такие слитые клетки также участвуют в мембранах многоклеточных животных, например, в тех, которые предотвращают проникновение химических веществ через плаценту и разделение тела мозга. [42] Были идентифицированы два вирусных компонента. Первый — синцитин , полученный из вируса. [44] Второй, идентифицированный в 2002 году, называется EFF-1 . [45] который помогает формировать кожуру Caenorhabditis elegans , члена целого семейства белков FF. Феликс Рей из Института Пастера в Париже построил трехмерную структуру белка EFF-1. [46] и показано, что он выполняет работу по связыванию одной клетки с другой при вирусных инфекциях.Тот факт, что все известные слитые клетки молекулы имеют вирусное происхождение, предполагает, что они были жизненно важны для систем межклеточной коммуникации, которые сделали возможным многоклеточное существование. Без способности к слиянию клеток могли бы образоваться колонии, но что-то даже столь сложное, как губка, было бы невозможно. [47]
Гипотеза доступности кислорода
[ редактировать ]Эта теория предполагает, что кислород, доступный в атмосфере ранней Земли, мог быть ограничивающим фактором для возникновения многоклеточной жизни. [48] Эта гипотеза основана на корреляции между возникновением многоклеточной жизни и повышением уровня кислорода в это время. Это должно было произойти после Великого события окисления , но до недавнего повышения уровня кислорода. Миллс [49] приходит к выводу, что количество кислорода, присутствовавшее во время эдиакарского периода, не было необходимым для сложной жизни и, следовательно, вряд ли было движущим фактором возникновения многоклеточности. [ нужна ссылка ]
Гипотеза Земли-снежка
[ редактировать ]Земля -снежок — это геологическое явление, при котором вся поверхность Земли покрыта снегом и льдом. Этот термин может относиться либо к отдельным событиям (которых было как минимум два), либо к более крупному геологическому периоду, в течение которого произошли все известные полные оледенения.
Самый последний снежный ком на Земле произошел в криогенный период и состоял из двух глобальных событий оледенения, известных как Стуртовское и Мариноское оледенения. Сяо и др . [50] предполагают, что между периодом времени, известным как « Скучный миллиард » и Землей-снежком, простая жизнь могла успеть обновиться и развиться, что позже могло привести к эволюции многоклеточности.
Гипотеза Земли-снежка в отношении многоклеточности предполагает, что криогенный период в истории Земли мог стать катализатором эволюции сложной многоклеточной жизни. Брокс [51] предполагает, что время между Стуртским ледниковым периодом и более поздним Мариноским ледниковым периодом позволило планктонным водорослям доминировать в морях, открывая путь быстрому разнообразию жизни как растений, так и животных. Сложная жизнь быстро возникла и разнообразилась в ходе так называемого Кембрийского взрыва вскоре после Маринойского взрыва. [ нужна ссылка ]
Гипотеза хищничества
[ редактировать ]Гипотеза хищничества предполагает, что, чтобы избежать поедания хищниками, простые одноклеточные организмы развили многоклеточность, чтобы их было труднее съесть в качестве добычи. Херрон и др. [52] провел лабораторные эксперименты по эволюции на одноклеточной зеленой водоросли Chlamydomonas Reinhardtii , используя инфузорию в качестве хищника. Они обнаружили, что в присутствии этого хищника C. Reinhardtii действительно приобретает простые многоклеточные особенности. [ нужна ссылка ]
Экспериментальная эволюция
[ редактировать ]Невозможно узнать, что произошло, когда отдельные клетки превратились в многоклеточные организмы сотни миллионов лет назад. Однако мы можем выявить мутации , способные превратить одноклеточные организмы в многоклеточные. Это продемонстрировало бы возможность такого события. Одноклеточные виды могут относительно легко приобретать мутации, которые заставляют их прикрепляться друг к другу, что является первым шагом на пути к многоклеточности. В ходе эволюции появилось множество обычно одноклеточных видов, демонстрирующих такие ранние стадии:
- Давно известно, что дрожжи обладают флокуляцией . Одним из первых генов дрожжей, вызывающих этот фенотип, является FLO1. [53] Более ярко скомканный фенотип , называемый «снежинкой», возникает из-за потери одного транскрипционного фактора Ace2 . Дрожжи-снежинки растут в многоклеточные кластеры, которые быстро оседают; они были идентифицированы путем направленной эволюции. [54] Совсем недавно (2024 г.) дрожжи-снежинки претерпели более 3000 поколений дальнейшей направленной эволюции, образуя макроскопические ансамбли размером в миллиметры . Были выявлены изменения во многих генах. Кроме того, авторы сообщили, что только анаэробные культуры снежных дрожжей развили этот признак, а аэробные - нет. [55]
- Ряд видов зеленых водорослей были экспериментально выведены для формирования более крупных скоплений. Когда Chlorella vulgaris выращивают вместе с хищником Ochromonas vallescia , она начинает образовывать небольшие колонии, которые труднее проглотить из-за большего размера. То же самое относится и к Chlamydomonas reinhardtii, находящимся под хищничеством Brachionus calyciflorus и Paramecium Tetraurelia .
