Диктиостелиум дискоидеум
| Диктиостелиум дискоидеум | |
|---|---|
 | |
| Плодовые тела D. discoideum | |
| Продолжительность: 23 секунды. | |
| Мигрирующий D. discoideum , граница которого окрашена кривизной, масштаб: 5 мкм, продолжительность: 22 секунды. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Тип: | амебозоа |
| Сорт: | Диктиостелия |
| Заказ: | Диктиостелида |
| Семья: | Диктиостелииды |
| Род: | Диктиостелиум |
| Разновидность: | Д. дискоидеум |
| Биномиальное имя | |
| Диктиостелиум дискоидеум | |

Dictyostelium discoideum — вид почвенных амеб, принадлежащий к типу Amoebozoa , infraphylum Mycetozoa . , обычно называемый слизевиком , D. discoideum представляет собой эукариот , который в течение своей жизни превращается из скопления одноклеточных амеб в многоклеточную слизню, а затем в плодовое тело. Его уникальный бесполый жизненный цикл состоит из четырех стадий: вегетативной, агрегационной, миграции и кульминации. Жизненный цикл D. discoideum относительно короткий, что позволяет своевременно просматривать все стадии. Клетки, участвующие в жизненном цикле, подвергаются движению, химической передаче сигналов и развитию, что применимо к исследованиям рака у человека. Простота жизненного цикла делает D. discoideum ценным модельным организмом для изучения генетических, клеточных и биохимических процессов в других организмах. [2]
Естественная среда обитания и рацион
[ редактировать ]В дикой природе D. discoideum можно найти в почве и влажной листовой подстилке. Его основной рацион состоит из бактерий , таких как Escherichia coli , обитающих в почве и разлагающихся органических веществах. Одноядерные амебы D. discoideum потребляют бактерии, встречающиеся в их естественной среде обитания, включая почву лиственных лесов и гниющие листья. [3]
Жизненный цикл и размножение
[ редактировать ]Жизненный цикл начинается D. discoideum , когда споры высвобождаются из зрелого сорокарпа (плодового тела). Миксамебы вылупляются из спор в теплых и влажных условиях. На вегетативной стадии миксамебы делятся митозом , питаясь бактериями. Бактерии выделяют фолиевую кислоту , которая привлекает миксамеб. Когда запас бактерий истощается, миксамебы вступают в стадию агрегации.
Во время агрегации голодание инициирует выработку белковых соединений, таких как гликопротеины и аденилатциклаза . [4] Гликопротеины обеспечивают межклеточную адгезию , а аденилатциклаза создает циклический АМФ . Циклический АМФ секретируется амебами для привлечения соседних клеток к центральному месту. Двигаясь к сигналу, они сталкиваются друг с другом и слипаются за счет гликопротеиновых молекул адгезии.
Стадия миграции начинается, когда амебы образовали плотный агрегат и удлиненный холмик клеток опрокидывается и плашмя лежит на земле. Амебы действуют вместе как подвижный псевдоплазмодий, также известный как слизень. Слизень имеет длину около 2–4 мм и состоит из до 100 000 клеток. [5] и способен двигаться, образуя клетках целлюлозную в своих передних оболочку , через которую движется слизняк. [6] Часть этой оболочки остается в виде скользкого следа, когда она движется к аттрактантам, таким как свет , тепло и влажность , только в прямом направлении. [6] Циклический АМФ и вещество, называемое фактором, индуцирующим дифференцировку , помогают формировать различные типы клеток. [6] Слизень дифференцируется на предстебельковые и преспоровые клетки, которые перемещаются к переднему и заднему концам соответственно. Как только слизень нашел подходящую среду, передний конец слизняка образует стебель плодового тела, а задний конец образует споры плодового тела. [6] Переднеподобные клетки, которые были обнаружены лишь недавно, также рассеяны по всей задней части слизняка. Эти переднеподобные клетки образуют самое дно плодового тела и шляпки спор. [6] После того, как слизняк оседает в одном месте, задний конец расправляется, а передний конец поднимается в воздух, образуя так называемую «мексиканскую шляпу», и начинается кульминационная стадия.
На кульминационной стадии предстебельковые клетки и преспоровые клетки меняют положение, образуя зрелое плодовое тело. [6] Передний конец мексиканской шляпы образует целлюлозную трубку, которая позволяет более задним клеткам перемещаться вверх по внешней стороне трубки к верху, а предстебельчатым клеткам двигаться вниз. [6] В результате этой перестройки формируется ножка плодового тела, состоящая из клеток переднего конца слизняка, а клетки заднего конца слизняка оказываются наверху и теперь образуют споры плодового тела. В конце этого 8–10-часового процесса полностью формируется зрелое плодовое тело. [6] Это плодовое тело имеет высоту 1–2 мм и теперь может начать весь цикл заново, выпустив зрелые споры, которые становятся миксамебами.
