Амеба

Амеба ( ( / ə ˈ m iː b ə / пишется амеба или amœba ; мн .: амебы (реже амебы ) или амебы amebae ) ; реже / ə ˈ m iː b i / ), [1] часто называемый амебоидом , представляет собой тип клетки или одноклеточного организма , способный изменять свою форму, в первую очередь путем вытягивания и втягивания ложноножек . [2] Амебы не образуют единой таксономической группы ; вместо этого они встречаются во всех основных линиях эукариотических организмов . Амебоидные клетки встречаются не только среди простейших , но и у грибов , водорослей и животных . [3] [4] [5] [6] [7]
Микробиологи часто используют термины «амебоид» и «амебоид» как синонимы для любого организма, демонстрирующего амебоидное движение . [8] [9]
В старых классификационных системах большинство амеб относили к классу или подтипу Sarcodina — группе одноклеточных организмов , которые обладают ложноножками или передвигаются за счет протоплазматического потока. Однако молекулярно-филогенетические исследования показали, что Sarcodina не является монофилетической группой, члены которой имеют общее происхождение . Следовательно, амебоидные организмы больше не относят к одной группе. [10]
Наиболее известными амебоидными протистами являются Chaos carolinense и Amoeba proteus , оба из которых широко культивируются и изучаются в классах и лабораториях. [11] [12] Другие хорошо известные виды включают так называемую «амёбу, пожирающую мозг» Naegleria fowleri , кишечный паразит Entamoeba histolytica , вызывающий амебную дизентерию , и многоклеточную «социальную амебу» или слизевую плесень Dictyostelium discoideum .
Форма, движение и питание
[ редактировать ]
У амеб нет клеточных стенок, что позволяет им свободно передвигаться. Амебы передвигаются и питаются с помощью ложноножек, которые представляют собой выпуклости цитоплазмы, образующиеся в результате скоординированного действия актиновых микрофиламентов, выталкивающих плазматическую мембрану , окружающую клетку. [13] По внешнему виду и внутреннему строению ложноножек можно отличить группы амеб друг от друга. Виды амебозойных животных , например представители рода Amoeba , обычно имеют луковицеобразные (лопастные) ложноножки, закругленные на концах и примерно трубчатые в поперечном сечении. У церкозойных амебоидов, таких как Euglypha и Gromia , есть тонкие, нитевидные (нитевидные) ложноножки. Фораминиферы выделяют мелкие ветвящиеся ложноножки, которые сливаются друг с другом, образуя сетчатые (сетчатые) структуры. Некоторые группы, такие как Radiolaria и Heliozoa , имеют жесткие, игольчатые, расходящиеся аксоподии (actinopoda), поддерживаемые изнутри пучками микротрубочек . [3] [14]


Свободноживущие амебы могут быть « раковинными » (заключенными в твердую оболочку) или «голыми» (также известными как гимнамебные , не имеющие твердого покрытия). Раковины раковинных амеб могут состоять из различных веществ, включая кальций , кремнезем , хитин , или из скоплений найденных материалов, таких как мелкие песчинки и панцири диатомовых водорослей . [15]
Для регулирования осмотического давления у большинства пресноводных амеб имеется сократительная вакуоль , которая выводит избыток воды из клетки. [16] Эта органелла необходима, потому что пресная вода имеет более низкую концентрацию растворенных веществ (например, соли), чем собственные внутренние жидкости амебы ( цитозоль ). Поскольку окружающая вода гипотонична по отношению к содержимому клетки, вода переносится через клеточную мембрану амебы путем осмоса . Без сократительной вакуоли клетка наполнилась бы избытком воды и в конечном итоге лопнула бы. Морские амебы обычно не имеют сократительной вакуоли, поскольку концентрация растворенных веществ внутри клетки находится в балансе с тоничностью окружающей воды. [17]
Диета
[ редактировать ]
Источники питания амеб различаются. Некоторые амебы являются хищниками и живут, поедая бактерии и других протистов . Некоторые из них являются детритофагами и питаются мертвым органическим материалом.
