Слизистая плесень

Слизевик или слизевик — неофициальное название, данное полифилетическому скоплению неродственных эукариотических организмов в Stramenopiles , Rhizaria , Discoba , Amoebozoa и Holomycota кладах . Большинство из них микроскопические; те, что обитают в Myxogastria, образуют более крупные плазмодии слизевиков, видимые невооруженным глазом. слизевика Жизненный цикл включает стадию свободноживущей одноклеточной формы и образование спор. Споры часто образуются в макроскопических многоклеточных или многоядерных плодовых телах, которые могут образовываться путем агрегации или слияния; агрегация управляется химическими сигналами, называемыми акразинами . Слизевики способствуют разложению отмершей растительности; некоторые являются паразитическими .
Большинство слизевиков являются наземными и свободноживущими, обычно во влажных тенистых местах обитания, например, внутри или на поверхности гниющей древесины. Некоторые миксогастрийцы и протостелии ведут водный или полуводный образ жизни. Фитомиксеи являются паразитами и живут внутри своих растений- хозяев . Географически слизевики космополитично распространены . Небольшое количество видов встречается в таких засушливых регионах, как пустыня Атакама , и в таких холодных регионах, как Арктика ; их много в тропиках , особенно в тропических лесах .
Слизевики обладают разнообразным поведением, которое можно наблюдать у животных с мозгом. Такие виды, как Physarum polycephalum, использовались для моделирования транспортных сетей. Некоторые виды традиционно употреблялись в пищу в таких странах, как Эквадор.
Эволюция
[ редактировать ]Таксономическая история
[ редактировать ]
Первым отчетом о слизевиках было Томасом Панкоу в 1654 году обсуждение Lycogala epidendrum . Он назвал его Fungus cito crescentes , «быстрорастущий гриб». [2] [1]
Немецкий миколог Генрих Антон де Бари в 1860 и 1887 годах классифицировал Myxomycetes (плазмодиальные слизевики) и Acrasieae (клеточные слизевики) как Mycetozoa, новый класс. Он также представил раздел «Сомнительные Mycetozoa» для Plasmodiophora (теперь в Phytomyxea ) и Labyrinthula , подчеркнув их отличие от растений и грибов. [3] [4] В 1880 году французский ботаник Филипп ван Тигем дополнительно проанализировал эти две группы. [4] В 1868 году немецкий биолог Эрнст Геккель поместил Mycetozoa в королевство, которое он назвал Protista . [4] В 1885 году британский зоолог Рэй Ланкестер сгруппировал Mycetozoa вместе с Proteomyxa как часть Gymnomyxa в типе Protozoa . [4] Артур и Гулиэльма Листер опубликовали монографии группы в 1894, 1911 и 1925 годах. [5] [6]
В 1932 и 1960 годах американский миколог Джордж Уиллард Мартин утверждал, что слизевики произошли от грибов. [7] [8] В 1956 году американский биолог Герберт Коупленд поместил Mycetozoa (миксомицеты и плазмодиофориды) и Sarkodina (лабиринтулиды и клеточные слизевики) в тип под названием Protoplasta, который он поместил рядом с грибами и водорослями в новое царство Protoctista. . [4] [9]
В 1969 году систематик Р. Х. Уиттакер заметил, что слизевики были очень заметны и различимы внутри группы грибов, к которой их затем отнесли. Он согласился с предложением Линдси С. Олив реклассифицировать Gymnomycota, включающую слизевики, в категорию Protista. [10] Уиттекер поместил три типа, а именно Myxomycota, Acrasiomycota и Labyrinthulomycota, в подцарство Gymnomycota внутри грибов. [4] В том же году Мартин и Алексопулос опубликовали свой влиятельный учебник «Миксомицеты» . [6]
В 1975 году Олив выделил диктиостелид и акрасид в отдельные группы. [4] В 1992 г. Дэвид Дж. Паттерсон и М.Л. Согин предположили, что диктиостелиды разошлись раньше растений, животных и грибов. [11]
Филогения
[ редактировать ]Слизевики практически не имеют ископаемой истории, как и следовало ожидать, учитывая их небольшие размеры и мягкое тело. [12] Эта группировка полифилетическая , состоящая из множества клад (выделенных на филогенетическом дереве ), широко разбросанных по эукариотам. Парафилетические группы показаны в кавычках: [13] [14]
| Эукариоты |
| |||||||||||||||||||||||||||

Разнообразие
[ редактировать ]Различные оценки количества видов слизевиков сходятся в том, что существует около 1000 видов, большинство из которых — Myxogastria . Коллекция ДНК окружающей среды дает более высокую оценку - от 1200 до 1500 видов. [6] Они разнообразны как таксономически, так и по внешнему виду, причем самый крупный и наиболее знакомый вид принадлежит Myxogasteria. Наиболее часто встречающимися формами роста являются спорангии , спорообразующие тела, часто имеющие примерно сферическую форму; они могут находиться непосредственно на поверхности, например, на гниющей древесине, или могут находиться на тонком стебле, который поднимает споры для высвобождения над поверхностью. У других видов споры собраны в большую массу, которую насекомые могут посещать в пищу; они рассеивают споры, когда уходят. [15]
Макроскопические плазмодиальные слизевики: Миксогастрия.
[ редактировать ]Миксогастрия, или плазмодии, являются единственными макроскопического масштаба слизевиками ; они дали группе неофициальное название, поскольку часть своего жизненного цикла они скользкие на ощупь. [16] Миксогастрий состоит из крупной клетки с тысячами ядер внутри одной мембраны без стенок, образующих синцитий . [17] Большинство из них меньше нескольких сантиметров, но некоторые виды могут достигать размеров до нескольких квадратных метров, а в случае Brefeldia maxima - массы до 20 килограммов (44 фунта). [18] [19] [20]
 Стемонит показывает стебельчатые спорангии для распространения спор по воздуху.
Стемонит показывает стебельчатые спорангии для распространения спор по воздуху.
 Клетки Fuligo septica объединяются, образуя мягкую массу.
Клетки Fuligo septica объединяются, образуя мягкую массу.
