Кокколитофор
| Кокколитофор Временной диапазон:
| |
|---|---|

| |
| Кокколитус пелагический | |
| Научная классификация | |
| Домен: | |
| (без рейтинга): | |
| (без рейтинга): | |
| (без рейтинга): | |
| Сорт: | |
| Заказ: | |

Кокколитофориды , или кокколитофориды , — одноклеточные организмы , входящие в состав фитопланктона , автотрофного (самопитающегося) компонента планктонного сообщества. Они образуют группу, насчитывающую около 200 видов, и принадлежат либо к королевству Protista , согласно Уиттакера Роберта системе пяти королевств , либо к кладе Hacrobia , согласно более новой системе биологической классификации. В пределах Hacrobia кокколитофоры относятся к типу или разделу Haptophyta , классу Prymnesiophyceae (или Coccolithophyceae ). почти исключительно морские , фотосинтезирующие и существуют в больших количествах по всей солнечной зоне океана Кокколитофоры .
Кокколитофороры — наиболее продуктивные кальцинирующие организмы на планете, покрытые оболочкой из карбоната кальция, называемой коккосферой . Однако причины их кальцификации остаются неясными. Одной из ключевых функций может быть то, что коккосфера обеспечивает защиту от хищничества микрозоопланктона , который является одной из основных причин гибели фитопланктона в океане. [ 1 ]
Кокколитофоры экологически важны и биогеохимически играют важную роль в морском биологическом насосе и углеродном цикле . [ 2 ] [ 1 ] В зависимости от среды обитания они могут производить до 40 процентов местной первичной морской продукции . [ 3 ] Они представляют особый интерес для тех, кто изучает глобальное изменение климата , поскольку по мере повышения кислотности океана их кокколиты могут стать еще более важными поглотителями углерода . [ 4 ] Стратегии управления используются для предотвращения цветения кокколитофоров, связанного с эвтрофикацией , поскольку это цветение приводит к уменьшению потока питательных веществ в нижние уровни океана. [ 5 ]
Самый распространенный вид кокколитофоры, Emiliania huxleyi , принадлежит к порядку Isochrysidales и семейству Noëlaerhabdaceae . [ 6 ] Встречается в умеренных , субтропических и тропических океанах. [ 7 ] Это делает E. huxleyi важной частью планктонной основы значительной части морских пищевых сетей . Это также самый быстрорастущий кокколитофор в лабораторных культурах. [ 8 ] Его изучают на предмет обширного цветения, которое оно образует в водах, обедненных питательными веществами, после реформирования летнего термоклина . [ 9 ] [ 10 ] и за производство молекул, известных как алкеноны , которые обычно используются учеными-землеведами в качестве средства оценки прошлых температур поверхности моря . [ 11 ]
Обзор
[ редактировать ]Кокколитофориды (или кокколитофориды, от прилагательного [ 12 ] ) образуют группу, насчитывающую около 200 видов фитопланктона. [ 13 ] Они принадлежат либо к королевству Протиста , согласно Уиттакера Роберта классификации Пяти королевств , либо к кладе Хакробия , согласно новой биологической системе классификации. В пределах Hacrobia кокколитофоры относятся к типу или разделу Haptophyta , классу Prymnesiophyceae (или Coccolithophyceae ). [ 6 ] Кокколитофоры отличаются особыми пластинками (или чешуйками) карбоната кальция неопределенной функции, называемыми кокколитами , которые также являются важными микрофоссилиями . Однако существуют виды Prymnesiophyceae, лишенные кокколитов (например, в роде Prymnesium ), поэтому не каждый представитель Prymnesiophyceae является кокколитофором. [ 14 ]
Кокколитофоры — одноклеточный фитопланктон , образующий мелкие чешуйки карбоната кальция (CaCO 3 ) ( кокколиты ), которые покрывают поверхность клеток в виде сферического покрытия, называемого коккосферой . Они были неотъемлемой частью морских планктонных сообществ начиная с юрского периода . [ 15 ] [ 16 ] Сегодня кокколитофоры способствуют ~ 1–10% фиксации неорганического углерода (кальцификации) и общей фиксации углерода (кальцификации плюс фотосинтеза) в поверхностном океане. [ 17 ] и ~50% – пелагическим осадкам CaCO 3 . [ 18 ] Их известковая оболочка увеличивает скорость погружения фотосинтетически связанного CO 2 в глубины океана за счет балластирования органического вещества. [ 19 ] [ 20 ] В то же время биогенное осаждение карбоната кальция при формировании кокколита снижает общую щелочность морской воды и выделяет CO 2 . [ 21 ] [ 22 ] Таким образом, кокколитофоры играют важную роль в морском углеродном цикле , влияя на эффективность биологического углеродного насоса и поглощение океаном атмосферного CO 2 . [ 1 ]
По состоянию на 2021 год неизвестно, почему кокколитофоры кальцифицируются и как их способность производить кокколиты связана с их экологическим успехом. [ 23 ] [ 24 ] [ 25 ] [ 26 ] [ 27 ] Наиболее вероятным преимуществом наличия коккосферы является защита от хищников и вирусов. [ 28 ] [ 26 ] Вирусная инфекция является важной причиной гибели фитопланктона в океанах. [ 29 ] недавно было показано, что кальцификация может влиять на взаимодействие между кокколитофором и его вирусом. [ 30 ] [ 31 ] Основными хищниками морского фитопланктона являются микрозоопланктон, такой как инфузории и динофлагелляты . По оценкам, они потребляют около двух третей первичной продукции океана. [ 32 ] а микрозоопланктон может оказывать сильное пастбищное давление на популяции кокколитофоров. [ 33 ] Хотя кальцификация не предотвращает хищничество, утверждается, что коккосфера снижает эффективность выпаса, затрудняя хищнику использование органического содержимого кокколитофоров. [ 34 ] Гетеротрофные протисты способны выборочно выбирать добычу на основе ее размера или формы, а также с помощью химических сигналов. [ 35 ] [ 36 ] и, таким образом, может отдавать предпочтение другой добыче, которая доступна и не защищена кокколитами. [ 1 ]
Структура
[ редактировать ]
Кокколитофоры представляют собой сферические клетки диаметром около 5–100 микрометров, окруженные известковыми пластинками, называемыми кокколитами , диаметром около 2–25 микрометров. Каждая клетка содержит два коричневых хлоропласта , окружающих ядро . [ 39 ]
коккосфере заключена одна клетка с мембраносвязанными В каждой органеллами . Два крупных хлоропласта с коричневым пигментом расположены по обе стороны клетки и окружают ядро , митохондрии , аппарат Гольджи , эндоплазматическую сеть и другие органеллы. Каждая клетка также имеет две жгутиковые структуры, которые участвуют не только в подвижности, но и в митозе и формировании цитоскелета . [ 40 ] функциональная или рудиментарная гаптонема . У некоторых видов также присутствует [ 41 ] Эта структура, уникальная для гаптофитов , скручивается и разворачивается в ответ на раздражители окружающей среды. Хотя это плохо изучено, было предложено участвовать в захвате добычи. [ 40 ]
Экология
[ редактировать ]Стратегия истории жизни
[ редактировать ]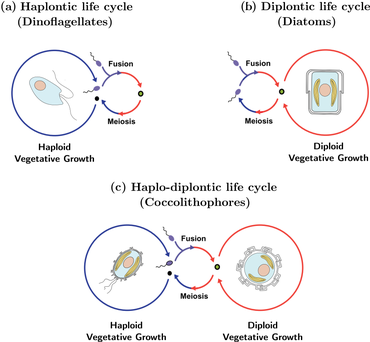
Сложный жизненный цикл кокколитофоров известен как гаплодиплонтный жизненный цикл и характеризуется чередованием как бесполых, так и половых фаз. Бесполая фаза известна как гаплоидная фаза, а половая фаза известна как диплоидная фаза. Во время гаплоидной фазы кокколитофороры производят гаплоидные клетки посредством митоза . Эти гаплоидные клетки затем могут делиться дальше посредством митоза или подвергаться половому размножению с другими гаплоидными клетками. Полученная диплоидная клетка проходит мейоз и снова производит гаплоидные клетки, начиная цикл заново. У кокколитофоров бесполое размножение путем митоза возможно на обеих фазах жизненного цикла, что контрастирует с большинством других организмов, имеющих чередующиеся жизненные циклы. [ 42 ] Как абиотические , так и биотические факторы могут влиять на частоту возникновения каждой фазы. [ 43 ]
Кокколитофоры размножаются бесполым путем путем бинарного деления. В этом процессе кокколиты родительской клетки делятся между двумя дочерними клетками. Были предположения о возможном наличии процесса полового размножения за счет диплоидных стадий кокколитофоров, но этот процесс никогда не наблюдался. [ 44 ]
