Бурые водоросли
| Бурые водоросли Временной диапазон: от поздней юры до настоящего времени | |
|---|---|
 | |
| Гигантские водоросли ( Macrocystispyrifera ) | |
| Научная классификация | |
| Домен: | Эукариоты |
| Клэйд : | потогонные средства |
| Клэйд : | САР |
| Клэйд : | Страменопилес |
| Тип: | Гириста |
| Подтип: | Охрофитина |
| Инфрафилум: | Крисиста |
| Суперкласс: | Фукистия |
| Сорт: | феофицеи Кьельман , 1891 г. [3] |
| Заказы | |
См. классификацию | |
| Синонимы | |
| |
Бурые водоросли ( син.: alga ) — большая группа многоклеточных водорослей, входящая в класс Phaeophyceae . К ним относятся многие водоросли, обитающие в более холодных водах Северного полушария . Бурые водоросли являются основными водорослями умеренных и полярных регионов. Многие бурые водоросли, например представители отряда Fucales , обычно растут вдоль скалистых берегов моря. Большинство бурых водорослей обитают в морской среде, где они играют важную роль как в качестве пищи, так и в качестве потенциальной среды обитания . Например, Macrocystis , водоросли отряда Laminariales , могут достигать 60 м (200 футов) в длину и образуют заметные подводные леса из водорослей , которые содержат высокий уровень биоразнообразия. [4] Другим примером является Саргассум , который создает уникальные плавающие маты из морских водорослей в тропических водах Саргассова моря , которые служат средой обитания для многих видов. Некоторые представители этого класса, например водоросли, используются человеком в пищу.
от 1500 до 2000 видов бурых водорослей. Во всем мире известно [5] Некоторые виды, такие как Ascophyllum nodosum , сами по себе стали объектами обширных исследований из-за их коммерческого значения. Они также имеют экологическое значение благодаря фиксации углерода . [4]
Бурые водоросли принадлежат к Stramenopiles , кладе эукариотических симбиотических организмов, которые отличаются от зеленых растений наличием хлоропластов, окруженных четырьмя мембранами, что позволяет предположить, что они были приобретены вторично в результате отношений между базальным эукариотом и красной или зеленой водорослью. Большинство бурых водорослей содержат пигмент фукоксантин , который отвечает за характерный зеленовато-коричневый цвет, давший им такое название. Бурые водоросли уникальны среди Stramenopiles, поскольку развиваются в многоклеточные формы с дифференцированными тканями , но размножаются с помощью жгутиковых спор и гамет , очень напоминающих клетки одноклеточных Stramenopiles. Генетические исследования показывают, что их ближайшими родственниками являются желто-зеленые водоросли .
Морфология
[ редактировать ]Бурые водоросли существуют в широком диапазоне размеров и форм. Самые маленькие представители группы вырастают в виде крошечных перистых пучков нитевидных клеток длиной не более нескольких сантиметров (несколько дюймов). [6] У некоторых видов есть стадия жизненного цикла, состоящая всего из нескольких клеток, что делает всю водоросль микроскопической. Другие группы бурых водорослей вырастают до гораздо больших размеров. и Морские водоросли кожистые водоросли часто являются наиболее заметными водорослями в их среде обитания. [7] высотой 60 см (2 фута) Размер водорослей может варьироваться от морской пальмы Postelsia до гигантской водоросли Macrocystispyrifera , длина которой достигает более 50 м (150 футов). [8] [9] и является самой крупной из всех водорослей. По форме бурые водоросли варьируются от небольших корок или подушечек. [10] до листовых свободно плавающих ковриков, образованных видами Sargassum . Они могут состоять из тонких, похожих на войлок нитей клеток, как у Ectocarpus , или из уплощенных ветвей длиной 30 см (1 фут), напоминающих веер, как у Padina .
Независимо от размера и формы, две видимые особенности отличают Phaeophyceae от всех других водорослей. Во-первых, члены группы обладают характерным цветом, который варьируется от оливково-зеленого до различных оттенков коричневого . Конкретный оттенок зависит от количества фукоксантина , присутствующего в водорослях. [11] Во-вторых, все бурые водоросли многоклеточные . Не существует известных видов, существующих в виде отдельных клеток или колоний клеток. [11] а бурые водоросли — единственная крупная группа морских водорослей , не включающая такие формы. Однако это может быть результатом классификации, а не следствием эволюции, поскольку все группы, предположительно ближайшие родственники коричневых, включают одноклеточные или колониальные формы. [ нужна ссылка ] Они могут менять цвет в зависимости от солености: от красноватого до коричневого.
Видимые структуры
[ редактировать ]
Какова бы ни была форма, тело всех бурых водорослей называется слоевищем , что указывает на отсутствие сложной ксилемы и флоэмы сосудистых растений . Это не означает, что у бурых водорослей полностью отсутствуют специализированные структуры. Но поскольку некоторые ботаники определяют «настоящие» стебли, листья и корни по наличию этих тканей, их отсутствие у бурых водорослей означает, что стеблеобразные и листовидные структуры, обнаруженные в некоторых группах бурых водорослей, должны описываться с помощью разная терминология. [12] Хотя не все бурые водоросли имеют сложную структуру, те, которые обычно имеют одну или несколько характерных частей.
Крепление — это корневидная структура , присутствующая в основании водоросли. Подобно корневой системе растений, опора служит для закрепления водоросли на субстрате, где она растет, и, таким образом, предотвращает унос водорослей течением. В отличие от корневой системы, опора обычно не служит основным органом поглощения воды и не получает питательные вещества из субстрата. Общий внешний вид держателя различается в зависимости от различных бурых водорослей и различных субстратов. Он может быть сильно разветвленным или иметь чашеобразную форму. У одной водоросли обычно есть только одна опора, хотя у некоторых видов из ее опоры растет более одной ножки.