Половое размножение
[ редактировать ]Хотя D. discoideum обычно размножается бесполым путем , D. discoideum все же способен к половому размножению при соблюдении определенных условий. У D. discoideum есть три различных типа спаривания, и исследования выявили половой локус, который определяет эти три типа спаривания. Штаммы типа I определяются геном, называемым MatA, штаммы типа II имеют три разных гена: MatB (гомологичный Mat A), Mat C и Mat D, а штаммы типа III имеют гены Mat S и Mat T (которые гомологичны Мат C и Мат D). [7] Представители этого пола могут спариваться только с представителями двух разных полов, а не со своим собственным. [7]
При инкубации с бактериальной пищей может происходить гетероталличное или гомоталличное половое развитие, приводящее к образованию диплоидной зиготы. [8] [9] Гетероталлическое спаривание происходит, когда две амебы разных типов спаривания присутствуют в темной и влажной среде, где они могут сливаться во время агрегации, образуя гигантскую клетку-зиготу. Затем гигантская клетка высвобождает цАМФ для привлечения других клеток, а затем каннибалистически поглощает другие клетки в совокупности. Съеденные клетки служат для того, чтобы покрыть весь агрегат толстой целлюлозной стенкой и защитить его. Это известно как макрокиста . Внутри макроцисты гигантская клетка делится сначала посредством мейоза , а затем посредством митоза, образуя множество гаплоидных амеб, которые будут высвобождаться для питания, как это делают нормальные амебы. Гомоталлические штаммы D. discoideum AC4 и ZA3A также способны продуцировать макроцисты. [10] Каждый из этих штаммов, в отличие от гетероталличных штаммов, вероятно, экспрессирует аллели обоих типов спаривания ( matA и mata ). в лабораторных условиях встречается очень редко Хотя половое размножение возможно, успешное прорастание макроцист D. discoideum . Тем не менее, рекомбинация широко распространена в природных популяциях D. discoideum , что указывает на то, что пол, вероятно, является важным аспектом их жизненного цикла. [9]
Использование в качестве модельного организма
[ редактировать ]Поскольку многие из его генов гомологичны генам человека, но его жизненный цикл прост, D. discoideum обычно используется в качестве модельного организма . Его можно наблюдать на организменном, клеточном и молекулярном уровнях, прежде всего из-за ограниченного числа типов клеток и поведения, а также их быстрого роста. [6] Он используется для изучения дифференцировки клеток, хемотаксиса и апоптоза , которые являются нормальными клеточными процессами. Он также используется для изучения других аспектов развития, включая сортировку клеток, формирование паттернов, фагоцитоз, подвижность и передачу сигналов. [11] Эти процессы и аспекты развития либо отсутствуют, либо их слишком сложно увидеть у других модельных организмов. D. discoideum тесно связан с высшими многоклеточными животными. Он несет схожие гены и пути, что делает его хорошим кандидатом на нокаут генов . [12]
Процесс дифференциации клеток происходит, когда клетка становится более специализированной для развития в многоклеточный организм. Изменения размера, формы, метаболической активности и реакции могут происходить в результате корректировок экспрессии генов. Разнообразие и дифференциация клеток у этого вида включает в себя решения, принимаемые на основе межклеточных взаимодействий на путях либо к клеткам стебля, либо к клеткам спор. [13] Судьба этих клеток зависит от их окружения и формирования структуры. Таким образом, организм является прекрасной моделью для изучения клеточной дифференцировки.
