Смена поколений

Смена поколений (также известная как метагенез или гетерогенез ) [ 1 ] — преобладающий тип цикла растений водорослей и жизненного . У растений обе фазы многоклеточные : гаплоидная половая фаза — гаметофит — чередуется с диплоидной бесполой фазой — спорофит .
Зрелый спорофит производит гаплоидные споры путем мейоза — процесса, который уменьшает количество хромосом вдвое, с двух наборов до одного. Образующиеся гаплоидные споры прорастают и превращаются в многоклеточные гаплоидные гаметофиты. По достижении зрелости гаметофит производит гаметы путем митоза — нормального процесса деления клеток у эукариот, который сохраняет исходное количество хромосом. Две гаплоидные гаметы (происходящие из разных организмов одного и того же вида или из одного и того же организма) сливаются , образуя диплоидную зиготу , которая неоднократно делится митозом, развиваясь в многоклеточный диплоидный спорофит. Этот цикл от гаметофита к спорофиту (или, в равной степени, от спорофита к гаметофиту) представляет собой путь наземных растений и большинства водорослей всех полового размножения .
Соотношение между фазами спорофита и гаметофита варьируется у разных групп растений. У большинства водорослей спорофит и гаметофит представляют собой отдельные независимые организмы, которые могут иметь или не иметь сходный внешний вид. У печеночников , мхов и роголистников спорофит развит хуже, чем гаметофит, и во многом зависит от него. Хотя спорофиты мхов и роголистников могут фотосинтезировать, им требуется дополнительный фотосинтез гаметофита для поддержания роста и развития спор, и они зависят от него в качестве источника воды, минеральных питательных веществ и азота. [ 2 ] [ 3 ] Напротив, у всех современных сосудистых растений гаметофит развит хуже, чем спорофит, хотя их девонские предки имели гаметофиты и спорофиты примерно одинаковой сложности. [ 4 ] У папоротников гаметофит представляет собой небольшой уплощенный автотрофный заросток , от которого на короткое время зависит питание молодого спорофита. У цветковых растений редукция гаметофита гораздо более выражена; он состоит всего из нескольких клеток, которые полностью растут внутри спорофита.
Животные развиваются по-разному. Они непосредственно производят гаплоидные гаметы. Гаплоидные споры, способные к делению, не образуются, поэтому многоклеточная гаплоидная фаза обычно отсутствует. У некоторых насекомых есть система определения пола , при которой гаплоидные самцы производятся из неоплодотворенных яиц; однако самки, полученные из оплодотворенных яиц, диплоидны.
Жизненные циклы растений и водорослей с чередованием гаплоидных и диплоидных многоклеточных стадий называются диплогаплонтическими . Эквивалентные термины гаплодиплонтный , дилобионтический и дибионтный также используются для описания такого организма как имеющего двухфазный онтогенез . [ 5 ] Жизненные циклы животных, в которых имеется только диплоидная многоклеточная стадия, называются диплоидными . Жизненные циклы, в которых присутствует только гаплоидная многоклеточная стадия, называются гаплонтическими .
Определение
[ редактировать ]Смена поколений определяется как чередование многоклеточных диплоидных и гаплоидных форм в жизненном цикле организма независимо от того, являются ли эти формы свободноживущими. [ 6 ] У некоторых видов, таких как водоросль Ulva Lactuca , диплоидная и гаплоидная формы действительно представляют собой свободноживущие независимые организмы, по существу идентичные по внешнему виду и поэтому называемые изоморфными . У многих водорослей свободноплавающие гаплоидные гаметы образуют диплоидную зиготу, которая прорастает в многоклеточный диплоидный спорофит. Спорофит в результате мейоза производит свободно плавающие гаплоидные споры , которые прорастают в гаплоидные гаметофиты. [ 7 ]
Однако у наземных растений ни спорофит, ни гаметофит сильно редуцированы и не способны к свободной жизни. Например, у всех мохообразных доминирует гаметофитное поколение и от него зависит спорофит. Напротив, у всех семенных растений гаметофиты сильно редуцированы, хотя ископаемые свидетельства указывают на то, что они произошли от изоморфных предков. [ 4 ] У семенных растений женский гаметофит полностью развивается внутри спорофита, который защищает и питает его, а также эмбриональный спорофит, который он производит. Пыльцевые зерна, являющиеся мужскими гаметофитами, сокращаются до нескольких клеток (во многих случаях всего до трех клеток). Здесь понятие двух поколений менее очевидно; как говорят Бейтман и Димишель, «спорофит и гаметофит эффективно функционируют как единый организм». [ 8 ] Альтернативный термин «чередование фаз» может оказаться более подходящим. [ 9 ]
История
[ редактировать ]У животных