K или r-выбранные стратегии кокколитофоров зависят от стадии их жизненного цикла. Когда кокколитофороры диплоидны, они подвергаются r-отбору. На этом этапе они переносят более широкий диапазон питательных составов. Когда они гаплоидны, они подвергаются K-отбору и часто более конкурентоспособны в стабильной среде с низким содержанием питательных веществ. [ 44 ] Большинство кокколитофоров являются стратегами калия и обычно встречаются в бедных питательными веществами поверхностных водах. Они являются плохими конкурентами по сравнению с другим фитопланктоном и процветают в средах обитания, где другой фитопланктон не выжил бы. [ 45 ] Эти две стадии жизненного цикла кокколитофоров происходят сезонно, когда больше питательных веществ доступно в более теплые сезоны и меньше — в более прохладные. Этот тип жизненного цикла известен как сложный гетероморфный жизненный цикл. [ 44 ]
Глобальное распространение
[ редактировать ]
Кокколитофоры встречаются во всем Мировом океане. Их распределение варьируется по вертикали в зависимости от стратифицированных слоев океана и географически в зависимости от временных зон. [ 46 ] В то время как большинство современных кокколитофоров могут находиться в связанных с ними стратифицированных олиготрофных условиях, наиболее многочисленные районы кокколитофоров, где наблюдается наибольшее видовое разнообразие, расположены в субтропических зонах с умеренным климатом. [ 47 ] Хотя температура воды и интенсивность света, попадающего на поверхность воды, являются более влиятельными факторами на определение местонахождения видов, океанские течения также могут определять местонахождения определенных видов кокколитофоров. [ 48 ]
Хотя подвижность и образование колоний варьируются в зависимости от жизненного цикла различных видов кокколитофоров, часто происходит чередование подвижной, гаплоидной фазы и неподвижной диплоидной фазы. На обеих фазах распространение организма во многом происходит за счет океанских течений и особенностей циркуляции. [ 49 ]
В Тихом океане было идентифицировано около 90 видов в шести отдельных зонах, относящихся к различным тихоокеанским течениям, которые содержат уникальные группы разных видов кокколитофоров. [ 50 ] Наибольшее разнообразие кокколитофоров в Тихом океане наблюдалось в районе океана, считающемся Центрально-Северной зоной, которая представляет собой территорию между 30 тот Н и 5 тот N, состоит из Северного экваториального течения и экваториального противотечения. Эти два течения движутся в противоположных направлениях, на восток и запад, что обеспечивает сильное смешивание вод и позволяет большому разнообразию видов заселить эту территорию. [ 50 ]
В Атлантическом океане наиболее многочисленными видами являются E. huxleyi и Florisphaera profunda с меньшими концентрациями видов Umbellosphaera ignores , Umbellosphaera tenuis и различных видов Gephyrocapsa . [ 50 ] На численность глубоководных видов кокколитофоров большое влияние оказывают глубины нутриклина и термоклина . Численность этих кокколитофоров увеличивается, когда нутриклин и термоклин глубоки, и уменьшается, когда они неглубоки. [ 51 ]

Полное распространение кокколитофоров в настоящее время неизвестно, а некоторые регионы, такие как Индийский океан, изучены не так хорошо, как другие места в Тихом и Атлантическом океанах. Также очень сложно объяснить распределение из-за множества постоянно меняющихся факторов, влияющих на свойства океана, таких как прибрежный и экваториальный апвеллинг , фронтальные системы, донная среда, уникальная топография океана и очаги изолированных высоких или низких температур воды. [ 53 ]
Верхняя фототическая зона имеет низкую концентрацию питательных веществ, высокую интенсивность и проникновение света и обычно более высокую температуру. Нижняя фотозона отличается высокой концентрацией питательных веществ, низкой интенсивностью и проникновением света и относительно прохладной. Средняя фотозона — это область, которая содержит одинаковые значения между нижней и верхней фотозоной. [ 47 ]



Большой кальцитовый пояс
[ редактировать ]Большой кальцитовый пояс Южного океана — это регион повышенной концентрации кальцита в верхних слоях океана в летнее время, происходящий от кокколитофоров, несмотря на то, что этот регион известен преобладанием диатомовых водорослей . Перекрытие двух основных групп фитопланктона, кокколитофор и диатомей, в динамических фронтальных системах, характерных для этого региона, обеспечивает идеальную среду для изучения окружающей среды. влияет на распространение различных видов внутри этих таксономических групп. [ 56 ]
Большой кальцитовый пояс, определяемый как повышенное содержание неорганического углерода в виде частиц (PIC), встречающееся наряду с сезонным повышенным содержанием хлорофилла а южной весной и летом в Южном океане. [ 57 ] играет важную роль в колебаниях климата, [ 58 ] [ 59 ] занимая более 60% площади Южного океана (30–60° ю.ш.). [ 60 ] В регионе между 30° и 50° ю.ш. наблюдается самое высокое поглощение антропогенного углекислого газа (CO 2 ) наряду с Северной Атлантикой и северной частью Тихого океана. [ 61 ]
Влияние глобального изменения климата на распространение
[ редактировать ]Недавние исследования показывают, что изменение климата оказывает прямое и косвенное влияние на распространение и продуктивность кокколитофор. На них неизбежно повлияют повышение температуры и термическая стратификация верхнего слоя океана, поскольку они являются основными средствами контроля их экологии, хотя неясно, приведет ли глобальное потепление к чистому увеличению или уменьшению численности кокколитофоров. Поскольку они являются кальцифицирующими организмами, было высказано предположение, что закисление океана из-за увеличения количества углекислого газа может серьезно повлиять на кокколитофоры. [ 51 ] Недавнее увеличение выбросов CO 2 привело к резкому увеличению популяции кокколитофоров. [ 62 ]
Роль в пищевой сети
[ редактировать ]

Кокколитофоры — одни из наиболее распространенных первичных производителей в океане. Как таковые, они вносят большой вклад в первичную продуктивность тропических и субтропических океанов, однако какой именно объем еще предстоит зарегистрировать. [ 66 ]
Зависимость от питательных веществ
[ редактировать ]Соотношение концентраций азота , фосфора и силикатов в отдельных районах океана диктует конкурентное доминирование внутри фитопланктонных сообществ. Каждое соотношение по существу склоняет шансы в пользу либо диатомовых водорослей , либо других групп фитопланктона, таких как кокколитофоры. Низкое соотношение силикатов к азоту и фосфору позволяет кокколитофорам вытеснять другие виды фитопланктона; однако, когда соотношение силикатов, фосфора и азота высокое, кокколитофоры вытесняются диатомовыми водорослями. Увеличение сельскохозяйственных процессов приводит к эвтрофикации вод и, таким образом, к цветению кокколитофоров в этих средах с высоким содержанием азота и фосфора и низким содержанием силиката. [ 5 ]
Влияние на продуктивность водного столба
[ редактировать ]Кальцит . в карбонате кальция позволяет кокколитам рассеивать больше света, чем поглощать Это имеет два важных последствия: 1) поверхностные воды становятся ярче, что означает, что у них более высокое альбедо , и 2) происходит фотоингибирование , то есть производство фотосинтеза уменьшается из-за избытка света. В случае 1) высокая концентрация кокколитов приводит к одновременному повышению температуры поверхностных вод и снижению температуры более глубоких вод. Это приводит к большей стратификации толщи воды и уменьшению вертикального перемешивания питательных веществ. Однако исследование 2012 года показало, что общее влияние кокколитофоров на увеличение радиационного воздействия на океан меньше, чем влияние антропогенных факторов. [ 67 ] Таким образом, общим результатом массового цветения кокколитофор является снижение продуктивности водной толщи, а не вклад в глобальное потепление.
Взаимодействие хищник-жертва
[ редактировать ]Их хищники включают обычных хищников всего фитопланктона, включая мелкую рыбу, зоопланктон и личинки моллюсков. [ 45 ] [ 68 ] Вирусы, специфичные для этого вида, были выделены из нескольких мест по всему миру и, по-видимому, играют важную роль в динамике весеннего цветения.
