Вампиреллида
| Вампиреллида | |
|---|---|
| |
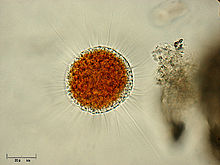
| Вампирелла латериция | |
| Научная классификация | |
| Домен: | Эукариоты |
| Клэйд : | потогонные средства |
| Клэйд : | САР |
| Тип: | Эндомикса |
| Сорт: | Идея вампира Кавалер-Смит 2018 [ 2 ] |
| Заказ: | Вампиреллида Запад 1901 года, ред. Хесс и др. 2012 год [ 1 ] |
| Клады [ 3 ] | |
| Разнообразие [ 3 ] | |
| 48 видов | |
| Синонимы [ 4 ] | |
|
Акончулинида Де Саделеер 1934 г. | |
Вампиреллиды отряд ( классифицированных Vampyrellida , класс Vampyrellidea ), в просторечии известные как вампирские амебы , представляют собой группу свободноживущих хищных амеб, как часть линии Endomyxa . От других групп амеб их отличает неправильная форма клеток со склонностью к слиянию и расщеплению, как у плазмодий , а также их жизненный цикл со стадией пищеварительной кисты , которая переваривает собранную пищу. Они появляются по всему миру в морских , солоноватых , пресноводных и почвенных средах обитания. Они являются важными хищниками огромного разнообразия микроскопических организмов, от водорослей до грибов и животных . [ 3 ] Они также известны как аконхулинидные амебы (отряд Aconchulinida ). [ 4 ]
Морфология и движение клеток
[ редактировать ]Вампиреллиды традиционно считаются нитевидными амебами, т.е. они образуют тонкие псевдоподии ( филоподии ). Они голые, лишены внешних структур, таких как чешуя, клеточные оболочки или гликокаликс может присутствовать временная слизистая , хотя на стадии трофозоита оболочка . Трофозоиты сильно различаются по форме, размеру и цвету у разных видов, но их можно сгруппировать в три клеточных состояния или «морфотипа»: изодиаметрический, расширенный и «филофлабеллятный». [ 1 ] [ 3 ]
- Изодиаметрический (сферический) морфотип, распространенный у альгиворных Vampyrella и Lateromyxa , с радиальными филоподиями. Некоторые виды плавают в толще воды, напоминая солнечники по форме . Другие ползают по поверхности, концентрируя жесткие филоподии в передней части клетки, прикрепляя их к поверхности, втягивая и перемещая их в заднюю область. [ 3 ]
- Расширенный морфотип, наиболее распространенный, прикрепленный к поверхности, с разнообразной формой (например, веерообразной или разветвленной у Leptophrys ; с крупными, гиалиновыми пластинками с нитевидными филоподиями у Sericomyxa ; сильно разветвленными или сетчатыми у Platyreta). и талассомикса ). [ 3 ]
- Филофлабеллятный морфотип, обнаруженный только у Placopus , с уплощенными эллиптическими, сферическими или веерообразными клетками, которые демонстрируют четкое разделение между горбом гранулоплазматических клеток и гиалоплазматическими пластинками, иногда называемыми « ламеллиподиями ». На вентральной стороне клетки имеются многочисленные филоподии. Некоторые из этих трофозоитов напоминают амебозоев, таких как ванеллиды , за исключением наличия филоподий. Они передвигаются, перекатываясь по филоподиям, прикрепленным к субстрату. [ 3 ]

Жизненный цикл
[ редактировать ]


Этапы питания
[ редактировать ]Все известные вампиреллиды — гетеротрофные амебы со свободноживущим (непаразитарным ) жизненным циклом , лишенным жгутиковых стадий, за исключением Lateromyxa Gallica , и характеризующимся чередованием подвижных и неподвижных клеточных стадий: [ 3 ]
- Подвижные амебоидные клетки, называемые « трофозоитами » или «роевиками». [ а ] в старой литературе. Их основная деятельность — рассеивание, поиск и сбор пищи посредством фагоцитоза . [ 3 ]
