РАД52
| РАД52 | ||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | RAD52 , гомолог RAD52, белок репарации ДНК | |||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 600392 ; МГИ : 101949 ; Гомологен : 31118 ; Генные карты : RAD52 ; ОМА : RAD52 – ортологи | |||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||






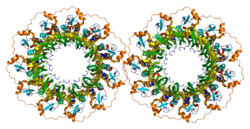
Гомолог RAD52 (S. cerevisiae) , также известный как RAD52 , представляет собой белок , который у человека кодируется RAD52 геном . [ 5 ] [ 6 ]
Функция
[ редактировать ]Белок, кодируемый этим геном, имеет сходство с Saccharomyces cerevisiae Rad52, белком, важным для ДНК восстановления двухцепочечных разрывов и гомологичной рекомбинации . Было показано, что этот генный продукт связывает концы одноцепочечной ДНК и опосредует взаимодействие ДНК-ДНК, необходимое для отжига комплементарных цепей ДНК. Также было обнаружено, что он взаимодействует с белком рекомбинации ДНК RAD51 , что предполагает его роль в рекомбинации и репарации ДНК, связанной с RAD51. [ 6 ]
Роль в рекомбинационной репарации ДНК
[ редактировать ]RAD52 опосредует функцию RAD51 в гомологичной рекомбинационной репарации (HRR) как в дрожжах Saccharomyces cerevisiae , так и в клетках млекопитающих, мышей и человека. Однако белок RAD52 выполняет совершенно разные функции в HRR дрожжей и человека. У S. cerevisiae белок Rad52, действуя отдельно, облегчает загрузку белка Rad51 на одноцепочечную ДНК, предварительно покрытую репликационным белком А, в пресинаптической фазе рекомбинации. [ 7 ] [ 8 ]
Однако у мышей и людей BRCA2 в первую очередь опосредует упорядоченную сборку RAD51 на оцДНК, форме, которая активна для гомологичного спаривания и инвазии цепей. [ 9 ] BRCA2 также перенаправляет RAD51 из дцДНК и предотвращает диссоциацию от оцДНК. [ 9 ] Кроме того, четыре паралога RAD51 RAD51L3 , состоящие из RAD51B ( RAD51L1 ), RAD51C (RAD51L2), RAD51D ( ) , XRCC2, образуют комплекс, называемый комплексом BCDX2. Этот комплекс участвует в рекрутировании или стабилизации RAD51 в местах повреждения. [ 10 ] Комплекс BCDX2, по-видимому, действует, облегчая сборку или стабильность нуклеопротеиновой нити RAD51 . Однако при наличии мутации BRCA2 человеческий RAD52 может опосредовать сборку RAD51 на оцДНК и заменять BRCA2 при гомологичной рекомбинационной репарации ДНК. [ 11 ] хотя и с более низкой эффективностью, чем BRCA2.
Кроме того, человеческий RAD52 в сочетании с ERCC1 способствует предрасположенному к ошибкам пути репарации гомологичной ДНК при одноцепочечном отжиге. [ 12 ] Хотя этот путь восстановления подвержен ошибкам, он может быть необходим для выживания клеток с повреждением ДНК, которое иначе не поддается восстановлению.
Человеческий RAD52 также играет важную роль в восстановлении двухцепочечных разрывов ДНК в активных сайтах транскрипции во время фазы G0/G1 клеточного цикла. Репарация этих двухцепочечных разрывов, по-видимому, использует механизм рекомбинации на основе матрицы РНК, зависящий от RAD52. [ 13 ] Белок синдрома Кокейна B (CSB) (кодируемый ERCC6 ) локализуется в двухцепочечных разрывах в сайтах активной транскрипции, за которым следуют RAD51 , RAD51C и RAD52 для осуществления гомологичной рекомбинационной репарации с использованием вновь синтезированной РНК в качестве матрицы. [ 13 ]
микроРНК и риск рака
[ редактировать ]Три первичных нетранслируемых региона (3'UTR) информационных РНК (мРНК) часто содержат регуляторные последовательности, которые могут вызывать посттранскрипционное молчание РНК . Такие 3'-UTR часто содержат сайты связывания микроРНК (миРНК). Связываясь со специфическими сайтами внутри 3'-UTR, микроРНК могут снижать экспрессию генов различных мРНК, либо ингибируя трансляцию , либо непосредственно вызывая деградацию транскрипта.