Ножка — это стебель или стеблевидная структура, присутствующая в водоросли. Он может вырасти как короткая структура у основания водоросли (как у Laminaria ) или может развиться в большую сложную структуру, пронизывающую тело водоросли (как у Sargassum или Macrocystis ). У наиболее структурно дифференцированных бурых водорослей (таких как фукус ) ткани внутри ножки разделены на три отдельных слоя или области. Эти области включают центральную сердцевину, окружающую кору и внешний эпидермис, каждая из которых имеет аналог в стебле сосудистого растения. У некоторых бурых водорослей сердцевинная область включает ядро из удлиненных клеток, напоминающих флоэму по строению и функциям сосудистых растений. У других (например, Nereocystis ) центр ножки полый и заполнен газом, который поддерживает плавучесть этой части водоросли. Ножка может быть относительно гибкой и эластичной у таких видов, как Macrocystispyrifera , которые растут в условиях сильного течения, или может быть более жесткой у таких видов, как Postelsia palmaeformis. которые подвергаются воздействию атмосферы во время отлива.
Многие водоросли имеют уплощенную часть, которая может напоминать лист, и ее называют пластинкой , пластинкой или листвой . Название «лезвие» чаще всего применяется к одной неделимой структуре, тогда как «вайя» может применяться ко всему или большей части уплощенного тела водоросли, но это различие не применяется повсеместно. Название пластинка относится к той части структурно дифференцированной водоросли, которая уплощена. Это может быть одиночная или разделенная структура, которая может распространяться на значительную часть водоросли. Например, у водорослей пластинка представляет собой широкое тканевое крыло, которое непрерывно проходит вдоль обеих сторон разветвленной средней жилки . Средняя жилка и пластинка вместе составляют почти всю водоросль, так что пластинка распространена по всей водоросли, а не существует как локализованная ее часть.

У некоторых бурых водорослей имеется одна пластинка или пластинка, у других может быть множество отдельных пластинок. Даже у тех видов, которые изначально образуют одну лопасть, структура может разорваться при сильном течении или в процессе созревания с образованием дополнительных лопастей. Эти лезвия могут быть прикреплены непосредственно к ножке, к креплению без ножки или между ножкой и лезвием может быть воздушная камера. Поверхность пластинки или лезвия может быть гладкой или морщинистой; его ткани могут быть тонкими и гибкими или толстыми и кожистыми. У таких видов, как Egregia menziesii , эта характеристика может меняться в зависимости от турбулентности вод, в которых они растут. [6] У других видов поверхность пластинки покрыта слизью, чтобы препятствовать прикреплению эпифитов или отпугивать травоядных животных . Пластинки также часто являются частями водорослей, несущими репродуктивные структуры.
Газонаполненные поплавки, называемые пневмоцистами, обеспечивают плавучесть многим водорослям и представителям Fucales . Эти структуры, похожие на пузырь, возникают внутри пластинки или рядом с ней , поэтому она удерживается ближе к поверхности воды и, таким образом, получает больше света для фотосинтеза. Пневматоцисты чаще всего имеют сферическую или эллипсоидную форму , но у разных видов могут различаться по форме. Такие виды, как Nereocystis luetkeana и Pelagophycus porra, несут одну большую пневматоцисту между вершиной ножки и основанием лопастей. Напротив, гигантская водоросль Macrocystispyrifera имеет множество пластинок вдоль ножки, с пневматоцистой у основания каждой пластинки, где она прикрепляется к основной ножке. Виды Sargassum также несут много пластинок и пневматоцист, но оба вида структур прикреплены к ножке отдельно короткими ножками. У видов Fucus пневматоцисты развиваются внутри самой пластинки либо в виде отдельных сферических пузырей, либо в виде удлиненных газонаполненных областей, повторяющих контуры пластинки, в которой они развиваются.
Рост
[ редактировать ]
Бурые водоросли включают в себя самые крупные и быстрорастущие морские водоросли. [6] Вайи Macrocystis могут вырасти до 50 см (20 дюймов) в день, а ножки - на 6 см (2,4 дюйма) за один день. [13]
Рост у большинства бурых водорослей происходит на кончиках структур в результате делений одной апикальной клетки или ряда таких клеток. Это одноклеточные организмы. [7] По мере деления этой апикальной клетки новые клетки, которые она производит, развиваются во все ткани водоросли. Ветвления и другие боковые структуры появляются, когда апикальная клетка делится с образованием двух новых апикальных клеток. Однако некоторые группы (например, Ectocarpus ) растут за счет диффузного, нелокализованного производства новых клеток, которые могут возникать в любом месте слоевища. [11]
Организация ткани
[ редактировать ]Простейшие коричневые нитчатые, то есть их клетки удлиненные и имеют перегородки, разрезающие по ширине. Они ветвятся, расширяясь на кончике, а затем разделяя расширение. [14]
Эти нити могут быть гаплостихическими или полистихозными, многоосными или одноосными, образующими или не образующими псевдопаренхиму . [15] [16] Помимо листьев имеются крупные паренхиматозные водоросли с трехмерным развитием и ростом и различными тканями ( меристодерма , кора и мозговое вещество ), которые можно было бы считать морскими деревьями. [17] [18] Существуют также Fucales и Dictyotales, меньшие по размеру, чем водоросли, но все же паренхимные с такими же отчетливыми тканями.