- Неподвижная, но высоко метаболически активная стадия «пищеварительной кисты », возникающая после кормления. У некоторых видов это называется «фазой покоя», но она отличается от настоящей покоящейся кисты (или споры ), которая метаболически неактивна, чтобы выжить в неблагоприятных условиях. Чтобы достичь этой стадии, трофозоит втягивает свои филоподии , выделяет слоистую клеточную стенку и прочно прикрепляется к субстрату или свободно плавает. либо центральная главная вакуоль По-видимому, переваривает пищу , либо несколько отдельных вакуолей. Цвет цитоплазмы может измениться на ярко-красный, оранжевый или желтый цвет или остаться бесцветным. Когда фаза пищеварения завершается, один или несколько трофозоитов вылупляются из кисты через отверстия в клеточной стенке. [ 3 ]
Воспроизведение
[ редактировать ]У некоторых видов, ближе к концу стадии пищеварительной кисты, бесполое размножение внутри кисты происходит посредством деления клеток (так называемая «внутренняя плазмотомия »), в результате чего образуются 2–4 дочерние клетки . Эти клетки высвобождаются через отверстия в виде молодых трофозоитов. Другие виды не делятся внутри закрытой кисты, а делятся во время или после процесса вылупления («внешняя плазмотомия»). Lateromyxa Gallica демонстрирует необычный способ размножения: питаясь внутренней частью клеток водоросли , плазмодии выделяются и развиваются в пищеварительные кисты. [ 3 ]
недостаточно Доказательств полового размножения вампиреллид , за исключением некоторых мейотических стадий в покоящихся цистах, выявленных у Lateromyxa Gallica посредством ультраструктурных исследований . [ 5 ]
Плазмодиальное поведение
[ редактировать ]Многие виды вампиреллид имеют более одного ядра и ведут себя как плазмодии . Они могут сливать свои клетки при контакте и разделяться при движении в противоположных направлениях. Некоторые виды легко выращивают плазмодии размером с чашку Петри в лабораторных условиях, в то время как другие сливаются только при высокой плотности клеток и низкой доступности пищи. Неизвестно, в какой степени это может произойти в естественной среде. Напротив, виды Placopus редко когда-либо встречаются с более чем двумя ядрами. [ 3 ]
Стадии отдыха
[ редактировать ]В неблагоприятных условиях внешней среды вампиреллиды могут переходить в несколько типов стадий покоя: [ 3 ]
- Гипноцисты, тонкостенные и лишенные пищевого содержания, образуются при нарушении трофозоита внешним стрессом. [ 3 ]
- Вторичные цисты, тонкостенные и лишенные пищевого содержимого, образовались в результате голодания. [ 3 ]
- Настоящие кисты покоя, также называемые в старой литературе «спороцистами» или « спорами ». Они образуются в природных образцах и старых культурах, когда нет пищи или условия неблагоприятны. Они строят несколько стенок кисты и уплотняют их клеточное содержимое. Они могут пережить высыхание или замерзание как минимум до трех лет. [ 3 ]
Экология
[ редактировать ]Распределение
[ редактировать ]Вампиреллиды имеют космополитическое распространение : они встречаются на всех континентах, кроме Антарктиды и всех морских экосистем. Они населяют широкий спектр морских , солоноватых и пресноводных местообитаний и часто выделяются из почвы . образцов [ 3 ] Морские экосистемы отличаются удивительно высоким разнообразием. [ 6 ] и они встречаются в основном в донных средах обитания (например, приливные бассейны , диатомовые лужайки, связанные с красными водорослями ...). Существует значительная положительная корреляция между разнообразием Vampyrellida и наличием питательных веществ в отложениях. [ 7 ] Согласно данным экологического секвенирования, вампиреллиды колонизируют неотропическую почву. [ 8 ] ледниковые криоконитовые системы, [ 9 ] листья крестоцветных , [ 10 ] Сфагновые – обитаемые торфяники, [ 11 ] гидротермальные отложения [ 6 ] и глубокое море . [ 12 ]
Трофическое разнообразие
[ редактировать ]
