Фотосинтетический реакционный центр

Фотосинтетический реакционный центр представляет собой комплекс нескольких белков, пигментов и других сопутствующих факторов, которые вместе выполняют первичные реакции преобразования энергии фотосинтеза . Молекулярные возбуждения, возникающие либо непосредственно от солнечного света, либо передаваемые в виде энергии возбуждения через светособирающие антенные системы , вызывают реакции переноса электронов по пути ряда связанных с белками кофакторов. Этими кофакторами являются светопоглощающие молекулы (также называемые хромофорами или пигментами ), такие как хлорофилл и феофитин , а также хиноны . Энергия фотона используется для возбуждения электрона пигмента. затем Созданная свободная энергия используется через цепочку близлежащих акцепторов электронов для переноса атомов водорода (как протонов и электронов) от H 2 O или сероводорода к диоксиду углерода, в конечном итоге производя глюкозу . Эти этапы переноса электронов в конечном итоге приводят к преобразованию энергии фотонов в химическую энергию.
Преобразование световой энергии в разделение зарядов
[ редактировать ]Реакционные центры имеются у всех зеленых растений , водорослей и многих бактерий . разнообразные светособирающие комплексы У фотосинтезирующих видов существуют . Зеленые растения и водоросли имеют два разных типа реакционных центров, которые являются частью более крупных суперкомплексов, известных как P700 в Фотосистеме I и P680 в Фотосистеме II . Структуры этих суперкомплексов большие и включают в себя множество светособирающих комплексов . Реакционный центр, обнаруженный у бактерий Rhodopseudomonas , в настоящее время лучше всего изучен, поскольку он был первым реакционным центром известной структуры и имел меньше полипептидных цепей , чем примеры у зеленых растений. [1]
Реакционный центр устроен таким образом, что он улавливает энергию фотона с помощью молекул пигмента и превращает ее в пригодную для использования форму. Как только световая энергия поглощается непосредственно молекулами пигмента или передается им путем резонансной передачи от окружающего светособирающего комплекса , они высвобождают электроны в цепь переноса электронов и передают энергию донору водорода, такому как H 2 O, для извлечения. электроны и протоны от него. У зеленых растений цепь переноса электронов имеет множество акцепторов электронов, включая феофитин , хинон , пластохинон , цитохром bf и ферредоксин , что в конечном итоге приводит к восстановленной молекуле НАДФН , в то время как энергия, используемая для расщепления воды, приводит к высвобождению кислорода . Прохождение электрона через цепь переноса электронов также приводит к перекачке протонов (ионов водорода) из в стромы хлоропласта , просвет , что приводит к образованию протонного градиента через тилакоидную мембрану который можно использовать для синтеза АТФ с использованием АТФ-синтаза молекула. И АТФ, и НАДФН используются в цикле Кальвина для связывания углекислого газа в триозные сахара.
Классификация
[ редактировать ]Выделяют два класса реакционных центров. Тип I, обнаруженный у зелено-серных бактерий , гелиобактерий и растений/цианобактерий PS-I, использует железо-серные кластеры в качестве акцепторов электронов. Тип II, обнаруженный в хлорофлексусах , пурпурных бактериях и растительных/цианобактериях PS-II, использует хиноны. Не только все члены каждого класса имеют общее происхождение, но и эти два класса благодаря общей структуре кажутся родственными. [2] [3]
Цианобактерии, предшественники хлоропластов, обнаруженные в зеленых растениях, имеют обе фотосистемы с обоими типами реакционных центров. Объединение двух систем позволяет производить кислород. [3]
У пурпурных бактерий (тип II)
[ редактировать ]В этом разделе рассматривается система типа II, обнаруженная у пурпурных бактерий. [3]
Структура
[ редактировать ]
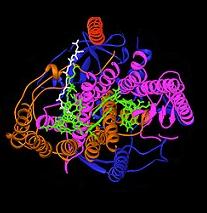
Бактериальный фотосинтетический реакционный центр стал важной моделью для понимания структуры и химии биологического процесса захвата световой энергии. В 1960-х годах Родерик Клейтон первым очистил комплекс реакционных центров от пурпурных бактерий. Однако первая кристаллическая структура (верхнее изображение справа) была определена в 1984 году Хартмутом Мишелем , Иоганном Дайзенхофером и Робертом Хубером. [4] за что они разделили Нобелевскую премию в 1988 году. [5] Это также было важно, поскольку это была первая трехмерная кристаллическая структура любого мембранного белкового комплекса.