C. reinhartii обычно начинается как подвижный одноклеточный побег ; эта единственная клетка бесполым образом размножается, проходя 2–5 раундов митоза в виде небольшого скопления неподвижных клеток, затем все клетки становятся одноклеточными пропагулами, и скопление растворяется. Через несколько поколений под воздействием хищников Paramecium «комок» становится устойчивой структурой: только некоторые клетки становятся размножениями. Некоторые популяции пошли еще дальше и развили многоклеточные побеги: вместо отделения отдельных клеток от комка, комок теперь размножается путем отделения более мелких комков. [56]
Преимущества
[ редактировать ]Многоклеточность позволяет организму превышать пределы размера, обычно налагаемые диффузией : отдельные клетки увеличенного размера имеют уменьшенное соотношение поверхности к объему и испытывают трудности с поглощением достаточного количества питательных веществ и транспортировкой их по клетке. Таким образом, многоклеточные организмы обладают конкурентными преимуществами увеличения размеров без каких-либо ограничений. Они могут иметь более длительную продолжительность жизни, поскольку могут продолжать жить, даже когда отдельные клетки умирают. Многоклеточность также позволяет увеличивать сложность, позволяя дифференцировать типы клеток внутри одного организма. [ нужна ссылка ]
Однако можно ли рассматривать все это как преимущества, остается спорным: подавляющее большинство живых организмов являются одноклеточными, и даже с точки зрения биомассы одноклеточные организмы гораздо более успешны, чем животные, хотя и не растения. [57] Вместо того, чтобы рассматривать такие черты, как более продолжительная продолжительность жизни и больший размер, как преимущество, многие биологи рассматривают их только как примеры разнообразия с соответствующими компромиссами. [ нужна ссылка ]
Экспрессия генов меняется при переходе от одно- к многоклеточности
[ редактировать ]В ходе эволюционного перехода от одноклеточных организмов к многоклеточным, вероятно, изменилась экспрессия генов, связанных с размножением и выживанием. [58] В одноклеточном состоянии гены, связанные с размножением и выживанием, экспрессируются таким образом, что повышается приспособленность отдельных клеток, но после перехода к многоклеточному состоянию характер экспрессии этих генов должен был существенно измениться, чтобы отдельные клетки стали более специализированными на их функция относительно воспроизводства и выживания. [58] С появлением многоклеточного организма паттерны экспрессии генов стали разделены между клетками, специализирующимися на размножении ( клетки зародышевой линии ), и клетками, специализирующимися на выживании ( соматические клетки ). По мере продвижения перехода специализированные клетки имели тенденцию терять свою индивидуальность и больше не могли ни выживать, ни воспроизводиться вне контекста группы. [58]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Беккер, Уэйн М.; и др. (2008). Мир клетки . Пирсон Бенджамин Каммингс . п. 480. ИСБН 978-0-321-55418-5 .