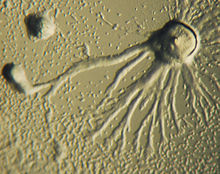
Хемотаксис определяется как переход организма к химическому раздражителю или от него по градиенту химической концентрации. Некоторые организмы демонстрируют хемотаксис, когда они движутся к запасу питательных веществ. У D. discoideum амеба секретирует сигнал цАМФ из клетки, привлекая другие амебы к миграции к источнику. Каждая амеба движется к центральной амебе, той, которая выделяет наибольшее количество секреции цАМФ. Секреция цАМФ затем проявляется у всех амеб и является для них сигналом к началу агрегации. Эти химические выбросы и движение амеб происходят каждые шесть минут. Амебы движутся в направлении градиента концентрации в течение 60 секунд и останавливаются до тех пор, пока не будет выпущен следующий секрет. Такое поведение отдельных клеток имеет тенденцию вызывать колебания в группе клеток, и химические волны с различной концентрацией цАМФ распространяются через группу по спирали. [14] : 174–175
Элегантный набор математических уравнений, воспроизводящий спирали и потоки D. discoideum, был обнаружен биологами-математиками Томасом Хёфером и Мартином Бёрлийстом. Биолог-математик Корнелис Дж. Вейер доказал, что подобные уравнения могут моделировать его движение. На уравнения этих закономерностей в основном влияют плотность популяции амеб, скорость продукции циклического АМФ и чувствительность отдельных амеб к циклическому АМФ. Спиральный узор формируется амебами в центре колонии, которые вращаются, посылая волны циклического АМФ. [15] [16]
Использование цАМФ в качестве хемотаксического агента не установлено ни в одном другом организме. В биологии развития это один из понятных примеров хемотаксиса, который важен для понимания воспаления у человека, артрита, астмы, транспорта лимфоцитов и управления аксонами. Фагоцитоз используется для иммунного надзора и презентации антигенов, тогда как определение типа клеток, сортировка клеток и формирование структуры являются основными особенностями эмбриогенеза, которые можно изучать с помощью этих организмов. [6]
Однако обратите внимание, что колебания цАМФ могут не быть необходимыми для коллективной миграции клеток на многоклеточных стадиях. Исследование показало, что на многоклеточной стадии D. discoideum передача сигналов, опосредованная цАМФ, меняется от распространяющихся волн к устойчивому состоянию. [17]
Термотаксис – движение по градиенту температуры. Было показано, что слизни мигрируют по чрезвычайно пологим градиентам, всего 0,05 ° C/см, но выбранное направление сложно; кажется, что температура здесь примерно на 2 ° C ниже той, к которой они были акклиматизированы. Это сложное поведение было проанализировано с помощью компьютерного моделирования поведения и периодического изменения температуры в почве, вызванного ежедневными изменениями температуры воздуха. Вывод состоит в том, что такое поведение перемещает слизней на несколько сантиметров ниже поверхности почвы вверх на поверхность. Это удивительно сложное поведение примитивного организма, лишенного явного чувства гравитации. [14] : 108–109
Апоптоз (запрограммированная гибель клеток) – нормальная часть развития вида. [4] Апоптоз необходим для правильного расположения и формирования сложных органов. Около 20% клеток D. discoideum альтруистически жертвуют собой при формировании зрелого плодового тела. На стадии псевдоплазмодия (слизняка или грекса) своего жизненного цикла в организме образовались три основных типа клеток: предстебельковые, преспоровые и переднеподобные клетки. Во время кульминации предстебельковые клетки выделяют целлюлозную оболочку и проходят через грекс в виде трубки. [4] По мере дифференцировки они образуют вакуоли и увеличиваются в размерах, поднимая вверх клетки преспор. Клетки стебля подвергаются апоптозу и погибают, когда клетки преспор поднимаются высоко над субстратом. Преспоровые клетки затем превращаются в споровые клетки, каждая из которых при расселении становится новой миксамебой. [6] Это пример того, как апоптоз используется при формировании репродуктивного органа – зрелого плодового тела.
Недавний значительный вклад в исследования Dictyostelium внесли новые методы, позволяющие визуализировать активность отдельных генов в живых клетках. [18] Это показало, что транскрипция происходит в виде «всплесков» или «импульсов» ( всплеск транскрипции ), а не в результате простого вероятностного или непрерывного поведения. В настоящее время взрывная транскрипция, по-видимому, сохраняется между бактериями и людьми. Еще одной замечательной особенностью этого организма является наличие в нем набора ферментов репарации ДНК, обнаруженных в клетках человека, которых нет во многих других популярных модельных системах многоклеточных животных. [19] Дефекты в репарации ДНК приводят к разрушительным раковым заболеваниям у человека, поэтому возможность изучать репарационные белки человека на простой и понятной модели окажется неоценимой.
Лабораторное выращивание
[ редактировать ]Способность этого организма легко изолироваться и культивироваться в лаборатории. [6] [20] добавляет ему привлекательности в качестве модельного организма. Хотя D. discoideum можно выращивать в жидкой культуре, обычно его выращивают в чашках Петри, содержащих питательный агар , а поверхности поддерживают влажными. Культуры лучше всего растут при температуре 22–24 °C (комнатная температура). D. discoideum питаются в основном кишечной палочкой , которой хватает на всех стадиях жизненного цикла. Когда запасы пищи уменьшаются, миксамебы объединяются, образуя псевдоплазмодии. Вскоре блюдо проходит различные стадии жизненного цикла. Проверка блюда часто позволяет детально наблюдать за развитием. Клетки можно собирать на любой стадии развития и быстро выращивать.