Было обнаружено, что четыре различных субъединицы важны для функции фотосинтетического реакционного центра. Субъединицы L и M , показанные синим и фиолетовым цветом на изображении структуры, охватывают липидный бислой плазматической мембраны. Они структурно похожи друг на друга, оба имеют по 5 трансмембранных альфа-спиралей . [6] четыре бактериохлорофилла b молекулы бактериофеофитина b (BChl-b), две молекулы (BPh), два хинона (Q A и Q B С субъединицами L и M связаны ) и ион двухвалентного железа. Субъединица H, показанная золотом, расположена на цитоплазматической стороне плазматической мембраны. Субъединица цитохрома, здесь не показанная, содержит четыре гема c-типа и расположена на периплазматической поверхности (внешней) мембраны. Последняя субъединица не является общим структурным мотивом фотосинтезирующих бактерий. Субъединицы L и M связывают функциональные и взаимодействующие со светом кофакторы, показанные здесь зеленым цветом.
Реакционные центры разных видов бактерий могут содержать слегка измененные хромофоры бактериохлорофилла и бактериофеофита в качестве функциональных кофакторов. Эти изменения вызывают изменения цвета света, который может быть поглощен. Реакционный центр содержит два пигмента, которые служат для сбора и передачи энергии поглощения фотонов: BChl и Bph. БХл примерно напоминает молекулу хлорофилла, обнаруженную в зеленых растениях, но из-за незначительных структурных различий его пиковая длина волны поглощения сдвинута в инфракрасную область , с длиной волны до 1000 нм. Bph имеет ту же структуру, что и BChl, но центральный ион магния заменен двумя протонами. Это изменение вызывает как сдвиг максимума поглощения, так и снижение окислительно-восстановительного потенциала.
Механизм
[ редактировать ]
Процесс начинается, когда свет поглощается двумя молекулами БХл, расположенными вблизи периплазматической стороны мембраны. Эта пара молекул хлорофилла, часто называемая «специальной парой», поглощает фотоны с длиной волны 870 или 960 нм, в зависимости от вида, и, таким образом, называется P870 (для Rhodobacter sphaeroides ) или P960 (для Blastochromis viridis ), где P стоит «пигмент»). Как только P поглощает фотон, он выбрасывает электрон, который передается через другую молекулу Bchl на BPh в L-субъединице. Это первоначальное разделение зарядов дает положительный заряд на P и отрицательный заряд на BPH. Этот процесс происходит за 10 пикосекунд (10 −11 секунды). [1]
Обвинения по делу П. + и БФ − В этом состоянии может произойти рекомбинация зарядов, что приведет к потере энергии и преобразованию ее в тепло . Этому препятствуют несколько факторов строения реакционного центра. Во-первых, перенос электрона от БФ − до P960 + протекает относительно медленно по сравнению с двумя другими окислительно-восстановительными реакциями в реакционном центре. Более быстрые реакции связаны с переносом электрона от БФ. − (Бакалавр Ph − окисляется до BPh) до акцептора электронов хинона (Q A ), и происходит перенос электрона на P960 + (P960 + восстанавливается до P960) из гема в субъединице цитохрома над реакционным центром.