Вампиреллиды демонстрируют большое трофическое разнообразие. Они являются хищниками длинного списка организмов разнообразного эволюционного сходства, структуры и размеров, включая хлорофитные и стрептофитные зеленые водоросли, диатомовые , водоросли хризофиты , криптофиты , эвглениды , гетеротрофные жгутиконосцы , реснитчатые цисты , грибковые гифы и споры , дрожжи и даже микрометазоа , такие как как нематоды и яйца коловраток . Бактериальность встречается редко и в основном связана с нитчатыми цианобактериями . Хотя существуют универсальные всеядные хищники, такие как Leptophrys , некоторые виды вампиреллид являются специализированными хищниками; например, альгиворные Vampyrella и Placopus ограничены несколькими видами зеленых водорослей с твердыми стенками, в то время как Arachnomyxa и Planctomyxa предпочитают Volvocales и эвглениды . [ 3 ]
Стратегии кормления
[ редактировать ]Вампиреллиды развили стратегии борьбы с относительно крупной и громоздкой добычей, которую трудно съесть. Они демонстрируют по крайней мере четыре различные стратегии питания, чтобы поглотить всю добычу или поглотить содержимое других эукариотических клеток . Эти стратегии питания не являются взаимоисключающими, и один и тот же вид может демонстрировать разные типы добычи. [ 3 ]
- Бесплатный захват. Подобно амебам других супергрупп , они ловят и заключают добычу в пищевую вакуоль путем обычного фагоцитоза . Некоторые могут парализовать свою жертву еще до того, как ее закроют. Размеры оболочек весьма разнообразны: от многочисленных одновременно мелких клеток до целых нематод или колониальных зеленых водорослей. [ 3 ]
- Вторжение в колонию . Они прикрепляются к колониям вольвокальных водорослей, растворяются и проникают во внеклеточный студенистый слизистый матрикс, а также фагоцитируют отдельные клетки внутри колонии. Возможно, для защиты от хищников, внутри колонии они превращаются в пищеварительные кисты. [ 3 ]
- Извлечение протопластов , самая известная стратегия. Они целенаправленно удаляют, заглатывают и переваривают клеточное содержимое своей добычи, всегда растворяя органическую клеточную стенку жертвы или просто смещая кремниевую стенку жертвы, и проникая через псевдоподии (называемые «каликулоподиями»), чтобы удалить содержимое клетки. Некоторые виды выбрасывают цитоплазму жертвы, оказывая давление - процесс, известный как «плазмоптизис», за которым следует быстрое образование большой вакуоли. Этот процесс напоминает сосательные движения и, вероятно, является причиной их сравнения с вампирами . У морских видов плазмоптизис не наблюдается, что позволяет предположить, что осмотическое давление , обусловленное соленостью . для плазмоптиза важно [ 3 ]
- Инфильтрация добычи . Подобно экстракторам протопластов, они прокалывают клеточную стенку водоросли-жертвы, но проникают в саму клетку и завершают цикл внутри нее. Некоторые из них способны перемещаться латерально от одной клетки к другой у нитчатой добычи. Они делятся на более мелкие части, которые превращаются в пищеварительные кисты. [ 3 ]
История исследований
[ редактировать ]
Вампиреллиды имеют долгую историю исследований. Они известны вампироподобной привычкой питания некоторых амеб-вампиреллид, которые прокалывают клеточные стенки других эукариотических клеток, чтобы питаться именно содержимым клетки, - механизм питания, известный как экстракция протопластов. Это сходство привело к возникновению названия их самого популярного рода, Vampyrella , и их разговорного названия « вампирские амебы ». [ 3 ]
Одним из самых ранних недвусмысленных сообщений о вампиреллидах является описание Amoeba Lateritia (теперь известное как Vampyrella Lateritia ) в середине XIX века немецким ботаником Георгом Фрезениусом . [ 13 ] Первая обширная документация об их истории жизни и пищевом поведении была предоставлена в 1865 году польским протозоологом Леоном Циенковским , который создал род Vampyrella и отнес его к подгруппе «монад». [ 14 ] полифилетическая совокупность паразитоидных простейших . В последующих работах и монографиях описаны многочисленные виды водных вампиреллид с важными наблюдениями за их поведением и экологией. В 1885 году немецкий миколог Вильгельм Цопф продемонстрировал наличие ядер у вампиреллид и выделил первое семейство Vampyrellidae . [ 15 ] [ 16 ] [ 3 ]