При культивировании D. discoideum в лаборатории важно учитывать его поведенческие реакции. Например, он имеет сродство к свету, более высоким температурам, высокой влажности, низкой концентрации ионов и кислой стороне градиента pH. Часто проводятся эксперименты, чтобы увидеть, как манипуляции с этими параметрами препятствуют, останавливают или ускоряют развитие. Вариации этих параметров могут изменить скорость и жизнеспособность роста культуры. Кроме того, плодовые тела, поскольку это самая высокая стадия развития, очень чувствительны к потокам воздуха и физическим раздражителям. Неизвестно, существует ли стимул, связанный с выбросом спор.
Исследования экспрессии белка
[ редактировать ]Детальный анализ экспрессии белка у Dictyostelium затруднен из-за значительных сдвигов в профиле экспрессии белка между различными стадиями развития и общей нехваткой коммерчески доступных антител к антигенам Dictyostelium . [21] В 2013 году группа онкологического центра Beatson West of Scotland сообщила о стандарте визуализации белков без антител для иммуноблоттинга, основанном на обнаружении MCCC1 с использованием конъюгатов стрептавидина . [22]
болезнь легионеров
[ редактировать ]Род бактерий Legionella включает виды, вызывающие у человека болезнь легионеров . D. discoideum также является хозяином легионеллы и является подходящей моделью для изучения инфекционного процесса. [23] В частности, D. discoideum имеет сходный с клетками-хозяевами млекопитающих цитоскелет и клеточные процессы, имеющие отношение к инфекции Legionella , включая фагоцитоз , мембранный транспорт, эндоцитоз , сортировку везикул и хемотаксис.
«Фермерство»
[ редактировать ]В отчете журнала Nature за 2011 год были опубликованы результаты, демонстрирующие «примитивное земледельческое поведение» колоний D. discoideum . [24] [25] Описываемый как « симбиоз » между D. discoideum и бактериальной добычей, около трети собранных в дикой природе колоний D. discoideum занимаются « разведением » бактерий, когда бактерии попадают в плодовые тела слизевиков. [25] Включение бактерий в плодовые тела позволяет «засеять» источник пищи в месте распространения спор, что особенно ценно, если в новом месте мало пищевых ресурсов. [25] Колонии, полученные из «фермерских» спор, обычно демонстрируют такое же поведение при спорообразовании. Это объединение связано с издержками: те колонии, которые не потребляют все бактерии-жертвы, производят более мелкие споры, которые не могут распространяться так широко. Кроме того, гораздо меньше пользы приносят споры, содержащие бактерии, которые попадают в регион, богатый пищевыми продуктами. Такой баланс издержек и выгод от такого поведения может способствовать тому, что меньшинство колоний D. discoideum практикует эту практику. [24] [25]
D. discoideum известен тем, что поедает как грамположительные , так и грамотрицательные бактерии , но некоторые из фагоцитируемых бактерий, в том числе некоторые человеческие патогены, [26] способны жить в амебах и выходить из них, не убивая клетку. Когда они попадают в камеру, где они проживают и когда покидают камеру, неизвестно. Исследование еще не является окончательным, но можно нарисовать общий жизненный цикл D. discoideum, адаптированный для фермерских клонов, чтобы лучше понять этот симбиотический процесс.

