Диктиостелид
| Диктиостелид | |
|---|---|

| |
| Диктиостелиум дискоидеум | |
| Научная классификация | |
| Домен: | Эукариоты |
| Тип: | амебозоа |
| Подтип: | Коноса |
| Инфрафилум: | Эумицетозоа |
| Сорт: | Диктиостелия Листер 1909, ред. Олива 197 |
| Заказы | |
Диктиостелиды Dictyosteliomycetes ( /Dictyostelea , ICZN , Dictyostelia , ICBN ) или клеточные слизевики представляют собой группу слизевиков или социальных амеб .
Многоклеточное поведение
[ редактировать ]
Когда пища (обычно бактерии) легко доступна, диктиостелиды ведут себя как отдельные амебы, которые нормально питаются и делятся. Однако, когда запасы пищи исчерпаны, они объединяются, образуя многоклеточную совокупность, называемую псевдоплазмодием, грексом или слизнем (не путать с брюхоногим моллюском , называемым слизнем ). Грекс имеет четко выраженную переднюю и заднюю часть, реагирует на градиенты света и температуры и обладает способностью мигрировать. При правильных обстоятельствах грекс созревает, образуя сорокарпий (плодовое тело) со стеблем, поддерживающим один или несколько сорусов (шариков спор). Эти споры представляют собой неактивные клетки, защищенные устойчивыми клеточными стенками, и становятся новыми амебами, как только становится доступной пища.
У Acytostelium сорокарпий целлюлозы поддерживается стеблем, состоящим из , но у других диктиостелид стебель состоит из клеток, иногда занимающих большую часть исходных амеб. За некоторыми исключениями, эти клетки погибают при образовании стебля, и между частями грекса и частями плодового тела существует определенное соответствие. Агрегация амеб обычно происходит в сходящихся ручьях. Амебы передвигаются с помощью нитевидных ложноножек и привлекаются к химическим веществам, вырабатываемым другими амебами. У Dictyostelium discoideum агрегация сигнализируется цАМФ , но другие используют другие химические вещества. У вида Dictyostelium purpureum группировка осуществляется по родству, а не только по близости.
Использование в качестве модельного организма
[ редактировать ]Диктиостелиум использовался в качестве модельного организма в молекулярной биологии и генетике и изучается как пример клеточной коммуникации , дифференциации и запрограммированной гибели клеток . Это также интересный пример эволюции сотрудничества и мошенничества. [ 1 ] [ 2 ] [ 3 ] Большой объем исследовательских данных, касающихся D. discoideum , доступен в Интернете на сайте DictyBase .
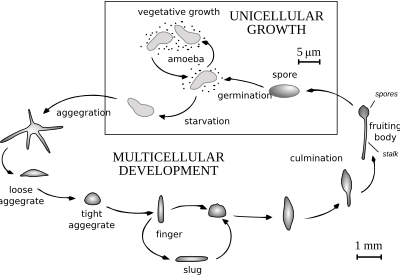
Механизм агрегации у Dictyostelium discoideum
[ редактировать ]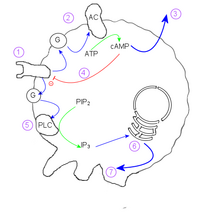
Механизм агрегации амеб основан на циклическом аденозинмонофосфате (цАМФ) в качестве сигнальной молекулы. Одна клетка, основательница колонии, в ответ на стресс начинает секретировать цАМФ. Другие обнаруживают этот сигнал и реагируют двумя способами:
- Амеба движется навстречу сигналу.
- Амеба выделяет больше цАМФ для усиления сигнала.
Эффект этого состоит в том, чтобы передать сигнал на близлежащую популяцию амеб и вызвать движение внутрь к области с наибольшей концентрацией цАМФ.
Внутри отдельной клетки механизм следующий:
- Прием цАМФ на клеточной мембране активирует G-белок.
- Белок G стимулирует аденилатциклазу
- цАМФ диффундирует из клетки в среду
- Внутренний цАМФ инактивирует внешний рецептор цАМФ.
- Другой g-белок стимулирует фосфолипазу C.
- IP 3 индуцирует высвобождение ионов кальция
- Ионы кальция действуют на цитоскелет , вызывая расширение псевдоподий .