Амебы обычно поглощают пищу путем фагоцитоза , расширяя ложноножки, чтобы окружить и поглотить живую добычу или частицы собранного материала. Амебоидные клетки не имеют рта или цитостома , и на клетке нет фиксированного места, в котором обычно происходит фагоцитоз. [18]
Некоторые амебы также питаются путем пиноцитоза , поглощая растворенные питательные вещества через пузырьки, образующиеся внутри клеточной мембраны. [19]
Диапазон размеров
[ редактировать ]
Размер амебоидных клеток и видов чрезвычайно изменчив. Морской амебоид Massisteria voersi всего 2,3–3 микрометра . имеет диаметр [20] в пределах размера многих бактерий. [21] Напротив, раковины глубоководных ксенофиофоров могут достигать 20 см в диаметре. [22] Большинство свободноживущих пресноводных амеб, обычно встречающихся в прудовой воде , канавах и озерах, микроскопичны , но некоторые виды, такие как так называемые «гигантские амебы» Pelomyxa palustris и Chaos carolinense , могут быть достаточно большими, чтобы их можно было увидеть обнаженными. глаз.
| Вид или тип клетки | Размер в микрометрах |
|---|---|
| Массистерия voersi [20] | 2.3–3 |
| Неглерия Фаулери [23] | 8–15 |
| Нейтрофил (лейкоцит) [24] | 12–15 |
| Акантамеба [25] | 12–40 |
| Энтамеба гистолитическая [26] | 15–60 |
| Арселла обыкновенная [27] | 30–152 |
| Амеба протей [28] | 220–760 |
| Каролина хаос [29] | 700–2000 |
| Пеломикса болотная [30] | до 5000 |
| Очень хрупкий Сирингамин [22] | до 200 000 |
Амебы как специализированные клетки и стадии жизненного цикла
[ редактировать ]
Некоторые многоклеточные организмы имеют амебоидные клетки только в определенные фазы жизни или используют амебоидные движения для выполнения специализированных функций. В иммунной системе человека и других животных амебоидные лейкоциты преследуют вторгшиеся организмы, такие как бактерии и патогенные протисты, и поглощают их путем фагоцитоза . [31]
Амебоидные стадии встречаются также у многоклеточных грибоподобных простейших, так называемых слизевиков . Как плазмодиальные слизевики, в настоящее время отнесенные к классу Myxogastria , так и клеточные слизевики групп Acrasida и Dictyosteliida , на стадии питания живут как амебы. Амебоидные клетки первых объединяются, образуя гигантский многоядерный организм. [32] в то время как клетки последних живут отдельно, пока не закончится пища, в это время амебы объединяются, образуя многоклеточный мигрирующий «слизень», функционирующий как единый организм. [8]
Другие организмы также могут представлять амебоидные клетки на определенных стадиях жизненного цикла, например, гаметы некоторых зеленых водорослей ( Zygnematophyceae ). [33] и пеннатные диатомеи , [34] споры (или фазы распространения) некоторых Mesomycetozoea , [35] [36] и спороплазмы стадия Myxozoa и Ascetosporea . [37]
Ставки амеб
[ редактировать ]Ранняя история и происхождение Саркодины
[ редактировать ]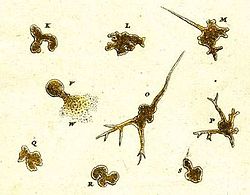

Самая ранняя запись об амебоидном организме была сделана в 1755 году Августом Иоганном Рёзелем фон Розенгофом , который назвал свое открытие «Der Kleine Proteus» («Маленький Протей»). [38] На иллюстрациях Рёзеля изображена неопознанная пресноводная амеба, внешне похожая на обычный вид, ныне известный как Amoeba proteus . [39] Термин «Proteus Animalcule» использовался на протяжении 18 и 19 веков как неофициальное название любого крупного свободноживущего амебоида. [40]
В 1822 году род Amiba (от греческого ἀμοιβή amoibe , что означает «изменение») был выделен французским натуралистом Бори де Сен-Винсентом . [41] [42] Современник Бори, К. Г. Эренберг , принял этот род в свою собственную классификацию микроскопических существ, но изменил написание на Amoeba . [43]
В 1841 году Феликс Дюжарден ввёл термин « саркод » (от греческого σάρξ sarx — «плоть» и εἶδος eidos — «форма») для обозначения «толстого, клейкого, однородного вещества», заполняющего тела простейших клеток. [44] Хотя этот термин первоначально относился к протоплазме любого простейшего, вскоре он стал использоваться в ограниченном смысле для обозначения студенистого содержимого амебоидных клеток. [10] Тридцать лет спустя австрийский зоолог Людвиг Карл Шмарда использовал «саркод» в качестве концептуальной основы для своего подразделения Sarcodea, группы уровня типа , состоящей из «нестабильных, изменчивых» организмов с телами, в основном состоящими из «саркода». [45] Более поздние исследователи, в том числе влиятельный систематик Отто Бючли , внесли поправки в эту группу, создав класс Sarcodina. [46] таксон , который широко использовался на протяжении большей части 20 века.