На картинке можно увидеть разные этапы. Во-первых, на стадии голодания бактерии заключены внутри D. discoideum , [26] после проникновения в амебы в фагосоме слияние с лизосомами блокируется, и эти незрелые фагосомы окружены органеллами клетки-хозяина, такими как митохондрии, везикулы и многослойная мембрана, происходящая из шероховатой эндоплазматической сети (RER) амеб. Роль РЭР во внутриклеточной инфекции неизвестна, но РЭР не требуется в качестве источника белков для бактерий. [27] Бактерии обитают внутри этих фагосом на стадиях агрегации и многоклеточного развития. Амебы сохраняют свою индивидуальность, и у каждой амебы есть своя бактерия. На кульминационной стадии, когда образуются споры, бактерии переходят из клетки в сорус с помощью структуры цитоскелета, которая предотвращает разрушение клетки-хозяина. [28] Некоторые результаты показывают, что бактерии используют экзоцитоз, не убивая клетку. [28] Свободноживущие амебы, по-видимому, играют решающую роль в сохранении и распространении некоторых патогенов в окружающей среде. Сообщалось о временной ассоциации с амебами для ряда различных бактерий, включая Legionella pneumophila , многие Mycobacterium виды , Francesella tularensis и Escherichia coli , среди других. [27] Сельское хозяйство, по-видимому, играет решающую роль в выживании патогенов, поскольку они могут жить и размножаться внутри D. discoideum , занимаясь животноводством. Отчет природы сделал важный шаг вперед в изучении поведения амеб, а знаменитая испанская фраза, переведенная как «ты глупее амебы», теряет смысл, поскольку амебы являются прекрасным примером социального поведения с удивительной координацией и чувство жертвенности ради блага вида. [ нужна ссылка ]
Дозорные клетки
[ редактировать ]Сигнальные клетки Dictyostelium discoideum представляют собой фагоцитирующие клетки, ответственные за удаление токсического материала со стадии слизняка в социальном цикле. Обычно эти клетки имеют круглую форму и находятся внутри оболочки слизней, где они свободно циркулируют. Процесс детоксикации происходит, когда эти клетки поглощают токсины и патогены внутри слизняка посредством фагоцитоза . Затем клетки группируются в группы по пять-десять клеток, которые затем прикрепляются к внутренней оболочке слизняка. Оболочка отслаивается, когда слизень мигрирует на новое место в поисках пищевых бактерий.
Сигнальные клетки составляют примерно 1% от общего числа клеток-слизней, и количество сигнальных клеток остается постоянным даже во время их высвобождения. Это указывает на постоянную регенерацию сторожевых клеток внутри слизней по мере их удаления вместе с токсинами и патогенами. Сигнальные клетки присутствуют в слизне, даже если нет токсинов или патогенов, которые необходимо удалить. Сигнальные клетки были обнаружены у пяти других видов Dictyostelia , что позволяет предположить, что сигнальные клетки можно описать как общую характеристику врожденной иммунной системы социальных амеб. [29]
Влияние статуса фермерства на сторожевые клетки
[ редактировать ]Число сторожевых клеток варьируется в зависимости от статуса выращивания дикого D. discoideum . При воздействии токсичной среды, созданной при использовании бромистого этидия, было показано, что количество сторожевых клеток на миллиметр было ниже у фермеров, чем у нефермеров. К такому выводу пришли наблюдения за следами, оставленными после миграции слизней, и подсчет количества присутствующих сторожевых клеток в миллиметре. Однако количество сторожевых клеток не влияет на продукцию и жизнеспособность спор у фермеров. Фермеры, подвергшиеся воздействию токсичной среды, производят такое же количество спор, как и фермеры в нетоксичной среде, а жизнеспособность спор была одинаковой у фермеров и нефермеров. Когда у фермеров удаляли Clade 2 Burkholderia , или бактерии, связанные с фермерами, производство и жизнеспособность спор были аналогичны таковым у нефермеров. Таким образом, предполагается, что бактерии, переносимые фермерами, обеспечивают дополнительную защиту фермеров от потенциального вреда, вызванного токсинами или патогенами. [30]
Классификация и филогения
[ редактировать ]В старых классификациях Dictyostelium относили к несуществующему полифилетическому классу Acrasiomycetes . Это был класс клеточных слизевиков, для которого характерно агрегирование отдельных амеб в многоклеточное плодовое тело, что делает его важным фактором, связывающим акразиды с диктиостелидами. [31]
Более поздние геномные исследования показали, что Dictyostelium сохранил большую часть своего предкового разнообразия генома, чем растения и животные, хотя филогения на основе протеома подтверждает, что амебозоа отделились от линии животных и грибов после разделения растений и животных. [32] Подкласс Dictyosteliidae, отряд Dictyosteliales представляет собой монофилетическое собрание внутри Mycetozoa, группы, в которую входят протостелиды, диктиостелиды и миксогастриды.Анализ данных фактора элонгации-1α (EF-1α) подтверждает, что Mycetozoa является монофилетической группой, хотя деревья рРНК относят ее к полифилетической группе. Кроме того, эти данные подтверждают идею о том, что диктиостелиды и миксогастриды более тесно связаны друг с другом, чем протостелиды. Анализ EF-1α также определил Mycetozoa как непосредственную внешнюю группу животно-грибной клады. [33] Последние филогенетические данные относят диктиостелиды к супергруппе Amoebozoa , наряду с миксомицетами . Между тем, протостелиды оказались полифилетическими, их плодовые тела со стебельками являются конвергентным признаком множества неродственных линий. [34]
Геном
[ редактировать ]Проект D. discoideum секвенирования генома был завершен и опубликован в 2005 году международным сотрудничеством институтов. Это был первый полностью секвенированный геном свободноживущих простейших. D. discoideum состоит из гаплоидного генома размером 34 МБ с базовым составом 77% [A+T] и содержит шесть хромосом, которые кодируют около 12 500 белков. [3] Секвенирование генома D. discoideum обеспечивает более сложное изучение его клеточной биологии и биологии развития.