Высокоэнергетический электрон, находящийся на прочно связанной молекуле хинона Q A, переносится на обменную молекулу хинона Q B . Эта молекула слабо связана с белком и ее довольно легко отсоединить. необходимы два электрона Для полного восстановления Q B до QH 2 , при этом забирая два протона из цитоплазмы. Восстановленный хинон QH 2 диффундирует через мембрану к другому белковому комплексу ( цитохрома bc 1 комплексу ), где окисляется. При этом восстанавливающая способность QH 2 используется для перекачки протонов через мембрану в периплазматическое пространство. Электроны из комплекса цитохрома bc 1 затем передаются через растворимый промежуточный продукт цитохрома с, называемый цитохромом с 2 , в периплазме к субъединице цитохрома.
У цианобактерий и растений
[ редактировать ]Цианобактерии, предшественники хлоропластов, обнаруженные в зеленых растениях, имеют обе фотосистемы с обоими типами реакционных центров. Объединение двух систем позволяет производить кислород.
Кислородный фотосинтез
[ редактировать ]В 1772 году химик Джозеф Пристли провел серию экспериментов, касающихся газов, участвующих в дыхании и горении. В своем первом эксперименте он зажег свечу и поставил ее под перевернутую банку. Через небольшой промежуток времени свеча перегорела. Аналогичный эксперимент он провел с мышами в замкнутом пространстве горящей свечи. Он обнаружил, что мышь умерла вскоре после того, как погасла свеча. Однако он мог оживить грязный воздух, посадив здесь зеленые растения и выставив их на свет. Наблюдения Пристли были одними из первых экспериментов, продемонстрировавших активность фотосинтетического реакционного центра.
В 1779 году Ян Ингенхауз провел более 500 экспериментов в течение 4 месяцев, пытаясь понять, что происходит на самом деле. Свои открытия он описал в книге « Опыты над овощами » . Ингенхауз взял зеленые растения и погрузил их в воду в прозрачный резервуар. Он наблюдал множество пузырьков, поднимающихся с поверхности листьев всякий раз, когда растения подвергались воздействию света. Ингенхауз собрал газ, выделяемый заводами, и провел несколько различных тестов, пытаясь определить, что это был за газ. Тест, который, наконец, выявил подлинность газа, заключался в помещении тлеющей свечи в образец газа и повторном зажигании. Это испытание показало, что это был кислород, или, как назвал его Джозеф Пристли, «дефлогистизированный воздух ».
В 1932 году Роберт Эмерсон и его ученик Уильям Арнольд использовали метод повторяющихся вспышек для точного измерения небольших количеств кислорода, выделяемого хлорофиллом в водорослях хлорелле . Их эксперимент доказал существование фотосинтетической единицы. Позже Гаффрон и Воль интерпретировали эксперимент и поняли, что свет, поглощенный фотосинтетической единицей, передавался. [7] Эта реакция происходит в реакционном центре Фотосистемы II и протекает у цианобактерий, водорослей и зеленых растений. [8]
Фотосистема II
[ редактировать ]
Фотосистема II — это фотосистема, генерирующая два электрона, которые в конечном итоге восстанавливают НАДФ. + в ферредоксин-НАДФ-редуктазе. Фотосистема II присутствует на мембранах тилакоидов внутри хлоропластов, места фотосинтеза зеленых растений. [9] Структура Фотосистемы II удивительно похожа на бактериальный реакционный центр, и предполагается, что они имеют общего предка.
Ядро Фотосистемы II состоит из двух субъединиц, называемых D1 и D2 . Эти две субъединицы подобны субъединицам L и M, присутствующим в бактериальном реакционном центре. Фотосистема II отличается от бактериального реакционного центра тем, что имеет множество дополнительных субъединиц, которые связывают дополнительные хлорофиллы для повышения эффективности. Общая реакция, катализируемая Фотосистемой II, следующая:
- 2Q + 2H 2 О + hν → О 2 + 2QH 2
Q представляет собой окисленную форму пластохинона, а QH 2 представляет собой его восстановленную форму. Этот процесс восстановления хинона сравним с тем, который происходит в бактериальном реакционном центре. Фотосистема II получает электроны путем окисления воды в процессе, называемом фотолизом . Молекулярный кислород является побочным продуктом этого процесса, и именно эта реакция снабжает атмосферу кислородом . Тот факт, что кислород зеленых растений происходит из воды, впервые установил американский биохимик канадского происхождения Мартин Дэвид Кэмен . Он использовал стабильный изотоп кислорода. 18 О, проследить путь кислорода от воды до газообразного молекулярного кислорода. Эта реакция катализируется реактивным центром Фотосистемы II, содержащим четыре марганца иона .