[ редактировать ]Первоначально Адельберт фон Шамиссо (изучение сальп , колониальных морских животных между 1815 и 1818 годами). [ 10 ] ) и Япетус Стенструп (изучавший в 1842 году развитие трематод , а также оболочников и книдарий ) описывали смену разноорганизованных поколений (половых и бесполых) у животных как «чередование поколений». [ 11 ] Позднее явление у животных стало известно как гетерогамия , тогда как термин «чередование поколений» ограничился жизненными циклами растений, имея в виду именно чередование гаплоидных гаметофитов и диплоидных спорофитов. [ 11 ]
В растениях
[ редактировать ]В 1851 году Вильгельм Гофмейстер продемонстрировал морфологическую смену поколений у растений. [ 12 ] между спороносным поколением (спорофит) и гаметоносным поколением (гаметофит). [ 13 ] [ 14 ] К этому времени возникла дискуссия, сосредоточенная на происхождении бесполого поколения наземных растений (т. е. спорофита) и условно характеризуемая как конфликт между теориями антитетического ( Ладислав Йозеф Челаковский , 1874) и гомологического ( Натанаэль Прингсхайм , 1876) смена поколений. [ 11 ]
В 1874 году Эдуард Страсбургер обнаружил чередование диплоидных и гаплоидных ядерных фаз. [ 11 ] также называется цитологическим чередованием ядерных фаз. [ 15 ] Хотя чаще всего они совпадают, морфологическое чередование и чередование ядерных фаз иногда независимы друг от друга, например, у многих красных водорослей одна и та же ядерная фаза может соответствовать двум разным морфологическим поколениям. [ 15 ] У некоторых папоротников , утративших половое размножение, изменения ядерной фазы не происходит, но сохраняется чередование поколений. [ 16 ]
Смена поколений у растений
[ редактировать ]Фундаментальные элементы
[ редактировать ]На схеме выше показаны основные элементы смены поколений у растений. Существует множество вариаций у разных групп растений. Речь идет о следующих процессах: [ 17 ]
- Две одноклеточные гаплоидные гаметы, каждая из которых содержит n непарных хромосом, сливаются, образуя одноклеточную диплоидную зиготу, которая теперь содержит n пар хромосом, т. е. 2 n хромосом. всего [ 17 ]
- Одноклеточная диплоидная зигота прорастает, делясь путем нормального процесса ( митоза ), при котором сохраняется число хромосом на уровне 2 n . В результате образуется многоклеточный диплоидный организм, называемый спорофитом (поскольку при зрелости он производит споры). [ 17 ]
- Достигая зрелости, спорофит производит один или несколько спорангиев (единственное число: спорангий), которые являются органами, производящими диплоидные материнские клетки спор (спороциты). Они делятся посредством особого процесса ( мейоза ), который уменьшает число хромосом вдвое. Первоначально в результате образуются четыре одноклеточные гаплоидные споры, каждая из которых содержит n непарных хромосом. [ 17 ]
- Одноклеточная гаплоидная спора прорастает, делясь обычным процессом (митозом), при котором сохраняется число хромосом на уровне n . В результате образуется многоклеточный гаплоидный организм, называемый гаметофитом (поскольку в зрелом возрасте он производит гаметы). [ 17 ]
- Когда гаметофит достигает зрелости, он производит один или несколько гаметангиев (единственное число: гаметангиум), которые являются органами, производящими гаплоидные гаметы. По крайней мере, один вид гамет обладает каким-то механизмом достижения другой гаметы и слияния с ней. [ 17 ]
Таким образом, «чередование поколений» в жизненном цикле происходит между диплоидным (2 n ) поколением многоклеточных спорофитов и гаплоидным ( n ) поколением многоклеточных гаметофитов. [ 17 ]

Ситуация совершенно отличается от ситуации у животных, где фундаментальный процесс заключается в том, что многоклеточная диплоидная (2 n ) особь непосредственно производит гаплоидные ( n ) гаметы путем мейоза. У животных споры (т.е. гаплоидные клетки, способные подвергаться митозу) не образуются, поэтому бесполое многоклеточное поколение отсутствует. У некоторых насекомых есть гаплоидные самцы, развивающиеся из неоплодотворенных яиц, но все самки диплоидны. [ 17 ]
Вариации
[ редактировать ]Диаграмма, показанная выше, хорошо отображает жизненный цикл некоторых многоклеточных водорослей (например, рода Cladophora ), у которых спорофиты и гаметофиты почти одинакового внешнего вида и которые не имеют разных видов спор или гамет. [ 18 ]
Однако существует множество возможных вариаций фундаментальных элементов жизненного цикла, в котором происходит чередование поколений. Каждая вариация может возникать отдельно или в комбинации, что приводит к ошеломляющему разнообразию жизненных циклов. Термины, используемые ботаниками при описании этих жизненных циклов, могут сбивать с толку. Как говорят Бейтман и Димишель, «[...] чередование поколений превратилось в терминологическую трясину; часто один термин представляет несколько понятий или одно понятие представлено несколькими терминами». [ 19 ]
Возможные варианты:
- Относительное значение спорофита и гаметофита.
