Оогоний
| Оогоний | |
|---|---|
| Идентификаторы | |
| МеШ | D009867 |
| ФМА | 83673 |
| Анатомическая терминология | |
Оогоний ) ( мн.: оогония ) — небольшая диплоидная клетка, которая при созревании образует примордиальный фолликул у плода женского пола или женский (гаплоидный или диплоидный гаметангий некоторых таллофитов .
У плода млекопитающих
[ редактировать ]Оогонии образуются в большом количестве в результате митоза на ранних стадиях развития плода из примордиальных половых клеток . У человека они начинают развиваться между 4 и 8 неделями и присутствуют у плода между 5 и 30 неделями.
Структура
[ редактировать ]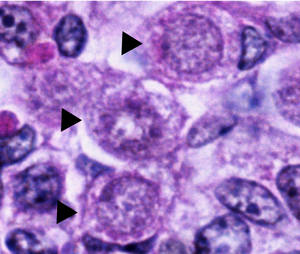
Нормальные оогонии в яичниках человека имеют сферическую или яйцевидную форму и обнаруживаются среди соседних соматических клеток и ооцитов на разных стадиях развития. Оогонии можно отличить от соседних соматических клеток под электронным микроскопом , наблюдая за их ядрами . Оогониальные ядра содержат беспорядочно рассеянный фибриллярный и зернистый материал, тогда как соматические клетки имеют более конденсированное ядро, которое под микроскопом создает более темный контур. Оогониальные ядра также содержат плотные выступающие ядрышки . Хромосомный везикулами материал в ядре митотически делящихся оогониев представляет собой плотную массу, окруженную или двойными мембранами. [1]
Цитоплазма митохондрии оогониев похожа на цитоплазму окружающих соматических клеток и также содержит большие круглые с латеральными кристами . Однако эндоплазматическая сеть (ЭР) оогониев очень слабо развита и состоит из нескольких мелких пузырьков. Некоторые из этих мелких пузырьков содержат цистерны с рибосомами и располагаются вблизи аппарата Гольджи . [1]
Оогонии, претерпевающие дегенерацию, под электронным микроскопом выглядят несколько иначе. В этих оогониях хромосомы слипаются в неразличимую массу внутри ядра, а митохондрии и ЭР кажутся набухшими и разрушенными. Дегенерирующие оогонии обычно обнаруживаются частично или полностью поглощенными соседними соматическими клетками, что определяет фагоцитоз как способ элиминации. [1]
Развитие и дифференциация
[ редактировать ]В бластоцисте млекопитающих эмбриона примордиальные половые клетки возникают из проксимальных эпибластов под влиянием внеэмбриональных сигналов. Эти зародышевые клетки затем посредством амебоидного движения перемещаются к генитальному гребню и, в конечном итоге, в недифференцированные гонады плода. [2] На 4-5-й неделе развития гонады начинают дифференцироваться. При отсутствии Y-хромосомы гонады дифференцируются в яичники. По мере дифференцировки яичников развиваются врастания, называемые кортикальными тяжами. Здесь собираются первичные половые клетки. [3] [4]
На 6–8-й неделе женского (XX) эмбрионального развития примордиальные зародышевые клетки растут и начинают дифференцироваться в оогонии. Оогонии размножаются путем митоза на 9-22 неделе эмбрионального развития. Оогониев может быть до 600 000 к 8-й неделе развития и до 7 000 000 к 5-му месяцу. [3]
В конце концов, оогонии либо дегенерируют, либо далее дифференцируются в первичные ооциты посредством асимметричного деления. Асимметричное деление — это процесс митоза, при котором один оогоний делится неравномерно с образованием одной дочерней клетки, которая в конечном итоге станет ооцитом в процессе оогенеза , и одной дочерней клетки, которая является идентичным оогонию родительской клетке. Это происходит в период с 15-й недели по 7-й месяц эмбрионального развития. [2] Большинство оогониев к моменту рождения либо дегенерировали, либо дифференцировались в первичные ооциты. [3] [5]