Реакция начинается с возбуждения пары молекул хлорофилла, аналогичных молекулам бактериального реакционного центра. Благодаря наличию хлорофилла а , в отличие от бактериохлорофилла , Фотосистема II поглощает свет с более короткой длиной волны. Пару молекул хлорофилла в реакционном центре часто называют P680 . [1] Когда фотон поглощается, образующийся высокоэнергетический электрон передается ближайшей молекуле феофитина. Он находится выше и правее пары на диаграмме и окрашен в серый цвет. Электрон проходит от молекулы феофитина через две молекулы пластохинона, первая прочно связанная, вторая слабосвязанная. Прочно связанная молекула показана над молекулой феофитина и окрашена в красный цвет. Слабо связанная молекула находится слева от нее и также окрашена в красный цвет. Этот поток электронов подобен потоку бактериального реакционного центра. Два электрона необходимы для полного восстановления слабосвязанной молекулы пластохинона до QH 2, а также для поглощения двух протонов.
Отличие Фотосистемы II от бактериального реакционного центра заключается в источнике электронов, нейтрализующих пару молекул хлорофилла а . В бактериальном реакционном центре электрон получается из восстановленной гем-группы в субъединице цитохрома или из водорастворимого белка цитохрома-с.
Каждый раз, когда P680 поглощает фотон, он отдает электрон феофитину, получая положительный заряд. После фотоиндуцированного разделения зарядов P680 + является очень сильным окислителем высокой энергии. Он передает свою энергию молекулам воды , которые связаны в марганцевом центре непосредственно под парой, и извлекает из них электрон. Этот центр, расположенный ниже и слева от пары на диаграмме, содержит четыре иона марганца, ион кальция , ион хлорида и остаток тирозина . Марганец хорошо участвует в этих реакциях, поскольку он способен существовать в четырех степенях окисления: Mn 2+ , Мн 3+ , Мн 4+ и Мн 5+ . Марганец также образует прочные связи с кислородсодержащими молекулами, такими как вода. Процесс окисления двух молекул воды с образованием молекулы кислорода требует четырех электронов. Молекулы воды, окисляющиеся в марганцевом центре, являются источником электронов, которые восстанавливают две молекулы Q до QH 2 . На сегодняшний день этот каталитический центр расщепления воды не воспроизведен ни одним искусственным катализатором.
Фотосистема I
[ редактировать ]После того, как электрон покидает Фотосистему II, он переносится на комплекс цитохрома b6f , а затем на пластоцианин , белок синей меди и переносчик электронов. Комплекс пластоцианина несет электрон, который нейтрализует пару в следующем реакционном центре, Фотосистеме I.