МикроРНК (миРНК), по-видимому, регулируют экспрессию более 60% генов, кодирующих белки генома человека. [ 14 ] Одна микроРНК, миР-210 , репрессирует RAD52. [ 15 ] Как отметили Devlin et al., уровень экспрессии миР-210 повышается в большинстве солидных опухолей и отрицательно влияет на клинический исход. [ 16 ]
3'-UTR RAD52 также имеет сайт связывания микроРНК let-7 . Женщины с однонуклеотидным полиморфизмом (SNP) в сайте связывания let-7 (rs7963551), который вызывает снижение связывания let-7, вероятно, имеют повышенную экспрессию RAD52 (как было показано для этого SNP в печени). [ 17 ] ). Женщины с этим SNP в 3'UTR RAD52 продемонстрировали снижение риска рака молочной железы с отношением шансов 0,84, 95% доверительным интервалом 0,75-0,95. [ 18 ]
В китайской популяции хань тот же SNP, что и выше, в 3'-UTR сайта связывания RAD52 для let-7 (rs7963551) снижал риск развития глиомы. Риск глиомы, связанной с генотипом RAD52 rs7963551, имел отношение шансов (по сравнению с лицами без SNP) 0,44 для лиц старше 41 года и отношение шансов 0,58 для лиц 41 года и моложе. [ 19 ]
Ли и др. [ 17 ] обнаружили значительно сниженный риск развития клеточной карциномы печени среди людей с генотипом RAD52 rs7963551 CC (тот же SNP, что и выше) по сравнению с лицами с генотипом AA в китайской популяции. Они также обнаружили, что в 44 образцах нормальной ткани печени человека присутствие SNP rs7963551 было связано со значительным увеличением экспрессии мРНК RAD52.
Таким образом, повышенная экспрессия RAD52 защищает от различных видов рака.
Другое исследование измененных сайтов связывания микроРНК в RAD52 и их влияния на предрасположенность к раку было проведено Naccarati et al. [ 20 ] Они обнаружили два сайта связывания микроРНК RAD52, которые часто менялись и влияли на риск рака толстой кишки. Лица с гомозиготным или гетерозиготным SNP в rs1051669 подвергались повышенному риску рака толстой кишки (ОШ 1,78, 95% ДИ 1,13–2,80, р = 0,01 для гомозигот и ОШ 1,72, 95% ДИ 1,10–2,692, р = 0,02 для гетерозигот). Гетерозиготные носители другого SNP RAD52 (rs11571475) имели пониженный риск рака толстой кишки (ОШ 0,76, 95% ДИ 0,58–1,00, p = 0,05). Из 21 гена гомологичного пути рекомбинационной репарации и 7 генов негомологичного пути соединения концов единственными SNP, обнаруженными в областях связывания микроРНК, которые имели достаточно высокую частоту для оценки и которые влияли на риск рака толстой кишки, были два в RAD52 и один в MRE11A .
Повреждение ДНК, по-видимому, является основной причиной рака. [ 21 ] а нарушения репарации ДНК, по-видимому, лежат в основе многих форм рака. [ 22 ] Если репарация ДНК недостаточна, повреждения ДНК имеют тенденцию накапливаться. Такое избыточное повреждение ДНК может увеличить мутационные ошибки во время репликации ДНК из-за склонного к ошибкам синтеза транслейкоза . Чрезмерное повреждение ДНК может также усилить эпигенетические изменения из-за ошибок во время репарации ДНК. [ 23 ] [ 24 ] Такие мутации и эпигенетические изменения могут привести к раку . Частое увеличение или дефицит RAD52 - опосредованной репарации ДНК, вызванной микроРНК, из-за изменений связывания микроРНК, вероятно, способствует предотвращению или прогрессированию рака молочной железы, головного мозга, печени или толстой кишки.
Взаимодействия
[ редактировать ]Было показано, что RAD52 взаимодействует с RAD51. [ 25 ] Rad52 облегчит загрузку Rad51 в оцДНК, взаимодействуя с белком RPA .
Внутригенная комплементация
[ редактировать ]Когда несколько копий полипептида, кодируемого геном, образуют агрегат, такая белковая структура называется мультимером. Когда мультимер образуется из полипептидов, продуцируемых двумя разными мутантными аллелями конкретного гена, смешанный мультимер может проявлять большую функциональную активность, чем несмешанные мультимеры, образованные каждым из мутантов по отдельности. В таком случае это явление называется внутригенной комплементацией . другие миссенс мутантные Было обнаружено, что мутантный аллель RAD52 Saccharomyces cerevisiae, экспрессирующий усеченный на С-конце белок, дополняет RAD52 - аллели . [ 26 ] Это открытие внутригенной комплементации предполагает, что белок RAD52 имеет мультимерную структуру, которая обеспечивает кооперативные взаимодействия между составляющими его мономерами.