- ^ Чимилески, Скотт; Колтер, Роберто (2017). Жизнь на грани зрения: фотографическое исследование мира микробов . Издательство Гарвардского университета. ISBN 9780674975910 .
- ^ Перейти обратно: а б с Лайонс, Николас А.; Колтер, Роберто (апрель 2015 г.). «Об эволюции многоклеточности бактерий» . Современное мнение в микробиологии . 24 : 21–28. дои : 10.1016/j.mib.2014.12.007 . ISSN 1879-0364 . ПМК 4380822 . ПМИД 25597443 .
- ^ С.М. Миллер (2010). «Вольвокс, Хламидомонада и эволюция многоклеточности». Природное образование . 3 (9): 65.
- ^ Перейти обратно: а б Брайан Кейт Холл; Бенедикт Халлгримссон; Монро В. Стрикбергер (2008). Эволюция Стрикбергера: интеграция генов, организмов и популяций (4-е изд.). Холл/Халлгримссон. п. 149 . ISBN 978-0-7637-0066-9 .
- ^ Адл, Сина; и др. (октябрь 2005 г.). «Новая классификация эукариот более высокого уровня с акцентом на таксономию протистов» . Дж. Эукариот. Микробиол . 52 (5): 399–451. дои : 10.1111/j.1550-7408.2005.00053.x . ПМИД 16248873 . S2CID 8060916 .
- ^ Перейти обратно: а б Гросберг, РК; Стратманн, Р.Р. (2007). «Эволюция многоклеточности: незначительный крупный переход?» (PDF) . Annu Rev Ecol Evol Syst . 38 : 621–654. doi : 10.1146/annurev.ecolsys.36.102403.114735 .
- ^ Парфри, LW; Лар, DJG (2013). «Многоклеточность возникала несколько раз в эволюции эукариот» (PDF) . Биоэссе . 35 (4): 339–347. дои : 10.1002/bies.201200143 . ПМИД 23315654 . S2CID 13872783 .
- ^ Поппер, Зои А.; Мишель, Гурван; Эрве, Сесиль; Домозич, Дэвид С.; Уиллатс, Уильям Г.Т.; Туохи, Мария Г.; Клоарег, Бернард; Стенгель, Дагмар Б. (2011). «Эволюция и разнообразие стенок растительных клеток: от водорослей до цветковых растений». Ежегодный обзор биологии растений . 62 : 567–590. doi : 10.1146/annurev-arplant-042110-103809 . hdl : 10379/6762 . ПМИД 21351878 . S2CID 11961888 .
- ^ Перейти обратно: а б Никлас, KJ (2014). «Эволюционно-развитие истоков многоклеточности» . Американский журнал ботаники . 101 (1): 6–25. дои : 10.3732/ajb.1300314 . ПМИД 24363320 .
- ^ Боннер, Джон Тайлер (1998). «Истоки многоклеточности» (PDF) . Интегративная биология . 1 (1): 27–36. doi : 10.1002/(SICI)1520-6602(1998)1:1<27::AID-INBI4>3.0.CO;2-6 . ISSN 1093-4391 . Архивировано из оригинала 8 марта 2012 года.
{{cite journal}}: CS1 maint: неподходящий URL ( ссылка ) - ^ Маргулис, Л .; Чепмен, MJ (2009). Королевства и владения: иллюстрированный путеводитель по типам жизни на Земле (4-е изд.). Амстердам, Нидерланды: Академическая пресса / Elsevier. п. 116.
- ^ Серавин, Л.Н. (2001). «Принцип встречной морфологической эволюции и его значение для построения мегасистемы протистов и других эукариот» . Протистология . 2 :6–14.
- ^ Парфри, Л.В. и Лар, DJG (2013), с. 344.
- ^ Медина, М.; Коллинз, АГ; Тейлор, Дж.В.; Валентин, JW; Липпс, Дж. Х.; Зеттлер, Лос-Анджелес Амарал; Согин, М.Л. (2003). «Филогения Opisthokonta и эволюция многоклеточности и сложности у грибов и многоклеточных животных» . Международный журнал астробиологии . 2 (3): 203–211. Бибкод : 2003IJAsB...2..203M . дои : 10.1017/s1473550403001551 .