- Равные ( гомоморфия или изоморфность ).
Нитчатые водоросли рода Cladophora , встречающиеся преимущественно в пресных водах, имеют диплоидные спорофиты и гаплоидные гаметофиты, внешне неразличимые. [ 20 ] Ни одно живое наземное растение не имеет одинаково доминирующих спорофитов и гаметофитов, хотя некоторые теории эволюции чередования поколений предполагают, что у предковых наземных растений они были. - Неравные ( гетероморфность или анизоморфия ).

Гаметофит Mnium Hornum , мох. - Доминантный гаметофит ( гаметофит ).
У печеночников, мхов и роголистников доминирующей формой является гаплоидный гаметофит. Диплоидный спорофит не способен к самостоятельному существованию, получая большую часть питания от родительского гаметофита и не имея хлорофилла при созревании. [ 21 ]
Спорофит Ломарии обесцвечивает , папоротник - Доминирующий спорофит ( спорофит ).
У папоротников как спорофит, так и гаметофит способны жить самостоятельно, но доминирующей формой является диплоидный спорофит. Гаплоидный гаметофит значительно меньше и проще по строению. У семенных растений гаметофит еще более редуцирован (минимум всего до трех клеток), получая все питание от спорофита. Чрезвычайное уменьшение размеров гаметофита и сохранение его внутри спорофита означает, что применительно к семенным растениям термин «чередование поколений» несколько вводит в заблуждение: «[s]порофит и гаметофит эффективно функционируют как единый организм». [ 8 ] Некоторые авторы предпочитают термин «чередование фаз». [ 9 ]
- Доминантный гаметофит ( гаметофит ).
- Равные ( гомоморфия или изоморфность ).
- Дифференциация гамет.
- Обе гаметы одинаковы ( изогамия ).
Как и другие виды Cladophora , C. callicoma имеет жгутиковые гаметы, идентичные по внешнему виду и способности двигаться. [ 20 ] - Гаметы двух разных размеров ( анизогамия ).
- Оба имеют схожую подвижность .
Виды Ulva , морского салата, имеют гаметы, все из которых имеют два жгутика и поэтому подвижны. Однако они бывают двух размеров: более крупные «женские» гаметы и меньшие «мужские» гаметы. [ 22 ] - Один большой и сидячий, другой маленький и подвижный ( оогаминый ). Более крупные сидячие мегагаметы представляют собой яйцеклетки (яйцеклетки), а более мелкие подвижные микрогаметы — это сперматозоиды (сперматозоиды, сперматозоиды). Степень подвижности сперматозоидов может быть весьма ограниченной (как в случае с цветковыми растениями), но все они способны двигаться к сидячим яйцеклеткам. Когда (что почти всегда бывает) сперма и яйцеклетки производятся в разных видах гаметангиев, те, которые производят сперму, называются антеридиями (единственный антеридиум), а продуцирующие яйцеклетки - архегониями (единственный архегоний).

Гаметофит Pellia epiphylla со спорофитами, растущими из остатков архегониев. - Антеридии и архегонии встречаются на одном гаметофите, который тогда называют однодомным . (Многие источники, в том числе посвященные мохообразным, используют термин «однодомный» для этой ситуации и «раздельнополый» — для противоположного. [ 23 ] [ 24 ] Здесь слова «однодомный» и «раздельнополый» используются только для спорофитов.)
У печеночницы Pellia epiphylla гаметофит является доминирующим поколением. Оно одноцветное: маленькие красноватые антеридии, производящие сперму, разбросаны вдоль средней жилки, а архегонии, производящие яйца, растут ближе к кончикам частей растения. [ 25 ] - Антеридии и архегонии встречаются на разных гаметофитах, которые тогда называются двудомными .
У мха Mnium Hornum гаметофит является доминирующим поколением. Оно двудомное: мужские растения образуют только антеридии в концевых розетках, женские растения - только архегонии в виде черешковых коробочек. [ 26 ] Гаметофиты семенных растений также двудомные. Однако родительский спорофит может быть однодомным, дающим как мужские, так и женские гаметофиты, или двудомным, дающим гаметофиты только одного пола. Гаметофиты семенных растений чрезвычайно уменьшены в размерах; архегоний состоит лишь из небольшого числа клеток, а весь мужской гаметофит может быть представлен всего двумя клетками. [ 27 ]
- Антеридии и архегонии встречаются на одном гаметофите, который тогда называют однодомным . (Многие источники, в том числе посвященные мохообразным, используют термин «однодомный» для этой ситуации и «раздельнополый» — для противоположного. [ 23 ] [ 24 ] Здесь слова «однодомный» и «раздельнополый» используются только для спорофитов.)