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с ГРЧ38: Версия Ensembl 89: ENSG00000002016 – Ensembl , май 2017 г.
- ^ Перейти обратно: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000030166 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Шен З., Денисон К., Лобб Р., Гейтвуд Дж. М., Чен DJ (январь 1995 г.). «Человеческие и мышиные гомологи дрожжевого гена RAD52: клонирование кДНК, анализ последовательности, отнесение к хромосоме человека 12p12.2-p13 и экспрессия мРНК в тканях мыши» . Геномика . 25 (1): 199–206. дои : 10.1016/0888-7543(95)80126-7 . PMID 7774919 — через Zenodo .
- ^ Перейти обратно: а б EntrezGene 5893, также известный как гомолог RAD52, белок репарации ДНК [Homo sapiens (человек)]
- ^ Шинохара А., Огава Т. (1998). «Стимуляция Rad52 дрожжевой Rad51-опосредованной рекомбинации». Природа . 391 (6665): 404–7. Бибкод : 1998Natur.391..404S . дои : 10.1038/34943 . ПМИД 9450759 . S2CID 4304549 .
- ^ Нью Дж.Х., Сугияма Т., Зайцева Е., Ковальчиковски С.К. (1998). «Белок Rad52 стимулирует обмен цепей ДНК с помощью Rad51 и репликационного белка A». Природа . 391 (6665): 407–10. Бибкод : 1998Natur.391..407N . дои : 10.1038/34950 . ПМИД 9450760 . S2CID 4408959 .
- ^ Перейти обратно: а б Холломан В.К. (2011). «Разгадка механизма гомологичной рекомбинации BRCA2» . Нат. Структура. Мол. Биол. 18 (7): 748–54. дои : 10.1038/nsmb.2096 . ПМЦ 3647347 . ПМИД 21731065 .
- ^ Чун Дж., Бючелмайер Э.С., Пауэлл С.Н. (2013). «Паралоговые комплексы Rad51 BCDX2 и CX3 действуют на разных стадиях BRCA1-BRCA2-зависимого пути гомологичной рекомбинации» . Мол. Клетка. Биол . 33 (2): 387–95. дои : 10.1128/MCB.00465-12 . ПМЦ 3554112 . ПМИД 23149936 .
- ^ Фэн З., Скотт С.П., Буссен В., Шарма Г.Г., Го Дж., Пандита Т.К., Пауэлл С.Н. (2011). «Инактивация Rad52 синтетически смертельна при дефиците BRCA2» . Учеб. Натл. акад. наук. США . 108 (2): 686–91. дои : 10.1073/pnas.1010959107 . ПМК 3021033 . ПМИД 21148102 .
- ^ Старк Дж. М., Пирс А. Дж., О Дж., Пастинк А., Джасин М. (2004). «Генетические этапы гомологичной репарации млекопитающих с отчетливыми мутагенными последствиями» . Мол. Клетка. Биол . 24 (21): 9305–16. дои : 10.1128/MCB.24.21.9305-9316.2004 . ПМК 522275 . ПМИД 15485900 .
- ^ Перейти обратно: а б Вэй Л., Накадзима С., Бём С., Бернштейн К.А., Шен З., Цанг М., Левин А.С., Лан Л. (2015). «Повреждение ДНК во время фазы G0/G1 запускает РНК-матрицу, синдром Коккейна B-зависимую гомологичную рекомбинацию» . Учеб. Натл. акад. наук. США . 112 (27): E3495–504. Бибкод : 2015PNAS..112E3495W . дои : 10.1073/pnas.1507105112 . ПМК 4500203 . ПМИД 26100862 .
- ^ Фридман Р.К., Фарх К.К., Бердж CB, Бартель Д.П. (2009). «Большинство мРНК млекопитающих являются консервативными мишенями микроРНК» . Геном Рез . 19 (1): 92–105. дои : 10.1101/гр.082701.108 . ПМЦ 2612969 . ПМИД 18955434 .
- ^ Кросби М.Э., Кулшрешта Р., Иван М., Глейзер П.М. (2009). «Регуляция микроРНК экспрессии генов репарации ДНК при гипоксическом стрессе» . Рак Рез . 69 (3): 1221–9. дои : 10.1158/0008-5472.CAN-08-2516 . ПМК 2997438 . ПМИД 19141645 .