Токсичность
[ редактировать ]Никаких свидетельств токсичности кокколитофоров из окружающей среды не поступало, но они принадлежат к классу Prymnesiophyceae, который включает отряды с токсичными видами. Токсичные виды обнаружены у родов Prymnesium Massart и Chrysochromulina Lackey. представители рода Prymnesium Было обнаружено, что продуцируют гемолитические соединения - агент, ответственный за токсичность. Некоторые из этих токсичных видов являются причиной гибели крупных рыб и могут накапливаться в таких организмах, как моллюски; перенося его по пищевой цепи. Лабораторные тесты на токсичность показали, что представители океанических кокколитофор родов Emiliania, Gephyrocapsa, Calcidiscus и Coccolithus нетоксичны, как и виды прибрежного рода Hymenomonas , однако несколько видов Pleurochrysis и Jomonlithus , оба прибрежных рода, были токсичны для артемии . [ 68 ]
Взаимодействие с сообществом
[ редактировать ]Кокколитофориды преимущественно встречаются в виде одиночных свободно плавающих гаплоидных или диплоидных клеток. [ 46 ]
Соревнование
[ редактировать ]Большинству фитопланктона для выживания необходим солнечный свет и питательные вещества из океана, поэтому они процветают в районах с большим количеством богатой питательными веществами воды, поднимающейся с нижних уровней океана. Большинству кокколитофоров требуется солнечный свет только для производства энергии, и у них более высокий коэффициент поглощения нитратов по сравнению с поглощением аммония (азот необходим для роста и может использоваться непосредственно из нитрата, но не из аммония). Из-за этого они процветают в тихой, бедной питательными веществами среде, где другой фитопланктон голодает. [ 69 ] Компромиссы, связанные с такими более высокими темпами роста, включают меньший радиус клеток и меньший объем клеток, чем у других типов фитопланктона.
Вирусная инфекция и коэволюция
[ редактировать ]гигантские ДНК-содержащие вирусы Известно, что литически заражают кокколитофоры, особенно E. huxleyi . Эти вирусы, известные как вирусы E. huxleyi (EhV), по-видимому, почти исключительно заражают покрытую коккосферой диплоидную фазу жизненного цикла. Было высказано предположение, что, поскольку гаплоидный организм не инфицирован и, следовательно, не затронут вирусом, коэволюционная « гонка вооружений » между кокколитофорами и этими вирусами не следует классической эволюционной схеме Красной Королевы , а вместо этого следует «Чеширскому коту». экологическая динамика. [ 70 ] Более поздние работы показали, что вирусный синтез сфинголипидов и индукция запрограммированной гибели клеток обеспечивают более прямую связь для изучения коэволюционной гонки вооружений, подобной Красной Королеве, по крайней мере, между кокколитовирусами и диплоидным организмом. [ 43 ]
Эволюция и разнообразие
[ редактировать ]Кокколитофоры являются членами клады Haptophyta , которая является сестринской кладой Centrohelida , которые оба находятся в Haptista . [ 71 ] Самые древние из известных кокколитофоров известны из позднего триаса, на границе Нория и Рэта . [ 72 ] Разнообразие неуклонно увеличивалось в течение мезозоя, достигнув своего апогея в позднем меловом периоде . произошло резкое падение Однако во время мел-палеогенового вымирания , когда вымерло более 90% видов кокколитофоров. Кокколиты достигли другого, более низкого пика разнообразия во время палеоцен-эоценового термического максимума , но впоследствии их численность сократилась с олигоцена из-за снижения глобальных температур, при этом наиболее сильно пострадали виды, производившие крупные и сильно кальцинированные кокколиты. [ 26 ]


Раковины кокколитофоров
[ редактировать ]| Часть серии, посвященной |
| Биоминерализация |
|---|
 |
- Экзоскелет: коккосферы и кокколиты.
Каждый кокколитофор заключен в защитную оболочку из кокколитов , кальцинированных чешуек, которые составляют его экзоскелет или коккосферу . [ 73 ] Кокколиты создаются внутри клетки кокколитофора, и хотя некоторые виды поддерживают один слой на протяжении всей жизни, производя только новые кокколиты по мере роста клетки, другие постоянно производят и сбрасывают кокколиты.
Состав
[ редактировать ]Основным компонентом кокколитов является карбонат кальция или мел . Карбонат кальция прозрачен, поэтому фотосинтетическая активность организмов не снижается из-за инкапсуляции в коккосферу. [ 45 ]
Формирование
[ редактировать ]Кокколиты образуются в результате процесса биоминерализации, известного как кокколитогенез. [ 39 ] Обычно кальцификация кокколитов происходит в присутствии света, и эти чешуйки образуются гораздо чаще во время экспоненциальной фазы роста, чем в стационарной фазе. [ 74 ] Хотя процесс биоминерализации еще не до конца понятен, он жестко регулируется передачей сигналов кальция . Образование кальцита начинается в комплексе Гольджи , где белковые матрицы способствуют образованию кристаллов CaCO 3 , а сложные кислые полисахариды контролируют форму и рост этих кристаллов. [ 49 ] По мере образования каждой чешуи она экспортируется в везикулу, полученную из аппарата Гольджи , и добавляется на внутреннюю поверхность коккосферы. Это означает, что кокколиты, образовавшиеся совсем недавно, могут лежать под более старыми кокколитами. [ 42 ] В зависимости от стадии жизненного цикла фитопланктона могут образовываться два разных типа кокколитов. Голококколиты образуются только в гаплоидной фазе, лишены радиальной симметрии и состоят из от сотен до тысяч одинаковых мельчайших (около 0,1 мкм) ромбических кальцита кристаллов . Считается, что эти кристаллы формируются, по крайней мере частично, вне клетки. Гетерококколиты встречаются только в диплоидной фазе, имеют радиальную симметрию и состоят из относительно небольшого числа сложных кристаллических единиц (менее 100). Хотя они и редки, в планктоне наблюдались комбинированные коккосферы, содержащие как голококколиты, так и гетерококколиты, регистрирующие переходы жизненного цикла кокколитофоров. Наконец, коккосферы некоторых видов сильно модифицированы различными придатками, состоящими из специализированных кокколитов. [ 53 ]
Функция
[ редактировать ]Хотя точная функция коккосферы неясна, было предложено множество потенциальных функций. Наиболее очевидно, что кокколиты могут защищать фитопланктон от хищников. Похоже также, что это помогает им создать более стабильный уровень pH . В ходе фотосинтеза углекислый газ удаляется из воды, делая ее более щелочной. Кальцификация также удаляет углекислый газ, но химические процессы, стоящие за ней, приводят к противоположной реакции pH; это делает воду более кислой. Таким образом, сочетание фотосинтеза и кальцификации уравновешивает друг друга в отношении изменений pH. [ 75 ] Кроме того, эти экзоскелеты могут давать преимущество в производстве энергии, поскольку кокколитогенез, по-видимому, тесно связан с фотосинтезом. Органическое осаждение карбоната кальция из раствора бикарбоната приводит к образованию свободного углекислого газа непосредственно внутри клеточного тела водоросли, этот дополнительный источник газа затем доступен кокколитофору для фотосинтеза. Было высказано предположение, что они могут обеспечивать барьер, подобный клеточной стенке, для изоляции внутриклеточных химических веществ от морской среды. [ 76 ] Более специфические защитные свойства кокколитов могут включать защиту от осмотических изменений, химического или механического шока и коротковолнового света. [ 41 ] Также было высказано предположение, что дополнительный вес нескольких слоев кокколитов позволяет организму погружаться в более низкие, более богатые питательными веществами слои воды и, наоборот, что кокколиты добавляют плавучести, не давая клетке опускаться на опасные глубины. [ 77 ] Также предполагается, что придатки кокколита выполняют несколько функций, например, препятствуют выпасу зоопланктона. [ 53 ]
Использование
[ редактировать ]Кокколиты являются основным компонентом Мела , позднемелового горного образования, которое широко выходит на поверхность на юге Англии и образует Белые скалы Дувра , а также других подобных пород во многих других частях мира. [ 10 ] В настоящее время осадочные кокколиты являются основным компонентом известковых илов , которые покрывают до 35% площади дна океана и местами достигают километровой мощности. [ 49 ] Из-за своего обилия и широкого географического распространения кокколиты, составляющие слои этого ила, и меловой осадок, образующийся при его уплотнении, служат ценными микрофоссилиями .
