амебозоа
| амебозоа Временной диапазон:
| |
|---|---|

| |
| Каролинский хаос | |
| Научная классификация | |
| Домен: | Эукариоты |
| Клэйд : | Аморфея |
| Тип: | амебозоа Люэ , 1913 год , ред. Кавальер-Смит , 1998 г. [ 3 ] |
| Клады [ 4 ] [ 5 ] | |
| Синонимы | |
| |
Amoebozoa — крупная таксономическая группа, содержащая около 2400 описанных видов амебоидных простейших . [ 7 ] часто имеют тупые, пальцеобразные, лопастные ложноножки и трубчатые митохондриальные кристы . [ 6 ] [ 8 ] В традиционных классификационных схемах Amoebozoa обычно относят к царства типу Protista . [ 9 ] или царство простейших . [ 10 ] [ 11 ] В классификации, одобренной Международным обществом протистологов, он сохраняется как « супергруппа » без ранга среди эукариот. [ 6 ] Молекулярно-генетический анализ подтверждает, что Amoebozoa является монофилетической кладой . Современные исследования филогенетических деревьев эукариот идентифицируют его как сестринскую группу Opisthokonta , другой крупной клады, которая включает как грибы , так и животных , а также несколько других клад, включающих около 300 видов одноклеточных эукариот. [ 7 ] [ 8 ] Amoebozoa и Opisthokonta иногда группируются в таксон высокого уровня , называемый по-разному Unikonta . [ 10 ] Аморфея [ 6 ] или Опимода . [ 12 ]
Amoebozoa включает в себя многие из наиболее известных амебоидных организмов, таких как Chaos , Entamoeba , Pelomyxa и сам род Amoeba . Виды Amoebozoa могут быть либо покрыты панцирем (раковиной), либо голыми, а клетки могут иметь жгутики . Свободноживущие виды распространены как в соленой, так и в пресной воде, а также в почве, мхах и опавших листьях. Некоторые из них живут как паразиты или симбионты других организмов, а некоторые, как известно, вызывают заболевания у людей и других организмов.
Хотя большинство видов амебозойных являются одноклеточными, группа также включает несколько клад слизевиков , которые имеют макроскопическую, многоклеточную стадию жизни, в течение которой отдельные амебоидные клетки остаются вместе после множественного деления клеток, образуя макроскопический плазмодий или, в клеточных слизевиках. , объединяются в одно.
Амебозоа сильно различаются по размеру. Некоторые из них имеют диаметр всего 10–20 мкм, а другие относятся к числу крупнейших простейших. Хорошо известный вид Amoeba proteus , длина которого может достигать 800 мкм, часто изучается в школах и лабораториях как репрезентативная клетка или модельный организм , отчасти из-за его удобного размера. Многоядерные амебы, такие как Chaos и Pelomyxa, могут достигать нескольких миллиметров в длину, а некоторые многоклеточные амебозои, такие как слизевик «собачьей рвоты» Fuligo septica , могут занимать площадь в несколько квадратных метров. [ 13 ]
Морфология
[ редактировать ]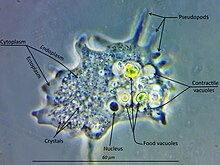
Amoebozoa — большая и разнообразная группа, но некоторые черты являются общими для многих ее представителей. Клетка амебозои обычно делится на зернистую центральную массу, называемую эндоплазмой , и чистый внешний слой, называемый эктоплазмой. Во время передвижения эндоплазма течет вперед, а эктоплазма движется назад вдоль внешней стороны клетки. В движении многие амебозои имеют четко выраженную переднюю и заднюю часть и могут принимать «моноподиальную» форму, при этом вся клетка функционирует как одна ложноножка. Большие псевдоподии могут образовывать многочисленные четкие выступы, называемые субпсевдоподиями (или детерминированные псевдоподии ), которые вытягиваются на определенную длину, а затем втягиваются либо с целью передвижения, либо с целью приема пищи. Клетка может также образовывать множественные неопределенные псевдоподии, через которые все содержимое клетки течет в направлении движения. Они более или менее трубчатые и большей частью заполнены зернистой эндоплазмой. Клеточная масса перетекает в ведущую ложноножку, а остальные в конечном итоге втягиваются, если только организм не меняет направление. [ 14 ]
В то время как большинство амебозойных животных «голые», как известные нам Amoeba и Chaos , или покрыты рыхлой оболочкой из мелких чешуек, как Cochliopodium и Korotnevella , представители отряда Arcellinida образуют жесткие раковины, или панцири , снабженные единственным отверстием, через которое появляются ложноножки. Арцеллинидные тесты могут выделяться из органических материалов, как у Arcella , или состоять из собранных частиц, сцементированных вместе, как у Difflugia .