Первичные ооциты подвергаются оогенезу, в ходе которого они вступают в мейоз . Однако первичные ооциты задерживаются в профазе 1 первого мейоза и остаются на этой стадии до тех пор, пока у взрослой женщины не начнется половое созревание. [6] В этом отличие от мужских первичных зародышевых клеток, которые задерживаются на сперматогониальной стадии при рождении и не вступают в сперматогенез и мейоз для производства первичных сперматоцитов до полового созревания у взрослого мужчины. [3]
Регуляция дифференцировки оогониев и вступление в оогенез
[ редактировать ]Регуляция и дифференцировка половых клеток в первичные гаметоциты в конечном итоге зависят от пола эмбриона и дифференцировки гонад. У самок мышей белок RSPO1 отвечает за дифференцировку женских (XX) гонад в яичники . RSPO1 активирует сигнальный путь β-катенина путем активации Wnt4 , что является важным этапом дифференцировки яичников. Исследования показали, что в яичниках, лишенных Rspo1 или Wnt4, происходит смена пола гонад, образование яйцеклеток и дифференцировка соматических клеток Сертоли , которые способствуют развитию сперматозоидов . [4]
После того, как женские (XX) зародышевые клетки собираются в недифференцированных гонадах, необходима активация Stra8 для дифференцировки зародышевых клеток в оогоний и, в конечном итоге, для вступления в мейоз. Одним из основных факторов, который способствует активации Stra8 , является инициация сигнального пути β-Catenin через RSPO1, который также отвечает за дифференцировку яичников. Поскольку RSPO1 вырабатывается в соматических клетках, этот белок действует на половые клетки паракринным способом . Rspo1 , однако, не является единственным фактором регуляции Stra8 . Многие другие факторы находятся под пристальным вниманием, и этот процесс все еще оценивается. [4]
Оогониальные стволовые клетки
[ редактировать ]Предполагается, что огонии либо дегенерируют, либо дифференцируются в первичные ооциты , которые вступают в оогенез и останавливаются в профазе I первого мейоза послеродового . Поэтому считается, что у взрослых самок млекопитающих отсутствует популяция зародышевых клеток, которые могут обновляться или регенерировать, и вместо этого имеется большая популяция первичных ооцитов, которые задерживаются в первом мейозе до полового созревания . [2] В период полового созревания один первичный ооцит продолжает мейоз каждый менструальный цикл . Поскольку у человека отсутствуют регенерирующие зародышевые клетки и оогонии, количество первичных ооцитов уменьшается после каждого менструального цикла до менопаузы , когда у женщины больше нет популяции первичных ооцитов. [2]
Однако недавние исследования показали, что возобновляемые оогонии могут присутствовать в слизистой оболочке женских яичников человека, приматов и мышей. [2] [7] [8] Считается, что эти зародышевые клетки могут быть необходимы для поддержания репродуктивных фолликулов и развития ооцитов даже во взрослом возрасте. Также было обнаружено, что некоторые стволовые клетки могут мигрировать из костного мозга в яичники в качестве источника экстрагениальных половых клеток. Эти митотически активные зародышевые клетки, обнаруженные у взрослых млекопитающих, были идентифицированы путем отслеживания нескольких маркеров, которые были распространены в ооцитах. Эти потенциальные возобновляемые зародышевые клетки были идентифицированы как положительные по этим важным маркерам ооцитов. [2]