В середине 20 века были сделаны первые открытия почвенных вампиреллид. Была создана первая лабораторная культура вампиреллид , содержащая почвенную амебу Theratomyxa weberi, питавшуюся нематодами . Подобные почвенные амебы были выделены позже и изучены как возможная борьба с вредителями против нематод, патогенных для растений. [ 17 ] Другие исследования выявили, что гигантский почвенный вампиреллид является организмом, ответственным за перфорации, обнаруженные в грибных спорах . [ 18 ] [ 3 ]
В начале 1980-х годов процесс питания и жизненный цикл альгиворной пресноводной Vampyrella Lateritia был заснят с непревзойденной детализацией. [ 19 ] [ 20 ] род крупных плазмодийных амеб Thalassomyxa . был обнаружен В то же время в морских водах отдаленных уголков мира [ 21 ]
До проведения генетического анализа таксономическое размещение вампиреллид было затруднено: их считали родственниками миксомицетовых слизевиков . [ 16 ] гелиозоа , [ 22 ] протеомиксиды , [ 23 ] нитевидные корненожки [ 24 ] и даже Монера . [ 25 ] В 2009 году загадка была раскрыта благодаря филогении генов 18S рибосомальной РНК , что сделало вампиреллид частью Rhizaria . [ 26 ] Пересмотренная таксономия в 2012 году воссоздала отряд Vampyrellida . [ 1 ] В 2013 году было обнаружено огромное неожиданное разнообразие морских вампиреллид. [ 6 ] [ 3 ]
Эволюция и систематика
[ редактировать ]Внешние отношения
[ редактировать ]Vampyrellida представляет собой одну из основных групп свободноживущих амеб , филогенетически отделенную от других групп амеб, таких как Amoebozoa , Heterolobosea и Nucleariidae . Вместо этого Vampyrellida представляет собой изолированную кладу внутри Rhizaria супергруппы . [ 26 ] Они являются ближайшими родственниками Phytomyxea , паразитов растений и которые, в отличие от вампиреллид, в течение своего жизненного цикла рассеиваются через жгутиковые стадии и водорослей , проводят большую часть своей активной жизни внутри клеток-хозяев. [ 3 ] Современные классификации относят Vampyrellida и Phytomyxea вместе с другими небольшими группами Rhizaria к типу Endomyxa . [ 4 ] Несколько филогенетических анализов выявили родство сестринской группы между Vampyrellida и Phytomyxea и дали их кладе название Proteomyxia. [ 2 ] или Фиториза . [ 27 ]
Внутренняя классификация
[ редактировать ]| Филогения вампиреллид | ||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||
| Филогения Vampyrellida, опубликованная в 2023 году, основана на рРНК SSU последовательностях гена . [ 28 ] Линии B1, B2 и B4 представляют собой клады, которые содержат только последовательности ДНК окружающей среды , без описанных видов. [ 3 ] |
В настоящее время существует 48 заслуживающих доверия видов вампиреллид, распределенных в 10 родах , разбросанных по пяти хорошо устоявшимся кладам, обнаруженным с помощью генетических данных , четыре из которых являются семействами . Несмотря на достижения, большая часть разнообразия вампиреллид все еще неизвестна или неописана. [ 3 ]
- Семейство Vampyrellidae Zopf 1885, исправлено. Гесс, Саусен и Мелконян 2012 г. [ 1 ]
- Vampyrella Cienkowski 1865 (19 видов)
- Семейство Leptophryidae Hess, Sausen & Melkonian 2012. [ 1 ]
- Arachnomyxa Hess 2017 (1 вид)
- Leptophrys Hertwig & Lesser 1874 (3 вида)
- Planctomyxa Hess 2017 (1 вид)
- Platyreta Cavalier-Smith & Bass 2008 (1 вид)
- Псевдовампирелла Suthaus & Hess 2023 (2 вида) [ 28 ]
- Theratromyxa Zwillenberg 1952 (1 вид)
- Верналофрис Гонг и др. 2015 г. (1 вид)
- Семейство Placopodidae Jahn 1928 = Hyalodiscidae Poche 1913 = линия B3.
- Семейство Sericomyxidae More, Simpson & Hess, 2021 г.