![Спорангий Enteridium lycoperdon. Споры могут распространяться в воздухе или воде, а также с помощью слизевиковых мух.[21]](//upload.wikimedia.org/wikipedia/commons/thumb/c/c2/Enteridium_lycoperdon%2C_%28Bull.%29_M.L._Farr%2C_1976_%28Reticularia_lycoperdon%29_%28cropped%29.JPG/175px-Enteridium_lycoperdon%2C_%28Bull.%29_M.L._Farr%2C_1976_%28Reticularia_lycoperdon%29_%28cropped%29.JPG) Enteridium lycoperdon Спорангий . Споры могут распространяться в воздухе или воде, а также с помощью мух-слизевиков. [21]
Enteridium lycoperdon Спорангий . Споры могут распространяться в воздухе или воде, а также с помощью мух-слизевиков. [21] Metatrichia vesparium имеет небольшие круглые спорангии со спиральными выступами , позволяющими выбрасывать крышки и рассеивать споры. [22]
Metatrichia vesparium имеет небольшие круглые спорангии со спиральными выступами , позволяющими выбрасывать крышки и рассеивать споры. [22] Ракообразные Mucilago, агрегирующие из струящегося плазмодия (сеть нитей) в спорангий (большая масса)
Ракообразные Mucilago, агрегирующие из струящегося плазмодия (сеть нитей) в спорангий (большая масса)




![Спорангий Enteridium lycoperdon. Споры могут распространяться в воздухе или воде, а также с помощью слизевиковых мух.[21]](http://upload.wikimedia.org/wikipedia/commons/thumb/c/c2/Enteridium_lycoperdon%2C_%28Bull.%29_M.L._Farr%2C_1976_%28Reticularia_lycoperdon%29_%28cropped%29.JPG/175px-Enteridium_lycoperdon%2C_%28Bull.%29_M.L._Farr%2C_1976_%28Reticularia_lycoperdon%29_%28cropped%29.JPG)


Клеточные слизевики: Dictyosteliida.
[ редактировать ]Dictyosteliida , или клеточная слизевика не образуют огромных ценоцитов как Myxogastria; их амебы большую часть жизни остаются индивидуальными как отдельные одноклеточные протисты , питающиеся микроорганизмами. Когда пища исчерпана и они готовы образовывать спорангии, они образуют рой. Амебы — объединяются в крошечного многоклеточного слизняка, который выползает на открытое освещенное место и вырастает в плодовое тело сорокарпия . Некоторые из амеб становятся спорами, чтобы дать начало следующему поколению, но другие жертвуют собой, становясь мертвым стеблем, поднимая споры в воздух. [23] [24]
 Dictyostelium discoideum — микроскопический организм. Клетки могут объединяться, образуя грекса или слизняка, а затем в сорокарпия или плодовое тело (показано) на нежном стебле.
Dictyostelium discoideum — микроскопический организм. Клетки могут объединяться, образуя грекса или слизняка, а затем в сорокарпия или плодовое тело (показано) на нежном стебле.

Протостелида
[ редактировать ]Protosteliida , полифилетическая группа, имеют промежуточные признаки между двумя предыдущими группами, но они намного меньше, а плодовые тела образуют лишь от одной до нескольких спор . [25]
 Ceratiomyxa микроскопическая; на каждом стебле находится только одна или очень мало спор.
Ceratiomyxa микроскопическая; на каждом стебле находится только одна или очень мало спор.

Копромикса
[ редактировать ]Лобозаны слизевики , парафилетическая группа амеб, включают . Copromyxa [26] [27]
Неамебозойные слизевики
[ редактировать ]К неамебозойным слизевикам относятся Acrasids , у которых есть слизеподобные амебы. амеб При передвижении псевдоподии являются эруптивными, то есть спереди появляются полусферические выпуклости. [28] Phytomyxea — облигатные паразиты , хозяевами которых являются растения, диатомовые водоросли , оомицеты и бурые водоросли . Они вызывают такие заболевания растений, как клубень капусты и мучнистую паршу . [29] Лабиринтуломицеты представляют собой сети морской слизи, образующие лабиринтные сети трубок, по которым могут перемещаться амебы без ложноножек. [30] Fonticulida — это клеточные слизевики, образующие плодовое тело в форме «вулкана». [31]
 Лабиринтуломицет — Aplanochytrium морской протист.
Лабиринтуломицет — Aplanochytrium морской протист.

Распространение, среда обитания и экология.