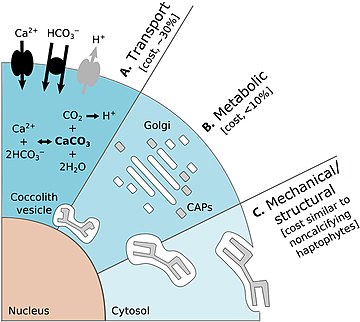
Кальцификация , биологическое производство карбоната кальция (CaCO 3 ), является ключевым процессом в морском углеродном цикле . Кокколитофоры являются основной планктонной группой, ответственной за пелагическое производство CaCO 3 . [ 78 ] [ 79 ] На диаграмме справа показаны энергетические затраты на кальцификацию кокколитофоров:
- (А) Процессы транспорта включают транспорт в клетку из окружающей морской воды субстратов первичной кальцификации Ca 2+ и HCO 3. − (черные стрелки) и удаление конечного продукта H + из ячейки (серая стрелка). Транспорт Ca 2+ через цитоплазму в сердечно-сосудистый сосуд является основной затратой, связанной с кальцификацией. [ 26 ]
- (B) Метаболические процессы включают синтез CAP (серые прямоугольники) комплексом Гольджи (белые прямоугольники), которые регулируют зарождение и геометрию кристаллов CaCO 3 . Завершенный кокколит (серая пластинка) представляет собой сложную структуру из сложно расположенных CAP и CaCO 3 . кристаллов [ 26 ]
- (C) Механические и структурные процессы ответственны за секрецию завершенных кокколитов, которые транспортируются из своего исходного положения, прилегающего к ядру, на периферию клетки, где они переносятся на поверхность клетки. Затраты, связанные с этими процессами, вероятно, будут сопоставимы с экзоцитозом органического масштаба в некальцифицирующихся гаптофитных водорослях. [ 26 ]

На диаграмме слева показаны преимущества кальцификации кокколитофоров. (A) Ускоренный фотосинтез включает CCM (1) и повышенное поглощение света за счет рассеяния редких фотонов для глубоко обитающих видов (2). (B) Защита от фотоповреждений включает защиту солнцезащитного козырька от ультрафиолетового (УФ) света и фотосинтетического активного излучения (ФАР) (1), а также рассеивание энергии в условиях яркого освещения (2). (C) Броневая защита включает защиту от вирусных/бактериальных инфекций (1) и выпаса селективными (2) и неселективными (3) травоядными. [ 26 ]
Степень, в которой кальцификация может адаптироваться к закислению океана, в настоящее время неизвестна. Физиологические исследования клеток обнаружили существенный H + выбросы (в результате использования HCO 3 − для внутриклеточной кальцификации) станет более дорогостоящим из-за продолжающегося закисления океана, поскольку электрохимический H + Градиент наизнанку уменьшается и пассивный отток протонов затрудняется. [ 80 ] Адаптированным клеткам придется активировать протонные каналы чаще , регулировать свой мембранный потенциал и/или снижать внутренний pH . [ 81 ] Снижение внутриклеточного pH серьезно повлияет на весь клеточный механизм и потребует совместной адаптации других процессов (например, фотосинтеза ), чтобы сохранить H. + истечение живое. [ 82 ] [ 83 ] Обязательный Н + Таким образом, отток, связанный с кальцификацией, может представлять собой фундаментальное ограничение для адаптации, что потенциально может объяснить, почему «кризис кальцификации» был возможен во время длительных (тысячи лет) CO 2 . событий возмущения [ 84 ] [ 85 ] хотя эволюционная адаптация к изменяющимся условиям карбонатного химического состава возможна в течение одного года. [ 84 ] [ 85 ] Раскрытие этих фундаментальных ограничений и пределов адаптации должно быть в центре внимания будущих исследований кокколитофоров, поскольку их знание является ключевой информацией, необходимой для понимания того, в какой степени реакция кальцификации на химические возмущения карбонатов может быть компенсирована эволюцией. [ 86 ]
Функциональные группы, защищенные силикатом или целлюлозой, такие как диатомовые водоросли и динофлагелляты, не нуждаются в поддержании H, связанного с кальцификацией. + отток. Таким образом, им, вероятно, не нужно адаптироваться, чтобы сохранить низкие затраты на производство конструктивных элементов. Напротив, динофлагелляты (кроме кальцинирующих видов; [ 87 ] с обычно неэффективными CO 2 фиксирующими ферментами RuBisCO, [ 88 ] могут даже извлечь выгоду из химических изменений, поскольку фотосинтетическая фиксация углерода как источника структурных элементов в форме целлюлозы должна облегчаться за счет удобрения CO 2 , связанного с подкислением океана . [ 89 ] [ 90 ] Если предположить, что любая форма панциря/экзоскелета защищает фитопланктон от хищников. [ 28 ] неизвестковая броня может быть предпочтительным решением для обеспечения защиты в будущем океане. [ 86 ]

Диаграмма справа показывает, как действуют сравнительные энергетические усилия для построения панциря у диатомовых водорослей, динофлагеллят и кокколитофоров. Панцирь (панцирь диатомовой водоросли ) кажется самым недорогим панцирем при любых обстоятельствах, поскольку диатомовые водоросли обычно вытесняют все другие группы, когда доступен силикат. Коккосфера относительно недорога при достаточном [CO 2 ], высоком [HCO 3 − ] и низкий [H + ] потому что субстрат насыщает и протоны легко высвобождаются в морскую воду. [ 80 ] Напротив, при высоких H следует отдавать предпочтение построению текальных элементов, которые представляют собой органические ( целлюлозные ) пластинки, составляющие оболочку динофлагеллят. + концентрации, поскольку они обычно совпадают с высокими концентрациями [CO 2 ]. В этих условиях динофлагелляты могут подавлять энергозатратную работу механизмов концентрации углерода, необходимых для производства органического исходного материала для их панциря. Следовательно, сдвиг условий карбонатной химии в сторону высоких значений [CO 2 ] может повысить их конкурентоспособность по сравнению с кокколитофорами. влияет огромное количество факторов, помимо химического состава карбонатов . Однако такое гипотетическое повышение конкурентоспособности за счет изменения условий карбонатного химического состава не приведет автоматически к доминированию динофлагеллят, поскольку на видовой состав также [ 86 ] [ 91 ]
Защита от хищников
[ редактировать ]В настоящее время данные, подтверждающие или опровергающие защитную функцию коккосферы от хищников, ограничены. Некоторые исследователи обнаружили, что общий уровень хищничества микрозоопланктона снижался во время цветения кокколитофоры Emiliania huxleyi . [ 92 ] [ 93 ] в то время как другие обнаружили высокие показатели выпаса микрозоопланктона в естественных сообществах кокколитофоров. [ 94 ] В 2020 году исследователи обнаружили, что in situ скорость поступления микрозоопланктона на E. huxleyi существенно не отличается от таковой на некальцифицирующемся фитопланктоне аналогичного размера. [ 95 ] В лабораторных экспериментах гетеротрофная динофлагеллята Oxyris marina некальцинированным предпочитала кальцинированные клетки E. huxleyi , что, как предполагалось, было связано с селективным пищевым поведением по размеру, поскольку кальцинированные клетки крупнее, чем некальцинированные E. huxleyi . [ 96 ] В 2015 году Харви и др. исследовали хищничество динофлагелляты O. marina на различных генотипах некальцифицирующегося E. huxleyi , а также на кальцинированных штаммах, различающихся степенью кальцинирования. [ 97 ] Они обнаружили, что частота употребления O. marina генотипа E. huxleyi зависела от предложенного , а не от степени их кальцификации. Однако в том же исследовании авторы обнаружили, что хищники, которые охотились на некальцифицирующиеся генотипы, росли быстрее, чем те, которых кормили кальцинированными клетками. [ 97 ] В 2018 году Стром и др. сравнили уровень хищничества динофлагелляты Amphidinium longum на кальцинированной добыче по сравнению с голой добычей E. huxleyi и не обнаружили никаких доказательств того, что коккосфера предотвращает проглатывание травоядным. [ 98 ] Вместо этого уровень потребления зависел от предложенного генотипа E. huxleyi. [ 98 ] В целом, эти два исследования предполагают, что генотип оказывает сильное влияние на поедание видами микрозоопланктона, но не удалось полностью выяснить, защищает ли кальцификация кокколитофоры от хищничества микрозоопланктона и каким образом. [ 1 ]