- ^ Зекбах, Джозеф, Чепмен, Дэвид Дж. [ред.]. (2010). Красные водоросли в эпоху генома . Нью-Йорк, штат Нью-Йорк, США: Springer, с. 252, [1] .
- ^ Кокуит, Э.; Вербрюгген, Х.; Лелиарт, Ф.; Де Клерк, О. (2010). «Эволюция и цитологическое разнообразие зеленых морских водорослей (Ulvophyceae)» . Мол. Биол. Эвол . 27 (9): 2052–2061. дои : 10.1093/molbev/msq091 . ISSN 0737-4038 . ПМИД 20368268 .
- ^ Рихтер, Дэниел Джозеф: Генный состав разнообразных хоанофлагеллят проливает свет на происхождение животных , 2013.
- ^ «Миксозоа» . tolweb.org . Проверено 14 апреля 2018 г.
- ^ Дэвис, PCW; Лайнуивер, Швейцария (2011). «Раковые опухоли как Metazoa 1.0: использование генов древних предков» . Физическая биология . 8 (1): 015001. Бибкод : 2011PhBio...8a5001D . дои : 10.1088/1478-3975/8/1/015001 . ПМК 3148211 . ПМИД 21301065 .
- ^ Домазет-Лосо, Т.; Таутц, Д. (2010). «Филостратиграфическое отслеживание раковых генов предполагает связь с появлением многоклеточности у многоклеточных животных» . БМК Биология . 8 (66): 66. дои : 10.1186/1741-7007-8-66 . ПМК 2880965 . ПМИД 20492640 .
- ^ Жак, Ф.; Баратчарт, Э.; Пиента, К.; Хаммарлунд, Э. (2022). «Происхождение и эволюция многоклеточности животных в свете филогеномики и генетики рака» . Медицинская онкология . 39 (160): 1–14. дои : 10.1007/s12032-022-01740-w . ПМЦ 9381480 . ПМИД 35972622 . .
- ^ Рихтер, DJ (2013), с. 11.
- ^ Гаспар, Т.; Хагеге, Д.; Кеверс, К.; Пенель, К.; Кревекёр, М.; Энгельманн, И.; Греппин, Х.; Фойдар, Дж. М. (1991). «Когда тератомы растений превращаются в рак в отсутствие патогенов». Физиология Плантарум . 83 (4): 696–701. дои : 10.1111/j.1399-3054.1991.tb02489.x .
- ^ Лаукнер, Г. (1980). Болезни простейших. В: Болезни морских животных . Кинн, О. (ред.). Том. 1, с. 84 , John Wiley & Sons, Чичестер, Великобритания.
- ^ Райкер, Эй Джей (1958). «Растительные опухоли: Введение» . Труды Национальной академии наук Соединенных Штатов Америки . 44 (4): 338–9. Бибкод : 1958PNAS...44..338R . дои : 10.1073/pnas.44.4.338 . ПМК 335422 . ПМИД 16590201 .
- ^ Дунан, Дж.; Хант, Т. (1996). «Клеточный цикл. Почему растения не болеют раком?» . Природа . 380 (6574): 481–2. дои : 10.1038/380481a0 . ПМИД 8606760 . S2CID 4318184 .
- ^ Ридли М (2004) Эволюция, 3-е издание. Блэквелл Паблишинг, с. 295–297.
- ^ Фэрклаф, Стивен Р.; Дайел, Марк Дж.; Кинг, Николь (26 октября 2010 г.). «Многоклеточное развитие хоанофлагелляты» . Современная биология . 20 (20): 875–876 р. Бибкод : 2010CBio...20.R875F . дои : 10.1016/j.cub.2010.09.014 . ПМЦ 2978077 . ПМИД 20971426 .