- Оба имеют схожую подвижность .
- Обе гаметы одинаковы ( изогамия ).
- Дифференциация спор.
- Все споры одинакового размера ( гомоспория или изоспория).
Хвощи (виды Equisetum ) имеют споры одинакового размера. [ 28 ] - Споры двух разных размеров ( гетероспория и анизоспория): более крупные мегаспоры и более мелкие микроспоры . Когда два вида спор образуются в разных видах спорангиев, их называют мегаспорангиями и микроспорангиями . Мегаспора часто (но не всегда) развивается за счет трех других клеток, образующихся в результате мейоза, который прерывается.
- Мегаспорангии и микроспорангии возникают на одном и том же спорофите, который тогда называют однодомным .
Большинство цветущих растений попадают в эту категорию. Таким образом, цветок лилии содержит шесть тычинок (микроспорангии), которые производят микроспоры, которые развиваются в пыльцевые зерна (микрогаметофиты), и три сросшихся плодолистика, которые образуют покровные мегаспорангии (семяпочки), каждая из которых образует мегаспору, которая развивается внутри мегаспорангия и дает образование мегагаметофит. У других растений, например у лещины, одни цветки имеют только тычинки, другие — только плодолистики, но одно и то же растение (т. е. спорофит) имеет оба вида цветков и поэтому является однодомным.
Цветы падуба европейского, раздельнополого вида: вверху мужские, внизу женские (листья срезаны, чтобы цветы были более четко видны) - Мегаспорангии и микроспорангии встречаются на разных спорофитах, которые тогда называются раздельнополыми .
Отдельное дерево падуба европейского ( Ilex aquifolium ) дает либо «мужские» цветки, которые имеют только функциональные тычинки (микроспорангии), образующие микроспоры, которые развиваются в пыльцевые зерна (микрогаметофиты), либо «женские» цветки, которые имеют только функциональные плодолистики, образующие покровные мегаспорангии (семязачатки). ), содержащие мегаспору, которая развивается в многоклеточный мегагаметофит.
- Мегаспорангии и микроспорангии возникают на одном и том же спорофите, который тогда называют однодомным .
- Все споры одинакового размера ( гомоспория или изоспория).




Между этими вариациями есть некоторые корреляции, но это всего лишь корреляции, а не абсолютные. Например, у цветковых растений микроспоры в конечном итоге производят микрогаметы (сперматозоиды), а мегаспоры в конечном итоге производят мегагаметы (яйца). Однако у папоротников и их родственных групп имеются группы с недифференцированными спорами, но с дифференцированными гаметофитами. Например, у папоротника Ceratopteris thalictrioides есть споры только одного вида, размеры которых постоянно различаются. Меньшие споры имеют тенденцию прорастать в гаметофиты, которые производят только антеридии, продуцирующие сперму. [ 28 ]
Сложный жизненный цикл
[ редактировать ]
Жизненный цикл растений может быть сложным. Смена поколений может происходить у растений, которые одновременно гетероморфны, спорофитны, оогаметны, двудомны, гетероспоричны и двудомны, например, у ивы ( поскольку большинство видов рода Salix раздельнополы). [ 29 ] Задействованные процессы:
- Неподвижная яйцеклетка, содержащаяся в архегонии, сливается с подвижным спермием, выделяющимся из антеридия. В результате образуется зигота мужского или женского пола.
- Мужская зигота путем митоза развивается в микроспорофит, который при зрелости производит один или несколько микроспорангиев. Микроспоры развиваются внутри микроспорангия путем мейоза.
У ивы (как и у всех семенных растений) зигота сначала развивается в зародыш микроспорофита внутри семяпочки (мегаспорангия, заключенного в один или несколько защитных слоев ткани, известных как покровы). По мере созревания эти структуры становятся семенем . Позже семя проливается, прорастает и вырастает в зрелое дерево. Мужская ива (микроспорофит) дает цветки только с тычинками, пыльниками которых являются микроспорангии. - Микроспоры прорастают, образуя микрогаметофиты; в зрелом возрасте образуется один или несколько антеридиев. Сперматозоиды развиваются внутри антеридиев.
У ивы микроспоры не высвобождаются из пыльника (микроспорангия), а развиваются внутри него в пыльцевые зерна (микрогаметофиты). Все пыльцевое зерно перемещается (например, насекомым или ветром) в семязачаток (мегагаметофит), где вырабатывается сперматозоид, который перемещается по пыльцевой трубке и достигает яйцеклетки. - Женская зигота путем митоза развивается в мегаспорофит, который при зрелости производит один или несколько мегаспорангиев. Мегаспоры развиваются внутри мегаспорангия; обычно одна из четырех спор, образующихся в результате мейоза, набирает массу за счет остальных трех, которые исчезают.