Как и в случае с Фотосистемой II и бактериальным реакционным центром, пара молекул хлорофилла А инициирует фотоиндуцированное разделение зарядов. Эта пара называется P700 , где 700 — это длина волны , при которой молекулы хлорофилла максимально поглощают свет. P700 находится в центре белка. Как только фотоиндуцированное разделение зарядов инициировано, электрон проходит путь через молекулу хлорофилла α, расположенную непосредственно над P700, через молекулу хинона, расположенную непосредственно над ней, через три кластера 4Fe-4S и, наконец, к взаимозаменяемому комплексу ферредоксина. [10] Ферредоксин представляет собой растворимый белок, содержащий кластер 2Fe-2S, координированный четырьмя остатками цистеина. Положительный заряд высокоэнергетического P700 + нейтрализуется переносом электрона от пластоцианина , который получает энергию, которая в конечном итоге используется для преобразования QH 2 обратно в Q. Таким образом, общая реакция, катализируемая Фотосистемой I, такова:
- ПК (Cu + ) + Fd [ox] + hν → Pc(Cu 2+ ) + Fd [красный]
Сотрудничество Фотосистем I и II создает поток электронов и протонов от H 2 O к НАДФ. + , производя НАДФН, необходимый для синтеза глюкозы. Этот путь называется « Z-схемой », потому что окислительно-восстановительная диаграмма от H 2 O до НАДФ + через P680 и P700 напоминает букву Z. [11]
См. также
[ редактировать ]- Дикислород в биологических реакциях (кислород в биологических процессах)
- Светоуборочный комплекс
- Фотосинтез
- Фотосистема
- Фикобилисома
- Семейство белков фотосинтетического реакционного центра
Ссылки
[ редактировать ]- ^ Jump up to: а б с Берг Дж. М., Тимочко Дж. Л., Страйер Л. (2002). «Глава 19: Световые реакции фотосинтеза» . Биохимия (5-е изд.). Нью-Йорк: WH Freeman.
- ^ Садекар, С; Раймонд, Дж; Бланкеншип, RE (ноябрь 2006 г.). «Консервация отдаленно родственных мембранных белков: фотосинтетические реакционные центры имеют общее структурное ядро» . Молекулярная биология и эволюция . 23 (11): 2001–7. дои : 10.1093/molbev/msl079 . ПМИД 16887904 .
- ^ Jump up to: а б с Орф Г.С., Жизриэль С., Реддинг К.Э. (октябрь 2018 г.). «Эволюция фотосинтетических реакционных центров: понимание структуры гелиобактериального реакционного центра». Исследования фотосинтеза . 138 (1): 11–37. дои : 10.1007/s11120-018-0503-2 . ОСТИ 1494566 . ПМИД 29603081 . S2CID 4473759 .
- ^ Дайзенхофер Дж., Эпп О., Мики К., Хубер Р., Мишель Х. (декабрь 1984 г.). «Рентгеноструктурный анализ мембранного белкового комплекса. Карта электронной плотности с разрешением 3 А и модель хромофоров фотосинтетического реакционного центра Rhodopseudomonas viridis». Журнал молекулярной биологии . 180 (2): 385–98. дои : 10.1016/S0022-2836(84)80011-X . ПМИД 6392571 .
- ^ «Нобелевская премия по химии 1988 года» . NobelPrize.org .
- ↑ Фотосинтетические реакционные центры пурпурных бактерий. Архивировано 14 мая 2006 г. в Wayback Machine (2 февраля 1999 г.). Проверено 28 февраля 2010 г.
- ^ Говинджи Р. (2000). «Глава 1: Вехи фотосинтеза» . В Юнус М., Патре У., Моханти П. (ред.). Зондирующий фотосинтез: механизмы, регуляция и адаптация . Лондон: Тейлор и Фрэнсис. ISBN 978-0-7484-0821-4 .
- ^ Кайзер Г.Е. (24 февраля 2003 г.). «Кислородный фотосинтез: рост бактерий и микробный метаболизм» . Архивировано из оригинала 4 мая 2006 года.
- ^ «Хлоропласт» . Ультранет Биология . Архивировано из оригинала 3 августа 2003 года.
- ^ Джаганнатан Б., Голбек Дж. (2009). «Фотосинтез: микробный». В Шехтере М. (ред.). Энциклопедия микробиологии (3-е изд.). стр. 325–341. дои : 10.1016/B978-012373944-5.00352-7 . ISBN 978-0-12-373944-5 .
- ^ Говинджи Р. «Z-схема фотосинтеза» . Университет Иллинойса в Урбана-Шампейн.
Внешние ссылки
[ редактировать ]- Орр Л., Говинджи Р. (2013). «Веб-ресурсы по фотосинтезу» . Исследования фотосинтеза . 115 (2–3). Университет Иллинойса в Урбана-Шампейн: 179–214. дои : 10.1007/s11120-013-9840-3 . ПМИД 23708976 . S2CID 254943144 .