- ^ Кэрролл, Шон Б. (14 декабря 2010 г.). «Одноклеточный хищник: ключ к зарождению царства животных» . Нью-Йорк Таймс .
- ^ А. Х. Нолл, 2003. Жизнь на молодой планете . Издательство Принстонского университета. ISBN 0-691-00978-3 (твердый переплет), ISBN 0-691-12029-3 (мягкая обложка). Превосходная книга по ранней истории жизни, очень доступная неспециалисту; включает в себя обширные обсуждения ранних следов, окаменения и организации жизни.
- ^ Эль-Альбани, Абдерразак; и др. (1 июля 2010 г.). «Крупные колониальные организмы с скоординированным ростом в насыщенной кислородом среде 2,1 миллиарда лет назад». Природа . 466 (7302): 100–104. Бибкод : 2010Natur.466..100A . дои : 10.1038/nature09166 . ISSN 0028-0836 . ПМИД 20596019 . S2CID 4331375 .
- ^ Чен, Л.; Сяо, С.; Панг, К.; Чжоу, К.; Юань, X. (2014). «Дифференциация клеток и разделение зародышей и сомы в окаменелостях, похожих на эмбрионы эдиакарских животных». Природа . 516 (7530): 238–241. Бибкод : 2014Natur.516..238C . дои : 10.1038/nature13766 . ПМИД 25252979 . S2CID 4448316 .
- ^ Маргулис, Линн (1998). Симбиотическая планета: новый взгляд на эволюцию . Нью-Йорк: Основные книги . п. 160. ИСБН 978-0-465-07272-9 . Архивировано из оригинала 20 апреля 2010 г. Проверено 5 сентября 2017 г.
- ^ Хикман CP, Хикман FM (8 июля 1974 г.). Комплексные принципы зоологии (5-е изд.). Мосби . п. 112. ИСБН 978-0-8016-2184-0 .
- ^ Вулперт, Л.; Сатмари, Э. (2002). «Многоклеточность: Эволюция и яйцо» . Природа . 420 (6917): 745. Бибкод : 2002Natur.420..745W . дои : 10.1038/420745a . ПМИД 12490925 . S2CID 4385008 .
- ^ Кирк, Д.Л. (2005). «Программа из двенадцати шагов по развитию многоклеточности и разделения труда». Биоэссе . 27 (3): 299–310. doi : 10.1002/bies.20197 . ПМИД 15714559 .
- ^ База водорослей. Вольвокс Линней, 1758: 820.
- ^ Михайлов Кирилл В.; Константинова Анастасия В.; Никитин Михаил А.; Трошин Петр Владимирович; Русин Леонид Ю.; Любецкий, Василий А.; Панчин Юрий В.; Мыльников Александр П.; Мороз Леонид Л.; Кумар, Судхир; Алеошин, Владимир В. (2009). «Происхождение Metazoa: переход от временной к пространственной дифференциации клеток» (PDF) . Биоэссе . 31 (7): 758–768. doi : 10.1002/bies.200800214 . ПМИД 19472368 . S2CID 12795095 . Архивировано из оригинала (PDF) 5 марта 2016 г.
- ^ Эрвин, Дуглас Х. (9 ноября 2015 г.). «Ранняя жизнь многоклеточных животных: дивергенция, окружающая среда и экология» . Фил. Пер. Р. Сок. Б. 370 (20150036): 20150036. doi : 10.1098/rstb.2015.0036 . ПМК 4650120 . ПМИД 26554036 .
- ^ Циммер, Карл (7 января 2016 г.). «Генетический переворот помог организмам перейти от одной клетки к множеству» . Нью-Йорк Таймс . Проверено 7 января 2016 г.
- ^ Перейти обратно: а б Кунин, Е.В. (2016). «Вирусы и мобильные элементы как драйверы эволюционных переходов» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 371 (1701). дои : 10.1098/rstb.2015.0442 . ПМЦ 4958936 . ПМИД 27431520 .
- ^ Летцтер, Рафи (2 февраля 2018 г.). «Древний вирус может быть ответственен за человеческое сознание» . Живая наука . Проверено 05 сентября 2022 г.