Открытие этих активных половых клеток и оогоний у взрослых женщин может быть очень полезным для развития исследований фертильности и лечения бесплодия. [2] [8] Зародышевые клетки были извлечены, изолированы и успешно выращены in vitro . [8] Эти зародышевые клетки использовались для восстановления фертильности у мышей, способствуя образованию и поддержанию фолликулов у ранее бесплодных мышей. Также проводятся исследования возможной регенерации зародышевой линии у приматов. Митотически активные женские зародышевые клетки человека могут оказаться очень полезными для нового метода развития эмбриональных стволовых клеток, который включает перенос ядра в зиготу . Использование этих функциональных оогоний может помочь создать с помощью этого метода линии стволовых клеток, специфичные для пациента. [2]
Споры
[ редактировать ]Существуют серьезные разногласия относительно существования оогониальных стволовых клеток млекопитающих. Споры заключаются в отрицательных данных, полученных из многих лабораторий США. Множественные подходы к проверке существования оогониальных стволовых клеток дали отрицательные результаты, и ни одна исследовательская группа в США не смогла воспроизвести первоначальные результаты. [9] [10] [11]
У некоторых таллофитов
[ редактировать ]
В психологии и микологии если оогонием называют женский гаметангий, слияние мужской (подвижной или неподвижной) и женской гаметы . внутри этой структуры происходит [12] [13]
У Oomycota и некоторых других организмов женские оогонии и мужские эквивалентные антеридии являются результатом полового споруляции, т.е. развития структур, внутри которых происходит мейоз. Гаплоидные ядра (гаметы) образуются в результате мейоза внутри антеридиев и оогоний, а когда происходит оплодотворение, диплоидная ооспора , которая в конечном итоге прорастает в диплоидную соматическую стадию жизненного цикла таллофита. образуется [14]
У многих водорослей (например, Chara ) основное растение гаплоидное; оогонии и антеридии образуют и производят гаплоидные гаметы. Единственной диплоидной частью жизненного цикла является спора (оплодотворенная яйцеклетка), которая подвергается мейозу с образованием гаплоидных клеток, из которых развиваются новые растения. Это гаплонтический жизненный цикл (с зиготическим мейозом ).
Структура
[ редактировать ]Оогонии некоторых видов Thallophyte. [ который? ] обычно округлые или яйцевидные, содержимое которых разделено на несколько одноядерных оосфер . В отличие от мужских антеридиев, которые удлинены и содержат несколько ядер. [14]
У гетероталличных видов оогонии и антеридии располагаются на гифальных ветвях разных колоний таллофитов. Оогонии этого вида могут оплодотворяться только антеридиями из другой колонии и гарантируют, что самооплодотворение невозможно. [ нужны разъяснения ] Напротив, гомоталличные виды имеют оогонии и антеридии либо на одной и той же ветви гиф, либо на отдельных ветвях гиф, но в пределах одной колонии. [14]
Оплодотворение
[ редактировать ]При обычном способе оплодотворения, обнаруженном у некоторых видов Thallophytes, антеридии связываются с оогониями. Затем антеридии образуют трубки оплодотворения, соединяющие цитоплазму антеридиев с каждой оосферой внутри оогоний. Гаплоидное ядро (гамета) из антеридия затем переносится через трубку оплодотворения в оосферу и сливается с гаплоидным ядром оосферы, образуя диплоидную ооспору. Затем ооспора готова прорасти и развиться во взрослую диплоидную соматическую стадию. [14]
См. также
[ редактировать ]Список различных типов клеток в организме взрослого человека
Ссылки
[ редактировать ]- ^ Jump up to: а б с Бейкер, Т.Г.; Л.Л. Франки (1967). «Тонкая структура ооцитов огониев в яичниках человека» . Журнал клеточной науки . 2 (2): 213–224. дои : 10.1242/jcs.2.2.213 . ПМИД 4933750 . Архивировано из оригинала 4 марта 2016 года . Проверено 6 апреля 2012 г.
- ^ Jump up to: а б с д и ж г час «Герминные стволовые клетки. Научное резюме» . Медицинская школа Нью-Джерси. Архивировано из оригинала 12 января 2012 года . Проверено 6 апреля 2012 г.