Поскольку внутренняя концентрация цАМФ инактивирует рецептор внешнего цАМФ, отдельная клетка демонстрирует колебательное поведение. Такое поведение создает красивые спирали, наблюдаемые в сближающихся колониях, и напоминает реакцию Белоусова-Жаботинского и двумерные циклические клеточные автоматы .
Геном
[ редактировать ]Полный геном Dictyostelium discoideum был опубликован в журнале Nature в 2005 году генетиком Людвигом Эйхингером и его коллегами. [ 4 ] Гаплоидный геном содержит около 12 500 генов в 6 хромосомах. Для сравнения: диплоидный геном человека имеет 20 000–25 000 генов (представленных дважды) на 23 парах хромосом. Существует высокий уровень нуклеотидов аденозина и тимидина (~ 77%), что приводит к использованию кодонов, которое благоприятствует большему количеству аденозина и тимидина в третьей позиции. Тандемные повторы тринуклеотидов широко распространены в Dictyostelium, которые у людей вызывают нарушения тринуклеотидных повторов .
Половое размножение
[ редактировать ]Половое развитие может происходить, когда амебоидные клетки испытывают нехватку бактериальной пищи и присутствуют темные и влажные условия. [ 5 ] Спариванию могут подвергаться как гетероталличные, так и гомоталличные штаммы Dictostelium. Гетероталлическое половое развитие наиболее полно изучено у D. discoideum , а гомоталличное половое развитие наиболее хорошо изучено у D. mucoroides . [ 6 ] Гетероталлические спаривания инициируются слиянием гаплоидных клеток (гамет) двух линий противоположного типа спаривания. Это контрастирует с гомоталличными штаммами, которые, по-видимому, экспрессируют оба типа спаривания. [ 7 ]
Спаривание инициируется гаметогенезом, в результате которого образуются маленькие подвижные гаметы, которые сливаются, образуя небольшую двуядерную клетку . Затем объем двуядерной клетки увеличивается, образуя гигантскую двуядерную клетку. По мере роста ядра набухают, а затем сливаются, образуя настоящую диплоидную гигантскую клетку зиготы. Пока это происходит, у амеб происходит индуцированный цАМФ хемотаксис по направлению к поверхности гигантских клеток. Это образует клеточный агрегат, и в центре агрегата гигантская клетка зиготы поглощает окружающие амебы. За фагоцитозом следует переваривание проглоченных амеб. Затем зигота образует макроцисту, окруженную внеклеточной целлюлозной оболочкой. После образования макроцисты она обычно остается в состоянии покоя в течение некоторого времени, прежде чем может произойти прорастание. [ 8 ] Внутри макроцисты диплоидная зигота подвергается мейозу с последующими митотическими делениями. Когда макроциста прорастает, она высвобождает множество гаплоидных амебоидных клеток.
Таксономия
[ редактировать ]История классификации
[ редактировать ]Дерево филогении Dictyostelium претерпело множество изменений за последние десятилетия. Первым диктиостелидом, который был описан Оскером Брефельдом в 1869 году, был Dictyostelium mucoroides , а первоначальное открытие Dictyostelium discoideum произошло в 1935 году. [ 9 ] с дальнейшими открытиями Кеннета Рэпера , за которыми последовали глобальные усилия под руководством Джеймса Кавендера и его сотрудников. Dictyostelium discoideum первоначально был отнесен к «низшим грибам», но с тех пор классификация была переведена в категорию Amoebozoa, где она и находится в настоящее время. [ 4 ]
Группировки внутри дерева филогении диктиостелид часто менялись в связи с появлением новых данных. В большинстве общепринятых в настоящее время филогений диктиостелид используется секвенирование генома и рибосомальная ДНК малых субъединиц (ssu-рДНК). Диктиостелиды можно разделить на четыре группы. В частности, группа 4 содержит виды Dictyostelium discoideum и отличается от других групп использованием цАМФ в качестве аттрактанта, выделяющегося при агрегации. [ 10 ]
Эволюция
[ редактировать ]Калибровка окаменелостей показывает, что класс диктиостелид первоначально разделился на две основные ветви примерно 0,52 миллиарда лет назад. Современные теории предполагают, что образование стеблей и спор диктиостелид изначально возникло как адаптация к глобальным ледниковым образованиям. Дальнейшее подразделение видов диктиостелид, вероятно, возникло по мере таяния большинства ледниковых образований. Большинство видов в основных группах 1, 2 и 3 демонстрируют способность к инцистированию, которая позволяет людям выживать при низких температурах, но споры продемонстрировали повышенную способность противостоять более низким температурам. Группа 4 отличается от других основных групп отсутствием способности к инцистированию, но ее споры продемонстрировали лучшую устойчивость к более низким температурам по сравнению со спорами из других групп. [ 11 ]
Внутренняя филогения диктиостелид показана на кладограмме . [ 12 ]
| Диктиостелида | |
Таксономия
[ редактировать ]Класс Dictyostelia Lister 1909 г. эм. Олива 1970 г. [ 13 ]
- Пол? Калоспейра Арно 1949 г.