В этом геноме очень много тандемных повторов тринуклеотидов; один класс генома сгруппирован, что заставляет исследователей полагать, что он служит центромерами. Повторы соответствуют повторяющимся последовательностям аминокислот и, как полагают, расширяются за счет расширения нуклеотидов. [3] Расширение тринуклеотидных повторов происходит и у людей, что в целом приводит ко многим заболеваниям. Изучение того, как клетки D. discoideum переносят эти аминокислотные повторы, может дать представление о том, как люди смогут их переносить.
Каждый секвенированный геном играет важную роль в идентификации генов, которые были приобретены или потеряны с течением времени. Сравнительные геномные исследования позволяют сравнивать геномы эукариот. Филогения, основанная на протеоме, показала, что амебозоа отклонились от животно-грибной линии после разделения растений и животных. [3] Геном D. discoideum примечателен тем, что многие кодируемые им белки обычно встречаются у грибов, растений и животных. [3]
Базы данных
[ редактировать ]- DictyBase - общая геномная база данных Dictyostelium discoideum.
- База данных мембранома предоставляет информацию об однопроходных трансмембранных белках Dictyostelium . и некоторых других организмов
Ссылки
[ редактировать ]- ^ Рапер, КБ (1935). « Dictyostelium discoideum , новый вид слизевика, образующегося на разлагающихся лесных листьях» . Журнал сельскохозяйственных исследований . 50 : 135–147. Архивировано из оригинала 8 декабря 2017 г. Проверено 20 января 2016 г.
- ^ Груши, Кэтрин Дж.; Гросс, Джулиан Д. (01 марта 2021 г.). «Профиль микроба: Dictyostelium discoideum: модельная система для развития, хемотаксиса и биомедицинских исследований» . Микробиология . 167 (3). дои : 10.1099/mic.0.001040 . ISSN 1350-0872 . ПМИД 33646931 . S2CID 232092012 .
- ^ Перейти обратно: а б с д и Эйхингер Л; Ногель, А.А. (2003). «Вползая в новую эру – проект генома Dictyostelium » . Журнал ЭМБО . 22 (9): 1941–1946. дои : 10.1093/emboj/cdg214 . ПМК 156086 . ПМИД 12727861 .
- ^ Перейти обратно: а б с Гилберт С.Ф. 2006. Биология развития. 8-е изд. Сандерленд (Массачусетс): Зинауер с. 36-39.
- ^ Купер, Джеффри М. (2000). «Глава 1. Обзор клеток и клеточных исследований» . Клетка (Работа на книжной полке NCBI ). Часть I. Введение (2-е изд.). Сандерленд, Массачусетс: Sinauer Associates . Клетки как экспериментальные модели . ISBN 978-0-87893-106-4 .
- ^ Перейти обратно: а б с д и ж г час я дж к л Тайлер М.С. 2000. Биология развития: Руководство для экспериментальных исследований. 2-е изд. Сандерленд (Массачусетс): Синауэр. п. 31-34. ISBN 0-87893-843-5
- ^ Перейти обратно: а б Блумфилд, Гарет; Скелтон, Джейсон; Ивенс, Аласдер; Танака, Ёсимаса; Кей, Роберт Р. (10 декабря 2010 г.). «Определение пола у социальной амебы Dictyostelium discoideum» . Наука . 330 (6010): 1533–1536. Бибкод : 2010Sci...330.1533B . дои : 10.1126/science.1197423 . ISSN 0036-8075 . ПМЦ 3648785 . ПМИД 21148389 .
- ^ О'Дей Д.Х., Кесей А. (май 2012 г.). «Сигнализация и пол у социальных амебозойных». Biol Rev Camb Philos Soc . 87 (2): 313–29. дои : 10.1111/j.1469-185X.2011.00200.x . ПМИД 21929567 . S2CID 205599638 .
- ^ Перейти обратно: а б Флауэрс Дж.М., Ли С.И., Статос А., Саксер Г., Островски Э.А., Квеллер Д.С., Страссманн Дж.Э., Пуруганан, доктор медицинских наук (июль 2010 г.). «Вариации, пол и социальное сотрудничество: молекулярная популяционная генетика социальной амебы Dictyostelium discoideum» . ПЛОС Генет . 6 (7): e1001013. дои : 10.1371/journal.pgen.1001013 . ПМЦ 2895654 . ПМИД 20617172 .