У всех амеб основным способом питания является фагоцитоз , при котором клетка окружает потенциальные частицы пищи своими ложноножками, запечатывая их в вакуоли, внутри которых они могут перевариваться и всасываться. У некоторых амебозоев есть задняя луковица, называемая уроидом, которая может служить для накопления отходов, периодически отделяющихся от остальной части клетки. [ нужна ссылка ] Когда пищи не хватает, большинство видов могут образовывать цисты , которые можно переносить по воздуху и заносить в новую среду обитания. [ нужна ссылка ] У слизевиков эти структуры называются спорами и образуются на стебельках, называемых плодовыми телами или спорангиями . обнаружены миксотрофные виды, живущие в симбиотических отношениях с микроводорослями рода Chlorella , обитающими внутри цитоплазмы хозяина У Arcellinida и Mayorella . [ 15 ] [ 16 ]
У большинства Amoebozoa отсутствуют жгутики , и, как правило, они не образуют микротрубочками, структур, поддерживаемых за исключением митоза . Однако жгутики встречаются среди архамеб , и многие слизевики производят двужгутиковые гаметы. [ нужна ссылка ] . Жгутик обычно закреплен конусом микротрубочек, что указывает на тесное родство с опистоконтами . [ нужна ссылка ] в Для митохондрий клетках амеб характерны ветвящиеся трубчатые кристы. Однако среди Archamoebae , адаптированных к бескислородным или микроаэрофильным местообитаниям, митохондрии утрачены.
Классификация
[ редактировать ]Место амебозоа на дереве эукариот
[ редактировать ]Похоже (на основании молекулярной генетики), что представители Amoebozoa образуют сестринскую группу животных и грибов, отделившись от этой линии после того, как она отделилась от других групп. [ 17 ] как показано ниже на упрощенной схеме:

Сильное сходство между Amoebozoa и Opisthokonts позволяет предположить, что они образуют отдельную кладу. [ 18 ] Томас Кавальер-Смит предложил название «униконты» (формально Unikonta) для этой ветви, члены которой, как полагали, произошли от общего предка, обладавшего единственным появившимся жгутиком, укорененным в одном базальном теле . [1][2] Однако, хотя тесная связь между Amoebozoa и Opisthokonta твердо поддерживается, недавние исследования показали, что гипотеза об одноресничном предке, вероятно, ошибочна. В своей пересмотренной классификации эукариот (2012) Adl et al. предложил Amorphea как более подходящее название для клады примерно того же состава, сестринской группы Diaphoretickes . [ 6 ] Более поздние работы помещают членов Amorphea вместе с малавимонидами и коллодиктионидами в предполагаемую кладу под названием Opimoda, которая включает в себя одну из двух основных линий, расходящихся у корня древа жизни эукариот, другая — Diphoda . [ 12 ]
Подтипы внутри Amoebozoa: Lobosa и Conosa.
[ редактировать ]Традиционно все амебозои с лопастными ложноножками были сгруппированы в класс Lobosea , помещенный вместе с другими амебоидами в тип Sarcodina или Rhizopoda , но они считались неестественными группами. Структурно-генетические исследования выделили перколозоев и несколько архамеб как самостоятельные группы. В филогениях, основанных на рРНК, их представители были отделены от других амеб и, по-видимому, расходились вблизи основания эукариотической эволюции, как и большинство слизевиков.
Однако деревья были исправлены Кавальер-Смитом и Чао в 1996 году. [ 19 ] предположили, что оставшиеся лобозаны действительно образуют монофилетическую группу, с которой Archamoebae и Mycetozoa были тесно связаны, хотя перколозоа - нет. Впоследствии они внесли изменения в тип Amoebozoa, включив в него как подтип Lobosa, так и новый подтип Conosa , включающий Archamoebae и Mycetozoa . [ 3 ]
Недавние молекулярно-генетические данные, по-видимому, подтверждают это первичное разделение Amoebozoa на Lobosa и Conosa. [ 8 ] Первый, по определению Кавалье-Смита и его сотрудников, состоит в основном из классических Lobosea: нежгутиковых амеб с тупыми лопастными ложноножками ( Amoeba , Acanthamoeba, Arcella, Difflugia и т. д. ). Последний состоит как из амебоидных, так и из жгутиковых клеток, для которых характерны более заостренные или слегка ветвящиеся субпсевдоподии (архемебы и мицетозойные слизевики).