- Sericomyxa More, Simpson & Hess 2021 (1 вид)
- « Клада Thalassomyxa » = линия B5
- Thalassomyxa Grell 1985 (3 вида)
- Роды, которые, вероятно, принадлежат к Vampyrellida, но для которых нет генетических данных и поэтому считаются incertae sedis :
- Арахнула Ценковски 1856 (2 вида)
- Asterocaelum Canter 1973 (1 вид)
- Гобиелла Циенковски 1881 (2 вида)
- Lateromyxa Hülsmann 1993 (1 вид)
- Монадопсис Кляйн 1993 (1 вид)
- Пенардия Кэш 1904 г. (2 вида)
Следующие таксоны были связаны с Vampyrellida, но их местонахождение неясно или могло не принадлежать к этой группе. [ 3 ]
- Гиалодиск ангеловика Сойер 1975 г.
- Hyalodiscus ceruleus Шеффер 1926 г.
- Гиалодиск листовидный Лепси 1931 г.
- Hyalodiscus korotnewi Мерешовский 1879 г.
- Гиалодиск макронуклеус Лепси 1931
- Гиалодиск минор Лепси 1960 г.
- Hyalodiscus placopus Hülssmann 1974 [не Paragocevia placopus (Page & Willumsen 1980) Страница 1987 ]
- Hyalodiscus simplex Вольфарт-Боттерманн 1977 г.
- Vampyrella chaetoceratis (Paulsen 1910) Ostenfeld 1913 [ Apodinium chaetoceratis Paulsen 1910 ; Paulsenella chaetoceratis (Paulsen 1910) Chatton 1920 ]
- Vampyrella лабиринтулоидес (Арчер 1875) Валканов 1940 [ Chlamydomyxa лабиринтулоидес Арчер 1875 ]
- Vampyrella montana (Ланкестер 1896) Валканов 1940 [ Chlamydomyxa montana Lankester 1896 ]
- Vampyrellidium perforans Surek & Melkonian 1980 ( нуклеарная амеба)
- Vampyrellidium vagans Zopf 1885 ( нуклеарная амеба)
- Vampyrellidium roseus (Trichense 1885) Schepotieff 1912 [ Protogenes roseus Trichense 1885 ]
- Вампирина бючли Френцель 1897 г.
Примечания
[ редактировать ]- ^ Термин «роевики» более специально используется для реснитчатых плавающих клеток, таких как гаметы , поэтому трофозоиты . более подходящим термином является [ 3 ]
- ^ Не путать с родом диатомовых водорослей Hyalodiscus Ehrenberg 1845 отдает предпочтение , который в биологической номенклатуре описанию ранее. [ 3 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и Хесс С., Саусен Н., Мелконян М. (2012). «Проливая свет на вампиров: новый взгляд на филогению амеб вампиреллид» . ПЛОС ОДИН . 7 (2): e31165. Бибкод : 2012PLoSO...731165H . дои : 10.1371/journal.pone.0031165 . ПМК 3280292 . ПМИД 22355342 .
- ^ Jump up to: а б Кавалер-Смит, Томас; Э. Чао, Эма; Льюис, Родри (2018), «Мультигенная филогения и клеточная эволюция хромистского инфракоцарства Rhizaria: контрастирующая клеточная организация сестринских типов Cercozoa и Retaria», Protoplasma , 255 (5): 1517–1574, doi : 10.1007/s00709-018-1241- 1 , ЧВК 6133090 , ПМИД 29666938
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в Гесс С., Зутхаус А. (2022). «Амебы-вампиреллиды (Vampyrellida, Rhizaria)» . Протист . 173 (1): 125854. doi : 10.1016/j.protis.2021.125854 . ПМИД 35091168 .
- ^ Jump up to: а б с Адл С.М., Басс Д., Лейн CE, Лукеш Дж., Шох К.Л., Смирнов А., Агата С., Берни С., Браун М.В., Бурки Ф., Карденас П., Чепичка И., Чистякова Л., дель Кампо Дж., Данторн М., Эдвардсен Б., Эглит. Ю, Гийу Л., Хампл В., Хейсс А.А., Хоппенрат М., Джеймс Тай, Карнковска А, Карпов С, Ким Э, Колиско М, Кудрявцев А, Лар DJG, Лара Е, Ле Галл Л, Линн Д.Х., Манн Д.Г., Массана Р., Митчелл И.Д., Морроу С., Парк Дж.С., Павловски Дж.В., Пауэлл М.Дж., Рихтер Д.Д. , Рюкерт С, Шедвик Л, Шимано С, Шпигель Ф.В., Торруэлла Г, Юсеф Н, Златогурский В., Чжан Ц (2019). «Пересмотр классификации, номенклатуры и разнообразия эукариот» . Журнал эукариотической микробиологии . 66 (1): 4–119. дои : 10.1111/jeu.12691 . ПМК 6492006 . ПМИД 30257078 .