В рамках традиционной саркодины амебы обычно делились на морфологические категории на основе формы и строения их ложноножек . Амебы с псевдоподиями, поддерживаемыми регулярными массивами микротрубочек (такие как пресноводные Heliozoa и морские радиолярии ), были классифицированы как Actinopoda ; тогда как те, у кого псевдоподии не имели опоры, были классифицированы как Rhizopoda . [47] В зависимости от морфологии псевдоподий корненожки подразделялись на лопастные, нитевидные и сетчатые амебы.
Демонтаж Саркодины
[ редактировать ]В последнее десятилетие 20-го века серия молекулярно-филогенетических анализов подтвердила, что саркодины не являются монофилетической группой. Ввиду этих результатов от старой схемы отказались, а амебы Sarcodina были рассеяны среди многих других таксономических групп высокого уровня. Сегодня большинство традиционных саркодинов относят к двум супергруппам эукариот : Amoebozoa и Rhizaria . Остальные распределены по раскопам , опистоконтам и страменопилям . Некоторые из них, например Centrohelida , еще не отнесены ни к одной супергруппе. [10] [48]
Классификация
[ редактировать ]Недавняя классификация относит различные роды амебоидов к следующим группам:
| Супергруппы | Основные группы и роды | Морфология |
|---|---|---|
| амебозоа |
|
|
| Ризария |
| |
| Раскопки |
|
|
| Гетероконт |
|
|
| Альвеолата |
| |
| Опистоконта |
|
|
| Разгруппировано / неизвестный |
Некоторые из приведенных амебоидных групп (например, часть хризофитов , часть ксантофитов , хлорарахниофитов ) традиционно не включались в состав Sarcodina, относя их к водорослям или жгутиковым простейшим.
Патогенные взаимодействия с другими организмами
[ редактировать ]
заражать другие организмы Некоторые амебы могут патогенно , вызывая заболевания: [52] [53] [54] [55]
- Entamoeba histolytica является причиной амебиаза или амебной дизентерии.
- Naegleria fowleri («амёба, поедающая мозг») — это вид, обитающий в пресной воде, который может быть смертельным для человека при попадании через нос.
- Акантамеба может вызывать амебный кератит и энцефалит . у человека
- Balamuthia mandrillaris является причиной (часто смертельного) гранулематозного амебного менингоэнцефалита .
Было обнаружено, что амебы собирают и выращивают бактерии, вызывающие чуму . [56] Амебы также могут быть хозяином микроскопических организмов, патогенных для людей, и способствовать распространению таких микробов. Бактериальные возбудители (например, легионеллы ) могут препятствовать усвоению пищи при поедании амеб. [57] В настоящее время широко используемыми и наиболее изученными амебами, являющимися хозяевами других организмов, являются Acanthamoeba castellanii и Dictyostelium discoideum. [58] Микроорганизмы, способные преодолеть защиту одноклеточных организмов, могут укрываться и размножаться внутри них, где их хозяева защищают их от неблагоприятных внешних условий.
Мейоз
[ редактировать ]Недавние данные показывают, что некоторые линии Amoebozoa подвергаются мейозу .