Клеточная стенка состоит из двух слоев; внутренний слой обладает прочностью и состоит из целлюлозы ; Внешний слой стенки состоит в основном из альгина и во влажном состоянии становится липким, но при высыхании становится твердым и ломким. [15] В частности, клеточная стенка бурых водорослей состоит из нескольких компонентов, основными ингредиентами которых являются альгинаты и сульфатированный фукан , до 40% каждого из которых. [19] Целлюлоза, основной компонент стенок большинства растительных клеток, присутствует в очень небольшом процентном соотношении, до 8%. Пути биосинтеза целлюлозы и альгината, по-видимому, были приобретены от других организмов посредством эндосимбиотического и горизонтального переноса генов соответственно, тогда как сульфатированные полисахариды имеют наследственное происхождение. [20] В частности, целлюлозосинтазы, по-видимому, происходят от эндосимбионта красных водорослей фотосинтезирующего предка страменопилес, а предок бурых водорослей приобрел ключевые ферменты для биосинтеза альгинатов от актинобактерии . Наличие и тонкий контроль структуры альгината в сочетании с существовавшей до него целлюлозой потенциально дали бурым водорослям способность развивать сложные по структуре многоклеточные организмы, такие как ламинарии. [21]
Эволюционная история
[ редактировать ]Генетические и ультраструктурные данные относят Phaeophyceae к гетероконтам (Stramenopiles). [22] большая совокупность организмов, включающая как фотосинтезирующие представители с пластидами (такие как диатомовые водоросли ), так и нефотосинтезирующие группы (такие как слизевые сети и водяные плесени ). Хотя у некоторых гетероконтных родственников бурых водорослей отсутствуют пластиды в клетках, ученые полагают, что это результат эволюционной потери этой органеллы в этих группах, а не независимого приобретения несколькими фотосинтезирующими членами. [23] Таким образом, считается, что все гетероконты произошли от одного гетеротрофного предка, который стал фотосинтезирующим, когда приобрел пластиды в результате эндосимбиоза другого одноклеточного эукариота. [24]
К ближайшим родственникам бурых водорослей относятся одноклеточные и нитчатые виды, однако одноклеточные виды бурых водорослей не известны. Однако большинство ученых предполагают, что Phaeophyceae произошли от одноклеточных предков. [25] Сравнение последовательностей ДНК также позволяет предположить, что бурые водоросли произошли от нитчатых Phaeothamniophyceae . [26] Ксантофицеи , [27] или Хризофицеи [28] между 150 [1] и 200 миллионов лет назад. [2] Во многом эволюция бурых водорослей параллельна эволюции зеленых и красных водорослей . [29] так как все три группы обладают сложными многоклеточными видами с чередованием поколений . Анализ последовательностей 5S рРНК показывает гораздо меньшие эволюционные расстояния между родами бурых водорослей, чем между родами красных или зеленых водорослей. [2] [30] это говорит о том, что бурые водоросли диверсифицировались гораздо позже, чем две другие группы.
Окаменелости
[ редактировать ]Phaeophyceae в виде окаменелостей встречается редко из-за их, как правило, мягкотелого характера. [31] и ученые продолжают спорить об идентификации некоторых находок. [32] Частично проблема с идентификацией заключается в конвергентной эволюции морфологии многих бурых и красных водорослей. [33] Большинство окаменелостей мягкотканых водорослей сохраняют лишь уплощенные очертания без микроскопических признаков, позволяющих надежно различить основные группы многоклеточных водорослей. Среди бурых водорослей только виды рода Padina откладывают значительные количества минералов внутри или вокруг клеточных стенок. [34] Другие группы водорослей, такие как красные и зеленые водоросли , содержат ряд известковых представителей. Из-за этого они с большей вероятностью оставят следы в летописи окаменелостей, чем мягкие тела большинства бурых водорослей, и их чаще можно точно классифицировать. [35]
Окаменелости, по морфологии сравнимые с бурыми водорослями, известны из слоев, древних как верхний ордовик . [36] но таксономическое родство этих окаменелостей-отпечатков далеко не несомненно. [37] Утверждают, что более ранние эдиакарские окаменелости представляют собой бурые водоросли. [38] с тех пор были уволены. [26] Хотя многие углеродистые окаменелости были описаны из докембрия , они обычно сохраняются в виде уплощенных очертаний или фрагментов длиной всего лишь в миллиметры. [39] Поскольку у этих окаменелостей отсутствуют признаки, диагностические для идентификации даже на самом высоком уровне, их относят к таксонам ископаемых форм в соответствии с их формой и другими общими морфологическими особенностями. [40] Ряд девонских окаменелостей, названных фукоидами , из-за их внешнего сходства с видами рода Fucus оказались скорее неорганическими, чем настоящими окаменелостями. [31] Девонский мегаископаемый Prototaxites , состоящий из массы нитей, сгруппированных в стволообразные оси, считался возможной бурой водорослью. [11] Однако современные исследования отдают предпочтение новой интерпретации этого ископаемого как наземного гриба или грибоподобного организма. [41] Точно так же ископаемая протосальвиния когда-то считалась возможной бурой водорослью, но теперь считается ранним наземным растением . [42]
Ряд палеозойских окаменелостей предварительно был отнесен к бурым водорослям, хотя большинство из них также сравнивали с известными видами красных водорослей. Phascolophyllaphycus обладает многочисленными удлиненными вздутыми лопастями, прикрепленными к ножке. Это самая распространенная из окаменелостей водорослей, найденных в коллекции, сделанной из каменноугольных отложений в Иллинойсе . [43] Каждая полая пластинка несет в основании до восьми пневматоцист , а ножки, по-видимому, также были полыми и раздутыми. Эта комбинация характеристик аналогична некоторым современным родам отряда Laminariales (ламинарии). Несколько окаменелостей Drydenia и единственный экземпляр Hungerfordia из верхнего девона Нью-Йорка также сравнивали как с бурыми, так и с красными водорослями. [33] Ископаемые Drydenia состоят из эллиптического лезвия, прикрепленного к разветвляющейся нитевидной опоре, мало чем отличающейся от некоторых видов Laminaria , Porphyra или Gigartina . Единственный известный экземпляр Hungerfordia дихотомически разветвляется на доли и напоминает такие роды, как Chondrus и Fucus. [33] или Диктиота . [44]
Самые ранние известные окаменелости, которые можно с уверенностью отнести к Phaeophyceae, происходят из миоценовых диатомита отложений формации Монтерей в Калифорнии . [24] несколько мягких бурых макроводорослей, таких как Julescraneia . Было обнаружено [45]
Классификация
[ редактировать ]Филогения
[ редактировать ]На основе работы Зильберфельда, Руссо и де Ревье, 2014 г. [46]
Таксономия
[ редактировать ]Это список отрядов класса Phaeophyceae: [46] [47]
- Класс Phaeophyceae Hansgirg 1886 [Fucophyceae; Melanophycidae Rabenhorst 1863 stat. ноябрь Кавалер-Смит, 2006 г. ]
- Подкласс Discosporangiophycidae Silberfeld, Rousseau & Reviers 2014
- Отряд Discosporangiales Schmidt 1937, поправки. Каваи и др. 2007 год
- Семейство Choristocarpaceae. Кьельман 1891 г.
- Семейство Discosporangiaceae Schmidt 1937.