- ^ Репсторф П., Хюльсманн Н., Хаусманн К. (1993). «Кариологические исследования филозной амебы вампиреллид Lateromyxa Gallica Hülsmann 1993». Европейский журнал протистологии . 29 (3): 302–310. дои : 10.1016/S0932-4739(11)80373-1 . ПМИД 23195644 .
- ^ Jump up to: а б с Берни С., Ромак С., Маэ Ф., Сантини С., Сиано Р., Басс Д. (2013). «Вампиры в океанах: хищные церкозойные амебы в морской среде обитания» . ИСМЕ Дж . 7 (12): 2387–2399. дои : 10.1038/ismej.2013.116 . ПМЦ 3834849 . ПМИД 23864128 .
- ^ Сун Х, Чжан Ю, Ленг Х, Чжан Ю, Ю Ю (2015). «Аконхулинидные раковинные амебы играют важную роль в круговороте питательных веществ в пресноводных экосистемах». Биология и биохимия почвы . 82 : 1–7. doi : 10.1016/j.soilbio.2014.11.009 .
- ^ Лентенду Дж., Маэ Ф., Басс Д., Рюкерт С., Сток Т., Данторн М. (2018). «Постоянные закономерности высокого альфа- и низкого бета-разнообразия тропических паразитических и свободноживущих простейших» . Молекулярная экология . 27 (13): 2846–2857. дои : 10.1111/mec.14731 . ПМИД 29851187 . S2CID 44119381 .
- ^ Вимеркати Л., Дарси Дж.Л., Шмидт С.К. (2019). «Исчезающая перигляциальная экосистема на вершине горы Килиманджаро поддерживает как космополитические, так и эндемичные микробные сообщества» . Научные отчеты . 9 (1): 10676. Бибкод : 2019НатСР...910676В . дои : 10.1038/s41598-019-46521-0 . ПМК 6650471 . ПМИД 31337772 .
- ^ Плох С., Роуз Л.Е., Басс Д., Бонковски М. (2016). «Высокое разнообразие выявлено у связанных с листьями протистов (Rhizaria: Cercozoa) Brassicaceae» . Журнал эукариотической микробиологии . 63 (5): 635–641. дои : 10.1111/jeu.12314 . ПМК 5031217 . ПМИД 27005328 .
- ^ Лара Э., Митчелл И.Д., Морейра Д., Гарсиа П.Л. (2011). «Очень разнообразное и сезонно динамичное сообщество протистов в нетронутом торфянике» (PDF) . Протист . 162 (1): 14–32. дои : 10.1016/j.protis.2010.05.003 . ПМИД 20692868 .
- ^ Шенле А., Хольфельд М., Херманс К., Маэ Ф., де Варгас К., Нитше Ф., Арндт Х. (2021). «Высокое и специфическое разнообразие простейших в глубоководных котловинах с доминированием диплонемид, кинетопластид, инфузорий и фораминифер» . Коммуникационная биология . 4 (1): 501. дои : 10.1038/s42003-021-02012-5 . ПМК 8065057 . ПМИД 33893386 .
- ^ Фрезениус, Георг (1858). «Вклад в знание микроскопических организмов». Трактаты Зенкенбергского общества естественных исследований (на немецком языке). 2 : 218–219, 241–242.
- ^ Ценковский, Лев (1865). «Вклад в знание монад» . Архив микроскопической анатомии (на немецком языке). 1 :203-232. дои : 10.1007/BF02961414 . S2CID 84323025 .
- ^ Цопф В. (1885). «Грибные животные или слизевики». В Шенк А (ред.). ) наук Справочник по ботанике (Энциклопедия естественных (на немецком языке). Том 3. Трюендт, Бреслау. стр. 1–174.