ортологи генов, участвующих в мейозе половых эукариот были идентифицированы акантамебы Недавно в геноме . Эти гены включали Spo11 , Mre11 , Rad50 , Rad51 , Rad52 , Mnd1, Dmc1 , Msh и Mlh . [59] Это открытие предполагает, что акантамеба способна к той или иной форме мейоза и может подвергаться половому размножению.
Специфичная для мейоза рекомбиназа Dmc1 гомологичной необходима для эффективной мейотической рекомбинации , а Dmc1 экспрессируется в Entamoeba histolytica . [60] Очищенный Dmc1 из E. histolytica образует пресинаптические нити и катализирует АТФ -зависимое гомологичное спаривание ДНК и обмен цепей ДНК по меньшей мере на нескольких тысячах пар оснований . [60] Реакции спаривания ДНК и обмена цепей усиливаются эукариотическим специфичным для мейоза дополнительным фактором рекомбинации (гетеродимером) Hop2-Mnd1. [60] Эти процессы играют центральную роль в мейотической рекомбинации, что позволяет предположить, что E. histolytica подвергается мейозу. [60]
Исследования Entamoeba Invendes показали, что при превращении из тетраплоидного одноядерного трофозоита к четырехядерной кисте гомологичная рекомбинация усиливается. [61] Экспрессия генов, функции которых связаны с основными этапами мейотической рекомбинации, также увеличивается во время инцистации. [61] Эти данные у E. Inventens в сочетании с данными исследований E. histolytica указывают на наличие мейоза у Entamoeba .
Dictyostelium discoideum в супергруппе Amoebozoa может подвергаться спариванию и половому размножению, включая мейоз, при нехватке пищи. [62] [63]
Поскольку Amoebozoa рано отошли от генеалогического древа эукариот , эти результаты позволяют предположить, что мейоз присутствовал на ранних этапах эволюции эукариот. Более того, эти результаты согласуются с предложением Lahr et al. [64] что большинство амебоидных линий в древности были половыми.
Ссылки
[ редактировать ]- ^ «Амеба». Архивировано 22 ноября 2015 г. в Wayback Machine на Oxforddictionaries.com.
- ^ Синглтон, Пол (2006). Словарь по микробиологии и молекулярной биологии, 3-е издание, переработанное . Чичестер, Великобритания: John Wiley & Sons. стр. 32 . ISBN 978-0-470-03545-0 .
- ^ Jump up to: а б Дэвид Дж. Паттерсон. «Амебы: протисты, которые передвигаются и питаются с помощью псевдоподий» . Веб-проект «Древо жизни». Архивировано из оригинала 15 июня 2010 года . Проверено 21 сентября 2009 г.
- ^ «Амебы» . Эдинбургский университет. Архивировано из оригинала 10 июня 2009 года.
- ^ Вим ван Эгмонд. «Солнечные зверекулы и амебы» . Микроскопия-Великобритания. Архивировано из оригинала 4 ноября 2005 года . Проверено 23 октября 2005 г.
- ^ Флор-Парра, Игнасио; Берналь, Мануэль; Журинский, Яков; Дага, Рафаэль Р. (17 декабря 2013 г.). «Миграция и деление клеток амебоидноподобных делящихся дрожжей» . Биология Открытая . 3 (1): 108–115. дои : 10.1242/bio.20136783 . ISSN 2046-6390 . ПМЦ 3892166 . ПМИД 24357230 .
- ^ Фридл, П.; Боргманн, С.; Брёкер, Э.Б. (1 октября 2001 г.). «Амебоидные лейкоциты, ползущие по внеклеточному матриксу: уроки парадигмы движения клеток Dictyostelium». Журнал биологии лейкоцитов . 70 (4): 491–509. дои : 10.1189/jlb.70.4.491 . ISSN 0741-5400 . ПМИД 11590185 . S2CID 28731650 .
- ^ Jump up to: а б Мари, Афанасий FM; Хогевег, Паулин (2001). «Как амебоиды самоорганизуются в плодовое тело: многоклеточная координация у Dictyostelium discoideum» . Труды Национальной академии наук . 98 (7): 3879–3883. дои : 10.1073/pnas.061535198 . ПМК 31146 . ПМИД 11274408 .