Значение в глобальном изменении климата
[ редактировать ]| Часть серии о |
| Планктон |
|---|
 |
Влияние на углеродный цикл
[ редактировать ]Кокколитофоры оказывают как долгосрочное, так и краткосрочное воздействие на углеродный цикл . Для образования кокколитов необходимо поглощение растворенного неорганического углерода и кальция. Карбонат кальция и диоксид углерода получают из кальция и бикарбоната в результате следующей химической реакции: [ 99 ]
- Что 2+ + 2HCO − 3 ⇌ CaCO 3 + CO 2 + H 2 O
Поскольку кокколитофоры являются фотосинтезирующими организмами, они способны использовать часть CO2 , выделяющегося в реакции кальцификации, для фотосинтеза . [ 100 ]
Однако производство карбоната кальция снижает щелочность поверхности, и в условиях низкой щелочности CO 2 вместо этого выбрасывается обратно в атмосферу. [ 101 ] В результате исследователи предположили, что массовое цветение кокколитофоров может способствовать глобальному потеплению в краткосрочной перспективе. [ 102 ] Однако более широко распространена идея, что в долгосрочной перспективе кокколитофоры способствуют общему снижению концентрации CO 2 в атмосфере . Во время кальцификации поглощаются два атома углерода, а один из них захватывается в виде карбоната кальция. Этот карбонат кальция опускается на дно океана в виде кокколитов и становится частью осадка; таким образом, кокколитофоры служат поглотителем выбрасываемого углерода, смягчая последствия выбросов парниковых газов. [ 102 ]
Эволюционные ответы на закисление океана
[ редактировать ]Исследования также показывают, что подкисление океана из-за увеличения концентрации CO 2 в атмосфере может повлиять на механизм кальцификации кокколитофоров. Это может не только повлиять на непосредственные события, такие как увеличение популяции или производства кокколитов, но также может вызвать эволюционную адаптацию видов кокколитофоров в течение более длительных периодов времени. Например, кокколитофоры используют H + ионные каналы для постоянной перекачки H + ионы из клетки во время образования кокколита. Это позволяет им избежать ацидоза , поскольку в противном случае образование кокколита привело бы к токсичному избытку H. + ионы. Когда функция этих ионных каналов нарушается, кокколитофоры останавливают процесс кальцификации, чтобы избежать ацидоза, образуя таким образом петлю обратной связи . [ 103 ] океана Низкая щелочность ухудшает функцию ионных каналов и, следовательно, оказывает эволюционное селективное давление на кокколитофороры и делает их (и другие кальцифицирующие организмы океана) уязвимыми для закисления океана. [ 104 ] В 2008 году полевые данные, свидетельствующие об увеличении кальцификации вновь образованных океанских отложений, содержащих кокколитофоры, подтвердили первые в истории экспериментальные данные, показывающие, что увеличение концентрации CO 2 в океане приводит к увеличению кальцификации этих организмов. Уменьшение массы кокколита связано как с увеличением концентрации CO 2 , так и с уменьшением концентрации CO 2 . CO 2− 3 в Мировом океане. Предполагается, что такая более низкая кальцификация ставит кокколитофоры в невыгодное экологическое положение. Однако некоторые виды, такие как Calcidiscus leptoporus , не подвергаются такому воздействию, в то время как наиболее распространенный вид кокколитофоров, E. huxleyi, может пострадать (результаты исследования неоднозначны). [ 103 ] [ 105 ] были обнаружены сильно кальцинированные кокколитофориды . CaCO 3 Кроме того, вопреки прогнозам, в условиях низкого насыщения [ 4 ] Понимание воздействия растущего закисления океана на виды кокколитофоров абсолютно необходимо для прогнозирования будущего химического состава океана, особенно его карбонатного состава. Жизнеспособные меры по сохранению и управлению станут результатом будущих исследований в этой области. Такие группы, как европейская CALMARO. [ 106 ] отслеживают реакцию популяций кокколитофоров на изменение pH и работают над определением экологически обоснованных мер контроля.
-
 Gephyrocapsa Oceanica (масштабная линейка 1 мкм)
Gephyrocapsa Oceanica (масштабная линейка 1 мкм) -

-




Влияние на летопись микрокаменелостей
[ редактировать ]Окаменелости кокколита представляют собой известные и ценные известковые микроокаменелости . Они являются крупнейшим глобальным источником биогенного карбоната кальция и вносят значительный вклад в глобальный углеродный цикл . [ 107 ] Они являются основной составляющей меловых отложений, таких как белые скалы Дувра .
Особый интерес представляют окаменелости, датируемые палеоцен-эоценовым термальным максимумом 55 миллионов лет назад. Считается, что этот период наиболее точно соответствует нынешнему уровню CO 2 в океане. [ 108 ] Наконец, полевые данные о окаменелостях кокколитофоров в горных породах были использованы, чтобы показать, что глубоководная летопись окаменелостей несет в себе предвзятость в летописи о горных породах, аналогичную той, которая широко распространена в отношении наземной летописи окаменелостей . [ 109 ]
Воздействие на океаны
[ редактировать ]Кокколитофориды помогают регулировать температуру океанов. Они процветают в теплых морях и выделяют диметилсульфид в воздух (ДМС), ядра которого помогают создавать более толстые облака, закрывающие солнце. [ 110 ] Когда океаны остывают, количество кокколитофорид уменьшается, а количество облаков также уменьшается. Когда меньше облаков, закрывающих солнце, температура также повышается. Таким образом, это поддерживает баланс и равновесие природы. [ 111 ] [ 112 ]
См. также
[ редактировать ]- Гипотеза КОГОТА
- Диметилсульфид
- Диметилсульфониопропионат
- Вирус Эмилиании Хаксли 86
- Плеврохризис картера
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и Хауност, Матиас; Рибезель, Ульф; Д'Амор, Франческо; Келтинг, Оле; Бах, Леннарт Т. (30 июня 2021 г.). «Влияние карбонатного кальция панциря кокколитофоров на питание и рост динофлагеллятного хищника» . Границы морской науки . 8 . Фронтирс Медиа С.А. дои : 10.3389/fmars.2021.664269 . ISSN 2296-7745 .
 Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Рост, Бьёрн; Рибезель, Ульф (2004). «Кокколитофоры и биологический насос: реакция на изменения окружающей среды». Кокколитофоры . Берлин, Гейдельберг: Springer Berlin Heidelberg. стр. 99–125. дои : 10.1007/978-3-662-06278-4_5 . ISBN 978-3-642-06016-8 .
- ^ Перейти обратно: а б де Врис, Йост; Монтейро, Фанни; Уиллер, Глен; Поултон, Алекс; Годриджан, Елена; Черино, Федерика; Малвинтер, Элиза; Лангер, Джеральд; Браунли, Колин (16 февраля 2021 г.). «Гапло-диплонтический жизненный цикл расширяет нишу кокколитофоров» . Биогеонауки . 18 (3). Коперник ГмбХ: 1161–1184. Бибкод : 2021BGeo...18.1161D . дои : 10.5194/bg-18-1161-2021 . ISSN 1726-4189 . S2CID 233976784 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Перейти обратно: а б Смит, HEK; и др. (2012), «Преобладание сильно кальцинированных кокколитофоров при низком насыщении CaCO3 зимой в Бискайском заливе», Proceedings of the National Academy of Sciences , 109 (23): 8845–8849, Bibcode : 2012PNAS..109.8845S , doi : 10.1073/pnas.1117508109 , PMC 3384182 , PMID 22615387
- ^ Перейти обратно: а б Юнев О.А.; и др. (2007), «Тенденции питательных веществ и фитопланктона на западном шельфе Черного моря в ответ на культурную эвтрофикацию и изменения климата», Estuarine, Coastal and Shelf Science , 74 (1–2): 63–67, Bibcode : 2007ECSS...74 ...63Y , doi : 10.1016/j.ecss.2007.03.030
- ^ Перейти обратно: а б Хэй, WW; Молер, HP; Рот, PH; Шмидт, Р.Р.; Будро, Дж. Э. (1967), «Зональность известкового наннопланктона кайнозоя побережья Мексиканского залива и Карибско-Антильской области и трансокеанская корреляция», Труды Ассоциации геологических обществ побережья Мексиканского залива , 17 : 428–480 .
- ^ «Биогеография и распространение микроорганизмов: обзор с акцентом на протистов», Acta Protozoologica , 45 (2): 111–136, 2005 г.