- ^ Ми, С.; Ли, X.; Ли, Х.; Вельдман, генеральный директор; Финнерти, Х.; Рэйси, Л.; Лавалли, Э.; Тан, XY; Эдуард, П.; Хоуз, С.; Кейт-младший, Джей Си; Маккой, Дж. М. (2000). «Синцитин представляет собой пленный белок оболочки ретровируса, участвующий в морфогенезе плаценты человека» . Природа . 403 (6771): 785–789. Бибкод : 2000Natur.403..785M . дои : 10.1038/35001608 . ПМИД 10693809 . S2CID 4367889 .
- ^ Молер, Уильям А.; Шемер, Гиди; дель Кампо, Джейкоб Дж.; Валанси, Клари; Опоку-Серебуох, Евгений; Скрэнтон, Виктория; Ассаф, Нирит; Уайт, Джон Г.; Подбилевич, Бенджамин (март 2002 г.). «Мембранный белок типа I EFF-1 необходим для слияния клеток в процессе развития» . Развивающая клетка . 2 (3): 355–362. дои : 10.1016/s1534-5807(02)00129-6 . ISSN 1534-5807 . ПМИД 11879640 .
- ^ Перес-Варгас, Химена; Крей, Томас; Вэлэнс, Клэри; Авиноам, Ори; Хауз, Ахмед; Джамин, Марк; Раве-Барак, Хадас; Подбилевич, Бенджамин; Рей, Феликс А. (2014). «Структурные основы слияния эукариотических клеток» . Клетка . 157 (2): 407–419. дои : 10.1016/j.cell.2014.02.020 . ПМИД 24725407 .
- ^ Слезак, Майкл (2016), «Нет вирусов? Ни кожи, ни костей» (New Scientist, № 2958, 1 марта 2014 г.), стр. 16
- ^ Нурсолл, младший (апрель 1959 г.). «Кислород как предпосылка происхождения многоклеточных животных» . Природа . 183 (4669): 1170–1172. Бибкод : 1959Natur.183.1170N . дои : 10.1038/1831170b0 . ISSN 1476-4687 . S2CID 4200584 .
- ^ Миллс, Д.Б.; Уорд, LM; Джонс, К.; Суитен, Б.; Форт, М.; Треуш, А.Х.; Кэнфилд, Делавэр (18 февраля 2014 г.). «Потребность в кислороде древнейших животных» . Труды Национальной академии наук . 111 (11): 4168–4172. Бибкод : 2014PNAS..111.4168M . дои : 10.1073/pnas.1400547111 . ISSN 0027-8424 . ПМЦ 3964089 . ПМИД 24550467 .
- ^ Лайонс, Тимоти В.; Дрозер, Мэри Л.; Лау, Кимберли В.; Портер, Сюзанна М.; Сяо, Шухай; Тан, Цин (28 сентября 2018 г.). «После скучного миллиарда и до замерзания миллионов: эволюционные закономерности и инновации в тонианский период» . Новые темы в науках о жизни . 2 (2): 161–171. дои : 10.1042/ETLS20170165 . hdl : 10919/86820 . ISSN 2397-8554 . ПМИД 32412616 . S2CID 90374085 .
- ^ Брокс, Йохен Дж.; Джарретт, Эмбер Дж. М.; Сирантуан, Ева; Халлманн, Кристиан; Хосино, Ёске; Лиянаге, Тарика (август 2017 г.). «Рост водорослей в криогенных океанах и появление животных» . Природа . 548 (7669): 578–581. Бибкод : 2017Natur.548..578B . дои : 10.1038/nature23457 . ISSN 1476-4687 . ПМИД 28813409 . S2CID 205258987 .