- ^ Маккеррас, MJ; Эрколе, QN (1947). «Наблюдения за действием палудрина на малярийных паразитов». Труды Королевского общества тропической медицины и гигиены . 41 (3): 365–376. дои : 10.1016/s0035-9203(47)90133-8 . ПМИД 18898714 .
- ^ Jump up to: а б с Ян Павловский: Сумерки Саркодины: молекулярный взгляд на полифилетическое происхождение амебоидных простейших . Протистология, Группа 5, 2008, С. 281–302. (pdf, 570 КБ). Архивировано 14 июня 2013 г. в Wayback Machine.
- ^ Тан; и др. (2005). «Простая массовая культура амебы Chaos carolinense: пересмотр» (PDF) . Протистология . 4 : 185–90. Архивировано (PDF) из оригинала 29 сентября 2017 года . Проверено 28 сентября 2017 г.
- ^ «Отношения с людьми» . Амеба протей . 12 апреля 2013 года. Архивировано из оригинала 29 сентября 2017 года . Проверено 28 сентября 2017 г.
- ^ Альбертс ред.; и др. (2007). Молекулярная биология клетки, 5-е издание . Нью-Йорк: Garland Science. п. 1037. ИСБН 9780815341055 .
- ^ Маргулис, Линн (2009). Королевства и владения . Академическая пресса. стр. 206–7 . ISBN 978-0-12-373621-5 .
- ^ Огден, CG (1980). Атлас пресноводной раковинной амебы . Оксфорд, Лондон и Глазго: Издательство Оксфордского университета, для Британского музея (естественная история). стр. 1–5. ISBN 978-0198585022 .
- ^ Альбертс ред.; и др. (2007). Молекулярная биология клетки, 5-е издание . Нью-Йорк: Garland Science. п. 663. ИСБН 9780815341055 .
- ^ Кудо, Ричард Роксабро. «Протозоология». Протозоология, 4-е издание (1954). п. 83
- ^ Торп, Джеймс Х. (2001). Экология и классификация пресноводных беспозвоночных Северной Америки. Сан-Диего: Академик. п. 71. ISBN 0-12-690647-5 .
- ^ Чон, Кван В. (1973). Биология амебы . Нью-Йорк: Академическая пресса. стр. 100 . ISBN 9780123848505 .
- ^ Jump up to: а б Мыльников Александр П.; Вебер, Феликс; Юргенс, Клаус; Вилезич, Клаудия (1 августа 2015 г.). «У Massisteria marina есть сестра: Massisteria voersi sp. nov., редкий вид, выделенный из прибрежных вод Балтийского моря». Европейский журнал протистологии . 51 (4): 299–310. дои : 10.1016/j.ejop.2015.05.002 . ISSN 1618-0429 . ПМИД 26163290 .
- ^ «Размер, форма и расположение бактериальных клеток» . Classes.midlandstech.edu . Архивировано из оригинала 9 августа 2016 года . Проверено 21 августа 2016 г.
- ^ Jump up to: а б Добрый день, Эй Джей; Аранда да Силва, А.; Павловский, Дж. (1 декабря 2011 г.). «Ксенофиофоры (Rhizaria, Foraminifera) из каньона Назаре (окраина Португалии, северо-восток Атлантики)». Глубоководные исследования. Часть II: Актуальные исследования в океанографии . Геология, геохимия и биология подводных каньонов к западу от Португалии. 58 (23–24): 2401–2419. Бибкод : 2011DSRII..58.2401G . дои : 10.1016/j.dsr2.2011.04.005 .
- ^ «Амеба, поедающая мозг (Naegleria Fowleri): причины и симптомы» . Архивировано из оригинала 21 августа 2016 года . Проверено 21 августа 2016 г.
- ^ «Анатомические атласы: Атлас микроскопической анатомии: Раздел 4: Кровь» . www.anatomyatlases.org . Архивировано из оригинала 19 августа 2016 года . Проверено 21 августа 2016 г.