Филогения и систематика Amoebozoa
[ редактировать ]Из более ранних исследований Кавальер-Смита, Чао и Льюиса, 2016 г. [ 20 ] и Силар 2016. [ 21 ] Также недавняя филогения указывает на то, что Lobosa являются парафилетическими: Conosa - сестра Cutosea. [ 5 ] [ 22 ] [ 23 ]
| Филогения амебозой | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
Тип Amoebozoa Lühe 1913, исправление. Кавальер-Смит 1998 [Амебобиота; Eumycetozoa Zopf 1884 исправить Olive 1975 ]
- Clade Discosea Cavalier-Smith 2004 стат. ноябрь Адл и др. 2018 год
- Заказ ? Стереомиксида Грелль 1971
- Заказ ? Стигомебида Смирнов и Кавальер-Смит 2011 г.
- Класс Centramoebia Cavalier-Smith et al. 2016 год
- Заказать Centramoebida Rogerson & Patterson 2002 em. Кавалер-Смит 2004 г.
- Отряд Himatismenida Страница 1987 [Cochliopodiida]
- Отряд Pellitida Страница 1987 [Cochliopodiida]
- Класс Флабеллиния Смирнов & Кавальер-Смит 2011г. Кудрявцев и др. 2014 год
- Заказать Thecamoebida Schaeffer 1926 г.эм. Смирнов и Кавальер-Смит 2011 г.
- Заказать Дермамоебида Кавальер-Смит 2004г.эм. Смирнов и Кавальер-Смит 2011 г.
- Заказать Ваннеллида Смирнова и др. 2005 г.
- Отряд Dactylopodida Смирнов и др. 2005 г.
- Кладе Тевоса Канг и др. 2017 год
- Clade Tubulinea Смирнов и др. 2005 стат. ноябрь Адл и др. 2018 год
- Класс Corycidia Kang et al. Начало 2017 года. 19 ноября Адл и др. 2018 год
- Орден Трихозида Мебиуса 1889 г.
- Семейство Microcoryciidae de Saedeleer, 1934 г.
- Класс Эхинамебия Кавальер-Смит стат. 2016 г. ноябрь Адл и др. 2018 год
- Заказать Echinamoebida Cavalier-Smith 2004 г.эм. 2011 год
- Class Elardia Kang et al. 2017 stat. nov. Adl et al. 2018
- Подкласс Лептомиксия Кавальер-Смит 2016 г.
- Отряд Lptomyxida Pussard & Pons 1976 em. Страница 1987 г.
- Подкласс Eulobosia Cavalier-Smith 2016
- Орден Эуамебида Лепши 1960 г.эм. Кавалер-Смит 2016
- Орден Арцеллиниды Кент 1880 г.
- Подкласс Лептомиксия Кавальер-Смит 2016 г.
- Класс Corycidia Kang et al. Начало 2017 года. 19 ноября Адл и др. 2018 год
- Кладе Эвосеа Канг и др. Статистика 2017 года. Ноябрь Адл и др. 2018 год
- Стат. Clade Cutosa Cavalier-Smith 2016 г. ноябрь
- Класс Cutosea Cavalier-Smith 2016
- Заказать Squamocutida Cavalier-Smith 2016 г.
- Класс Cutosea Cavalier-Smith 2016
- Подтип Conosa Cavalier-Smith, стат 1998. ноябрь
- Инфрафилум Archamoeba Cavalier-Smith 1993 г. стоит. нет. 1998 год
- Класс Archamoebea Cavalier-Smith 1983 стат. н. 2004 г.
- Семейство Tricholimacidae Кавальер-Смит 2013 г.
- Семейство Endamoebidae Calkins 1926 г.
- Орден Entamoebida Cavalier-Smith 1993 г.
- Отряд Pelobiontida. Страница 1976 г., исправлено. Кавалер Смит, 1987 год.
- Класс Archamoebea Cavalier-Smith 1983 стат. н. 2004 г.
- Инфрафилум семиконозия Кавальер-Смит 2013
- Класс Variosea Cavalier-Smith et al. 2004 г.
- Заказ ? Flamellidae Кавалер-Смит 2016
- Заказ ? Холомастигида Лаутерборн 1895 г. [Артодисцида Кавальер-Смит 2013 г. ]
- Орден Фаланстериида Гибберд 1983 г.
- Орден Рамамебида Кавальер-Смит 2016 г.
- Отряд Profilidae Kang et al. 2017 [Protosthelid Olive & Stoianovitch 1966 em. Шадвик и Шпигель, 2012 ].
- Отряд Fractovitellida Lahr et al. 2011 г. Канг и др. 2017 год
- Надкласс Mycetozoa de Bary, 1859 г., бывший Ростафинский, 1873 г.
- Класс Dictyostelea Hawksworth et al. 1983 год
- Орден Acytosteliales Baldauf, Sheikh & Thulin 2017
- Заказать Dictyosteliales Lister 1909 em. Олива 1970 г.