- ^ Херрон, Мэтью Д.; Борин, Джошуа М.; Босуэлл, Джейкоб К.; Уокер, Джиллиан; Чен, И.-Чен Кимберли; Нокс, Чарльз А.; Бойд, Маргрет; Розенцвейг, Фрэнк; Рэтклифф, Уильям К. (20 февраля 2019 г.). «Происхождение de novo многоклеточности в ответ на хищничество» . Научные отчеты . 9 (1): 2328. Бибкод : 2019NatSR...9.2328H . дои : 10.1038/s41598-019-39558-8 . ISSN 2045-2322 . ПМЦ 6382799 . ПМИД 30787483 .
- ^ Смукалла, Скотт; Кальдара, Марина; Почет, Натали; Бове, Энн; Гуаданьини, Стефани; Ян, Чен; Винсес, Марсело Д.; Янсен, Ан; Прево, Мари Кристина; Латже, Жан-Поль; Финк, Джеральд Р.; Фостер, Кевин Р.; Верстрепен, Кевин Дж. (14 ноября 2008 г.). «FLO1 — это изменчивый ген зеленой бороды, который стимулирует сотрудничество почкующихся дрожжей, подобное биопленке» . Клетка . 135 (4): 726–737. дои : 10.1016/j.cell.2008.09.037 . ISSN 1097-4172 . ПМК 2703716 . ПМИД 19013280 .
- ^ Уд, Барт; Гуадалупе-Медина, Виктор; Нейкамп, Юрген Ф.; Де Риддер, Дик; Пронк, Джек Т.; Ван Марис, Антониус Дж.А.; Даран, Жан-Марк (2013). «Дупликация генома и мутации в ACE2 вызывают многоклеточные фенотипы с быстрым оседанием у эволюционировавших Saccharomyces cerevisiae » . Труды Национальной академии наук . 110 (45): Е4223-31. Бибкод : 2013PNAS..110E4223O . дои : 10.1073/pnas.1305949110 . ПМЦ 3831460 . ПМИД 24145419 .
- ^ Боздаг, Г. Озан; Замани-Дахадж, Сейед Алиреза; Дэй, Томас С.; Кан, Пенелопа К.; Бернетти, Энтони Дж.; Лак, Дунг Т.; Тонг, Кай; Конлин, Питер Л.; Балвани, Айшвария Х.; Дайер, Ева Л.; Юнкер, Питер Дж.; Рэтклифф, Уильям К. (25 мая 2023 г.). «Эволюция de novo макроскопической многоклеточности» . Природа . 617 (7962): 747–754. Бибкод : 2023Natur.617..747B . дои : 10.1038/s41586-023-06052-1 . ISSN 0028-0836 . ПМЦ 10425966 . ПМИД 37165189 . S2CID 236953093 .
- ^ Херрон, Мэтью Д.; Борин, Джошуа М.; Босуэлл, Джейкоб К.; Уокер, Джиллиан; Чен, И-Чен Кимберли; Нокс, Чарльз А.; Бойд, Маргрет; Розенцвейг, Фрэнк; Рэтклифф, Уильям К. (2019). «Происхождение de novo многоклеточности в ответ на хищничество» . Научные отчеты . 9 (1): 2328. Бибкод : 2019NatSR...9.2328H . дои : 10.1038/s41598-019-39558-8 . ПМК 6382799 . ПМИД 30787483 .
- ^ Бар-Он, Инон М.; Филлипс, Роб; Майло, Рон (19 июня 2018 г.). «Распределение биомассы на Земле» . ПНАС . 115 (25): 6506–6511. Бибкод : 2018PNAS..115.6506B . дои : 10.1073/pnas.1711842115 . ПМК 6016768 . ПМИД 29784790 .
- ^ Перейти обратно: а б с Грочау-Райт З.И., Недельку А.М., Мишод Р.Э. Генетика реорганизации фитнеса при переходе к многоклеточности: regA-подобное семейство Volvocine как модель. Гены (Базель). 2023, 19 апреля;14(4):941. doi: 10.3390/genes14040941. ПМИД 37107699; PMCID: PMC10137558
Внешние ссылки
[ редактировать ]- Древо жизни эукариотов . Архивировано 29 января 2012 г. в Wayback Machine .