[ редактировать ]
Слизевики из-за их небольшого размера и влажной поверхности обитают в основном во влажных местах обитания, включая затененные леса, гниющую древесину, опавшие или живые листья, а также на мохообразных . [32] [18] Большинство миксогастрий являются наземными. [18] хотя некоторые из них, например Didymium aquatilis , ведут водный образ жизни. [33] [34] а D. nigripes ведет полуводный образ жизни. [34] Миксогастрия не ограничивается влажными регионами; Из Саудовской Аравии известно 34 вида, обитающих на коре, в растительной подстилке и гниющей древесине даже в пустынях . [35] Они также встречаются в пустыне Сонора в Аризоне (46 видов) и в исключительно сухой пустыне Атакама в Чили (24 вида). Напротив, в полузасушливом биосферном заповеднике Теуакан-Куикатлан в России и Казахстане насчитывается 105 видов, а в бассейне рек Волги - 158 видов. [35] В тропических лесах Латинской Америки такие виды, как Arcyria и Didymium, обычно являются эпифиллярными и растут на листьях печеночников . [36]
Диктиостелиды в основном ведут наземный образ жизни. [37] На горе Чанбайшань в Китае шесть видов диктиостелид были обнаружены в лесных почвах на высоте до 2038 м (6686 футов), причем самым высоким зарегистрированным видом там был Dictyostelium mucoroides . [38] Протостелиды живут в основном на отмерших растительных веществах, где они потребляют споры бактерий , дрожжей и грибов . [37] К ним относятся некоторые водные виды, которые живут на мертвых частях растений, погруженных в пруды. [33] Клеточные слизевики наиболее многочисленны в тропиках, их количество уменьшается с широтой , но они распространены космополитно и встречаются в почве даже в Арктике и Антарктике. [39] В тундре Аляски единственными слизевиками являются диктиостелиды D. mucoroides и D. sphaerocephalum . [36]
Виды Copromyxa копрофилы , питаются навозом. [27]
Споры некоторых миксогастрийцев разносятся животными. Слизевиковая муха Epicypta testata откладывает яйца в споровую массу Enteridium lycoperdon , которой питаются личинки. Они окукливаются, а вылупившиеся взрослые особи переносят и рассеивают прилипшие к ним споры. [21] В то время как различные насекомые поедают слизевиков, жуки-слизевики Sphindidae , как личинки, так и взрослые особи, питаются исключительно ими. [40]
Жизненный цикл
[ редактировать ]Плазмодиальные слизевики
[ редактировать ]
Плазмодиальные слизевики начинают жизнь амебоподобные как клетки . Эти одноклеточные амебы обычно гаплоидны и питаются мелкой добычей, такой как бактерии , дрожжевые клетки и споры грибов, путем фагоцитоза , поглощая их своей клеточной мембраной . Эти амебы могут спариваться, если встречаются с правильным типом спаривания , и образуют зиготы , которые затем превращаются в плазмодии . Они содержат множество ядер без клеточных мембран между ними и могут достигать метров в размерах. Вид Fuligo septica часто можно увидеть в виде слизистой желтой сети внутри и на гниющих бревнах. Амебы и плазмодии поглощают микроорганизмы. [41] Плазмодий превращается в взаимосвязанную сеть протоплазматических нитей. [42] Внутри каждой протоплазматической цепи цитоплазматическое содержимое быстро течет, периодически меняя направление. Поток протоплазмы внутри цепи плазмодия может достигать скорости до 1,35 мм в секунду у Physarum multicephalum , самой быстрой скорости для любого микроорганизма. [43]
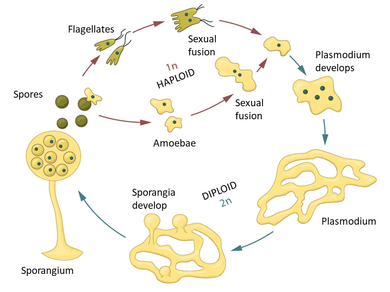
Слизевики изогамны , что означает, что их гаметы (репродуктивные клетки) имеют одинаковый размер, в отличие от яйцеклеток и сперматозоидов животных. [45] Physarum polycephalum имеет три гена, участвующих в размножении: mat A и mat B, каждый с тринадцатью вариантами, и mat C с тремя вариантами. Каждая репродуктивно зрелая слизевика диплоидна , то есть содержит по две копии каждого из трех репродуктивных генов. [46] Когда P. polycephalum готов к образованию репродуктивных клеток, у него вырастает луковицеобразное расширение тела, вмещающее их. [47] Каждая клетка имеет случайную комбинацию генов, которые слизевики содержат в своем геноме . Таким образом, он может создавать клетки, содержащие до восьми различных типов генов. Освободившиеся клетки затем независимо ищут другую совместимую клетку для слияния. Другие особи P. polycephalum могут содержать различные комбинации генов mat A, mat B и mat C, что допускает более 500 возможных вариаций. Для организмов с этим типом репродуктивных клеток выгодно иметь множество типов спаривания, поскольку вероятность того, что клетки найдут партнера, значительно увеличивается, а риск инбридинга резко снижается. [46]
Клеточные слизевики
[ редактировать ]Клеточные слизевики представляют собой группу, насчитывающую около 150 описанных видов. Встречаются преимущественно в гумусном слое лесных почв. [48] и питаются бактериями, но также встречаются в навозе животных и на сельскохозяйственных полях. Они существуют как одноклеточные организмы, пока еды много. Когда еды не хватает, многие одноклеточные амебы собираются и начинают двигаться как единое тело, называемое «слизнем». Способность одноклеточных организмов объединяться в многоклеточные формы является причиной того, что их еще называют социальными амебами. В этом состоянии они чувствительны к химическим веществам, переносимым по воздуху, и могут обнаруживать источники пищи. Они легко меняют форму и функции частей и могут образовывать стебли, из которых образуются плодовые тела, выделяющие бесчисленные споры, достаточно легкие, чтобы их можно было переносить ветром или проплывающими животными. [23] Клеточная слизевика Dictyostelium discoideum имеет множество различных типов спаривания. Когда этот организм вступил в стадию размножения, он выделяет химический аттрактант. [49] Когда приходит время слияния клеток, у Dictyostelium discoideum есть собственные типы спаривания, которые определяют, какие клетки совместимы друг с другом. Существует как минимум одиннадцать типов спаривания; макроцисты образуются после контакта клеток между совместимыми типами спаривания. [50]
Химические сигналы
[ редактировать ]
Химические вещества, которые агрегируют клеточные слизевики, представляют собой небольшие молекулы, называемые акразинами ; Движение навстречу химическому сигналу называется хемотаксисом . Первым открытым акрасином был циклический аденозинмонофосфат (циклический АМФ), общая сигнальная молекула клеток, у Dictyostelium discoideum . Во время фазы агрегации своего жизненного цикла амебы Dictyostelium discoideum общаются друг с другом, используя бегущие волны циклического АМФ. [51] [52] [53] Когда они агрегируются, происходит усиление циклического АМФ. [54] Предстебельковые клетки движутся в сторону циклического АМФ, но предспоровые клетки игнорируют сигнал. [55] Существуют и другие акрасины; акразин Polysphondylium violaceum , очищенный в 1983 году, представляет собой дипептид глорин. [56] Ионы кальция также служат для привлечения амеб слизевиков, по крайней мере, на коротких расстояниях. Было высказано предположение, что акразины могут быть таксонспецифичными, поскольку специфичность необходима для формирования агрегации генетически сходных клеток. Многие виды диктиостелид действительно не реагируют на циклический АМФ, но по состоянию на 2023 год их акразины оставались неизвестными. [57]