- Класс Protostelea Shadwick & Spiegel et al. 2012 год
- Отряд Protosteliida Shadwick & Spiegel et al. 2012 год
- Класс Ceratiomyxomycetes Hawksworth, Sutton & Ainsworth, 1983 г.
- Отряд Protosporangiida Shadwick & Spiegel 2012 г.
- Отряд Ceratiomyxida Martin 1961 от Farr & Alexopoulos.
- Класс Myxomycetes Link 1833 em. Геккель 1866 г. [ 24 ]
- Подкласс Lucisporomycetidae Леонтьев и др. 2019 год
- Суперотряд Cribrarianae Леонтьев 2015
- Отряд Cribrariales Macbr. 1922 год
- Суперотряд Trichianae Leontiev 2015
- Орден Retulariales Леонтьева 2015 г.
- Орден Лицеалес Ян 1928 г.
- Орден Трихиалеса Макбрайда 1922 г.
- Суперотряд Cribrarianae Леонтьев 2015
- Подкласс Columellomycetidae Леонтьев и др. 2019 год
- Заказ ? Echinosteliopsidales Щепин с соавт.
- Суперотряд Echinostelianae Leontiev 2015
- Отряд Echinosteliales Martin 1961 г.
- Надотряд Stemonitanae Leontiev 2015 [Fuscisporida Cavalier-Smith 2012 ]
- Отряд Clastodermatales Леонтьев 2015 г.
- Заказать Meridermatales Леонтьева 2015г.
- Орден Стемониталес Макбрайд 1922 г.
- Орден Фисаралеса Макбрайда 1922 г.
- Подкласс Lucisporomycetidae Леонтьев и др. 2019 год
- Класс Dictyostelea Hawksworth et al. 1983 год
- Класс Variosea Cavalier-Smith et al. 2004 г.
- Инфрафилум Archamoeba Cavalier-Smith 1993 г. стоит. нет. 1998 год
- Стат. Clade Cutosa Cavalier-Smith 2016 г. ноябрь
- Clade Tubulinea Смирнов и др. 2005 стат. ноябрь Адл и др. 2018 год
Ископаемая запись
[ редактировать ]вазообразной формы Микрофоссилии (VSM), обнаруженные по всему миру, показывают, что амебозои существовали с неопротерозойской эры. Ископаемые виды Melanocyrillium hexodiadema , Palaeoarcella athanata и Hemisphaeriella ornata происходят из пород возрастом 750 миллионов лет. Все три VSM имеют полусферическую форму, вдавленное отверстие и правильные углубления, которые сильно напоминают современных арцеллинид , — панцирных амебозойцев принадлежащих к классу Tubulinea . P. athanata , в частности, выглядит так же, как современный род Arcella . [ 1 ] [ 25 ]
Список амебозойных простейших, патогенных для человека
[ редактировать ]Мейоз
[ редактировать ]Недавно доступная акантамебы последовательность генома выявила несколько ортологов генов, участвующих в мейозе половых эукариот . Эти гены включали Spo11 , Mre11 , Rad50 , Rad51 , Rad52 , Mnd1, Dmc1 , Msh и Mlh . [ 26 ] Это открытие предполагает, что акантамеба способна к той или иной форме мейоза и может подвергаться половому размножению.
У эукариот, размножающихся половым путем, гомологичная рекомбинация (HR) обычно происходит во время мейоза. Специфичная для мейоза рекомбиназа Dmc1 необходима для эффективной мейотической HR, а Dmc1 экспрессируется в Entamoeba histolytica . [ 27 ] Очищенный Dmc1 из E. histolytica образует пресинаптические нити и катализирует АТФ -зависимое гомологичное спаривание ДНК и обмен цепей ДНК по меньшей мере на нескольких тысячах пар оснований . [ 27 ] Реакции спаривания ДНК и обмена цепей усиливаются эукариотическим специфичным для мейоза дополнительным фактором рекомбинации (гетеродимером) Hop2-Mnd1. [ 27 ] Эти процессы играют центральную роль в мейотической рекомбинации, что позволяет предположить, что E. histolytica подвергается мейозу. [ 27 ]
Исследования Entamoeba Inventens показали, что при превращении тетраплоидного одноядерного трофозоита в четырехядерную цисту гомологичная рекомбинация усиливается. [ 28 ] Экспрессия генов, функции которых связаны с основными этапами мейотической рекомбинации, также увеличивалась во время инцистаций. [ 28 ] Эти данные у E. Inventens в сочетании с данными исследований E. histolytica указывают на наличие мейоза у Entamoeba . Сравнительный генетический анализ показал, что мейотические процессы присутствуют во всех основных линиях амебозойных животных. [ 29 ]
Поскольку Amoebozoa рано отошли от генеалогического древа эукариот, эти результаты также позволяют предположить, что мейоз присутствовал на ранних этапах эволюции эукариот.