Женские деревья ивы (мегаспорофиты) дают цветы только с плодолистиками (модифицированными листьями, несущими мегаспорангии). - Мегаспоры прорастают, образуя мегагаметофиты; в зрелом возрасте образуется один или несколько архегониев. Яйца развиваются внутри архегониев.
Из плодолистиков ивы образуются семязачатки — мегаспорангии, заключенные в покровы. Внутри каждой семяпочки мегаспора путем митоза развивается в мегагаметофит. Внутри мегагаметофита развивается архегоний и производит яйцеклетку. Все гаметофитное поколение остается под защитой спорофита, за исключением пыльцевых зерен (которые сократились до трех клеток, содержащихся в стенке микроспор).
- Мужская зигота путем митоза развивается в микроспорофит, который при зрелости производит один или несколько микроспорангиев. Микроспоры развиваются внутри микроспорангия путем мейоза.
Жизненные циклы разных групп растений
[ редактировать ]Термин «растения» здесь используется для обозначения Archaeplastida , т.е. глаукофитов , красных и зеленых водорослей и наземных растений .
Смена поколений происходит почти у всех многоклеточных красных и зеленых водорослей, как пресноводных форм (например, Cladophora ), так и морских водорослей (например, Ulva ). У большинства поколения гомоморфны (изоморфны) и свободноживущие. У некоторых видов красных водорослей наблюдается сложная трехфазная смена поколений, при которой имеется фаза гаметофита и две четко выраженные фазы спорофита. Дополнительную информацию см. в разделе Красные водоросли: Размножение .
У всех наземных растений наблюдается гетероморфное (анизоморфное) чередование поколений, при котором спорофит и гаметофит резко различаются. У всех мохообразных , то есть печеночников , мхов и роголистников , поколение гаметофитов является наиболее заметным. В качестве иллюстрации рассмотрим одноцветный мох. На зрелом растении (гаметофите) развиваются антеридии и архегонии. В присутствии воды двужгутиковые спермии из антеридиев плывут к архегониям и происходит оплодотворение, приводящее к образованию диплоидного спорофита. Спорофит вырастает из архегония. Его тело представляет собой длинный стебель, увенчанный капсулой, внутри которой клетки, производящие споры, подвергаются мейозу с образованием гаплоидных спор. Большинство мхов рассеивают эти споры ветром, хотя Splachnum sphaericum энтомофилен и привлекает насекомых для распространения своих спор.
-
 Смена поколений у печеночников.
Смена поколений у печеночников. -
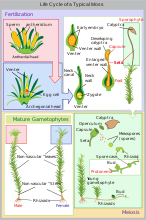 Жизненный цикл мха
Жизненный цикл мха -
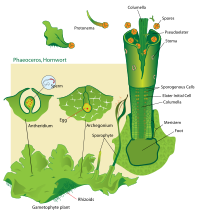 Жизненный цикл роголистника
Жизненный цикл роголистника



В жизненном цикле папоротников и их союзников, включая плауны и хвощи , наиболее заметным растением, наблюдаемым в полевых условиях, является диплоидный спорофит. Гаплоидные споры развиваются в сорусах на нижней стороне листьев и разносятся ветром (или, в некоторых случаях, плаванием по воде). При подходящих условиях спора прорастает и превращается в довольно незаметное тело растения, называемое проталлусом . Гаплоидный проталлус не похож на спорофит, поэтому папоротники и их союзники имеют гетероморфное чередование поколений. Заросток недолговечен, но осуществляет половое размножение, образуя диплоидную зиготу , которая затем вырастает из проталлома в виде спорофита.
-
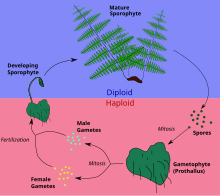 Смена поколений у папоротников
Смена поколений у папоротников -
 Гаметофит (проталлус) Диксонии .
Гаметофит (проталлус) Диксонии . -
 Спорофит Dicksonia antarctica.
Спорофит Dicksonia antarctica. -
 Вайя Dicksonia antarctica со спорообразующими структурами
Вайя Dicksonia antarctica со спорообразующими структурами




У сперматофитов , семенных растений, спорофит является доминирующей многоклеточной фазой; гаметофиты сильно уменьшены в размерах и весьма разнообразны по морфологии. Все поколение гаметофитов, за исключением пыльцевых зерен (микрогаметофитов), содержится внутри спорофита. Жизненный цикл двудомного цветкового растения (покрытосеменного), ивы, был подробно описан в предыдущем разделе ( «Сложный жизненный цикл »). Жизненный цикл голосеменных растений аналогичен. Однако у цветковых растений существует явление, называемое « двойное оплодотворение ». В процессе двойного оплодотворения в архегоний мегагаметофита попадают не один спермий, а два ядра спермия из пыльцевого зерна (микрогаметофита); одно сливается с ядром яйцеклетки, образуя зиготу, другое сливается с двумя другими ядрами гаметофита, образуя « эндосперм », который питает развивающийся эмбрион.
