Барьер Вейсмана
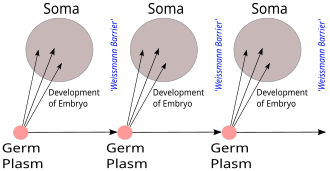
Барьер Вейсмана , предложенный Августом Вейсманом , представляет собой строгое различие между «бессмертными» линиями зародышевых клеток, производящих гаметы, и «одноразовыми» соматическими клетками у животных (но не растений), в отличие от Чарльзом Дарвином предложенного механизма пангенезиса для наследования. [1] [2]
Если говорить более точно, наследственная информация копируется только из зародышевых клеток в соматические клетки . Это означает, что новая информация от соматической мутации не передается зародышевой линии. Эта концепция барьера подразумевает, что соматические мутации не передаются по наследству. [3] [4]
Вейсман изложил эту концепцию в своей книге 1892 года «Das Keimplasma: einer Theorie der Vererbung» (по-немецки « Зародышевая плазма : теория наследования »). [5] Использование этой теории, обычно в контексте теории зародышевой плазмы конца 19-го века, до разработки более обоснованных и более сложных концепций генетики в начале 20-го века, иногда называют вейсманизмом . [6] Некоторые авторы отличают вейсманистское развитие ( преформистское или эпигенетическое ), при котором имеется отчетливая зародышевая линия, от соматического эмбриогенеза . [7] Этот тип развития коррелирует с эволюцией гибели соматической линии.
Барьер Вейсмана имел большое значение в свое время и, среди прочего, эффективно изгонял некоторые концепции Ламарка : в частности, он делал ламарковское наследование изменений в теле (соме) трудным или невозможным. [8] Это остается важным, но требует уточнения в свете современного понимания горизонтального переноса генов и некоторых других генетических и гистологических разработок. [9]
Бессмертие зародышевой линии
[ редактировать ]Российский биолог и историк Жорес А. Медведев , рассматривая теорию Вейсмана столетие спустя, считал, что точность репликации генома и других синтетических систем сама по себе не может объяснить «бессмертные» линии зародышевых клеток, предложенные Вейсманом. Скорее Медведев считал, что известные особенности биохимии и генетики полового размножения указывают на наличие уникальных процессов сохранения и восстановления информации на разных стадиях гаметогенеза . В частности, Медведев считал, что важнейшие возможности информационного обеспечения половых клеток создаются за счет рекомбинации при мейозе и репарации ДНК ; он рассматривал их как процессы внутри зародышевых клеток, которые были способны восстанавливать целостность ДНК и хромосом от тех типов повреждений, которые вызывали необратимое старение соматических клеток . [10]
Базальные животные
[ редактировать ]Базальные животные , такие как губки ( Porifera ) и кораллы ( Anthozoa ), содержат линии мультипотентных стволовых клеток, которые дают начало как соматическим, так и репродуктивным клеткам. Барьер Вейсмана, по-видимому, возник у животных более позднего эволюционного происхождения. [11]
Растения
[ редактировать ]У растений генетические изменения в соматических линиях могут привести и действительно приводят к генетическим изменениям в зародышевых линиях, поскольку зародышевые клетки производятся линиями соматических клеток (вегетативные меристемы ), которые могут быть достаточно старыми (много лет), чтобы с тех пор накопились множественные мутации. прорастание семян, некоторые из них подвергаются естественному отбору. [12] В этом контексте примечательно, что, вообще говоря, взрослые размножающиеся растения имеют тенденцию давать гораздо больше потомков, чем животные организмы.
См. также
[ редактировать ]- Альтернативы эволюции путем естественного отбора - Список альтернатив дарвиновскому естественному отбору.
- Эффект Болдуина - Влияние приобретенного поведения на эволюцию.
- Трансгенерационное эпигенетическое наследование - эпигенетическая передача без изменения первичной структуры ДНК.
- Ламаркизм - Научная гипотеза о наследственности.
- Пангенезис - предложенный Дарвином механизм наследственности.
Ссылки
[ редактировать ]- ^ Гейсон, Г.Л. (1969). «Дарвин и наследственность: эволюция его гипотезы пангенезиса». J Hist Med Allied Sci . XXIV (4): 375–411. дои : 10.1093/jhmas/XXIV.4.375 . ПМИД 4908353 .
- ^ Ты, Явен (26 января 2015 г.). «Зародышевая плазма: теория наследственности (1893 г.), Август Вейсманн» . Энциклопедия проекта «Эмбрион» (Университет штата Аризона) . Проверено 7 сентября 2018 г.
- ^ Готье, Питер (март – май 1990 г.). «Является ли эксперимент Вейсмана опровержением гипотезы Ламарка?». БИОС . 61 (1/2): 6–8. JSTOR 4608123 .
- ^ Это не относится к центральной догме молекулярной биологии , которая утверждает, что никакая последовательная информация не может передаваться от белка к ДНК или РНК , но обе гипотезы относятся к геноцентрическому взгляду на жизнь. Де Тиге, Алексис; Танхе, Коэн; Брэкман, Йохан; Ван де Пер, Ив (январь 2014 г.). «От ДНК-к NA-центризму и новые условия геноцентризма». Биология и философия . 29 (1): 55–69. дои : 10.1007/s10539-013-9393-z . S2CID 85866639 .
- ^ Вейсман, август (1892 г.). Зародышевая плазма: теория наследственности . Йена: Рыбак.
- ^ Романес, Джордж Джон (1893). Исследование вейсманизма . Чикаго: Открытый суд. ОЛ 23380098М .
- ^ Ридли, Марк (2004). Эволюция (3-е изд.). Блэквелл. стр. 295–297.
- ^ Романес, Джордж Джон (1893). Исследование вейсманизма . Открытый суд. ОЛ 23380098М .
- ^ Линдли, Робин А. «Как мутационные и эпигенетические изменения способствуют адаптивной эволюции» . Лабораторный журнал ГИТ .
- ^ Медведев, Жорес А. (1981). «О бессмертии зародышевой линии: генетические и биохимические механизмы. Обзор». Механизмы старения и развития . 17 (4): 331–359. дои : 10.1016/0047-6374(81)90052-X . ISSN 0047-6374 . ПМИД 6173551 . S2CID 35719466 .
- ^ Радзвилавичюс, Арунас Л.; Хадживасилиу, Зена; Помянковский, Эндрю; Лейн, Ник (20 декабря 2016 г.). «Отбор по качеству митохондрий стимулирует эволюцию зародышевой линии» . ПЛОС Биология . 14 (12): e2000410. дои : 10.1371/journal.pbio.2000410 . ISSN 1545-7885 . ПМК 5172535 . ПМИД 27997535 .
- ^ Уизем, Т.Г.; Слободчиков, CN (1981). «Эволюция особей, взаимодействие растений и травоядных и мозаика генетической изменчивости: адаптивное значение соматических мутаций у растений». Экология . 49 (3): 287–292. Бибкод : 1981Oecol..49..287W . дои : 10.1007/BF00347587 . ПМИД 28309985 . S2CID 20411802 .