- ^ Робсон Дж.Э., Уильямс К.Л. (апрель 1980 г.). «Система спаривания клеточной слизевика Dictyostelium discoideum». Курс. Жене . 1 (3): 229–32. дои : 10.1007/BF00390948 . ПМИД 24189663 . S2CID 23172357 .
- ^ Диктибаза, О диктиостелиуме. [Онлайн] (1 мая 2009 г.). http://dictybase.org/
- ^ Дилип К. Наг, Нарушение четырех генов кинезина в Dictyostelium. [Онлайн] (22 апреля 2008 г.). http://ukpmc.ac.uk/articlerender.cgi?artid=1529371 Архивировано 29 июля 2012 г. на archive.today.
- ^ Кей РР; Гаррод Д.; Тилли Р. (1978). «Требования к дифференцировке клеток Dictyostelium discoideum ». Природа . 211 (5640): 58–60. Бибкод : 1978Natur.271...58K . дои : 10.1038/271058a0 . ПМИД 203854 . S2CID 4160546 .
- ^ Перейти обратно: а б Дюсенбери, Дэвид Б. (1996). Жизнь в малых масштабах . Научная американская библиотека. Нью-Йорк. ISBN 978-0-7167-5060-4 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Ян Стюарт (ноябрь 2000 г.). «Образцы биоматематики: спиральная слизь. МАТЕМАТИЧЕСКИЕ ОТДЫХИ Яна Стюарта. Обнаружение математики в существах больших и малых» . Научный американец .
- ^ Ян Стюарт (2000). Какой формы снежинка? [ О снежных кристаллах и полосках зебры. Мир согласно математике ] (на голландском языке). Издательство Юниперс; Фонд Дэвида; Природа и технологии. стр. 96–97.
- ^ Уэда, Масахиро; Масато Ясуи; Моримото, Юсуке В.; Хашимура, Хиденори (24 января 2019 г.). «Коллективная миграция клеток Dictyostelium без колебаний цАМФ на многоклеточных стадиях» . Коммуникационная биология . 2 (1): 34. дои : 10.1038/s42003-018-0273-6 . ISSN 2399-3642 . ПМК 6345914 . ПМИД 30701199 .
- ^ Чабб, младший; Трчек, Т; Шеной, С.М.; Певица, Р.Х. (2006). «Транскрипционная пульсация гена развития» . Современная биология . 16 (10): 1018–25. Бибкод : 2006CBio...16.1018C . дои : 10.1016/j.cub.2006.03.092 . ПМЦ 4764056 . ПМИД 16713960 .
- ^ Хадсон Джей Джей; Сюй Д.В.; Го К.; Жуковская Н.; Лю ПХ; Уильямс Дж.Г.; Груши Си Джей; Лакин Н.Д. (2005). «ДНК-PKcs-зависимая передача сигналов о повреждении ДНК у Dictyostelium discoideum» . Курр Биол . 15 (20): 1880–5. Бибкод : 2005CBio...15.1880H . дои : 10.1016/j.cub.2005.09.039 . ПМИД 16243037 .
- ^ Кавендер, Джеймс С.; Насильник, Кеннет Б. (март 1965 г.). «Акрасии в природе. I. Изоляция» . Американский журнал ботаники . 52 (3): 294–296. дои : 10.1002/j.1537-2197.1965.tb06788.x . ISSN 0002-9122 .
- ^ «Иммуноблоттинг: равенство для слизевиков!». Биотехника (бумага). 55 (1): 9 июля 2013 г.
- ^ Дэвидсон, Эндрю Дж.; Кинг, Джейсон С.; Инсолл, Роберт Х. (июль 2013 г.). «Использование конъюгатов стрепавидина в качестве контроля нагрузки иммуноблоттинга митохондриальных маркеров для использования с Dictyostelium diskoidium» . Бенчмарки. Биотехника (бумага). 55 (1): 39–41. дои : 10.2144/000114054 . ПМИД 23834384 .
- ^ Брюн Х. 2008. Dictyostelium , управляемый модельный организм-хозяин легионеллы . В: Хойнер К., Суонсон М., редакторы. Легионелла : Молекулярная микробиология. Норидж (Великобритания): Caister Academic Press. ISBN 978-1-904455-26-4
- ^ Перейти обратно: а б «Амебы во время путешествий демонстрируют примитивное сельскохозяйственное поведение» , BBC News , 19 января 2011 г.