Изучать
[ редактировать ]Использование в исследованиях и преподавании
[ редактировать ]Практическому изучению слизевиков способствовало введение в 1933 году Х.К. Гилбертом и Г.В. Мартином «влажной культуральной камеры» . [58] Слизевики можно использовать для изучения конвергентной эволюции , поскольку привычка образовывать стебель со спорангием, который может выпускать споры в воздух, над землей, неоднократно развивалась, например, у миксогастрии (эукариоты) и миксобактерий ( прокариоты ). [59] Кроме того, как (макроскопические) диктиостелиды, так и (микроскопические) протостелиды имеют фазу с подвижными амебами и фазу со ножкой; у протостелид стебель крошечный, на нем находится только одна спора, но логика распространения спор по воздуху та же. [59]
О. Р. Коллинз показал, что слизевик Didymium iridis имеет два штамма (+ и -) клеток, эквивалентных гаметам, что они могут образовывать бессмертные клеточные линии в культуре и что система контролируется аллелями одного гена. Это сделало этот вид модельным организмом для изучения несовместимости, бесполого размножения и типов спаривания. [59]
Биохимия
[ редактировать ]Слизевики изучались на предмет производства необычных органических соединений, включая пигменты , антибиотики и противораковые препараты . [59] Пигменты включают нафтохиноны , физарохром А и соединения тетраминовой кислоты. Бисиндолилмалеимиды , продуцируемые Arcyria denudata, включают некоторые фосфоресцирующие соединения. [60] Спорофоры (плодовые тела) Arcyria denudata окрашены в красный цвет арцириафлавинами A–C, которые содержат необычное индоло[2,3- а ] карбазольное алкалоидное кольцо. [61] К 2022 году из слизевиков было выделено более 100 пигментов, в основном из спорофоров. Было высказано предположение, что многие пигменты от желтого до красного могут быть полезны в косметике . [15] Около 42% пациентов с сезонным аллергическим ринитом реагировали на споры миксогастрии, поэтому споры могут вносить значительный вклад в качестве аллергенов, передающихся по воздуху . [62]
Вычисление
[ редактировать ]Слизевики имеют некоторое сходство с нервной системой животных. [63] Мембраны как слизевиков, так и нервных клеток содержат рецепторные участки, которые изменяют электрические свойства мембраны при ее связывании. [64] Поэтому некоторые исследования ранней эволюции нервных систем животных вдохновлены слизевиками. [65] [66] [67] Когда масса или холмик слизевика физически разделяется, клетки возвращаются обратно, чтобы воссоединиться. Исследования Physarum polycephalum даже показали, что организм обладает способностью обучаться и прогнозировать периодические неблагоприятные условия в лабораторных экспериментах. [68] Джон Тайлер Боннер , профессор экологии, известный своими исследованиями слизевиков, утверждает, что они «не более чем мешок амеб, заключенный в тонкую слизистую оболочку, однако им удается вести себя так же, как и животные, которые обладают мышцами и нервами с ганглиями – то есть простым мозгом». [69]
слизевиков Алгоритм — это метаэвристический алгоритм , основанный на поведении агрегированных слизевиков в процессе поиска пищи. Он описывается как простой, эффективный и гибкий способ решения задач оптимизации , таких как поиск кратчайшего пути между узлами в сети. Однако он может попасть в ловушку локального оптимума . [70]
Тосиюки Накагаки и его коллеги изучают слизевики и их способность решать лабиринты, размещая узлы в двух точках, разделенных лабиринтом из пластиковой пленки. Форма исследовала все возможные пути и нашла кратчайший путь. [71]
Вдохновение для дорожной системы
[ редактировать ]
Ацуши Теро и его коллеги выращивали Physarum в плоской влажной чашке, поместив форму в центральное положение, символизируя Токио, а окружающие ее овсяные хлопья соответствовали расположению других крупных городов в районе Большого Токио. Поскольку Physarum избегает яркого света, свет использовался для имитации гор, воды и других препятствий в чашке. Плесень сначала плотно заполнила пространство плазмодиями, а затем истончила сеть, чтобы сосредоточить внимание на эффективно соединенных ветвях. Сеть очень напоминала железнодорожную систему Токио . [72] [73] P. polycephalum использовался в экспериментальных лабораторных аппроксимациях сетей автомагистралей в 14 географических регионах: Австралии, Африке, Бельгии, Бразилии, Канаде, Китае, Германии, Иберии, Италии, Малайзии, Мексике, Нидерландах, Великобритании и США. [74] [75] [76] Нитевидная структура P. polycephalum, образующая сеть источников пищи, аналогична крупномасштабной структуре галактических нитей во Вселенной . Это наблюдение побудило астрономов использовать моделирование, основанное на поведении слизевиков, для поиска темной материи . [77] [78]
Использовать в пищу
[ редактировать ]В центральной Мексике ложный дождевик Enteridium lycoperdon традиционно использовался в пищу; это был один из видов, которые собиратели грибов или хонгеро собирали во время поездок в лес в сезон дождей. Одно из его местных названий — «сырный гриб», названный так за текстуру и вкус при приготовлении. Его солили, заворачивали в кукурузный лист и запекали в золе костра; или варят и едят с кукурузными лепешками . Fuligo septica аналогичным образом собирали в Мексике, готовили с луком и перцем и ели в лепешке. В Эквадоре Lycogala epidendrum называли «якич» и ели в сыром виде в качестве закуски. [79]
В популярной культуре
[ редактировать ]Оскар Рекехо и Н. Флоро Андрес-Родригес предполагают, что Fuligo septica , возможно, вдохновила Ирвина Йиворта 1958 года на создание фильма «Капля» , в котором гигантская амеба из космоса начинает поглощать людей в маленьком американском городке. [79]
См. также
[ редактировать ]- Роевая подвижность – быстрое и скоординированное перемещение бактериальной популяции по твердым или полутвердым поверхностям.
- Водяная плесень — грибоподобный эукариотический микроорганизм.
Ссылки
[ редактировать ]- ^ Перейти обратно: а б Алексопулос, Константин Дж .; Мимс, Чарльз В.; Блэквелл, Мередит М. (1996). Вводная микология (4-е изд.). Нью-Йорк: Джон Уайли и сыновья. п. 776. ИСБН 978-0-471-52229-4 .