Здоровье человека
[ редактировать ]Амебиаз , также известный как амебиаз или энтамебиаз, [ 30 ] [ 31 ] – инфекция, вызываемая любым из амебозойных организмов группы Entamoeba . Симптомы наиболее распространены при заражении Entamoeba histolytica . Амебиаз может протекать без симптомов , в легкой или тяжелой форме . Симптомы могут включать боль в животе , легкую диарею , кровавую диарею или тяжелый колит с отмиранием тканей и перфорацией . Последнее осложнение может вызвать перитонит . У пострадавших людей может развиться анемия из-за потери крови. [ 32 ]
Инвазия слизистой оболочки кишечника вызывает амебную кровавую диарею или амебный колит . Если паразит попадает в кровоток, он может распространиться по организму, чаще всего попадая в печень, где вызывает амебные абсцессы печени . Абсцессы печени могут возникать без предшествующей диареи. Цисты Entamoeba могут сохраняться до месяца в почве или до 45 минут под ногтями. Важно различать амебиаз и бактериальный колит . Предпочтительным методом диагностики является исследование кала под микроскопом, но он требует квалифицированного микроскописта и не может быть надежным методом исключения инфекции. Однако этот метод может не обеспечивать разделение между конкретными типами. Увеличение количества лейкоцитов наблюдается в тяжелых случаях, но не в легких. Самый точный тест — на наличие антител в крови , но после лечения он может оставаться положительным. [ 32 ]
Профилактика амебиаза заключается в отделении пищи и воды от фекалий и принятии надлежащих санитарных мер. Вакцины нет. Существует два варианта лечения в зависимости от локализации инфекции. Амебиаз в тканях лечат метронидазолом , тинидазолом , нитазоксанидом , дегидроэметином или хлорохином , тогда как просветную инфекцию лечат фуроатом дилоксанида или йодхинолином . Чтобы лечение было эффективным против всех стадий амебы, может потребоваться комбинация лекарств. Инфекции без симптомов не требуют лечения, но инфицированные люди могут передавать паразита другим, и можно рассмотреть возможность лечения. Лечение других инфекций Entamoeba , кроме E. histolytica, не требуется. [ 32 ]
Амебиаз распространен во всем мире. [ 33 ] Около 480 миллионов человек инфицированы E. histolytica , что приводит к смерти от 40 000 до 110 000 человек каждый год. Большинство инфекций в настоящее время приписывается E. dispar . E. dispar чаще встречается в определенных районах, и случаев с симптомами может быть меньше, чем сообщалось ранее. Первый случай амебиаза был зарегистрирован в 1875 году, а в 1891 году заболевание было подробно описано, в результате чего появились термины амебная дизентерия и амебный абсцесс печени . Дальнейшие данные, полученные на Филиппинах в 1913 году, показали, что после проглатывания цист E. histolytica у добровольцев развилось заболевание. С 1897 года было известно, что по крайней мере один не вызывающий болезней вид Entamoeba существует ( Entamoeba coli ), но впервые официально признано ВОЗ в 1997 году, что E. histolytica представляет собой два вида, несмотря на то, что это было впервые предложено в 1997 году. 1925. Помимо ныне признанного E. dispar, данные показывают, что существуют по крайней мере два других вида Entamoeba , которые выглядят так же у человека - E. moshkovski. и Энтамеба бангладешская . Причина, по которой эти виды до недавнего времени не были дифференцированы, заключается в том, что они полагались на внешний вид. [ 32 ]
Галерея
[ редактировать ]-

-
 Арселла сп. тест (Лобоза: Tubulinea)
Арселла сп. тест (Лобоза: Tubulinea) -
 Акантамеба сп. (Лобоса: Дискозеа )
Акантамеба сп. (Лобоса: Дискозеа ) -
 Текамеба сп. (Лобоса: Дискозеа)
Текамеба сп. (Лобоса: Дискозеа) -

-
 Pelomyxa palustris (Conosa: Archamoebae)
Pelomyxa palustris (Conosa: Archamoebae) -
 Стемонит сп. (Коноза: Миксогастрия )
Стемонит сп. (Коноза: Миксогастрия ) -
 Диктиостелиум дискоидеум (Conosa: Dictyostelia )
Диктиостелиум дискоидеум (Conosa: Dictyostelia )








Примечания
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б Портер С.М., Мейстерфельд Р., Нолл А.Х. (май 2003 г.). «Микрокаменелости в форме вазы из неопротерозойской группы чуар, Гранд-Каньон: классификация, основанная на современных раковинных амеб» (PDF) . Журнал палеонтологии . 77 (3): 409–429. doi : 10.1666/0022-3360(2003)077<0409:VMFTNC>2.0.CO;2 .