- ^ Девлин С., Греко С., Мартелли Ф., Иван М. (2011). «МиР-210: Больше, чем тихий игрок в условиях гипоксии» . ИУБМБ Жизнь . 63 (2): 94–100. дои : 10.1002/iub.427 . ПМЦ 4497508 . ПМИД 21360638 .
- ^ Перейти обратно: а б Ли З, Го И, Чжоу Л, Ге Ю, Вэй Л, Ли Л, Чжоу С, Вэй Дж, Юань Q, Ли Дж, Ян М (2015). «Связь функционального генетического варианта RAD52, расположенного в сайте связывания микроРНК, с риском гепатоцеллюлярной карциномы, связанной с HBV» . Мол. Карциног . 54 (9): 853–8. дои : 10.1002/mc.22156 . ПМИД 24729511 . S2CID 25174260 .
- ^ Цзян Ю, Цинь Цз, Ху Цзы, Гуань Х, Ван Ю, Хэ Ю, Сюэ Дж, Лю Икс, Чэнь Дж, Дай Цз, Цзинь Г, Ма Х, Ван С, Шэнь Х (2013). «Генетическая вариация сайта связывания hsa-let-7 в RAD52 связана с предрасположенностью к раку молочной железы» . Канцерогенез . 34 (3): 689–93. дои : 10.1093/carcin/bgs373 . ПМИД 23188672 .
- ^ Лу С, Чен Ю.Д., Хан С., Вэй Дж., Ге Ю., Пан В., Цзян Т., Цю С.Г., Ян М. (2014). «Генетический вариант RAD52, расположенный в сайте связывания микроРНК, связан с риском глиомы у ханьцев». Дж. Нейронкол . 120 (1): 11–7. дои : 10.1007/s11060-014-1527-x . ПМИД 25012956 . S2CID 1082923 .
- ^ Наккарати А, Роза Ф, Выметалкова В, Бароне Е, Йираскова К, Ди Гаэтано С, Новотный Дж, Леви М, Водичкова Л, Джеминьяни Ф, Бухлер Т, Ланди С, Водика П, Пардини Б (2015). «Репарация двухцепочечного разрыва и колоректальный рак: варианты генов в пределах 3'-НТО и связывание микроРНК как модуляторы риска рака и клинического исхода» . Онкотаргет . 7 (17): 23156–69. дои : 10.18632/oncotarget.6804 . ПМК 5029617 . ПМИД 26735576 .
- ^ Кастан М.Б. (2008). «Реакция на повреждение ДНК: механизмы и роль в заболеваниях человека: лекция на премию Мемориала ГСГ Клоуза 2007 г.» . Мол. Рак Рез . 6 (4): 517–24. дои : 10.1158/1541-7786.MCR-08-0020 . ПМИД 18403632 .
- ^ Харпер Дж.В., Элледж С.Дж. (2007). «Реакция на повреждение ДНК: десять лет спустя» . Мол. Клетка . 28 (5): 739–45. doi : 10.1016/j.molcel.2007.11.015 . ПМИД 18082599 .
- ^ О'Хаган Х.М., Мохаммад Х.П., Бэйлин С.Б. (2008). «Двухнитевые разрывы могут инициировать молчание генов и SIRT1-зависимое начало метилирования ДНК на экзогенном промоторном острове CpG» . ПЛОС Генетика . 4 (8): е1000155. дои : 10.1371/journal.pgen.1000155 . ПМЦ 2491723 . ПМИД 18704159 .
- ^ Куоццо С, Порчеллини А, Ангризано Т, Морано А, Ли Б, Ди Пардо А, Мессина С, Юлиано Р, Фуско А, Сантильо М.Р., Мюллер М.Т., Кьяриотти Л., Готтесман М.Е., Авведименто Э.В. (июль 2007 г.). «Повреждение ДНК, репарация, направленная на гомологию, и метилирование ДНК» . ПЛОС Генетика . 3 (7): е110. дои : 10.1371/journal.pgen.0030110 . ЧВК 1913100 . ПМИД 17616978 .