- ^ Панков, Томас (1654). Переносной гербарий, или Книга проворных трав и растений . Берлин.
- ^ де Бари, А. (1860). «XXV.— О мицетозоях». Анналы и журнал естественной истории . 5 (28): 233–243. дои : 10.1080/00222936008697211 . ISSN 0374-5481 .
- ^ Перейти обратно: а б с д и ж г Олив, Линдси С.; Стоянович, Кармен (техническая помощь) (1975). Мицетозои . Академическая пресса . стр. 1–7. ISBN 978-0-1252-6250-7 .
- ^ Листер, Артур; Листер, Гулиэльма (1911). Монография Mycetozoa: описательный каталог видов в Гербарии Британского музея . Лондон: Отпечатано по заказу попечителей Британского музея. дои : 10.5962/bhl.title.21191 .
- ^ Перейти обратно: а б с Шниттлер, М.; Митчелл, Д.В. (2001) [2000]. «Видовое разнообразие миксомицетов на основе концепции морфологических видов - критическое исследование». В Новотны, Вольфганг; Эшт, Эрна (ред.). жизни между животными и растениями — формы Волчья кровь и коричневый цвет . Выставка в Биологическом центре Верхней Австрии. Государственный музей. Том 73. Государственная культура Верхней Австрии. стр. 39–53. ISBN 978-3854740568 .
- ^ Мартин, GW (1932). «Систематическое положение слизевиков и его влияние на классификацию грибов». Ботанический вестник . 93 (4): 421–335. дои : 10.1086/334272 . JSTOR 2471449 . S2CID 84506715 .
- ^ Мартин, GW (1932). «Систематическое положение миксомицетов». Микология . 93 (4): 119–129. дои : 10.2307/3756254 . JSTOR 3756254 .
- ^ Коупленд, Х.Ф. (1956). Классификация низших организмов . Пало-Альто, Калифорния: Pacific Books.
- ^ Уиттакер, Р.Х. (16 мая 1969 г.). «Ответ: переназначение Gymnomycota». Наука . 164 (3881). Американская ассоциация содействия развитию науки (AAAS): 857. doi : 10.1126/science.164.3881.857.b . ISSN 0036-8075 . S2CID 239845755 .
- ^ Паттерсон, диджей; Согин, М.Л. (1992). «Происхождение эукариот и разнообразие протистанов». Происхождение и эволюция прокариотических и эукариотических клеток . Нью-Джерси: World Scientific. стр. 13–46. ISBN 978-9-8102-1262-9 .
- ^ «Знакомство с «слизевиками » . Музей палеонтологии Калифорнийского университета .
- ^ Вальверду, Жорди; и др. (2018). «Слизевик: фундаментальные механизмы биологического познания». БиоСистемы . 165 (165): 57–70. arXiv : 1712.00414 . Бибкод : 2018BiSys.165...57В . doi : 10.1016/j.biosystems.2017.12.011 . ПМИД 29326068 . S2CID 3909678 .
- ^ Балдауф, СЛ; Дулиттл, WF (октябрь 1997 г.). «Происхождение и эволюция слизевиков (Mycetozoa)» . ПНАС . 94 (22): 12007–120012. Бибкод : 1997PNAS...9412007B . дои : 10.1073/pnas.94.22.12007 . ПМК 23686 . ПМИД 9342353 .
- ^ Перейти обратно: а б Стойнева-Гертнер, Майя; Узунов, Благой; Андров, Мирослав; Иванов, Кристиан; Гертнер, Георг (21 декабря 2022 г.). «Потенциал слизевиков как нового источника для косметической промышленности» . Косметика . 10 (1). MDPI AG: 3. doi : 10.3390/cosmetics10010003 . ISSN 2079-9284 .
- ^ Адамацкий, Эндрю (2016). Достижения в области машин Physarum: обнаружение и вычисление слизевиков . Спрингер. п. 4. ISBN 978-3-319-26662-6 .
- ^ Плес, Марек (11 ноября 2023 г.). «Лабораторные снимки Марека Плеса; Микробиология - биология на другом уровне» . сайт странной науки . Проверено 2 июля 2023 г.
- ^ Перейти обратно: а б с Инг, Б. (1999). Миксомицеты Великобритании и Ирландии: справочник по идентификации . Слау, Англия: Издательство Richmond Publishing. стр. 4, 9. ISBN 978-0-85546-251-2 .
- ^ Нанненга-Бремекамп, Н. (1974). Голландские миксомицеты . Зутпен: Королевское голландское общество естествознания. ISBN 978-90-03-93130-6 .
- ^ Жулидов Даниил А.; Робартс, Ричард Д.; Жулидов Александр В.; Жулидова Ольга Владимировна; Маркелов Данила А.; Русанов Виктор А.; Хедли, Джон В. (2002). «Накопление цинка слизевиком Fuligo septica (L.) Wiggers в бывшем Советском Союзе и Северной Корее». Журнал качества окружающей среды . 31 (3): 1038–1042. Бибкод : 2002JEnvQ..31.1038Z . дои : 10.2134/jeq2002.1038 . ПМИД 12026071 .
- ^ Перейти обратно: а б Стивенсон, Стивен Л. (15 июня 2000 г.). Миксомицеты . Портленд: Timber Press. п. 65. ИСБН 978-0-88192-439-8 .
- ^ Кривомаз, Т. І.; Мишо, А.; Минтер, Д.В. (2012). «Метатрихия веспариум» (PDF) .
- ^ Перейти обратно: а б Джейкобсон, Р. (5 апреля 2012 г.). «Слизевик: нет мозгов, нет ног, нет проблем» . Час новостей PBS.
- ^ Кин, К.; Шаап, П. (март 2021 г.). «Эволюция многоклеточной сложности у социальных амеб Dictyostelid» . Гены . 12 (4): 487. doi : 10.3390/genes12040487 . ПМК 8067170 . ПМИД 33801615 .