Эволюция доминирующей диплоидной фазы
[ редактировать ]Было высказано предположение, что основой появления диплоидной фазы жизненного цикла (спорофита) в качестве доминирующей фазы (например, как у сосудистых растений) является то, что диплоидия позволяет маскировать проявление вредных мутаций посредством генетической комплементации . [ 30 ] [ 31 ] Таким образом, если один из родительских геномов в диплоидных клетках содержал мутации, приводящие к дефектам одного или нескольких генных продуктов , эти недостатки могли быть компенсированы другим родительским геномом (который, тем не менее, может иметь свои собственные дефекты в других генах). Поскольку диплоидная фаза становилась преобладающей, эффект маскировки, вероятно, позволил увеличить размер генома и, следовательно, информационное содержание без необходимости повышения точности репликации ДНК. Возможность увеличить информационное содержание при низких затратах была выгодной, поскольку позволяла кодировать новые адаптации. Эта точка зрения была оспорена: данные показывают, что отбор не более эффективен на гаплоидных, чем на диплоидных фазах жизненного цикла мхов и покрытосеменных. [ 32 ]
-
 Жизненный цикл покрытосеменных
Жизненный цикл покрытосеменных -
 На кончике тычинки тюльпана видна пыльца (микрогаметофиты).
На кончике тычинки тюльпана видна пыльца (микрогаметофиты). -
 Семяпочки растений (мегагаметофиты): семяпочка голосеменных слева, семяпочка покрытосеменных (внутри завязи) справа.
Семяпочки растений (мегагаметофиты): семяпочка голосеменных слева, семяпочка покрытосеменных (внутри завязи) справа. -
 Двойное оплодотворение
Двойное оплодотворение




Подобные процессы у других организмов
[ редактировать ]Ризария
[ редактировать ]
Некоторые организмы, которые в настоящее время отнесены к кладе Rhizaria и, следовательно, не являются растениями в том смысле, который здесь используется, демонстрируют чередование поколений. У большинства фораминифер происходит гетероморфное чередование поколений между гаплоидными формами гамонта и диплоидными формами агамонта . Диплоидная форма обычно намного крупнее гаплоидной; эти формы известны как микросфера и мегасфера соответственно.
Грибы
[ редактировать ]Грибной мицелий обычно гаплоидный. Когда мицелии разных типов спаривания встречаются, они образуют две многоядерные шарообразные клетки, которые соединяются посредством «мостика спаривания». Ядра перемещаются из одного мицелия в другой, образуя гетерокарион (что означает «разные ядра»). Этот процесс называется плазмогамией . Фактическое слияние с образованием диплоидных ядер называется кариогамией и может не произойти до тех пор, пока спорангии не сформируются . Карогамия дает диплоидную зиготу, которая представляет собой недолговечный спорофит, который вскоре подвергается мейозу с образованием гаплоидных спор. Когда споры прорастают, они развиваются в новый мицелий.
Слизевики
[ редактировать ]Жизненный цикл слизевиков очень похож на жизненный цикл грибов. Гаплоидные споры прорастают с образованием роевых клеток или миксамеб . Они сливаются в процессе, называемом плазмогамией и кариогамией, с образованием диплоидной зиготы. Зигота развивается в плазмодий, а зрелый плазмодий дает, в зависимости от вида, от одного до многих плодовых тел, содержащих гаплоидные споры.
Животные
[ редактировать ]Чередование многоклеточного диплоидного и многоклеточного гаплоидного поколений у животных никогда не встречается. [ 33 ] У некоторых животных наблюдается чередование партеногенной и половой репродуктивной фаз ( гетерогамия ), например у сальп и долиолид (класс Thaliacea ). Обе фазы диплоидны. Иногда это называют «чередованием поколений». [ 34 ] но совсем другое. У некоторых других животных, таких как перепончатокрылые , самцы гаплоидны, а самки диплоидны, но это всегда так, а не происходит чередование отдельных поколений.
См. также
[ редактировать ]- Эволюционная история растений#жизненные циклы : Эволюционное происхождение чередования фаз
- Плоидия – количество наборов хромосом клетки.
- Апомиксис - замена нормального полового размножения бесполым без оплодотворения.
Примечания и ссылки
[ редактировать ]- ^ «Смена поколений | Определение и примеры» . Британская энциклопедия . Архивировано из оригинала 04 марта 2021 г. Проверено 25 февраля 2021 г.