- ^ Jump up to: а б Цопф В. (1885). По морфологии и биологии низших грибных животных (монадинов), одновременно вклад в фитопатологию [ По морфологии и биологии низших грибных животных (монадинов), одновременно вклад в фитопатологию ] (на немецком языке). Лейпциг: Veit и Comp. дои : 10.5962/bhl.title.945 .
- ^ Уинслоу Р.Д., Уильямс Т.Д. (1957). «Амебоидные организмы, атакующие личинки картофельного корневого нематода ( Heterodera rostochiensis Woll.) в Англии и свекловичного нематода ( H. schachtii Schm.) в Канаде». Журнал завода Зиктен 63 (5): 242–243. дои : 10.1007/BF01988794 . S2CID 21473045 .
- ^ Старый К.М., Дарбишир Дж.Ф. (1978). «Почвенные грибы как пища гигантским амебам». Биология и биохимия почвы . 10 (2): 93–100. дои : 10.1016/0038-0717(78)90077-9 .
- ^ Хюльсманн Н (1983). «О проникновении клеточных стенок водорослей при нападении вампиреллид». Журнал протозоологии . 30 : А50.
- ^ Хюльсманн Н (1985). «Развитие и особенности питания Vampyrella Lateritia Rhizopoda) » ( . Публикация научных Фильм, Сект. биол., Сер. 17, № 16/С . 17 :1–23.
- ^ Грелль К.Г. (1994). «Thalassomyxa canariensis n. sp. из приливного бассейна Тенерифе (Канарские острова)». Арка Протистенкд . 144 (3): 319–324. дои : 10.1016/S0003-9365(11)80146-6 .
- ^ Леблих А.Р., Леблих Л.А. «Номенклатура и таксономическое положение Pseudosporidae, Vampyrellidae и Acinetactidae». Процедура Биол Соц Стирка . 78 : 115–120.
- ^ Хонигберг Б.М., Баламут В., Бове Э.К., Корлисс Дж.О., Гойдичс М., Холл Р.П., Кудо Р.Р., Левин Н.Д., Леблих А.Р., Вайзер Дж., Венрих Д.Х. (1964). «Пересмотренная классификация типов простейших». Журнал протозоологии . 11 (1): 7–2 дои : 10.1111/j.1550-7408.1964.tb01715.x . ПМИД 14119564 .
- ^ Пейдж ФК (1987). «Классификация «голых» амеб (Phylum Rhizopada)». Архив для защиты . 133 (3–4): 199–217. дои : 10.1016/S0003-9365(87)80053-2 .
- ^ Геккель, Эрнст (1869). «Исследования Монеры и других протистов вместе с речью о ходе развития и функциях зоологии». Журнал Jena (на немецком языке). 5 . Лейпциг: Энгельманн: 353.
- ^ Jump up to: а б Басс Д., Чао Э.Э., Николаев С., Ябуки А., Исида К.И., Берни С., Пакзад У., Вилезич С., Кавальер-Смит Т. (2009). «Филогения новых голых Filose и Reticulose Cercozoa: Granofilosea cl. n. и Proteomyxidea Revised». Протист . 160 (1): 75–109. дои : 10.1016/j.protis.2008.07.002 . ISSN 1434-4610 . ПМИД 18952499 .
- ^ Сьерра Р., Каньяс-Дуарте С.Х., Бурки Ф., Швельм А., Фогельквист Дж., Дикселиус К., Гонсалес-Гарсия Л.Н., Джайл Г.Х., Сламовиц CH, Клопп С., Рестрепо С., Арзул И., Павловски Дж. (апрель 2016 г.). «Эволюционное происхождение ризарианских паразитов» . Молекулярная биология и эволюция . 33 (4): 980–983. дои : 10.1093/molbev/msv340 . ПМИД 26681153 .
- ^ Jump up to: а б Сухаус А., Гесс С. (2023). « Pseudovampyrella gen. nov.: Род Vampyrella -подобных экстракторов протопластов находит свое место в Leptophryidae» . Журнал эукариотической микробиологии . 00 . Международное общество протистологов: e13002. дои : 10.1111/jeu.13002 .