- ^ Чен Г, Юань СС, Лю В, Сюй Ю, Трухильо К, Сонг Б, Конг Ф, Гофф С.П., Ву Ю, Арлингхаус Р, Балтимор Д., Гассер П.Дж., Парк М.С., Сунг П., Ли Э.Ю. (апрель 1999 г.). «Радиационно-индуцированная сборка рекомбинационного комплекса Rad51 и Rad52 требует ATM и c-Abl» (PDF) . Журнал биологической химии . 274 (18): 12748–52. дои : 10.1074/jbc.274.18.12748 . ПМИД 10212258 . S2CID 2587580 .
- ^ Баунди-Миллс КЛ, Ливингстон DM. Аллель Saccharomyces cerevisiae RAD52, экспрессирующая С-концевой укороченный белок: активность и внутригенная комплементация миссенс-мутаций. Генетика. 1993;133(1):39-49.
Дальнейшее чтение
[ редактировать ]- Мурис Д.Ф., Беззубова О., Бюрстедде Дж.М., Врикен К., Баладжи А.С., Осгуд С.Дж., Троелстра С., Хоймейкерс Дж.Х., Остерманн К., Шмидт Х. (ноябрь 1994 г.). «Клонирование генов человека и мыши, гомологичных RAD52, дрожжевому гену, участвующему в репарации и рекомбинации ДНК» . Мутационные исследования . 315 (3): 295–305. дои : 10.1016/0921-8777(94)90040-х . hdl : 1765/3072 . ПМИД 7526206 .
- Шен З., Денисон К., Лобб Р., Гейтвуд Дж. М., Чен DJ (январь 1995 г.). «Человеческие и мышиные гомологи дрожжевого гена RAD52: клонирование кДНК, анализ последовательности, отнесение к хромосоме человека 12p12.2-p13 и экспрессия мРНК в тканях мыши» . Геномика . 25 (1): 199–206. дои : 10.1016/0888-7543(95)80126-7 . ПМИД 7774919 .
- Парк М.С. (июнь 1995 г.). «Экспрессия человеческого RAD52 придает устойчивость к ионизирующей радиации клеткам млекопитающих» . Журнал биологической химии . 270 (26): 15467–70. дои : 10.1074/jbc.270.26.15467 . ПМИД 7797537 .
- Шен З., Пардингтон-Пуртимун П.Е., Комо Дж.К., Мойзис Р.К., Чен DJ (сентябрь 1996 г.). «UBL1, человеческий убиквитиноподобный белок, ассоциированный с человеческими белками RAD51/RAD52» . Геномика . 36 (2): 271–9. дои : 10.1006/geno.1996.0462 . ПМИД 8812453 .
- Шен З., Пардингтон-Пуртимун П.Е., Комо Дж.К., Мойзис Р.К., Чен DJ (октябрь 1996 г.). «Ассоциации UBE2I с белками RAD52, UBL1, p53 и RAD51 в двугибридной системе дрожжей» . Геномика . 37 (2): 183–6. дои : 10.1006/geno.1996.0540 . ПМИД 8921390 .
- Чэнь Г, Юань СС, Лю В, Сюй Ю, Трухильо К, Сонг Б, Конг Ф, Гофф С.П., Ву Ю, Арлингхаус Р., Балтимор Д., Гассер П.Дж., Парк М.С., Сунг П., Ли Э.Ю. (апрель 1999 г.). «Радиационно-индуцированная сборка рекомбинационного комплекса Rad51 и Rad52 требует ATM и c-Abl» (PDF) . Журнал биологической химии . 274 (18): 12748–52. дои : 10.1074/jbc.274.18.12748 . ПМИД 10212258 . S2CID 2587580 .
- Кито К., Вада Х., Йе Э.Т., Камитани Т. (декабрь 1999 г.). «Идентификация новых изоформ человека RAD52». Биохимия и биофизика Acta (BBA) - Структура и экспрессия генов . 1489 (2–3): 303–14. дои : 10.1016/s0167-4781(99) 00214-6 ПМИД 10673031 .
- Стасяк А.З., Ларкет Э., Стасяк А., Мюллер С., Энгель А., Ван Дайк Э., Вест СК, Эгельман Э.Х. (март 2000 г.). «Человеческий белок Rad52 существует в виде гептамерного кольца» . Современная биология . 10 (6): 337–40. Бибкод : 2000CBio...10..337S . дои : 10.1016/S0960-9822(00)00385-7 . ПМИД 10744977 . S2CID 16087838 .
- Парсонс Калифорния, Бауманн П., Ван Дайк Э., Вест Южная Каролина (август 2000 г.). «Точное связывание концов одноцепочечной ДНК человеческим белком RAD52» . Журнал ЭМБО . 19 (15): 4175–81. дои : 10.1093/emboj/19.15.4175 . ПМК 306603 . ПМИД 10921897 .