- ^ Томас, Р.Дж.; Стэнтон, Д.С.; Лонгендорфер, Д.Х. и Фарр, МЭ (1978), «Физиологическая оценка пищевой автономии спорофита роголистника», Botanical Gazette , 139 (3): 306–311, doi : 10.1086/337006 , S2CID 84413961
- ^ Глайм, Дж. М. (2007), Экология мохообразных: Том. 1 Физиологическая экология (PDF) , Мичиганский технологический университет и Международная ассоциация бриологов, заархивировано (PDF) из оригинала 26 марта 2013 г. , получено 4 марта 2013 г.
- ^ Jump up to: а б Керп, Х.; Тревин, Н. Х. и Хасс, Х. (2003), «Новые гаметофиты из нижнего девона Райни Черт», Труды Королевского общества Эдинбурга: науки о Земле , 94 (4): 411–428, doi : 10.1017/S026359330000078X , S2CID 128629425
- ^ Клюге, Арнольд Г.; Штраус, Ричард Э. (1985). «Онтогенез и систематика» . Ежегодный обзор экологии и систематики . 16 : 247–268. doi : 10.1146/annurev.es.16.110185.001335 . ISSN 0066-4162 . JSTOR 2097049 . Архивировано из оригинала 27 января 2022 г. Проверено 25 февраля 2021 г.
- ^ Тейлор, Керп и Хасс, 2005 г.
- ^ " «Наука о растениях 4 У» . Архивировано из оригинала 18 августа 2016 года . Проверено 5 июля 2016 г.
- ^ Jump up to: а б Бейтман и Димишель 1994 , с. 403
- ^ Jump up to: а б Стюарт и Ротвелл, 1993 г.
- ^ «Объект месяца: Поэт и череп дельфина» . Центр культурной техники Германа фон Гельмгольца. 1 июля 2023 г. Проверено 4 мая 2024 г.
- ^ Jump up to: а б с д Хейг, Дэвид (2008), «Гомологическое и антитетическое чередование поколений и происхождение спорофитов» (PDF) , The Botanical Review , 74 (3): 395–418, doi : 10.1007/s12229-008-9012-x , S2CID 207403936 , заархивировано (PDF) из оригинала 19 августа 2014 г. , получено 17 августа 2014 г.
- ^ Сведелиус, Нильс (1927), «Смена поколений в связи с редукционным разделением», Botanical Gazette , 83 (4): 362–384, doi : 10.1086/333745 , JSTOR 2470766 , S2CID 84406292
- ^ Хофмайстер, В. (1851), Сравнительные исследования прорастания, развития и формирования плодов высших криптогамов (мхов, папоротников, хвощеобразных, корнеплодов и плаунов) и семенообразования хвойных пород (на немецком языке), Лейпциг: Ф. Хофмейстер , получено 17 августа 2014 г. Переведено как Карри, Фредерик (1862), О прорастании, развитии и плодоношении высших криптогамий и о плодоношении хвойных , Лондон: Роберт Хардвик, заархивировано из оригинала 19 августа 2014 г. , получено 17 августа 2014 г.
- ^ Фельдманн, Дж .; Фельдманн, Г. (1942), « Recherches sur les Bonnemaisoniacées et leur чередования поколений» [ Исследования Bonnemaisoniaceae и их чередования поколений] (PDF) , Ann. наук. Натл. Бот. , Серия 11 (на французском языке), 3 : 75–175, заархивировано из оригинала (PDF) 19 августа 2014 г. , получено 07 октября 2013 г. , стр. 157
- ^ Jump up to: а б Фельдманн, Дж. (1972), «Современные проблемы смены поколений у водорослей», Bulletin de la Société Botanique de France (на французском языке), 119 : 7–38, doi : 10.1080/00378941.1972.10839073
- ^ Шопфер, П.; Мор, Х. (1995). «Физиология развития» . Физиология растений . Берлин: Шпрингер. стр. 288–291 . ISBN 978-3-540-58016-4 .
- ^ Jump up to: а б с д и ж г час Фостер и Гиффорд 1974 , [ нужна страница ] Спорный 1974а [ нужна страница ] и Спорн, 1974b . [ нужна страница ]
- ^ Гири и Гири 2008
- ^ Бэйтман и Димишель 1994 , с. 347
- ^ Jump up to: а б Шьям 1980 г.
- ^ Уотсон 1981 , с. 2
- ^ Кирби 2001
- ^ Уотсон 1981 , с. 33
- ^ Белл и Хемсли 2000 , с. 104
- ^ Уотсон 1981 , стр. 425–6.
- ^ Уотсон 1981 , стр. 287–8.
- ^ Спорне 1974a , стр. 17–21.