- ^ Парфри Л.В., Лар DJ, Нолл А.Х., Кац Л.А. (август 2011 г.). «Оценка времени ранней диверсификации эукариот с помощью мультигенных молекулярных часов» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (33): 13624–9. Бибкод : 2011PNAS..10813624P . дои : 10.1073/pnas.1110633108 . ПМК 3158185 . ПМИД 21810989 .
- ^ Перейти обратно: а б Кавалер-Смит Т. (август 1998 г.). «Пересмотренная система жизни шести королевств». Биологические обзоры Кембриджского философского общества . 73 (3): 203–66. дои : 10.1111/j.1469-185X.1998.tb00030.x . ПМИД 9809012 . S2CID 6557779 .
- ^ Адл С.М., Басс Д., Лейн CE, Лукеш Дж., Шох К.Л., Смирнов А., Агата С., Берни С., Браун М.В., Бурки Ф., Карденас П., Чепичка И., Чистякова Л., дель Кампо Дж., Данторн М., Эдвардсен Б., Эглит. Ю, Гийу Л., Хампл В., Хейсс А.А., Хоппенрат М., Джеймс Тай, Карнковска А., Карпов С., Ким Е., Колиско М., Кудрявцев А., Лар DJG, Лара Е., Ле Галл Л., Линн Д.Х., Манн Д.Г., Массана Р. , Митчелл И.А.Д., Морроу С., Парк Дж.С., Павловски Дж.В., Пауэлл М.Дж., Рихтер Д.Д., Рюкерт С., Шедвик Л., Шимано С., Шпигель Ф.В., Торруэлла Г., Юсеф Н., Златогурский В., Чжан К. (2019). «Пересмотр классификации, номенклатуры и разнообразия эукариот» . Журнал эукариотической микробиологии . 66 (1): 4–119. дои : 10.1111/jeu.12691 . ПМК 6492006 . ПМИД 30257078 .
- ^ Перейти обратно: а б с Канг С., Тайс А.К., Шпигель Ф.В., Зильберман Дж.Д., Панек Т., Чепичка И. и др. (сентябрь 2017 г.). «Между капсулой и тяжелым испытанием: глубокая эволюция амеб» . Молекулярная биология и эволюция . 34 (9): 2258–2270. дои : 10.1093/molbev/msx162 . ПМК 5850466 . ПМИД 28505375 .
- ^ Перейти обратно: а б с д и Адл С.М., Симпсон А.Г., Лейн С.Э., Лукеш Дж., Басс Д., Баузер С.С. и др. (сентябрь 2012 г.). «Пересмотренная классификация эукариот» . Журнал эукариотической микробиологии . 59 (5): 429–93. дои : 10.1111/j.1550-7408.2012.00644.x . ПМЦ 3483872 . ПМИД 23020233 .
- ^ Перейти обратно: а б Павловски Дж., Аудик С., Адл С., Басс Д., Белбахри Л., Берни С. и др. (6 ноября 2012 г.). «Рабочая группа CBOL по протистам: штрих-кодирование эукариотического богатства за пределами царств животных, растений и грибов» . ПЛОС Биология . 10 (11): e1001419. дои : 10.1371/journal.pbio.1001419 . ПМК 3491025 . ПМИД 23139639 .
- ^ Перейти обратно: а б с Кавальер-Смит Т., Фиоре-Донно А.М., Чао Э., Кудрявцев А., Берни С., Снелл Э.А., Льюис Р. (февраль 2015 г.). «Мультигенная филогения решает проблему глубокого разветвления Amoebozoa» . Молекулярная филогенетика и эволюция . 83 : 293–304. дои : 10.1016/j.ympev.2014.08.011 . ПМИД 25150787 .
- ^ Корлисс Дж.О. (1984). «Королевство Протиста и его 45 типов». БиоСистемы . 17 (2): 87–126. дои : 10.1016/0303-2647(84)90003-0 . ПМИД 6395918 .
- ^ Перейти обратно: а б Кавалер-Смит Т. (2003). «Филогения протистов и классификация простейших высокого уровня». Европейский журнал протистологии . 39 (4): 338–348. дои : 10.1078/0932-4739-00002 .
- ^ Руджеро, Майкл А.; Гордон, Деннис П.; Оррелл, Томас М.; Байи, Николя; Бургуэн, Тьерри; Бруска, Ричард К.; Кавалер-Смит, Томас; Гири, Майкл Д.; Кирк, Пол М. (11 июня 2015 г.). «Коррекция: классификация всех живых организмов более высокого уровня» . ПЛОС ОДИН . 10 (6): e0130114. дои : 10.1371/journal.pone.0130114 . ISSN 1932-6203 . ПМК 5159126 . ПМИД 26068874 .