- ^ Jump up to: а б с д Джонс, Ричард Э. (1997). Репродуктивная биология человека, 2-е изд . Сан-Диего: Academic Press, Elsevier. стр. 26–40, 90–107, 117–125. ISBN 0-12-389775-0 .
- ^ Jump up to: а б с Чассо, А.А.; Грегуар, EP; Лавери, Р.; Такето, ММ; де Рой, генеральный директор; и др. (2011). «Сигнальный путь RSPO1/β-катенин регулирует дифференцировку оогоний и вступление в мейоз в яичниках плода мыши» . ПЛОС ОДИН . 6 (10): e25641. Бибкод : 2011PLoSO...625641C . дои : 10.1371/journal.pone.0025641 . ПМК 3185015 . ПМИД 21991325 .
- ^ «Эмриология человека, эмбриогенез» . Модуль 3, Гаметогенез . Проверено 6 апреля 2012 г.
- ^ «Генетика, мейоз и гетогенез» . www.emich.edu. Архивировано из оригинала 30 апреля 2012 года . Проверено 6 апреля 2012 г.
- ^ Телфер, Эвелин Э .; Дэвид Ф. Альбертини (2012). «В поисках стволовых клеток яичников человека». Природная медицина . 18 (3): 353–354. дои : 10.1038/нм.2699 . ПМИД 22395699 . S2CID 1289213 .
- ^ Jump up to: а б с Уайт, Ивонн А.Р.; Дори С. Вудс; Яшуши Такай; Осаму Исихара; Хироюки Секи; Джонатан Л. Тилли (2012). «Формирование ооцитов митотически активными половыми клетками, выделенными из яичников женщин репродуктивного возраста» . Природная медицина . 18 (3): 413–421. дои : 10.1038/нм.2669 . ПМК 3296965 . ПМИД 22366948 .
- ^ Чжан, Х; Панула, С; Петропулос, С; Эдсгард, Д; Бусаяваласа, К; Лю, Л; Ли, Х; Рисал, С; Шен, Ю; Шао, Дж; Лю, М; Ли, С; Чжан, Д; Чжан, X; Гернер, Р.Р.; Шейхи, М; Дамдимопулу, П; Сандберг, Р; Дуаги, я; Густафссон, Йо; Лю, Л; Ланнер, Ф; Оватта, О; Лю, К. (октябрь 2015 г.). «В яичниках взрослого человека и мыши отсутствуют функциональные оогониальные стволовые клетки, экспрессирующие DDX4». Нат. Мед . 21 (10): 1116–8. дои : 10.1038/нм.3775 . hdl : 10616/44674 . ПМИД 26444631 . S2CID 2949653 .
- ^ Лей, Л; Спрэдлинг, AC (2013). «Самкам мышей не хватает стволовых клеток взрослой зародышевой линии, но они поддерживают оогенез, используя стабильные примордиальные фолликулы» . Proc Natl Acad Sci США . 110 (21): 8585–90. Бибкод : 2013PNAS..110.8585L . дои : 10.1073/pnas.1306189110 . ПМЦ 3666718 . ПМИД 23630252 .
- ^ Хоран, CJ; Уильямс, Ю.А. (2017). «Стволовые клетки ооцитов: факт или фантазия?» . Размножение . 154 (1): Р23–Р35. дои : 10.1530/REP-17-0008 . ПМИД 28389520 .
- ^ Стегенга, Х. Болтон, Дж. Дж. и Андерсон, Р. Дж. 1997. Морские водоросли западного побережья Южной Африки. Болюсный гербарий, Кейптаунский университет. ISBN 0-7992-1793-X
- ^ Смит, генеральный директор 1955. Криптогамная ботаника. том. 1. Книжная компания McGraw-Hill
- ^ Jump up to: а б с д «Половое спорообразование у Oomycota» . Архивировано из оригинала 12 апреля 2012 года . Проверено 6 апреля 2012 г.