- ^ Перейти обратно: а б с д Брок Д.А., Дуглас Т.Е., Квеллер, Д.С., Страссманн Дж.Э. (20 января 2011 г.). «Примитивное земледелие в социальной амебе». Природа . 469 (7330): 393–396. Бибкод : 2011Natur.469..393B . дои : 10.1038/nature09668 . ПМИД 21248849 . S2CID 4333826 .
- ^ Перейти обратно: а б Кларк, Маргарет (2010). «Последние открытия о взаимодействии хозяина и патогена из Dyctiostelium» . Клеточная микробиология . 12 (3): 283–291. дои : 10.1111/j.1462-5822.2009.01413.x . ПМИД 19919566 .
- ^ Перейти обратно: а б Молмерет М., Хорн М., Вагнер М., Абу Квайк Ю. (январь 2005 г.). «Примитивные амебы как тренировочная площадка для внутриклеточных бактериальных патогенов» . Appl Environ Microbiol . 71 (1): 20–28. дои : 10.1128/АЕМ.71.1.20-28.2005 . ПМК 544274 . ПМИД 15640165 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Перейти обратно: а б Грант П.Оттом Мэри Ю.Ву; Маргарет Кларк; Хао Лу; О.Роджер Андерсон; Юбер Хильби; Говард А. Шуман; Ричард Х. Кессин (11 ноября 2003 г.). «Макроаутофагия необходима для внутриклеточной репликации Legionella pneumophila у Dictyostelium discoideum» . Молекулярная микробиология . 51 (1): 63–72. дои : 10.1046/j.1365-2958.2003.03826.x . ПМИД 14651611 . S2CID 22801290 .
- ^ Чен, Гуокай; Жученко Ольга; Куспа, Адам (3 августа 2007 г.). «Активность иммуноподобных фагоцитов у социальной амебы» . Наука . 317 (5838). Нью-Йорк, штат Нью-Йорк: 678–681. Бибкод : 2007Sci...317..678C . дои : 10.1126/science.1143991 . ISSN 0036-8075 . ПМК 3291017 . ПМИД 17673666 .
- ^ Брок, Дебра А.; Каллисон, В. Имон; Страссманн, Джоан Э.; Квеллер, Дэвид К. (27 апреля 2016 г.). «Сторожевые клетки, симбиотические бактерии и устойчивость к токсинам у социальной амебы Dictyostelium discoideum » . Учеб. Р. Сок. Б. 283 (1829): 20152727. doi : 10.1098/rspb.2015.2727 . ISSN 0962-8452 . ПМЦ 4855374 . ПМИД 27097923 .
- ^ Кавендер Джей Си; Шпигель Ф.; Суонсон А. (2002). «Таксономия, слизевики и вопросы, которые мы задаем». Микологическое общество Америки . 94 (6): 968–979. ПМИД 21156570 .
- ^ Эйхенгер Л.; и др. (2005). «Геном социальной амебы Dictyostelium discoideum » . Природа . 435 (7038): 34–57. Бибкод : 2005Natur.435...43E . дои : 10.1038/nature03481 . ПМЦ 1352341 . ПМИД 15875012 .
- ^ Балдауф С.Л.; Дулиттл В.Ф. (1997). «Происхождение и эволюция слизевиков (Mycetozoa)» . ПНАС . 94 (22): 12007–12012. Бибкод : 1997PNAS...9412007B . дои : 10.1073/pnas.94.22.12007 . ПМК 23686 . ПМИД 9342353 .
- ^ Шедвик, LL; Шпигель, ФРВ; Шедвик, доктор юридических наук; Браун, Миссури; Зильберман, доктор юридических наук (2009). «Eumycetozoa = Amoebozoa?: Филогения SSUrDNA протостелоидных слизевиков и ее значение для супергруппы амебозойных» . ПЛОС ОДИН . 4 (8): е6754. Бибкод : 2009PLoSO...4.6754S . дои : 10.1371/journal.pone.0006754 . ПМЦ 2727795 . ПМИД 19707546 .
Дальнейшее чтение
[ редактировать ]- Мэри С. Тайлер (2000). Биология развития: Руководство для экспериментальных исследований. 2-е изд . Синауэр Ассошиэйтс. стр. 31–34. ISBN 978-0-87893-843-8 .
- Скотт Ф. Гилберт (2006). Биология развития. 8-е изд . Синауэр. стр. 36–39. ISBN 978-0-87893-250-4 .
Внешние ссылки
[ редактировать ]- Модельные организмы для биомедицинских исследований: Dictyostelium discoideum.
- D. discoideum в MetaMicrobe: таксономия, факты, онтологии и ссылки
| Базы данных органов управления : Национальные |
|---|