- ^ Перейти обратно: а б Дерелле Р., Торруэлла Г., Климеш В., Бринкманн Х., Ким Э., Влчек Ч., Ланг Б.Ф., Элиаш М. (февраль 2015 г.). «Бактериальные белки определяют один эукариотический корень» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (7): E693–9. Бибкод : 2015PNAS..112E.693D . дои : 10.1073/pnas.1420657112 . ПМЦ 4343179 . ПМИД 25646484 .
- ^ Жулидов Д.А., Робартс Р.Д., Жулидов А.В., Жулидова О.В., Маркелов Д.А., Русанов В.А., Хедли Дж.В. (2002). «Накопление цинка слизевиком Fuligo septica (L.) Wiggers в бывшем Советском Союзе и Северной Корее». Журнал качества окружающей среды . 31 (3): 1038–42. дои : 10.2134/jeq2002.1038 . ПМИД 12026071 .
- ^ Чон К.В. (1973). Биология амебы . Нью-Йорк: Академическая пресса. стр. 100 . ISBN 978-0-323-14404-9 .
- ^ Хосина, Ре; Цуки, Юдзи; Харумото, Теруэ; Сузаки, Тошинобу (февраль 2021 г.). «Характеристика зеленого Стентора с симбиотическими водорослями, растущими в чрезвычайно олиготрофной среде и хранящими в своей цитоплазме большое количество крахмальных гранул» . Научные отчеты . 11 (2865). дои : 10.1038/s41598-021-82416-9 . ПМЦ 7859197 . ПМИД 33536497 .
- ^ Вайнер, Агнес К.М.; Каллисон, Билли; Дата, Шайлеш В.; Тымл, Томаш; Волланд, Жан-Мари; Войке, Таня; Кац, Лаура А.; Слейт, Робин С. (февраль 2022 г.). «Изучение связи между раковинной амебой Hyalosphenia papilio (Arcellinida, Amoebozoa) и связанными с ней внутриклеточными микроводорослями с использованием молекулярных и микроскопических методов» . Протист . 173 (1): 125853. doi : 10.1016/j.protis.2021.125853 . ПМЦ 9148389 . ПМИД 35030517 .
- ^ Эйхингер Л., Пачебат Дж.А., Глёкнер Г., Раджандрим М.А., Сукганг Р., Берриман М. и др. (май 2005 г.). «Геном социальной амебы Dictyostelium discoideum» . Природа . 435 (7038): 43–57. Бибкод : 2005Natur.435...43E . дои : 10.1038/nature03481 . ПМЦ 1352341 . ПМИД 15875012 .
- ^ Докинз, Ричард ; Вонг, Ян (2016). Рассказ предка . Хоутон Миффлин Харкорт. ISBN 978-0544859937 .
- ^ Кавалер-Смит Т., Чао Э.Э. (декабрь 1996 г.). «Молекулярная филогения свободноживущих архезоа Trepomonas agilis и природа первых эукариот». Журнал молекулярной эволюции . 43 (6): 551–62. Бибкод : 1996JMolE..43..551C . дои : 10.1007/BF02202103 . ПМИД 8995052 . S2CID 28992966 .
- ^ Кавальер-Смит Т., Чао Э.Э., Льюис Р. (июнь 2016 г.). «Филогения 187 генов простейших типа Amoebozoa открывает новый класс (Cutosea) глубоко разветвленных, ультраструктурно уникальных, покрытых оболочкой морских лобоз и проясняет эволюцию амеб» . Молекулярная филогенетика и эволюция . 99 : 275–296. дои : 10.1016/j.ympev.2016.03.023 . ПМИД 27001604 .
- ^ Силар П. (2016). «Эукариотические протисты: происхождение, эволюция и биология эукариотических микробов» (PDF) . HAL открывает архивы . Креативное сообщество. стр. 1–462. ISBN 978-2-9555841-0-1 .
- ^ Панек Т., Задробилкова Е., Уокер Г., Браун М.В., Гентекаки Е., Грудова М. и др. (май 2016 г.). «Первый мультигенный анализ Archamoebae (Amoebozoa: Conosa) достоверно раскрывает их филогению и показывает, что Entamoebidae представляют собой глубокую линию этой группы» . Молекулярная филогенетика и эволюция . 98 : 41–51. дои : 10.1016/j.ympev.2016.01.011 . ПМИД 26826602 .