- Род? Coenonia van Tieghem 1884 нон Вандамм и др. 1999 год
- Пол? Синстелиум Балдауф, Шейх и Тулин 2017
- Орден Actyosteliales Baldauf, Sheikh & Thulin 2017
- Семейство Cavendiaceae Baldauf, Sheikh & Thulin 2017
- Род Cavenderia Baldauf, Sheikh & Thulin 2017.
- Семейство Acytosteliidae Raper ex Raper & Quinlan, 1958 г.
- Род Acitostelium Raper 1956 г.
- Род Heterostelium Baldauf, Sheikh & Thulin 2017.
- Род Rostrostellium Baldauf, Sheikh & Thulin 2017.
- Семейство Cavendiaceae Baldauf, Sheikh & Thulin 2017
- Заказать Dictyosteliales Olive ex Kirk, Cannon & David 2001
- Пол? Coremyostelium Baldauf, Шейх и Тулин, 2017 г.
- Род? Гелиомикопсис Арно 1949 г.
- Род? Пигмомицеты Арно 1949 г.
- Род †? Миксомитодес Бенгтсон и др. 2007 год
- Род? Рабдоцистис Арно 1949
- Семейство Dictyosteliidae Rostafinski 1875 ex Cooke 1877
- Диктиостелиум Брефельд 1870 г.
- Полисфондилий Брефельд 1884 г.
- Семейство Raperosteliaceae Baldauf, Sheikh & Thulin 2017
- Род Hagiwaraea Baldauf, Sheikh & Thulin 2017.
- Род Raperostelium Baldauf, Sheikh & Thulin 2017.
- Род Speleostelium Baldauf, Sheikh & Thulin 2017.
- Род Tiehemostelium Baldauf, Sheikh & Thulin 2017.
Модельный организм-хозяин легионеллы
[ редактировать ]Dictyostelium имеет много общих молекулярных особенностей с макрофагами , человеческими хозяевами Legionella . Цитоскелетный состав D. discoideum подобен составу клеток млекопитающих, как и процессы, запускаемые этими компонентами, такие как фагоцитоз, мембранный транспорт, эндоцитарный транзит и сортировка везикул. Как и лейкоциты, D. discoideum обладают хемотаксической способностью. Таким образом, D. discoideum представляет собой подходящую модельную систему для установления влияния различных факторов клеток-хозяев во время инфекций Legionella . [ 14 ]
Ссылки
[ редактировать ]- ^ Страссман Дж. Э., Чжу Ю. и Квеллер, округ Колумбия. (2000)Альтруизм и социальное мошенничество у социальной амебы Dictyostelium discoideum . Природа
- ^ Дао Д.Н., Кессин Р.Х. и Эннис Х.Л. (2000). Обман развития и эволюционная биология Dictyostelium и Myxococcus . Микробиология
- ^ Брэннсрём О и Дикманн У (2005). Эволюционная динамика альтруизма и мошенничества среди социальных амеб. Труды Лондонского королевского общества Б.
- ^ Перейти обратно: а б Эйхингер, Л.; Пачебат, Дж.А.; Глекнер, Г.; Раджандрим, Массачусетс; Сукганг, Р.; Берриман, М.; Сонг, Дж.; Олсен, Р.; Шафрански, К.; Сюй, Кью; и др. (2005). «Геном социальной амебы Dictyostelium discoideum» . Природа . 435 (7038): 43–57. Бибкод : 2005Natur.435...43E . дои : 10.1038/nature03481 . ПМЦ 1352341 . ПМИД 15875012 .