- Отряд Discosporangiales Schmidt 1937, поправки. Каваи и др. 2007 год
- Подкласс Ishigeophycidae Silberfeld, Rousseau & Reviers 2014
- Орден Исигеалес Чо и Бу 2004 г.
- Семейство Ishigeaceae Окамура 1935 г.
- Семейство Petrodermataceae Silberfeld, Rousseau & Reviers, 2014 г.
- Орден Исигеалес Чо и Бу 2004 г.
- Подкласс Dictyotophycidae Silberfeld, Rousseau & Reviers 2014
- Орден Dictyotales Bory de Saint-Vincent 1828 ex Phillips et al.
- Семейство Dictyotaceae Lamouroux ex Dumortier 1822 [Scoresbyellaceae Womersley 1987 ; Диктиопсиды]
- Орден Onsloviales Draisma & Prud'homme van Reine 2008 г.
- Семейство Onslowiaceae Draisma & Prud'homme van Reine, 2001 г.
- Орден Sphacelariales Migula 1909 г.
- Семейство Cladostephaceae Oltmanns 1922.
- Семейство Lithodermataceae Хаук 1883 г.
- Семейство Phaeostrofiaceae Kawai et al. 2005 г.
- Семейство Sphacelariaceae Decaisne 1842 г.
- Семейство Sphacelodermaceae Draisma, Prud'homme & Kawai 2010
- Семейство Stypocaulaceae Oltmanns 1922.
- Отряд Syringodermatales Генри 1984 г.
- Семейство Syringodermataceae Генри 1984 г.
- Орден Dictyotales Bory de Saint-Vincent 1828 ex Phillips et al.
- Подкласс Fucophycidae Кавальер-Смит 1986 г.
- Орден Аскосейралеса Петрова 1964 г. Дополнить. Мо и Генри, 1982 г.
- Семейство Ascoseiraceae Скоттсберг 1907 г.
- Отряд Asterocladales T.Silberfeld et al. 2011 год
- Семейство Asterocladaceae Silberfeld et al. 2011 год
- Орден Desmarestiales Setchell & Gardner 1925 г.
- Семейство Arthroladiaceae Chauvin 1842 г.
- Семейство Desmarestiaceae (Туре) Кьельман 1880 г.
- Орден Ectocarpales Bessey 1907, исправить. Rousseau & Reviers 1999a [Chordariales Setchell & Gardner 1925 ; Dictyosiphonales Setchell & Gardner 1925 ; Scytosiphonales Feldmann 1949 ]
- Семейство Acinetosporaceae Hamel ex Feldmann 1937 [Pylaiellaceae; Пилаелловые]
- Семейство Adenocystaceae Rousseau et al. 2000 г., поправка. Зильберфельд и др. 2011 [Хордариопсидовые]
- Семейство Chordariaceae Greville 1830, исправлено. Петерс и Рамирес 2001 [Myrionemataceae]
- Семейство Ectocarpaceae Agardh 1828, исправление. Зильберфельд и др. 2011 год
- Семейство Petrospongiaceae Racault et al. 2009 год
- Семейство Scytosiphonaceae Ardissone & Straforello 1877 [Chnoosporaceae Setchell & Gardner 1925 ]
- Орден Fucales Bory de Saint-Vincent 1827 [Notheiales Womersley 1987 ; Дурвиллеалес Петров 1965 ]
- Семейство Bifurcariopsidaceae Cho et al. 2006 г.
- Семейство Durvilleeaceae (Oltmanns) Де Тони 1891 г.
- Семейство Fucaceae Адансон 1763 г.
- Семейство Himantaliaceae (Кьельман) Де Тони 1891 г.
- Семейство Hormosiraceae Fritsch 1945.
- Семейство Notheiaceae Шмидта 1938 г.
- Семейство Sargassaceae Kützing 1843 [Cystoseiraceae De Toni 1891 ]
- Семейство Seirococcaceae Низамуддин 1987 г.
- Семейство Xiphophoraceae Cho et al. 2006 г.
- Отряд Laminariales Migula 1909 [Phaeosiphoniellales Silberfeld, Rousseau & Reviers 2014 ord. Ноябрь реквизит. ]
- Семейство Agaraceae Postels & Ruprecht 1840 [Costariaceae]
- Семейство Akkesiphycaceae Kawai & Sasaki 2000 г.
- Семейство Alariaceae Setchell & Gardner, 1925 г.
- Семейство Aureophycaceae Kawai & Ridgway 2013
- Семейство Хордовые Дюмортье 1822 г.
- Семейство Laminariaceae Bory de Saint-Vincent 1827 [Arthrothamnaceae Petrov 1974 ]
- Семейство Lessoniaceae Setchell & Gardner, 1925 г.
- Семейство Pseudochordaceae Каваи и Куроги 1985 г.
- Отряд Nemodermatales Parente et al. 2008 год
- Семейство Nemodermataceae Kuckuck ex Feldmann 1937.
- Орден Phaeosiphoniellales Silberfeld, Rousseau & Reviers 2014
- Семейство Phaeosiphoniellaceae Phillips et al. 2008 год
- Заказать Ralfsiales Nakamura и Lim & Kawai 2007 г.
- Семейство Mesosporaceae Танака и Чихара 1982 г.
- Семейство Neoralfsiaceae Lim & Kawai 2007 г.
- Семейство Ralfsiaceae Farlow 1881 [Heterochordariaceae Setchell & Gardner 1925 ]
- Отряд Scytothamnales Peters & Clayton 1998, поправки. Зильберфельд и др. 2011 год
- Семейство Asteronemataceae Silberfeld et al. 2011 год
- Семейство Bachelotiaceae Silberfeld et al. 2011 год
- Семейство Splachnidiaceae Mitchell & Whitting 1892 [Scytothamnaceae Womersley 1987 ]
- Орден Sporochnales Sauvageau 1926 г.
- Семейство Sporochnaceae Гревиль 1830 г.
- Отряд Tilopteridales Bessey 1907 исправьте. Филлипс и др. 2008 [Кутлериалес Бесси 1907 ]
- Семейство Cutleriaceae Гриффит и Хенфри 1856 г.
- Семейство Halosiphonaceae Каваи и Сасаки.
- Семейство Phyllariaceae Tilden 1935.