- ^ « Акантамеба | Микромир» . www.arcella.nl . Архивировано из оригинала 18 августа 2016 года . Проверено 21 августа 2016 г.
- ^ «Микроскопия Entamoeba histolytica » . msu.edu . Архивировано из оригинала 5 октября 2016 года . Проверено 21 августа 2016 г.
- ^ « Arcella vulgaris | Микромир» . www.arcella.nl . Архивировано из оригинала 18 августа 2016 года . Проверено 21 августа 2016 г.
- ^ « Амеба протей | Микромир» . www.arcella.nl . Архивировано из оригинала 18 августа 2016 года . Проверено 21 августа 2016 г.
- ^ « Хаос | Микромир» . www.arcella.nl . Архивировано из оригинала 12 октября 2016 года . Проверено 21 августа 2016 г.
- ^ « Pelomyxa palustris | Микромир» . www.arcella.nl . Архивировано из оригинала 18 августа 2016 года . Проверено 21 августа 2016 г.
- ^ Фридл, Питер; Боргманн, Стефан; Ева-Б, Брёкер (2001). «Амебоидные лейкоциты, ползущие по внеклеточному матриксу: уроки парадигмы движения клеток Dictyostelium». Журнал биологии лейкоцитов . 70 (4): 491–509. дои : 10.1189/jlb.70.4.491 . ПМИД 11590185 . S2CID 28731650 .
- ^ Накагаки; и др. (2000). «Интеллект: решение лабиринта амебоидным организмом» . Природа . 407 (6803): 470. Бибкод : 2000Natur.407..470N . дои : 10.1038/35035159 . ПМИД 11028990 . S2CID 205009141 .
- ^ Вер, Джон Д. (2003). Пресноводные водоросли Северной Америки . Сан-Диего и Лондон: Академическая пресса. стр. 353 . ISBN 978-0-12-741550-5 .
- ^ «Мир водорослей: пол и жизненные циклы диатомей» . Мир водорослей . Королевский ботанический сад Эдинбурга. Архивировано из оригинала 23 сентября 2014 года . Проверено 1 марта 2015 г.
- ^ Валле, LG (2014). «Новые виды Paramoebidium (trichomycetes, Mesomycetozoea) из Средиземноморья с комментариями об амебоидных клетках Amoebidiales». Микология . 106 (3): 481–90. дои : 10.3852/13-153 . ПМИД 24895422 . S2CID 3383757 .
- ^ Тейлор, JW и Берби, ML (2014). Грибы от ПЦР к геномике: распространяющаяся революция в эволюционной биологии. В: Систематика и эволюция . Шпрингер Берлин Гейдельберг. п. 52, [1] Архивировано 30 июня 2015 г. в Wayback Machine.
- ^ Корлисс, Дж.О. (1987). «Протистанская филогения и эукариогенез» . Международный обзор цитологии . 100 : 319–370. дои : 10.1016/S0074-7696(08)61703-9 . ISBN 9780080586373 . ПМИД 3549607 .
- ^ Розенхоф, Р. (1755). Ежемесячные развлечения с насекомыми , том. 3, с. 621, [2] Архивировано 13 июля 2015 года в Wayback Machine .
- ^ Чон, Кван В. (1973). Биология амебы . Нью-Йорк: Академическая пресса. стр. 2–3, [3] . ISBN 9780123848505 .
- ^ Макэлпайн, Дэниел (1881). Биологический атлас: руководство к практическому изучению растений и животных . Эдинбург и Лондон: У. и А. К. Джонстон. стр. 17 .
- ^ Бори де Сен-Винсент, JBGM «Очерк классификации микроскопических животных». Агасс, Париж (1826).с. 28
- ^ МакГрат, Кимберли; Блахфорд, Стейси, ред. (2001). Научная энциклопедия Гейла Том. 1: Трубкозуб-катализатор (2-е изд.). Группа Гейл. ISBN 978-0-7876-4370-6 . OCLC 46337140 .
- ^ Эренберг, Кристиан Готфрид. Организация, систематика и географическое родство настойных животных: Две лекции, прочитанные в Академии наук в Берлине в 1828 и 1830 годах. Типография Королевской академии наук, 1832. с. 59
- ^ Дюжарден, Феликс (1841). Естественная история инфузорий зоофитов . Париж: Энциклопедия Библиотеки Роре. стр. 26 .