- ^ Бютенхейс, Эрик Т.; Пангерц, Таня; Франклин, Дэниел Дж.; Ле Кере, Коринн; Малин, Гилл (2008), «Скорость роста шести штаммов кокколиторипд как функция температуры», Лимнология и океанография , 53 (3): 1181–1185, Бибкод : 2008LimOc..53.1181B , doi : 10.4319/lo.2008.53. 3.1181 , S2CID 16601834
- ^ Эгге, Дж. К.; Акснес, Д.Л. (1992), «Силикат как регулирующее питательное вещество в конкуренции фитопланктона», Marine Ecology Progress Series , 83 (2): 281–289, Бибкод : 1992MEPS...83..281E , doi : 10.3354/meps083281
- ^ Перейти обратно: а б «Жизнь на грани зрения — Скотт Чимилески, Роберто Колтер | Издательство Гарвардского университета» . www.hup.harvard.edu . Проверено 26 января 2018 г.
- ^ Бенталеб, И.; и др. (1999), «Силикат как регулирующее питательное вещество в конкуренции фитопланктона», Marine Chemistry , 64 (4): 301–313, doi : 10.1016/S0304-4203(98)00079-6 .
- ^ «Международная ассоциация нанопланктона» .
- ^ Янг, младший; Гейзен, М.; Проберт, И. (2005). «Обзор избранных аспектов биологии кокколитофоров, имеющих значение для оценки палеобиоразнообразия» (PDF) . Микропалеонтология . 51 (4): 267–288. Бибкод : 2005MiPal..51..267Y . дои : 10.2113/gsmicropal.51.4.267 .
- ^ Шехтер, Моселио (2012). Эукариотические микробы . Академическая пресса. п. 239. ИСБН 978-0-12-383876-6 . Проверено 30 января 2015 г.
- ^ Баун, Пол Р.; Лиз, Джеки А.; Янг, Джереми Р. (2004). «Эволюция и разнообразие известкового наннопланктона с течением времени». Кокколитофоры . стр. 481–508. дои : 10.1007/978-3-662-06278-4_18 . ISBN 978-3-642-06016-8 .
- ^ Хэй, Уильям В. (2004). «Карбонатные потоки и известковый наннопланктон». Кокколитофоры . стр. 509–528. дои : 10.1007/978-3-662-06278-4_19 . ISBN 978-3-642-06016-8 .
- ^ Поултон, Алекс Дж.; Ади, Тим Р.; Балч, Уильям М.; Холлиган, Патрик М. (2007). «Связь скорости кальцификации кокколитофоров с динамикой сообщества фитопланктона: региональные различия и последствия для экспорта углерода». Глубоководные исследования, часть II: Актуальные исследования в океанографии . 54 (5–7): 538–557. Бибкод : 2007DSRII..54..538P . дои : 10.1016/j.dsr2.2006.12.003 .
- ^ Брокер, Уоллес; Кларк, Элизабет (2009). «Соотношение кокколита CaCO3 и фораминифер CaCO3 в глубоководных отложениях позднеголоцена» . Палеоокеанография . 24 (3). Бибкод : 2009PalOc..24.3205B . дои : 10.1029/2009PA001731 .
- ^ Клаас, Кристина; Арчер, Дэвид Э. (2002). «Связь тонущего органического вещества с различными типами минерального балласта в глубоком море: последствия для количества осадков» . Глобальные биогеохимические циклы . 16 (4): 1116. Бибкод : 2002GBioC..16.1116K . дои : 10.1029/2001GB001765 . S2CID 34159028 .
- ^ Хондзё, Сусуму; Манганини, Стивен Дж.; Кришфилд, Ричард А.; Франсуа, Роджер (2008). «Потоки твердых частиц органического углерода в недра океана и факторы, контролирующие биологический насос: синтез глобальных программ по улавливанию отложений с 1983 года». Прогресс в океанографии . 76 (3): 217–285. Бибкод : 2008Proce..76..217H . дои : 10.1016/j.pocean.2007.11.003 .
- ^ Франкиньюль, Мишель; Кэнон, Кристина; Гаттузо, Жан-Пьер (1994). «Морская кальцификация как источник углекислого газа: положительная обратная связь с увеличением содержания CO2 в атмосфере» . Лимнология и океанография . 39 (2): 458–462. Бибкод : 1994LimOc..39..458F . дои : 10.4319/lo.1994.39.2.0458 . hdl : 2268/246251 .
- ^ Рост, Бьёрн; Рибезель, Ульф (2004). «Кокколитофоры и биологический насос: реакция на изменения окружающей среды». Кокколитофоры . стр. 99–125. дои : 10.1007/978-3-662-06278-4_5 . ISBN 978-3-642-06016-8 .
- ^ Янг, младший (1987). Возможные функциональные интерпретации морфологии кокколита. Нью-Йорк: Springer-Verlag, 305–313.
- ^ Янг, младший (1994). «Функции кокколитов», в книге «Кокколитофоры», под редакцией А. Винтер и У. Г. Сиссера (Кембридж: издательство Кембриджского университета), 63–82.
- ^ Рэйвен, Дж.А.; Кроуферд, К. (2012). «Экологический контроль кальцификации кокколитофоров» . Серия «Прогресс в области морской экологии» . 470 : 137–166. Бибкод : 2012MEPS..470..137R . дои : 10.3354/meps09993 . hdl : 10453/114799 .
- ^ Перейти обратно: а б с д и ж г час я дж Монтейро, Фанни М.; Бах, Леннарт Т.; Браунли, Колин; Баун, Пол; Рикаби, Розалинда EM; Поултон, Алекс Дж.; Тиррелл, Тоби; Бофорт, Люк; Дуткевич, Стефани; Гиббс, Саманта; Гутовска, Магдалена А.; Ли, Рене; Рибезель, Ульф; Янг, Джереми; Риджвелл, Энди (2016). «Почему кальцинируется морской фитопланктон» . Достижения науки . 2 (7): e1501822. Бибкод : 2016SciA....2E1822M . дои : 10.1126/sciadv.1501822 . ПМЦ 4956192 . ПМИД 27453937 .
- ^ Мюллер, Мариус Н. (2019). «О генезисе и функции кальцификации кокколитофоров» . Границы морской науки . 6 . дои : 10.3389/fmars.2019.00049 .
- ^ Перейти обратно: а б Хамм, Кристиан; Сметачек, Виктор (2007). «Доспехи: почему, когда и как». Эволюция первичных продуцентов в море . стр. 311–332. дои : 10.1016/B978-012370518-1/50015-1 . ISBN 9780123705181 .
- ^ Брюссаард, Корина П.Д. (2004). «Вирусный контроль популяций фитопланктона - обзор1». Журнал эукариотической микробиологии . 51 (2): 125–138. дои : 10.1111/j.1550-7408.2004.tb00537.x . ПМИД 15134247 . S2CID 21017882 .
- ^ Джонс, Кристофер Т.; Грабб, Остин Р.; Нисимов, Юзеф И.; Натале, Фрэнк; Кнапп, Вики; Муи, Элвин; Фредрикс, Хелен Ф.; Ван Мой, Бенджамин А.С.; Бидл, Кей Д. (2019). «Взаимосвязь между кальцификацией и кокколитовирусной инфекцией» . Экологическая микробиология . 21 (6): 1896–1915. Бибкод : 2019EnvMi..21.1896J . дои : 10.1111/1462-2920.14362 . ПМЦ 7379532 . ПМИД 30043404 .
- ^ Хауност, Матиас; Рибезель, Ульф; Бах, Леннарт Т. (2020). «Оболочка карбоната кальция Emiliania huxleyi обеспечивает ограниченную защиту от вирусной инфекции» . Границы морской науки . 7 . дои : 10.3389/fmars.2020.530757 .
- ^ Кальбет, Альберт; Лэндри, Майкл Р. (2004). «Рост фитопланктона, выпас микрозоопланктона и круговорот углерода в морских системах». Лимнология и океанография . 49 (1): 51–57. Бибкод : 2004LimOc..49...51C . дои : 10.4319/lo.2004.49.1.0051 . hdl : 10261/134985 . S2CID 22995996 .
- ^ Майерс, KMJ; Поултон, Эй Джей; Дэниелс, CJ; Уэллс, СР; Вудворд, служба скорой помощи; Тарран, Джорджия; Виддикомб, CE; Мэр, диджей; Аткинсон, А.; Гиринг, SLC (2019). «Рост и смертность кокколитофоров весной в шельфовом море с умеренным климатом (Кельтское море, апрель 2015 г.)» . Прогресс в океанографии . 177 : 101928. Бибкод : 2019Proce.17701928M . дои : 10.1016/j.pocean.2018.02.024 . S2CID 135347218 .
- ^ Янг, младший (1994) «Функции кокколитов». В: Кокколитофоры , редакторы А. Винтер и У. Г. Сиссер (Кембридж: издательство Кембриджского университета), 63–82.