- Семейство Stschapoviaceae Kawai 2004 г.
- Семейство Tilopteridaceae Кьельман 1890 г.
- Орден Аскосейралеса Петрова 1964 г. Дополнить. Мо и Генри, 1982 г.
- Подкласс Discosporangiophycidae Silberfeld, Rousseau & Reviers 2014
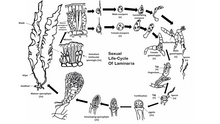
Жизненный цикл
[ редактировать ]Большинство бурых водорослей, за исключением Fucales , осуществляют половое размножение посредством спорового мейоза . [48] Между поколениями водоросли проходят отдельные фазы спорофита ( диплоидная ) и гаметофита ( гаплоидная ). Стадия спорофита часто более заметна из двух, хотя некоторые виды бурых водорослей имеют схожие диплоидную и гаплоидную фазы. Свободно плавающие формы бурых водорослей часто не подвергаются половому размножению до тех пор, пока не прикрепятся к субстрату. Гаплоидное поколение состоит из мужских и женских гаметофитов . [49] Оплодотворение яйцеклеток варьируется в зависимости от вида бурых водорослей и может быть изогамным , оогамным или анизогамным . Оплодотворение может происходить в воде яйцеклетками и подвижными сперматозоидами или внутри самого оогония .
Некоторые виды бурых водорослей могут также осуществлять бесполое размножение путем образования подвижных диплоидных зооспор . Эти зооспоры образуются в многогнездных спорангиях и могут сразу же перейти в фазу спорофита.
У репрезентативного вида Laminaria наблюдается заметное диплоидное поколение и более мелкие гаплоидные поколения. Мейоз происходит в нескольких одногнездных спорангиях вдоль пластинки водоросли, каждый из которых образует гаплоидные мужские или женские зооспоры . Затем споры высвобождаются из спорангиев и растут, образуя мужские и женские гаметофиты. Женский гаметофит производит яйцеклетку в оогонии, а мужской гаметофит выделяет подвижные сперматозоиды, которые оплодотворяют яйцеклетку. Оплодотворенная зигота затем превращается в зрелый диплоидный спорофит.
У отряда Fucales половое размножение оогамно , и зрелый диплоид — единственная форма для каждого поколения. Гаметы образуются в специализированных концептакулах , которые разбросаны по обеим поверхностям цветоложа, внешней части пластинок родительского растения. Яйцеклетки и подвижные сперматозоиды выходят из отдельных мешочков в зачатках родительских водорослей и объединяются в воде для завершения оплодотворения. Оплодотворенная зигота оседает на поверхности, а затем дифференцируется в листовидное слоевище и пальцеобразную опору. Свет регулирует дифференциацию зиготы на лезвие и опору. (самай великий)

Экология
[ редактировать ]Бурые водоросли адаптировались к широкому спектру морских экологических ниш, включая зону приливных брызг, каменные водоемы, всю приливную зону и относительно глубокие прибрежные воды. Они являются важным компонентом некоторых экосистем солоноватой воды и колонизировали пресную воду как минимум шесть известных случаев. [50] Большое количество Phaeophyceae обитает в приливной или верхней прибрежной зоне. [26] и это преимущественно прохладно- и холодноводные организмы, которые извлекают выгоду из питательных веществ из восходящих потоков холодной воды и притоков с суши; Саргассум является ярким исключением из этого обобщения.
Бурые водоросли, растущие в солоноватой воде, почти исключительно бесполы. [26]
Химия
[ редактировать ]| Группа водорослей | д 13 Диапазон С [51] |
|---|---|
| HCO 3 – с использованием красных водорослей | от −22,5 ‰ до −9,6 ‰ |
| CO 2 – с использованием красных водорослей | от −34,5 ‰ до −29,9 ‰ |
| Бурые водоросли | от −20,8‰ до −10,5‰ |
| Зеленые водоросли | от −20,3‰ до −8,8‰ |
Бурые водоросли имеют δ 13 Значение C находится в диапазоне от -30,0‰ до -10,5‰, в отличие от красных и зеленых водорослей. Это отражает их разные метаболические пути. [52]
Они имеют целлюлозные стенки с альгиновой кислотой , а также содержат полисахарид фукоидан в аморфных участках клеточных стенок. Некоторые виды ( Padina ) кальцифицируются с иглами арагонита . [26]
Помимо альгинатов, фукоидана и целлюлозы, углеводный состав бурых водорослей состоит из маннита , ламинарина и глюкана . [53]
Фотосинтетическая система бурых водорослей состоит из комплекса Р700 , содержащего хлорофилл а . Их пластиды также содержат хлорофилл С и каротиноиды (наиболее распространенные из них — фукоксантин ). [54]
Бурые водоросли производят особый тип танина, называемый флоротаннином, в больших количествах, чем красные водоросли.
Важность и использование
[ редактировать ]Бурые водоросли включают ряд съедобных морских водорослей . Все бурые водоросли содержат в клеточных стенках альгиновую кислоту (альгинат), которую добывают в коммерческих целях и используют в качестве промышленного загустителя в пищевых продуктах и для других целей. [55] Один из этих продуктов используется в литий-ионных аккумуляторах . [56] Альгиновая кислота используется в качестве стабильного компонента анода аккумуляторной батареи . Этот полисахарид является основным компонентом бурых водорослей и не встречается в наземных растениях.
Альгиновую кислоту также можно использовать в аквакультуре . Например, альгиновая кислота повышает иммунитет радужной форели. Молодые рыбы с большей вероятностью выживут, если на диете будет содержаться альгиновая кислота. [57]
Бурые водоросли, в том числе заросли ламинарии , также ежегодно фиксируют значительную часть углекислого газа на Земле посредством фотосинтеза. [58] Кроме того, они могут хранить большое количество углекислого газа, что может помочь нам в борьбе с изменением климата. [59] Было показано, что саргахроманол G , экстракт Sargassum siliquastrum , оказывает противовоспалительное действие. [60]
Съедобные бурые водоросли
[ редактировать ]Келп (Laminariales)[ редактировать ]
| Фукалес[ редактировать ]
|
|
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б Медлин, ЛК; и др. (1997). «Филогенетические взаимоотношения« золотых водорослей »(гаптофитов, гетероконтных хромофитов) и их пластид». Происхождение водорослей и их пластид (PDF) . Систематика и эволюция растений. Том. 11. С. 187–219. дои : 10.1007/978-3-7091-6542-3_11 . hdl : 10013/epic.12690 . ISBN 978-3-211-83035-2 . Архивировано (PDF) из оригинала 5 октября 2013 года . Проверено 27 августа 2012 г.