- ^ Шмарда, Людвиг Карл (1871). Зоология . В. Браумюллер. стр. 156 .
- ^ Бючли, Отто (1882). Классы и отряды царства животных I. Отдел: Саркодины и Спорозоа . Палеонтологическое развитие Rhisopoda К. Сквагером. п. 1.
- ^ Калкинс, Гэри Н. (1909). Протозоология . Нью-Йорк: Леа и Фебигер. стр. 38–40 .
- ^ Адл, Сина М.; и др. (2012). «Пересмотренная классификация эукариот» . Журнал эукариотической микробиологии . 59 (5): 429–93. дои : 10.1111/j.1550-7408.2012.00644.x . ПМЦ 3483872 . ПМИД 23020233 .
- ^ Jump up to: а б Парк, Дж.С.; Симпсон, AGB; Браун, С.; Чо, Британская Колумбия (2009). «Ультраструктура и молекулярная филогения двух гетеролобозных амеб, Euplaesiobystra Hypersalinica gen. Et sp. Nov. и Tulamoeba peronaphora gen. Et sp. Nov., изолированных из чрезвычайно гиперсоленой среды обитания». Протист . 160 (2): 265–283. дои : 10.1016/j.protis.2008.10.002 . ПМИД 19121603 .
- ^ Отт, Дональд В., Карла К. Олдхэм-Отт, Наталья Рыбалка и Томас Фридл. 2015. Ксантофитовые, эустигматофитовые и рафидофитовые водоросли. В: Вер, Дж.Д., Шит, Р.Г., Кочиолек, Дж.П. (ред.) Пресноводные водоросли Северной Америки: экология и классификация , 2-е издание. Академическое издательство, Амстердам, с. 483–534, [4] Архивировано из оригинала 22 января 2017 года в Wayback Machine .
- ^ Паттерсон, диджей; Симпсон, AGB; Роджерсон, А. (2000). «Амебы неопределенного родства». В: Ли, Джей-Джей; Лидейл, Г.Ф.; Брэдбери, П. Иллюстрированный справочник по простейшим , 2-е изд., Vol. 2, с. 804-827. Лоуренс, Канзас: Общество протозоологов/Allen Press. [5] Архивировано 8 марта 2016 года в Wayback Machine . Роды, которые этот источник считал разгруппированными/неизвестными в 2000 году, но которые с тех пор стали классифицированными, были перенесены в эти классификации в Википедии.
- ^ Casadevall A (2008)Эволюция внутриклеточных патогенов. Анну Преподобный Микробиол 62: 19–33. 10.1146/annurev.micro.61.080706.093305 [PubMed] [CrossRef] [Google Scholar]
- ^ Гимарайнш AJ, Гомес KX, Кортинес JR, Перальта JM, Перальта RHS (2016) Acanthamoeba spp. как универсальный хозяин для патогенных микроорганизмов: один мост от окружающей среды к вирулентности хозяина. Микробиологические исследования 193: 30–38. 10.1016/j.micres.2016.08.001 [PubMed] [CrossRef] [Google Scholar]
- ^ Хилби Х., Вебер С.С., Рагаз С., Найфелер Ю., Урвайлер С. (2007)Хищники окружающей среды как модели бактериального патогенеза. Экологическая микробиология 9: 563–575. 10.1111/j.1462-2920.2007.01238.x [PubMed] [CrossRef] [Google Scholar]
- ^ Греуб, Г; Рауль, Д. (2004). «Микроорганизмы, устойчивые к свободноживущим амебам» . Обзоры клинической микробиологии . 17 (2): 413–433. doi : 10.1128/CMR.17.2.413-433.2004 . ПМК 387402 . ПМИД 15084508 .
- ^ «Являются ли амебы безопасной гаванью для чумы? Новое исследование показывает, что бактерии чумы не только выживают, но и процветают и размножаются после попадания в организм амебы» .