- ^ Тиллманн, Урбан (2004). «Взаимодействие между планктонными микроводорослями и простейшими травоядными1». Журнал эукариотической микробиологии . 51 (2): 156–168. дои : 10.1111/j.1550-7408.2004.tb00540.x . ПМИД 15134250 . S2CID 36526359 .
- ^ Брекельс, Миннесота; Робертс, ЕС; Арчер, SD; Малин, Г.; Стейнке, М. (2011). «Роль растворенных инфохимических веществ в обеспечении взаимодействия хищник-жертва у гетеротрофной динофлагелляты Oxyris marina» . Журнал исследований планктона . 33 (4): 629–639. дои : 10.1093/plankt/fbq114 .
- ^ Алоизи, Г. (6 августа 2015 г.). «Ковариация скорости метаболизма и размера клеток у кокколитофоров» . Биогеонауки . 12 (15). Коперник ГмбХ: 4665–4692. Бибкод : 2015BGeo...12.4665A . дои : 10.5194/bg-12-4665-2015 . ISSN 1726-4189 . S2CID 6227548 .
- ^ Хендерикс, Йоринтье (2008). «Правила размера кокколитофоров — реконструкция геометрии древних клеток и доли клеточного кальцита на основе ископаемых кокколитов». Морская микропалеонтология . 67 (1–2). Эльзевир Б.В.: 143–154. Бибкод : 2008МарМП..67..143H . doi : 10.1016/j.marmicro.2008.01.005 . ISSN 0377-8398 .
- ^ Перейти обратно: а б Мохеймани, Северная Каролина; Уэбб, JP; Боровицка, Массачусетс (2012), «Биоремедиация и другие потенциальные применения кокколитофоридных водорослей: обзор. Биоремедиация и другие потенциальные применения кокколитофоридных водорослей: обзор», Algal Research , 1 (2): 120–133, doi : 10.1016/ j.algal.2012.06.002
- ^ Перейти обратно: а б Биллард, Шанталь; Иноуе, Исоа (17 августа 2004 г.). «Что нового в биологии кокколитофоров?». В Тирштейне, Ганс Р.; Янг, Джереми Р. (ред.). Кокколитофоры – от молекулярных процессов к глобальному воздействию . Берлин: Спринглер. стр. 1–29. ISBN 9783540219286 . .
- ^ Перейти обратно: а б Джордан, RW (2012), «Haptophyta», eLS , doi : 10.1002/9780470015902.a0001981.pub2 , ISBN 978-0470016176
- ^ Перейти обратно: а б Янг, младший; Карен, Х. (2003). «Биоминерализация внутри везикул: кальцит кокколитов». В Даве, PM ; Ёрео, Джей-Джей; Вайнер, С. (ред.). Обзоры по минералогии и геохимии . Вашингтон, округ Колумбия: Минералогическое общество Америки. стр. 189–216.
- ^ Перейти обратно: а б Варди, А.; и др. (2012), «Динамика вируса-хозяина и субклеточный контроль судьбы клеток в естественной популяции кокколитофоров», Proceedings of the National Academy of Sciences , 109 (47): 19327–19332, Bibcode : 2012PNAS..10919327V , doi : 10.1073/ pnas.1208895109 , PMC 3511156 , PMID 23134731
- ^ Перейти обратно: а б с Худан; Проберт, я; Затыльный, С; Верон, Б; Биллард, К; и др. (2006), "Экология океанических кокколитофоров. I. Пищевые предпочтения двух стадий жизненного цикла Coccolithus braarudii и Calcidiscus leptoporus", Aquatic Microbial Ecology , 44 : 291–301, doi : 10.3354/ame044291
- ^ Перейти обратно: а б с Хоган, МЦ «Кокколитофоры» . В Кливленде Катлер Дж. (ред.). Энциклопедия Земли . Вашингтон, округ Колумбия: Коалиция экологической информации, Национальный совет по науке и окружающей среде.
- ^ Перейти обратно: а б Гейзен, М.; и др. (17 августа 2004 г.). «Изменение видового уровня кокколитофоров =». В Тирштейне, Ганс Р.; Янг, Джереми Р. (ред.). Кокколитофоры – от молекулярных процессов к глобальному воздействию . Берлин: Спринглер. стр. 1–29. ISBN 9783540219286 . .
- ^ Перейти обратно: а б Джордан, RW; Чемберлен, AHL (1997), «Биоразнообразие гаптофитных водорослей», Biodiversity & Conservation , 6 (1): 131–152, doi : 10.1023/A:1018383817777 , S2CID 9564456
- ^ Бекель; Бауманн, Карл-Хайнц; Генрих, Рюдигер; Кинкель, Ханно; и др. (2006), «Схемы распределения кокколитов в поверхностных отложениях Южной Атлантики и Южного океана в зависимости от градиентов окружающей среды», Deep-Sea Research Part I: Oceanographic Research Papers , 53 (6): 1073–1099, Bibcode : 2006DSRI...53.1073 Б , doi : 10.1016/j.dsr.2005.11.006
- ^ Перейти обратно: а б с де Варгас, К.; Обри, член парламента; Проберт, И.; Янг, Дж. (2007). «От прибрежных охотников до океанических фермеров». В Фальковски, П.Г.; Нолл, А.Х. (ред.). Происхождение и эволюция кокколитофоров . Бостон: Эльзевир. стр. 251–285.
- ^ Перейти обратно: а б с Окада; Хондзё, Сусуму; и др. (1973), «Распределение океанических кокколитофоров в Тихом океане», Deep-Sea Research and Oceanographic Abstracts , 20 (4): 355–374, Бибкод : 1973DSRA...20..355O , doi : 10.1016/0011-7471 (73)90059-4
- ^ Перейти обратно: а б Кинкель, Х.; и др. (2000), «Кокколитофоры в экваториальной части Атлантического океана: реакция на сезонную и позднечетвертичную изменчивость поверхностных вод», Морская микропалеонтология , 39 (1–4): 87–112, Бибкод : 2000MarMP..39...87K , doi : 10.1016/с0377-8398(00)00016-5
- ^ Гафар, Н.А., Эйр, Б.Д. и Шульц, КГ (2019). «Сравнение видовой чувствительности к изменению света и химии карбонатов при кальцинировании морского фитопланктона». Научные отчеты , 9 (1): 1–12. два : 10.1038/s41598-019-38661-0 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Перейти обратно: а б с Янг, младший; и др. (2009), «Функция кокколита и морфогенез: данные о кокколитофорах, несущих придатки, семейства syracosphaeraceae (haptophyta)», Journal of Phycology , 45 (1): 213–226, doi : 10.1111/j.1529-8817.2008.00643. х , PMID 27033659 , S2CID 27901484
- ^ Дэниелс, К.Дж., Шевард, Р.М. и Поултон, А.Дж. (2014) «Биогеохимические последствия сравнительных темпов роста видов Emiliania huxleyi и Coccolithus ». Биогеонауки , 11 (23): 6915–6925. два : 10.5194/bg-11-6915-2014 .
- ^ Дурак, Г.М., Тейлор, А.Р., Уокер, К.Э., Проберт, И., Де Варгас, К., Аудик, С., Шредер, Д., Браунли, К. и Уилер, Г.Л. (2016) «Роль диатомовых водорослей -подобные кремниевым переносчикам в кальцинирующих кокколитофорах». Природные коммуникации , 7 :10543. два : 10.1038/ncomms10543 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Смит, Хелен ЭК; Поултон, Алекс Дж.; Гарли, Ребекка; Хопкинс, Джейсон; Любельчик, Лаура К.; Драпо, Дэйв Т.; Раушенберг, Сара; Твининг, Бен С.; Бейтс, Николас Р.; Балч, Уильям М. (2017). «Влияние изменчивости окружающей среды на биогеографию кокколитофоров и диатомей Большого кальцитового пояса» . Биогеонауки . 14 (21): 4905–4925. Бибкод : 2017BGeo...14.4905S . дои : 10.5194/bg-14-4905-2017 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Балч, ВМ; Гордон, Ховард Р.; Боулер, Британская Колумбия; Драпо, DT; Бут, ES (2005). «Измерения карбоната кальция на поверхности мирового океана на основе данных спектрорадиометра среднего разрешения» . Журнал геофизических исследований . 110 (С7): C07001. Бибкод : 2005JGRC..110.7001B . дои : 10.1029/2004JC002560 .