- ^ Перейти обратно: а б с Лим, Б.-Л.; Каваи, Х.; Хори, Х.; Осава, С. (1986). «Молекулярная эволюция 5S рибосомальной РНК красных и бурых водорослей» . Японский журнал генетики . 61 (2): 169–176. дои : 10.1266/jjg.61.169 . Архивировано из оригинала 10 января 2015 года . Проверено 27 августа 2012 г.
- ^ Кьельман, Ф.Р. (1891). «Phaeophyceae (Fucoideae)». В Энглер, А.; Прантл, К. (ред.). Семейства природных растений . Том 1. Лейпциг: Вильгельм Энгельманн . стр. 176–192.
- ^ Перейти обратно: а б Кок, Дж. Марк; Питерс, Акира Ф.; Коэльо, Сусана М. (9 августа 2011 г.). «Бурые водоросли» . Современная биология . 21 (15): R573–R575. дои : 10.1016/j.cub.2011.05.006 . ПМИД 21820616 .
- ^ Хук, Кристиан; ден Хёк, Хёк Ван; Манн, Дэвид; Янс, Х.М. (1995). Водоросли: введение в психологию . Издательство Кембриджского университета. п. 166. ИСБН 9780521316873 . OCLC 443576944 . Архивировано из оригинала 16 октября 2023 года . Проверено 26 апреля 2018 г.
- ^ Перейти обратно: а б с Коннор, Дж.; Бакстер, К. (1989). Келповые леса . Аквариум Монтерей Бэй . ISBN 978-1-878244-01-7 .
- ^ Перейти обратно: а б Диттмер, HJ (1964). Филогения и форма в царстве растений . Принстон, Нью-Джерси: Компания Д. Ван Ностранда . стр. 115–137 . ISBN 978-0-88275-167-2 .
- ^ Эбботт, Айова; Холленберг, Дж.Дж. (1976). Морские водоросли Калифорнии . Калифорния: Издательство Стэнфордского университета . ISBN 978-0-8047-0867-8 .
- ^ Крибб, AB (1953). « Macrocystispyrifera (L.) Ag. в водах Тасмании». Австралийский журнал исследований морской и пресной воды . 5 (1): 1–34. дои : 10.1071/MF9540001 .
- ^ Джонс, МЫ (1962). «Определитель родов британских морских водорослей» (PDF) . Полевые исследования . 1 (4): 1–32. [ постоянная мертвая ссылка ]
- ^ Перейти обратно: а б с д Смелый, ХК; Алексопулос, CJ; Делеворяс, Т. (1987). Морфология растений и грибов (5-е изд.). Нью-Йорк: Издательство Harper & Row . стр. 112–131, 174–186. ISBN 978-0-06-040839-8 .
- ^ Рэйвен, штат Пенсильвания; Эверт, РФ; Эйххорн, SE (2005). Биология растений (7-е изд.). Нью-Йорк: WH Freeman and Company . стр. 316–321 , 347. ISBN. 978-0-7167-1007-3 .
- ^ Раунд, FE (1981). Экология водорослей . Кембридж: Издательство Кембриджского университета . п. 103. ИСБН 978-0-521-26906-3 .
- ^ Винн, MJ (1981). «Биология морских водорослей» . В Лоббане, CS; Винн, MJ (ред.). Phaeophyta: морфология и классификация . Ботанические монографии. Том. 17. Издательство Калифорнийского университета . п. 52. ИСБН 978-0-520-04585-9 . Архивировано из оригинала 16 октября 2023 года . Проверено 11 мая 2020 г.
- ^ Перейти обратно: а б Шарма, ОП (1986). Учебник Водорослей . Тата МакГроу-Хилл . п. 298. ИСБН 978-0-07-451928-8 .
- ^ Грэм; Уилкокс; Грэм (2009). Водоросли, 2-е издание . Пирсон. ISBN 9780321603128 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Фрич, Ф.Е. 1945. Структура и размножение водорослей. Издательство Кембриджского университета, Кембридж.
- ^ Шарье, Бенедикт; Ле Бэйль, Од; де Ревье, Бруно (август 2012 г.). «Растение Протей: морфологическая пластичность бурых водорослей и основные механизмы развития» . Тенденции в науке о растениях . 17 (8): 468–477. doi : 10.1016/j.tplants.2012.03.003 . ISSN 1360-1385 . ПМИД 22513108 . Архивировано из оригинала 16 октября 2023 года . Проверено 13 ноября 2020 г.
- ^ Шарье, Бенедикт; Рабилле, Эрве; Биллоуд, Бернар (2019). «Наблюдение за расширением клеточной стенки под золотым светом» (PDF) . Тенденции в науке о растениях . 24 (2): 130–141. doi : 10.1016/j.tplants.2018.10.013 . ПМИД 30472067 . S2CID 53725259 . Архивировано (PDF) из оригинала 8 января 2022 года . Проверено 17 сентября 2022 г.
- ^ Мишель, Гурван; Тонон, Тьерри; Скорне, Дельфина; Кок, Дж. Марк; Клоарег, Бернард (2010). «Метаболизм полисахаридов клеточной стенки бурой водоросли Ectocarpus siliculosus. Взгляд на эволюцию полисахаридов внеклеточного матрикса у эукариот» . Новый фитолог . 188 (1): 82–97. дои : 10.1111/j.1469-8137.2010.03374.x . ПМИД 20618907 .
- ^ Денио-Буэ, Эстель; Керварец, Нелли; Мишель, Гурван; Тонон, Тьерри; Клоарег, Бернард; Эрве, Сесиль (2014). «Химическое и ферментативное фракционирование клеточных стенок Fucales: понимание структуры внеклеточного матрикса бурых водорослей» . Анналы ботаники . 114 (6): 1203–1216. дои : 10.1093/aob/mcu096 . ПМЦ 4195554 . ПМИД 24875633 .