- ^ Фиоре-Донно, Анна Мария; Николаев Сергей Иванович; Нельсон, Микаэла; Павловский, Ян; Кавальер-Смит, Томас ; Балдауф, Сандра Л. (январь 2010 г.). «Глубинная филогения и эволюция слизевиков (Mycetozoa)». Протист . 161 (1): 55–70. дои : 10.1016/j.protis.2009.05.002 . ПМИД 19656720 .
- ^ «Виды: Copromyxa arborescens M. Nesom & LS Olive» . База данных проекта Eumycetozoan . Университет Арканзаса . Проверено 26 июня 2010 г.
- ^ Перейти обратно: а б Браун, Мэтью В.; Зильберман, Джеффри Д.; Шпигель, Фредерик В. (2011). « Слизевики» среди Tubulinea (Amoebozoa): Молекулярная систематика и таксономия Copromyxa». Протист . 162 (2): 277–287. дои : 10.1016/j.protis.2010.09.003 . ISSN 1434-4610 . ПМИД 21112814 .
- ^ Браун, Мэтью В.; Зильберман, Джеффри Д.; Шпигель, Фредерик В. (2012). «Современная оценка акрасид (Acrasidae, Heterolobosea, Excavata)». Европейский журнал протистологии . 48 (2). Эльзевир Б.В.: 103–123. дои : 10.1016/j.ejop.2011.10.001 . ISSN 0932-4739 . ПМИД 22154141 .
- ^ Нойхаузер, Сигрид; Кирхмайр, Мартин; Булман, Саймон; Басс, Дэвид (2014). «Смена хозяев фитомиксидных паразитов между королевствами» . Эволюционная биология BMC . 14 (1): 33. Бибкод : 2014BMCEE..14...33N . дои : 10.1186/1471-2148-14-33 . ПМК 4016497 . ПМИД 24559266 .
- ^ Цуй, Клемент К.М.; Маршалл, Уит; Ёкояма, Ринка; Хонда, Дайске; Липпмайер, Дж. Кейси; Крэйвен, Келли Д.; Петерсон, Пол Д.; Берби, Мэри Л. (январь 2009 г.). «Филогения лабиринтуломицетов и ее значение для эволюционной потери хлоропластов и приобретения эктоплазматического скольжения». Молекулярная филогенетика и эволюция . 50 (1): 129–40. дои : 10.1016/j.ympev.2008.09.027 . ПМИД 18977305 .
- ^ Дизи, Мэри С.; Олив, Линдси С. (июль 1981 г.). «Роль аппарата Гольджи в сорогенезе клеточной слизевики Fonticula alba». Наука . 213 (4507): 561–563. Бибкод : 1981Sci...213..561D . дои : 10.1126/science.213.4507.561 . ПМИД 17794844 .
- ^ Глимн-Лейси, Дженис; Кауфман, Питер Б. (2006). «Слизевиковые формы». Иллюстрированная ботаника . Спрингер США. п. 45. дои : 10.1007/0-387-28875-9_45 . ISBN 978-0-387-28870-3 .
- ^ Перейти обратно: а б Линдли, Лора А.; Стивенсон, Стивен Л.; Шпигель, Фредерик В. (1 июля 2007 г.). «Протостелиды и миксомицеты, выделенные из водных местообитаний». Микология . 99 (4): 504–509. дои : 10.3852/микология.99.4.504 . ПМИД 18065001 .
- ^ Перейти обратно: а б Хоппе, Т.; Кучера, У. (27 августа 2022 г.). «Фенотипическая пластичность плазмодий слизевиков и молекулярная филогения наземных и водных видов» . Теория в биологических науках . 141 (3). Спрингер: 313–319. дои : 10.1007/s12064-022-00375-9 . ISSN 1431-7613 . ПМЦ 9474427 . ПМИД 36029433 .
- ^ Перейти обратно: а б Амин, Фуад; Альмансоб, Абобакр; Аль-Сабри, Ахмед (2020). «Записи о слизевиках (миксомицетах) из пустынь и других засушливых районов Саудовской Аравии». Сидовия (72). Верлаг Фердинанд Бергер и Зёне: 171–177. doi : 10.12905/0380.sydowia72-2020-0171 . ISSN 0082-0598 .
- ^ Перейти обратно: а б Глайм, Дж. М. (2019). «Слизевики: экология и среда обитания - меньшие среды обитания». Экология мохообразных . Том. 2. Бриологическое взаимодействие. Электронная книга спонсируется Мичиганским технологическим университетом и Международной ассоциацией бриологов.
- ^ Перейти обратно: а б Шпигель, Фредерик В.; Стивен Л. Стивенсон; Гарольд В. Келлер; Донна Л. Мур; Джеймс К. Кавендар (2004). «Мицетозои». У Грегори М. Мюллера; Джеральд Ф. Биллс; Мерседес С. Фостер (ред.). Биоразнообразие грибов: инвентаризация и методы мониторинга . Нью-Йорк: Elsevier Academic Press. стр. 547–576 . ISBN 0125095511 .
- ^ Цзоу, Юэ; Хоу, Цзянгань; Го, Суннин; Ли, Чангтянь; Ли, Чжуан; Стивенсон, Стивен Л.; Павлов Игорь Н.; Лю, Пу; Ли, Ю (26 октября 2022 г.). «Разнообразие клеточных слизевиков диктиостелид, включая два новых для науки вида, в лесных почвах горы Чанбайшань, Китай» . Микробиологический спектр . 10 (5). Американское общество микробиологии: e0240222. дои : 10.1128/spectrum.02402-22 . ISSN 2165-0497 . ПМЦ 9620775 . ПМИД 36190423 .
- ^ Боннер, Джон Тайлер (7 ноября 2015 г.). «Эволюция эволюции: глазами слизевика» . Бионаука . 65 (12). Издательство Оксфордского университета: 1184–1187. дои : 10.1093/biosci/biv154 . ISSN 1525-3244 .
- ^ Ли, Ян-Да; Тихелька, Эрик; Лю, Чжэнь-Хуа; Хуан, Ди-Ин; Цай, Чен-Ян (23 ноября 2021 г.). «Новые загадочные жуки-слизевики среднего мела и ранняя эволюция Sphindidae (Coleoptera: Cucujoidea)» . Систематика и филогения членистоногих . 79 : 587–597. дои : 10.3897/asp.79.e72724 . ISSN 1864-8312 .