- ^ Флауэрс Дж.М., Ли С.И., Статос А., Саксер Г., Островски Э.А., Квеллер Д.С., Страссманн Дж.Э., Пуруганан, доктор медицинских наук (июль 2010 г.). «Вариации, пол и социальное сотрудничество: молекулярная популяционная генетика социальной амебы Dictyostelium discoideum» . ПЛОС Генет . 6 (7): e1001013. дои : 10.1371/journal.pgen.1001013 . ПМЦ 2895654 . ПМИД 20617172 .
- ^ О'Дей Д.Х., Кесей А. (май 2012 г.). «Сигнализация и пол у социальных амебозойных». Biol Rev Camb Philos Soc . 87 (2): 313–29. дои : 10.1111/j.1469-185X.2011.00200.x . ПМИД 21929567 . S2CID 205599638 .
- ^ Робсон Дж.Э., Уильямс К.Л. (апрель 1980 г.). «Система спаривания клеточной слизевика Dictyostelium discoideum». Курс. Жене . 1 (3): 229–32. дои : 10.1007/BF00390948 . ПМИД 24189663 . S2CID 23172357 .
- ^ Никерсон А.В., Рэпер КБ. Макроцисты в жизненном цикле Dictyostelliaceae II. Прорастание макроцист. Являюсь. Дж. Бот. 1973 60 (3): 247–254.
- ^ Брефельд, О (1869). «Новый организм и взаимоотношения миксомицетов». Абх Зекенберг Натурфорш Гес . 7 :85–107.
- ^ Шильде, Кристина; Лаваль, Хагар М.; Кин, Корю; Сибано-Хаякава, Икуми; Иноуе, Кей; Шаап, Полина (2019). «Хорошо подтвержденная мультигенная филогения 52 диктиостелий» . Молекулярная филогенетика и эволюция . 134 : 66–73. дои : 10.1016/j.ympev.2019.01.017 . ISSN 1055-7903 . ПМК 6430600 . ПМИД 30711536 .
- ^ Лаваль, Хаджара М.; Шильде, Кристина; Кин, Корю; Браун, Мэтью В.; Джеймс, Джон; Прескотт, Алан Р.; Шаап, Полина (29 мая 2020 г.). «Адаптация к холодному климату является вероятной причиной эволюции многоклеточного спорообразования у Dictyostelia» . Научные отчеты . 10 (1): 8797. doi : 10.1038/s41598-020-65709-3 . ISSN 2045-2322 . ПМК 7260361 . ПМИД 32472019 .
- ^ Шейх, Санеа; Тулин, Матс; Кавендер, Джеймс С; Эскаланте, Рикардо; Каваками, Синъити; Ладо, Карлос; Ландольт, Джон; Нанджундия, Видьянанд; Квеллер, Дэвид; Штрассманн, Джоан; Шпигель, Фредерик В.; Стивенсон, Стив; Ваделл, Эдуардо М; Балдауф, Сандра (24 ноября 2017 г.). «Новая классификация диктиостелид» . Протисты . 169 (1): 1–28. дои : 10.1016/j.protis.2017.11.001 . ПМИД 29367151 .
- ^ Виджаявардене, Налин; Хайд, Кевин; Аль-Ани, ЛКТ; Долатабади, С; Стадлер, Марк; Хээлуотерс, Дэнни; и др. (2020). «Очерк грибов и грибоподобных таксонов» . Микосфера . 11 : 1060–1456. дои : 10.5943/микосфера/11/1/8 . hdl : 10481/61998 .
- ^ Брюн; и др. (2008). « Диктиостелиум , управляемая модель организма-хозяина легионеллы » . Легионелла : Молекулярная микробиология . Кайстер Академик Пресс. ISBN 978-1-904455-26-4 .
Внешние ссылки
[ редактировать ]- Диктиостелиум (2007)
- Низшее общество (2004)
- dictyBase Интернет-ресурс по информатике для Dictyostelium
- dictyBase wiki официальный вики-сайт dictyBase
- Dictyostelium discoideum Проект генома . Архивировано 6 декабря 2019 г. в Wayback Machine.
- Описание Dictyostelium discoideum , жизненный цикл