- ^ Адл, С.М.; и др. (2005). «Новая классификация эукариот более высокого уровня с упором на таксономию протистов» (PDF) . Журнал эукариотической микробиологии . 52 (5): 399–451. дои : 10.1111/j.1550-7408.2005.00053.x . ПМИД 16248873 . Архивировано (PDF) из оригинала 29 мая 2013 года . Проверено 27 августа 2012 г.
- ^ Лейн, CE; Арчибальд, Дж. М. (2008). «Эукариотическое древо жизни: эндосимбиоз берет свое TOL» (PDF) . Тенденции экологии и эволюции . 23 (5): 268–275. CiteSeerX 10.1.1.555.2930 . дои : 10.1016/j.tree.2008.02.004 . ПМИД 18378040 . Архивировано (PDF) из оригинала 20 июня 2013 года . Проверено 27 августа 2012 г.
- ^ Перейти обратно: а б ван ден Хук, К.; Манн, Д.Г.; Янс, Х.М. (1995). Водоросли: введение в психологию . Кембридж: Издательство Кембриджского университета . стр. 165–218. ISBN 978-0-521-31687-3 .
- ^ Никлас, KJ (1997). Эволюционная биология растений . Чикаго: Издательство Чикагского университета . п. 156. ИСБН 978-0-226-58082-1 .
- ^ Перейти обратно: а б с д и Ли, RE (2008). Психология (4-е изд.). Издательство Кембриджского университета . ISBN 978-0-521-63883-8 .
- ^ Аризтия, Е.В.; Андерсен, РА; Согин, М.Л. (1991). «Новая филогения хромофитных водорослей с использованием 16S-подобных последовательностей рРНК Mallomonas papillosa (Synurophyceae) и Tribonema aequale (Xanthophyceae)». Журнал психологии . 27 (3): 428–436. дои : 10.1111/j.0022-3646.1991.00428.x . S2CID 84693030 .
- ^ Тейлор, Теннесси; Тейлор, Э.Л. (1993). Биология и эволюция ископаемых растений . Энглвуд Клиффс, Нью-Джерси: Прентис Холл . стр. 128–131. ISBN 978-0-13-651589-0 .
- ^ Диттмер, HJ (1964). Филогения и форма в царстве растений . Принстон, Нью-Джерси: Компания Д. Ван Ностранда. стр. 115–137 . ISBN 978-0-88275-167-2 .
- ^ Хори, Х.; Осава, С. (1987). «Происхождение и эволюция организмов, выведенная на основе последовательностей 5S рибосомальных RNS» . Молекулярная биология и эволюция . 4 (5): 445–472. doi : 10.1093/oxfordjournals.molbev.a040455 . ПМИД 2452957 .
- ^ Перейти обратно: а б Арнольд, Калифорния (1947). Введение в палеоботанику . Нью-Йорк; Лондон: МакГроу-Хилл . п. 48 . ISBN 978-1-4067-1861-4 .
- ^ Койер, Дж. А.; Смит, Дж.Дж.; Андерсен, Р.А. (2001). «Эволюция видов Macrocystis (Phaeophyta), определенная по последовательностям ITS1 и ITS2» (PDF) . Журнал психологии . 37 (4): 574–585. дои : 10.1046/j.1529-8817.2001.037001574.x . S2CID 84074495 . Архивировано из оригинала 16 октября 2023 года . Проверено 27 августа 2012 г.
- ^ Перейти обратно: а б с Фрай, WL; Бэнкс, HP (1955). «Три новых рода водорослей из верхнего девона Нью-Йорка». Журнал палеонтологии . 29 (1): 37–44. JSTOR 1300127 .
- ^ Прескотт, GW (1968). Водоросли: обзор . Бостон: Компания Houghton Mifflin . стр. 207–231 , 371–372. ISBN 978-3-87429-244-3 .
- ^ Симпсон, Г.Г. (1953). Жизнь прошлого: введение в палеонтологию . Нью-Хейвен: Издательство Йельского университета . стр. 158–159 .
- ^ Фрай, WL (1983). «Водорослевая флора из верхнего ордовика региона озера Виннипег, Манитоба, Канада». Обзор палеоботаники и палинологии . 39 (3–4): 313–341. Бибкод : 1983RPaPa..39..313F . дои : 10.1016/0034-6667(83)90018-0 .
- ^ Шпеер, БР; Ваггонер, Б.М. (2000). «Phaeophyta: Летопись окаменелостей» . Архивировано из оригинала 9 июня 2007 года . Проверено 24 апреля 2007 г.
- ^ Леблих, Арканзас (1974). «Протистанская филогения, указанная в летописях окаменелостей». Таксон . 23 (2/3): 277–290. дои : 10.2307/1218707 . JSTOR 1218707 .
- ^ Хофманн, HJ (1985). «Докембрийские углеродистые мегаокаменелости». В Д. Ф. Туми; М. Х. Нитецкий (ред.). Палеоалгология: современные исследования и приложения . Берлин: Springer-Verlag . стр. 20–33.
- ^ Хофманн, HJ (1994). «Протерозойские углеродистые сжатия («метафиты» и «черви»)». В Бенгтсоне, С. (ред.). Жизнь на Земле . Нобелевский симпозиум. Том. 84. Нью-Йорк: Издательство Колумбийского университета . стр. 342–357.
- ^ Хубер, FM (2001). «Гнилая древесина-водоросль-гриб: история и жизнь прототакситов Доусона 1859». Обзор палеоботаники и палинологии . 116 (1): 123–158. Бибкод : 2001RPaPa.116..123H . дои : 10.1016/S0034-6667(01)00058-6 .
- ^ Тейлор, Вашингтон; Тейлор, Теннесси (1987). «Ультраструктура стенки спор протосальвинии » (PDF) . Американский журнал ботаники . 74 (3): 437–443. дои : 10.2307/2443819 . JSTOR 2443819 . Архивировано из оригинала (PDF) 17 июня 2010 года.