- ^ Линг, Х. (2012). «Миксомицеты: забытые местные растения» . Общество коренных растений Нью-Джерси . Архивировано из оригинала 9 июня 2015 года . Проверено 29 мая 2018 г.
- ^ Чимилески, Скотт; Колтер, Роберто. «Жизнь на краю поля зрения» . www.hup.harvard.edu . Издательство Гарвардского университета. Архивировано из оригинала 19 октября 2023 года . Проверено 26 января 2018 г.
- ^ Алексопулос, CJ (1962). Вводная микология (второе изд.). Нью-Йорк, штат Нью-Йорк: Джон Уайли и сыновья. п. 78.
- ^ Ди, Дженнифер (1960). «Система спаривания в бесклеточной слизевике». Природа . 185 (4715): 780–781. Бибкод : 1960Natur.185..780D . дои : 10.1038/185780a0 . S2CID 4206149 .
- ^ Москвич, Катя (9 июля 2018 г.). «Слизевики помнят, но учатся ли они?» . Журнал Кванта . Проверено 2 ноября 2019 г.
- ^ Перейти обратно: а б Джадсон, Оливия (2002). Секс-совет доктора Татьяны всему творению . Нью-Йорк: Генри Холт и компания. стр. 187–193. ISBN 978-0-8050-6332-5 .
- ^ Реннер, Б. (2006). «Размножение слизевиков» . БиоВеб . Система Университета Висконсина. Архивировано из оригинала 2 ноября 2019 года . Проверено 2 ноября 2019 г.
- ^ Кавендер, Джеймс С.; Насильник, Кеннет Б. (март 1965 г.). «Acrasieae в природе. II. Лесная почва как основная среда обитания» . Американский журнал ботаники . 52 (3): 297–302. дои : 10.1002/j.1537-2197.1965.tb06789.x . ISSN 0002-9122 .
- ^ Боннер, Дж.Т. (2009). Социальные амебы: биология клеточных слизевиков . Издательство Принстонского университета. ISBN 978-0-691-13939-5 . JSTOR j.ctt7s6qz .
- ^ Эрдос, Грегори В.; Рэпер, Кеннет Б.; Воген, Линда К. (июнь 1973 г.). «Типы спаривания и образование макроцист у Dictyostelium discoideum » . Труды Национальной академии наук Соединенных Штатов Америки . 70 (6): 1828–1830. Бибкод : 1973PNAS...70.1828E . дои : 10.1073/pnas.70.6.1828 . ПМК 433606 . ПМИД 16592095 .
- ^ Перейти обратно: а б Нестле, Мэрион; Сассман, Морис (август 1972 г.). «Влияние циклического АМФ на морфогенез и накопление ферментов у Dictyostelium discoideum». Биология развития . 28 (4): 545–554. дои : 10.1016/0012-1606(72)90002-4 . ПМИД 4340352 .
- ^ Левин, Герберт; Рейнольдс, Уильям (май 1991 г.). «Потоковая нестабильность агрегирующих амеб слизевиков». Письма о физических отзывах . 66 (18): 2400–2403. Бибкод : 1991PhRvL..66.2400L . doi : 10.1103/PhysRevLett.66.2400 . ПМИД 10043475 .
- ^ Тайсон, Джон Дж.; Александр, Кевин А.; Маноранджан, В.С.; Мюррей, доктор юридических наук (1 января 1989 г.). «Спиральные волны циклического усилителя в модели агрегации слизевиков». Физика D: Нелинейные явления . 34 (1): 193–207. Бибкод : 1989PhyD...34..193T . дои : 10.1016/0167-2789(89)90234-0 . ISSN 0167-2789 .
- ^ Роос, В.; Нанджундия, В.; Мальхов, Д.; Гериш, Г. (май 1975 г.). «Усиление сигналов циклического АМФ в агрегирующих клетках Dictyostelium discoideum». Письма ФЭБС . 53 (2): 139–142. дои : 10.1016/0014-5793(75)80005-6 . ПМИД 166875 . S2CID 29448450 .
- ^ Фухимори, Тайхэй; Накадзима, Акихико; Симада, Нао; Савай, Сатоши (март 2019 г.). «Самоорганизация тканей на основе коллективной миграции клеток путем контактной активации локомоции и хемотаксиса» . Труды Национальной академии наук Соединенных Штатов Америки . 116 (10): 4291–4296. Бибкод : 2019PNAS..116.4291F . дои : 10.1073/pnas.1815063116 . ПМК 6410881 . ПМИД 30782791 .
- ^ Боннер, Джон Тайлер (1983). «Химические сигналы социальных амеб» . Научный американец . 248 (4): 114–121. Бибкод : 1983SciAm.248d.114B . doi : 10.1038/scientificamerican0483-114 . ISSN 0036-8733 . JSTOR 24968880 .
- ^ Шейх, Санеа; Фу, Чэнцзе; Браун, Мэтью; Балдауф, Сандра (1 марта 2023 г.), Глубокие истоки многоклеточности эукариот, выявленные с помощью генома Acrasis kona и транскриптомов развития , doi : 10.21203/rs.3.rs-2587723/v1
- ^ Гилберт, ХК; Мартин, GW (1933). «Миксомицеты, обнаруженные на коре живых деревьев». Исследования Университета Айовы в области естественной истории . 15 : 3–8.
- ^ Перейти обратно: а б с д Келлер, Гарольд В.; Эверхарт, Сидней (2010). «Значение миксомицетов в биологических исследованиях и преподавании». Грибы . 3 (1 (зима 2010 г.)).
- ^ Стеглич, В. (1 января 1989 г.). «Слизевики (миксомицеты) как источник новых биологически активных метаболитов» . Чистая и прикладная химия . 61 (3). Вальтер де Грюйтер ГмбХ: 281–288. дои : 10.1351/pac198961030281 . ISSN 1365-3075 . S2CID 53663356 .