- ^ Леонтьев Дмитрий Владимирович; Шниттлер, Мартин; Стивенсон, Стивен Л.; Новожилов Юрий К.; Щепин, Олег Н. (март 2019 г.). «К филогенетической классификации миксомицетов». Фитотаксы . 399 (3): 209–238. дои : 10.11646/phytotaxa.399.3.5 . S2CID 108783142 .
- ^ Виджаявардене, Налин; Хайд, Кевин; Аль-Ани, ЛКТ; Долатабади, С; Стадлер, Марк; Хээлуотерс, Дэнни; и др. (2020). «Очерк грибов и грибоподобных таксонов» . Микосфера . 11 : 1060–1456. дои : 10.5943/микосфера/11/1/8 . hdl : 11336/151990 .
- ^ Портер С.М. (2006). «Протерозойская летопись окаменелостей гетеротрофных эукариот». В Сяо С., Кауфман А.Дж. (ред.). Неопротерозойская геолобиология и палеобиология . Темы геобиологии. Том. 27. Дордрехт, Нидерланды: Springer. стр. 1–21. дои : 10.1007/1-4020-5202-2 . ISBN 978-1-4020-5201-9 .
- ^ Хан Н.А., Сиддики Р. (июнь 2015 г.). «Есть ли свидетельства полового размножения (мейоза) у акантамебы?» . Патогены и глобальное здоровье . 109 (4): 193–5. дои : 10.1179/2047773215Y.0000000009 . ПМЦ 4530557 . ПМИД 25800982 .
- ^ Перейти обратно: а б с д Келсо А.А., Сэй А.Ф., Шарма Д., Ледфорд Л.Л., Терчик А., Саски К.А., Кинг А.В., Аттауэй CC, Темесвари Л.А., Сехорн М.Г. (2015). «Entamoeba histolytica Dmc1 катализирует гомологическое спаривание ДНК и обмен цепей, стимулируемый кальцием и Hop2-Mnd1» . ПЛОС ОДИН . 10 (9): e0139399. Бибкод : 2015PLoSO..1039399K . дои : 10.1371/journal.pone.0139399 . ПМК 4589404 . ПМИД 26422142 .
- ^ Перейти обратно: а б Сингх Н., Бхаттачарья А., Бхаттачарья С. (2013). «Гомологичная рекомбинация происходит у Entamoeba и усиливается во время стресса роста и конверсии стадий» . ПЛОС ОДИН . 8 (9): е74465. Бибкод : 2013PLoSO...874465S . дои : 10.1371/journal.pone.0074465 . ПМК 3787063 . ПМИД 24098652 .
- ^ Хофстаттер П.Г., Браун М.В., Лар DJG (ноябрь 2018 г.). «Сравнительная геномика поддерживает пол и мейоз у разнообразных амебозоа» . Геном Биол Эвол . 10 (11): 3118–3128. дои : 10.1093/gbe/evy241 . ПМК 6263441 . ПМИД 30380054 .
- ^ «Энтамебиаз – MeSH – NCBI» . www.ncbi.nlm.nih.gov . Проверено 21 июля 2015 г.
- ^ «Энтамебиаз» . сетка.киб.ки.се . Архивировано из оригинала 22 июля 2015 г. Проверено 21 июля 2015 г.
- ^ Перейти обратно: а б с д Фаррар Дж., Хотез П., Юнгханс Т., Канг Дж., Лаллоо Д., Уайт, Нью-Джерси (26 октября 2013 г.). Тропические болезни Мэнсона . Elsevier Науки о здоровье. стр. 664–671. ISBN 978-0-7020-5306-1 .
- ^ Бичинг Н., Гилл Дж. (17 апреля 2014 г.). «19». Конспект лекций: Тропическая медицина . Джон Уайли и сыновья. стр. 177–182. ISBN 978-1-118-73456-8 .
Дальнейшее чтение
[ редактировать ]- Шильде С., Шаап П. (2013). «Амебозоа». Протоколы Dictyostelium discoideum . Методы молекулярной биологии. Том. 983. стр. 1–15. дои : 10.1007/978-1-62703-302-2_1 . ISBN 978-1-62703-301-5 . ПМК 4037983 . ПМИД 23494299 .
Внешние ссылки
[ редактировать ]- Павловский Дж. «Молекулярная филогения амебоидных простейших - дерево амебозоа» . Группа молекулярной систематики (MSG) . Кафедра зоологии и биологии животных Женевского университета. Архивировано из оригинала 2 января 2005 года.
- Килинг П., Леандер Б.С., Симпсон А. «Древо жизни эукариотов» . Проект «Древо жизни».
- Лейди Дж (1879). «Тарелочки амебы» . Пресноводные корненожки Северной Америки . Вашингтон, округ Колумбия: Государственная типография.
- Амебозоа в приложении UniEuk Taxonomy.
| Базы данных органов управления : Национальные |
|---|