- ^ Лири, РЛ (1986). «Три новых рода ископаемых неизвестковых водорослей из слоев Валмейера (Миссисипи) штата Иллинойс». Американский журнал ботаники . 73 (3): 369–375. дои : 10.2307/2444080 . JSTOR 2444080 .
- ^ Смелый, ХК; Винн, MJ (1978). Введение в водоросли (2-е изд.). Прентис-Холл . п. 27 . ISBN 978-0-13-477786-3 .
- ^ Паркер, Британская Колумбия; Доусон, EY (1965). «Безизвестковые морские водоросли из отложений миоцена Калифорнии». Нова Хедвигия . 10 : 273–295, листы 76–96.
- ^ Перейти обратно: а б Зильберфельд, Томас; Руссо, Флоренция; де Ревье, Бруно (2014). «Обновленная классификация бурых водорослей (Ochrophyta, Phaeophyceae)». Криптогамия, Алгология . 35 (2): 117–156. дои : 10.7872/crya.v35.iss2.2014.117 . S2CID 86227768 .
- ^ Гири, доктор медицины; Гири, генеральный менеджер (2009). «АлгаБаза» . Национальный университет Ирландии . Архивировано из оригинала 12 октября 2009 года . Проверено 31 декабря 2012 г.
- ^ Смелый, Гарольд Чарльз; Винн, Майкл Джеймс (1 января 1985 г.). Знакомство с водорослями: строение и размножение . Прентис-Холл. ISBN 9780134777467 . Архивировано из оригинала 16 октября 2023 года . Проверено 11 мая 2020 г.
- ^ Лесли., Ловетт-Дуст, Джон и (1 января 1990 г.). Репродуктивная экология растений: закономерности и стратегии . Издательство Оксфордского университета. ISBN 9780198021926 . OCLC 437173314 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Диттами, С.М.; Хиш, С; Олсен, Дж.Л.; Коллен, Дж (2017). «Переходы между морской и пресноводной средой дают новые сведения о происхождении многоклеточных растений и водорослей» (PDF) . Дж. Фикол . 53 (4): 731–745. дои : 10.1111/jpy.12547 . ПМИД 28509401 . S2CID 43325392 . Архивировано (PDF) из оригинала 26 февраля 2020 г. Проверено 11 мая 2020 г.
- ^ Маберли, Южная Каролина; Рэйвен, Дж.А.; Джонстон, AM (1992). «Дискриминация между 12 С и 13 C морскими растениями». Oecologia . 91 (4): 481. doi : 10.1007/BF00650320 . JSTOR 4220100 .
- ^ Флетчер, Би Джей; Берлинг, диджей ; Чалонер, WG (2004). «Стабильные изотопы углерода и метаболизм наземного девонского организма Spongiophyton». Геобиология . 2 (2): 107–119. Бибкод : 2004Gbio....2..107F . дои : 10.1111/j.1472-4677.2004.00026.x . S2CID 85079041 .
- ^ Ли, Ю.; Чжэн, Ю.; Чжан, Ю.; Ян, Ю.; Ван, П.; Имре, Б.; Вонг, AC; Се, Ю.С.; Ван, Д. (2021). «Углеводы бурых водорослей: структура, фармацевтические свойства и проблемы исследования» . Морские наркотики . 19 (11): 620. дои : 10.3390/md19110620 . ПМЦ 8623139 . ПМИД 34822491 .
- ^ Беркалофф, Клэр (1990). «Субъединичная организация частиц PSI из бурых водорослей и диатомовых водорослей: полипептидный и пигментный анализ». Исследования фотосинтеза . 23 (2): 181–193. дои : 10.1007/BF00035009 . ПМИД 24421060 . S2CID 7160955 .
- ^ «Альгиновая кислота» . www.фао.орг . Архивировано из оригинала 20 апреля 2017 года . Проверено 20 апреля 2017 г.
- ^ Коваленко Игорь; Здырко, Богдан; Магасинский, Александр; Герцберг, Бенджамин; Миличев, Зоран; Буртовой, Руслан; Лузинов Игорь; Юшин, Глеб (1 января 2011 г.). «Основной компонент бурых водорослей для использования в литий-ионных батареях большой емкости» . Наука 334 (6052): 75–79. Бибкод : 2011Наука...334... 75K дои : 10.1126/science.1209150 . JSTOR 23059304 . ПМИД 21903777 . S2CID 6523029 .
- ^ Джоаккини, Джорджия; Ломбардо, Франческо; Авелла, Маттео Алессандро; Оливото, Айк; Карнавалы, Олиана (1 апреля 2010 г.). «Улучшение благосостояния молоди радужной форели (Oncorhynchus mykiss) с помощью альгиновой кислоты». Химия и экология . 26 (2): 111–121. Бибкод : 2010ЧЭко..26..111Г . дои : 10.1080/02757541003627738 . hdl : 11566/39838 . ISSN 0275-7540 . S2CID 86175912 .
- ^ Васкес, Хулио А.; Суньига, Серджио; Тала, Фадия; Пиаже, Николь; Родригес, Дени К.; Вега, Х. М. Алонсо (1 апреля 2014 г.). «Экономическая оценка лесов водорослей на севере Чили: ценность товаров и услуг экосистемы». Журнал прикладной психологии . 26 (2): 1081–1088. дои : 10.1007/s10811-013-0173-6 . ISSN 0921-8971 . S2CID 14492051 .
- ^ Краузе-Йенсен, Д.; Дуарте, К. (21 февраля 2020 г.). «Существенная роль макроводорослей в секвестрации углерода в море». Природа Геонауки . 9 (10): 737–742. дои : 10.1038/ngeo2790 .
- ^ Юн, Веон-Чон; Хо, Су-Джин; Хан, Санг-Чул; Ли, Хе-Джа; Кан, Кён Джин; Кан, Хи-Гён; Хён, Джин Вон; Кох, Янг-Санг; Ю, Ын Сук (1 августа 2012 г.). «Противовоспалительное действие саргахроманола G, выделенного из Sargassum siliquastrum в клетках RAW 264.7». Архивы фармацевтических исследований 35 (8): 1421–1430. дои : 10.1007/ s12272-012-0812-5 ISSN 0253-6269 . ПМИД 22941485 . S2CID 39748571 .