- ^ Дембицкий Валерий Михайлович; Ржежанка, Томас; Спижек, Ярослав; Хануш, Люмир О. (2005). «Вторичные метаболиты слизевиков (миксомицетов)». Фитохимия . 66 (7). Эльзевир Б.В.: 747–769. Бибкод : 2005PChem..66..747D . doi : 10.1016/j.phytochem.2005.02.017 . ISSN 0031-9422 . ПМИД 15797602 .
- ^ Лиерл, Мишель Б. (2013). «Споры миксомицета (слизевика): нераспознанные аэроаллергены?». Анналы аллергии, астмы и иммунологии . 111 (6). Эльзевир Б.В.: 537–541.e2. дои : 10.1016/j.anai.2013.08.007 . ISSN 1081-1206 . ПМИД 24267365 .
- ^ Карр, Уильям Э.С. (1989). «Химические сигнальные системы в низших организмах: прелюдия к эволюции химической коммуникации в нервной системе». В Андерсоне, Питер А.В. (ред.). Эволюция первых нервных систем . Бостон, Массачусетс: Спрингер. стр. 81–94. дои : 10.1007/978-1-4899-0921-3_6 . ISBN 978-1-4899-0921-3 .
- ^ Карр, Уильям Э.С.; Глисон, Ричард А.; Трапидо-Розенталь, Генри Г. (июнь 1990 г.). «Роль перирецепторных событий в хемосенсорных процессах». Тенденции в нейронауках . 13 (6): 212–215. дои : 10.1016/0166-2236(90)90162-4 . ПМИД 1694326 . S2CID 46452914 .
- ^ Линдси, Дж.; Ласкер, Р. (1974). «Химические сигналы в море: морская аллелохимика и эволюция» . Рыболовный вестник . 72 (1): 1–11.
- ^ Ленхофф, ХМ; Хиги, W (апрель 1977 г.). «Водные беспозвоночные: модельные системы для изучения активации рецепторов и эволюции рецепторных белков». Ежегодный обзор фармакологии и токсикологии . 17 (1): 243–258. дои : 10.1146/annurev.pa.17.040177.001331 . ПМИД 17353 .
- ^ Янссенс, премьер-министр; Ван Хаастерт, П.Дж. (декабрь 1987 г.). «Молекулярные основы трансмембранной передачи сигнала у Dictyostelium discoideum » . Микробиологические обзоры . 51 (4): 396–418. дои : 10.1128/мр.51.4.396-418.1987 . ПМЦ 373123 . ПМИД 2893972 .
- ^ Сайгуса, Тецу; Теро, Ацуши; Накагаки, Тосиюки; Курамото, Йошики (январь 2008 г.). «Амебы предвидят периодические события». Письма о физических отзывах . 100 (1): 018101. Бибкод : 2008PhRvL.100a8101S . doi : 10.1103/PhysRevLett.100.018101 . hdl : 2115/33004 . ПМИД 18232821 . S2CID 14710241 .
- Бароне, Дженнифер (9 декабря 2008 г.). «#71: Слизевики демонстрируют удивительную степень интеллекта» . Обнаружить . Архивировано из оригинала 10 декабря 2008 года.
- ^ Макферсон, Китта (21 января 2010 г.). «Султан слизи»: биолог продолжает восхищаться организмами после почти 70 лет исследований» . Принстонский университет.
- ^ Чжэн, Ронг; Цзя, Хеминг; Ауалига, Лейт; Лю, Цинсинь; Ван, Шуан (2021). «Глубокий ансамбль алгоритма слизевика и алгоритма арифметической оптимизации для глобальной оптимизации» . Процессы . 9 (10): 1774. дои : 10.3390/pr9101774 .
- ^ Накагаки, Тосиюки; Ямада, Хироясу; Тот, Агота (28 сентября 2000 г.). «Решение лабиринта амебоидным организмом» . Природа . 407 (6803): 470. дои : 10.1038/35035159 . ПМИД 11028990 .
- ^ Перейти обратно: а б Теро, А.; Такаги, С.; Сайгуса, Т.; и др. (январь 2010 г.). «Правила биологического адаптивного проектирования сетей» (PDF) . Наука . 327 (5964): 439–442. Бибкод : 2010Sci...327..439T . дои : 10.1126/science.1177894 . ПМИД 20093467 . S2CID 5001773 . Архивировано из оригинала (PDF) 21 апреля 2013 г.
- Йонг, Эд (21 января 2010 г.). «Атаки слизевиков имитируют железнодорожную сеть Токио» . Научные блоги .
- ^ Кристиансен Б. (25 января 2010 г.). «Инженерия сетей слизевиков» . Техновелия .
- ^ Маркс, П. (6 января 2010 г.). «Проектирование автомагистралей методом слизевиковой плесени» . Новый учёный .
- ^ Адамацкий, Эндрю ; Акл, С.; Алонсо-Санс, Р.; и др. (2013). «Являются ли автомагистрали рациональными с точки зрения слизевика?». Международный журнал параллельных, новых и распределенных систем . 28 (3): 230–248. arXiv : 1203.2851 . дои : 10.1080/17445760.2012.685884 . S2CID 15534238 .
- ^ Парр, Д. (18 февраля 2014 г.). «Города в движении: как слизевик может изменить наши железнодорожные и автомобильные карты» . Хранитель .
- ^ «Моделирование слизевиков, использованное для картирования темной материи» . НАСА . 10 марта 2020 г.
- ^ Венц, Дж. (12 марта 2020 г.). «Слизевик помогает астрономам составить карту темной материи» . Астрономический журнал .
- ^ Перейти обратно: а б Рекехо, Оскар; Андрес-Родригес, Н. Флоро (2019). «Соображения Etnobiologicas sobre los Myxomycetes» [Этнобиологические соображения по миксомицетам]. Бол. Мадрид (на испанском языке). 43 : 25–37.
Внешние ссылки
[ редактировать ]- Слизевики великолепны (вы просто этого не знали!) | Полевой путеводитель по Орегону на YouTube
- Серия фотографий слизевиков , автор Барри Уэбб, 2023 г.