- ^ Видьясагар, Апарна (апрель 2016 г.). «Что такое амеба?» . www.livscience.com . Проверено 8 ноября 2020 г.
- ^ Тьюс, Саша; Солдати, Тьерри; Эйхингер, Людвиг (2019). «Редакционная статья: Амебы как модели-хозяева для изучения взаимодействия с патогенами» . Границы клеточной и инфекционной микробиологии . 9 : 47. дои : 10.3389/fcimb.2019.00047 . ПМЦ 6433779 . ПМИД 30941316 .
- ^ Хан Н.А., Сиддики Р. (2015). «Есть ли свидетельства полового размножения (мейоза) у акантамебы?» . Патог Глоб Здоровье . 109 (4): 193–5. дои : 10.1179/2047773215Y.0000000009 . ПМЦ 4530557 . ПМИД 25800982 .
- ^ Jump up to: а б с д Келсо А.А., Сэй А.Ф., Шарма Д., Ледфорд Л.Л., Терчик А., Саски К.А., Кинг А.В., Аттауэй CC, Темешвари Л.А., Сехорн М.Г. (2015). «Entamoeba histolytica Dmc1 катализирует гомологическое спаривание ДНК и обмен цепей, стимулируемый кальцием и Hop2-Mnd1» . ПЛОС ОДИН . 10 (9): e0139399. Бибкод : 2015PLoSO..1039399K . дои : 10.1371/journal.pone.0139399 . ПМК 4589404 . ПМИД 26422142 .
- ^ Jump up to: а б Сингх Н., Бхаттачарья А., Бхаттачарья С. (2013). «Гомологичная рекомбинация происходит у Entamoeba и усиливается во время стресса роста и конверсии стадий» . ПЛОС ОДИН . 8 (9): е74465. Бибкод : 2013PLoSO...874465S . дои : 10.1371/journal.pone.0074465 . ПМК 3787063 . ПМИД 24098652 .
- ^ Флауэрс Дж.М., Ли С.И., Статос А., Саксер Г., Островский Э.А., Квеллер Д.С., Страссманн Дж.Э., Пуруганан, доктор медицинских наук (2010). «Вариации, пол и социальное сотрудничество: молекулярная популяционная генетика социальной амебы Dictyostelium discoideum» . ПЛОС Генет . 6 (7): e1001013. дои : 10.1371/journal.pgen.1001013 . ПМЦ 2895654 . ПМИД 20617172 .
- ^ О'Дэй Д.Х., Кесей А. (2012). «Сигнализация и пол у социальных амебозойных». Biol Rev Camb Philos Soc . 87 (2): 313–29. дои : 10.1111/j.1469-185X.2011.00200.x . ПМИД 21929567 . S2CID 205599638 .
- ^ Лар DJ, Парфри Л.В., Митчелл Э.А., Кац Л.А., Лара Э (2011). «Целомудрие амеб: переоценка доказательств пола у амебоидных организмов» . Учеб. Биол. Наука . 278 (1715): 2081–90. дои : 10.1098/rspb.2011.0289 . ПМК 3107637 . ПМИД 21429931 .
Дальнейшее чтение
[ редактировать ]- Валочник Дж. и Аспёк Х. (2007). Амебы: яркие примеры проблем филогенетики, классификации и номенклатуры . Денисия 20:323–350. (На немецком языке)
- Амебы: протисты, которые передвигаются и питаются с помощью псевдоподий в веб-проекте «Древо жизни»
- Павловский Дж. и Бурки Ф. (2009). Распутывание филогении амебоидных простейших . Журнал эукариотической микробиологии 56.1: 16–25.
Внешние ссылки
[ редактировать ]- Сименсма, Ф. Микромир: мир амебоидных организмов .
- Фелькер Э. и Клаус С. Визуальный ключ к амебоидным морфотипам . Пенард Лабс.
- Веб-сайт Amoebae лаборатории Maciver Lab Эдинбургского университета объединяет информацию из опубликованных источников.
- Цифровая видеогалерея молекулярных выражений: Жизнь в пруду – Амеба (Простейшие) – информативные видеоролики об амебах
| Базы данных органов управления : Национальные |
|---|