- ^ Jump up to: а б Бейтман и Димишель 1994 , стр. 350–1.
- ^ Чисхолм, Хью , изд. (1911). . Британская энциклопедия . Том. 28 (11-е изд.). Издательство Кембриджского университета. стр. 688–689.
- ^ Бернштейн, Х.; Байерс, Г.С. и Мишод, Р.Э. (1981), «Эволюция полового размножения: важность восстановления, комплементации и вариаций ДНК», The American Naturalist , 117 (4): 537–549, doi : 10.1086/283734 , S2CID 84568130
- ^ Мишод, Р.Э. и Гейли, Т.В. (1992), «Маскировка мутаций и эволюция пола», The American Naturalist , 139 (4): 706–734, doi : 10.1086/285354 , S2CID 85407883
- ^ Шовеньи, Питер; Рикка, Мариана; Хок, Жофия; Шоу, Джонатан А.; Симидзу, Кентаро К.; Вагнер, Андреас (2013), «Отбор на гаплоидных стадиях жизни покрытосеменных растений и мхов не более эффективен, чем на диплоидных», Molecular Biology and Evolution , 30 (8): 1929–39, doi : 10.1093/molbev/mst095 , ПМИД 23686659
- ^ Барнс и др. 2001 , с. 321
- ^ Скотт 1996 , с. 35
Библиография
[ редактировать ]- Барнс, РСК; Калоу, П.; Олив, PJW; Голдинг, Д.В. и Спайсер, Дж.И. (2001), Беспозвоночные: синтез , Оксфорд; Молден, Массачусетс: Блэквелл, ISBN 978-0-632-04761-1
- Бейтман, Р.М. и Димишель, Вашингтон (1994), «Гетероспория – наиболее повторяющаяся ключевая инновация в эволюционной истории царства растений» (PDF) , Biological Reviews of the Cambridge Philosophical Society , 69 (3): 345–417, doi : 10.1111/j.1469-185x.1994.tb01276.x , S2CID 29709953 , заархивировано из оригинала (PDF) 15 апреля 2012 г. , получено 30 декабря 2010 г.
- Белл, PR и Хемсли, AR (2000), Зеленые растения: их происхождение и разнообразие (2-е изд.), Кембридж и др.: Cambridge University Press, ISBN. 978-0-521-64109-8
- Фостер, А.С. и Гиффорд, Э.М. (1974), Сравнительная морфология сосудистых растений (2-е изд.), Сан-Франциско: WH Freeman, ISBN 978-0-7167-0712-7
- Гири, доктор медицины; Гири, GM (2008), «Cladophora» , AlgaeBase , всемирное электронное издание, Национальный университет Ирландии, Голуэй , получено 21 июля 2011 г.
- Кирби, А. (2001), Ульва, морской салат , Научно-исследовательский институт аквариумов Монтерей-Бей, заархивировано из оригинала 16 мая 2011 г. , извлечено 1 января 2011 г.
- Скотт, Томас (1996), Краткая энциклопедия биологии , Берлин: Вальтер де Грюйтер, ISBN 978-3-11-010661-9
- Шьям, Р. (1980), «О жизненном цикле, цитологии и таксономии Cladophora callicoma из Индии», American Journal of Botany , 67 (5): 619–24, doi : 10.2307/2442655 , JSTOR 2442655
- Спорн, КР (1974a), Морфология покрытосеменных , Лондон: Хатчинсон, ISBN 978-0-09-120611-6
- Спорн, КР (1974b), Морфология голосеменных растений (2-е изд.), Лондон: Хатчинсон, ISBN 978-0-09-077152-3
- Стюарт, В.Н. и Ротвелл, Г.В. (1993), Палеоботаника и эволюция растений (2-е изд.), Кембридж, Великобритания: Издательство Кембриджского университета, ISBN 978-0-521-38294-6
- Уотсон, EV (1981), Британские мхи и печеночники (3-е изд.), Кембридж, Великобритания: Cambridge University Press, ISBN 978-0-521-28536-0
- Тейлор, Теннесси; Керп, Х. и Хасс, Х. (2005), «Биология истории жизни ранних наземных растений: расшифровка фазы гаметофита», Труды Национальной академии наук Соединенных Штатов Америки , 102 (16): 5892–5897. , Bibcode : 2005PNAS..102.5892T , doi : 10.1073/pnas.0501985102 , PMC 556298 , PMID 15809414
| Субдисциплины | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| растений Группы | |||||||||||
| Анатомия растений |
| ||||||||||
| Физиология растений Материалы | |||||||||||
| Рост растений и привычка | |||||||||||
| Воспроизведение | |||||||||||
| Таксономия растений | |||||||||||
| Упражняться | |||||||||||
| |||||||||||
| Базы данных органов управления : Национальные |
|---|