- Мер Г, Бочкарев А, Гупта Р, Бочкарева Е, Фраппье Л, Инглес С.Дж., Эдвардс А.М., Чазин В.Дж. (октябрь 2000 г.). «Структурная основа распознавания белков репарации ДНК UNG2, XPA и RAD52 фактором репликации RPA» . Клетка . 103 (3): 449–56. дои : 10.1016/S0092-8674(00)00136-7 . ПМИД 11081631 . S2CID 16640087 .
- Ранатунга В., Джексон Д., Флауэрс II РА, Боргшталь GE (июль 2001 г.). «Человеческий белок RAD52 обладает чрезвычайной термостабильностью». Биохимия . 40 (29): 8557–62. CiteSeerX 10.1.1.566.9388 . дои : 10.1021/bi0155089 . ПМИД 11456495 .
- Ван Дайк Э., Стасяк А.З., Стасяк А., Вест СК (октябрь 2001 г.). «Визуализация промежуточных продуктов рекомбинации, полученных путем одноцепочечного отжига, опосредованного RAD52» . Отчеты ЭМБО . 2 (10): 905–9. doi : 10.1093/embo-reports/kve201 . ПМЦ 1084079 . ПМИД 11571269 .
- Ким П.М., Аллен С., Вагенер Б.М., Шен З., Николофф Дж.А. (ноябрь 2001 г.). «Сверхэкспрессия человеческих RAD51 и RAD52 снижает гомологичную рекомбинацию, индуцированную двухцепочечным разрывом, в клетках млекопитающих» . Исследования нуклеиновых кислот . 29 (21): 4352–60. дои : 10.1093/нар/29.21.4352 . ПМК 60192 . ПМИД 11691922 .
- Яньес Р.Дж., Портер AC (февраль 2002 г.). «Дифференциальные эффекты сверхэкспрессии Rad52p на нацеливание генов и внехромосомную гомологичную рекомбинацию в клеточной линии человека» . Исследования нуклеиновых кислот . 30 (3): 740–8. дои : 10.1093/нар/30.3.740 . ПМК 100286 . ПМИД 11809887 .
- Джексон Д., Дхар К., Валь Дж.К., Уолд М.С., Боргстал GE (август 2002 г.). «Анализ комплекса репликационного белка A человека: Rad52: доказательства перекрестных помех между RPA32, RPA70, Rad52 и ДНК». Журнал молекулярной биологии . 321 (1): 133–48. дои : 10.1016/S0022-2836(02)00541-7 . ПМИД 12139939 .
- Кагава В., Курумизака Х., Ишитани Р., Фукаи С., Нуреки О., Сибата Т., Ёкояма С. (август 2002 г.). «Кристаллическая структура домена гомологичных пар рекомбиназы Rad52 человека в ундекамерной форме» . Молекулярная клетка . 10 (2): 359–71. дои : 10.1016/S1097-2765(02)00587-7 . ПМИД 12191481 .
- Синглтон М.Р., Вентцелл Л.М., Лю Ю., Вест С.К., Вигли Д.Б. (октябрь 2002 г.). «Структура одноцепочечного домена отжига человеческого белка RAD52» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (21): 13492–7. Бибкод : 2002PNAS...9913492S . дои : 10.1073/pnas.212449899 . ПМК 129701 . ПМИД 12370410 .
- Лю Дж, Мэн X, Шэнь Z (октябрь 2002 г.). «Ассоциация человеческого белка RAD52 с факторами транскрипции». Связь с биохимическими и биофизическими исследованиями . 297 (5): 1191–6. дои : 10.1016/S0006-291X(02)02353-7 . ПМИД 12372413 .
- Хан Дж., Хэнкинсон С.Э., Де Виво I, Кольдиц Г.А., Хантер DJ (октябрь 2002 г.). «Нет связи между полиморфизмом стоп-кодонов в RAD52 и риском рака молочной железы». Эпидемиология рака, биомаркеры и профилактика . 11 (10, ч. 1): 1138–9. ПМИД 12376524 .
- Китао Х., Юань З.М. (декабрь 2002 г.). «Регуляция образования ядерных фокусов Rad52, индуцированного ионизирующим излучением, путем фосфорилирования, опосредованного c-Abl» . Журнал биологической химии . 277 (50): 48944–8. дои : 10.1074/jbc.M208151200 . ПМИД 12379650 .
галерея PDB |
|---|