- ^ Сармьенто, Хорхе Л.; Хьюз, Терция MC; Стоуффер, Рональд Дж.; Манабе, Сюкуро (1998). «Моделирование реакции углеродного цикла океана на антропогенное потепление климата». Природа . 393 (6682): 245–249. Бибкод : 1998Natur.393..245S . дои : 10.1038/30455 . S2CID 4317429 .
- ^ Сармьенто, JL; Слейтер, Р.; Барбер, Р.; Бопп, Л.; Дони, Южная Каролина; Херст, AC; Клейпас, Дж.; Матеар, Р.; Миколайевич, У.; Монфрей, П.; Солдатов В.; Сполл, ЮАР; Стоуффер, Р. (2004). «Реакция океанских экосистем на потепление климата» (PDF) . Глобальные биогеохимические циклы . 18 (3): н/д. Бибкод : 2004GBioC..18.3003S . дои : 10.1029/2003GB002134 . hdl : 1912/3392 . S2CID 15482539 .
- ^ Балч, ВМ; Драпо, DT; Боулер, Британская Колумбия; Лычковский, Э.; Бут, ES; Элли, Д. (2011). «Вклад кокколитофоров в балансы оптического и неорганического углерода во время эксперимента по газообмену в Южном океане: новые доказательства в поддержку гипотезы «Великого кальцитового пояса». Журнал геофизических исследований . 116 (С4): C00F06. Бибкод : 2011JGRC..116.0F06B . дои : 10.1029/2011JC006941 .
- ^ Сабина, CL; Фили, РА; Грубер, Н.; Ключ, РМ; Лук-порей.; Буллистер, Дж. Л.; Ваннинхоф, Р.; Вонг, CS; Уоллес, Д.В.; Тилбрук, Б.; Миллеро, Ф.Дж.; Пэн, TH; Козырь А.; Оно, Т.; Риос, АФ (2004). «Океанический поглотитель антропогенного CO2» (PDF) . Наука . 305 (5682): 367–371. Бибкод : 2004Sci...305..367S . дои : 10.1126/science.1097403 . ПМИД 15256665 . S2CID 5607281 .
- ^ Гитау, Беатрис (28 ноября 2015 г.). «Что способствует росту кокколитофоров в океанах?» . www.csmonitor.com . Христианский научный монитор . Проверено 30 ноября 2015 г.
- ^ «Вирусная зона» . ЭксПАСи . Проверено 15 июня 2015 г.
- ^ ICTV. «Таксономия вирусов: выпуск 2014 г.» . Проверено 15 июня 2015 г.
- ^ Крупнейшие известные вирусные геномы Giantviruses.org . Доступ: 11 июня 2020 г.
- ^ Рост, Б.; Рибеселл, У. (2004), «Кокколитофоры и биологический насос: реакция на изменения окружающей среды» (PDF) , Coccolithophores , 2 : 99–125, CiteSeerX 10.1.1.455.2864 , doi : 10.1007/978-3-662-06278 -4_5 , ISBN 978-3-642-06016-8 , заархивировано (PDF) из оригинала 10 ноября 2012 г.
- ^ Моррисси, Дж. Ф.; Сумич, Дж. Л. (2012). Введение в биологию морской жизни . п. 67.
- ^ Перейти обратно: а б Худан, А.; и др. (2004), «Токсичность прибрежных кокколитофоров (Prymnesiophyceae, Haptophyta)», Journal of Plankton Research , 26 (8): 875–883, doi : 10.1093/plankt/fbh079
- ^ Личман, Э.; и др. (2007), «Роль функциональных особенностей и компромиссов в структурировании сообществ фитопланктона: переход от клеточного к экосистемному уровню», Ecology Letters , 10 (12): 1170–1181, Bibcode : 2007EcolL..10.1170L , doi : 10.1111 /j.1461-0248.2007.01117.x , PMID 17927770
- ^ Фрада, М.; и др. (2008), «Стратегия побега кокколитофора Emiliania huxleyi «Чеширского кота» в ответ на вирусную инфекцию», Proceedings of the National Academy of Sciences , 105 (41): 15944–15949, Bibcode : 2008PNAS..10515944F , doi : 10.1073/pnas.0807707105 , PMC 2572935 , PMID 18824682
- ^ Тейлор, Элисон Р.; Браунли, Колин; Уиллер, Глен (3 января 2017 г.). «Биология кокколитофорных клеток: достижение прогресса» (PDF) . Ежегодный обзор морской науки . 9 (1). Годовые обзоры : 283–310. Бибкод : 2017ARMS....9..283T . doi : 10.1146/annurev-marine-122414-034032 . ISSN 1941-1405 . ПМИД 27814031 . Архивировано (PDF) из оригинала 16 июля 2021 г.
- ^ Гарден, Сильвия; Кристин, Леопольд; Ришос, Сильвен; Бартолини, Аннакьяра; Гальбрун, Бруно (октябрь 2012 г.). «Где и когда появились самые ранние кокколитофоры?: Где и когда появились самые ранние кокколитофоры?» . Летайя . 45 (4): 507–523. дои : 10.1111/j.1502-3931.2012.00311.x .
- ^ Фальковски, П.Г.; Нолл, АХ (29 августа 2007 г.). Эволюция первичных продуцентов в море . Амстердам, Бостон: Elsevier Academic Press. ISBN 9780123705181 .
- ^ Линсхутен, Корнелис; и др. (1991), «Роль цикла света и темноты и состава среды в производстве кокколитов Emiliania huxleyi (haptophyceae)», Journal of Phycology , 27 (1): 82–86, doi : 10.1111/j.0022-3646.1991 .00082.x , S2CID 84368830
- ^ «Микроскопические морские растения биоинженерируют окружающую среду для ускорения собственного роста – Разговор» . 2 августа 2016 г.
- ^ Вестбрук, П.; и др. (1983), «Кальцификация у Coccolithophoridae: расточительно или функционально?», Экологические бюллетени : 291–299.
- ^ Ирие, Такахиро; и др. (2010), «Увеличение затрат из-за закисления океана приводит к более сильной кальцификации фитопланктона: оптимальная стратегия роста кокколитофоров», PLOS ONE , 5 (10): e13436, Bibcode : 2010PLoSO...513436I , doi : 10.1371/журнал. pone.0013436 , PMC 2955539 , PMID 20976167
- ^ Крумхардт, Кристен М.; Ловендуски, Николь С.; Иглесиас-Родригес, М. Дебора; Клейпас, Джоан А. (2017). «Рост кокколитофоров и кальцификация в меняющемся океане» . Прогресс в океанографии . 159 : 276–295. Бибкод : 2017Proce.159..276K . дои : 10.1016/j.pocean.2017.10.007 .
- ^ Дэниелс, Крис Дж.; Поултон, Алекс Дж.; Балч, Уильям М.; Мараньон, Эмилио; Эйди, Тим; Боулер, Брюс С.; Черменьо, Педро; Харалампопулу, Анастасия; Кроуфорд, Дэвид В.; Драпо, Дэйв; Фэн, Юаньюань; Фернандес, Ана; Фернандес, Эмилио; Фрагозо, Главия М.; Гонсалес, Наталья; Грациано, Лиза М.; Хеслоп, Рэйчел; Холлиган, Патрик М.; Хопкинс, Джейсон; Уэте-Ортега, Мария; Хатчинс, Дэвид А.; Лам, Фиби Дж.; Липсен, Майкл С.; Лопес-Сандовал, Даффни К.; Лукайдес, Сократ; Маркетти, Адриан; Майерс, Кайл М.Дж.; Рис, Эндрю П.; Собрино, Кристина; и др. (2018). «Глобальный сборник показателей кальцификации кокколитофоров» . Данные науки о системе Земли . 10 (4): 1859–1876. Бибкод : 2018ESSD...10.1859D . дои : 10.5194/essd-10-1859-2018 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Перейти обратно: а б Тейлор, Элисон Р.; Чрачри, Абдул; Уилер, Глен; Годдард, Хелен; Браунли, Колин (2011). "H, управляемый напряжением + Канал, лежащий в основе гомеостаза pH при кальцификации кокколитофоров» . PLOS Biology . 9 (6): e1001085. doi : 10.1371/ . PMC 3119654. . PMID 21713028 journal.pbio.1001085
- ^ Тейлор, Элисон Р.; Браунли, Колин; Уилер, Глен Л. (2012). «Протонные каналы в водорослях: причины для волнения». Тенденции в науке о растениях . 17 (11): 675–684. doi : 10.1016/j.tplants.2012.06.009 . ПМИД 22819465 .
- ^ Беннер, Ина; Дайнер, Рэйчел Э.; Лефевр, Стефан К.; Ли, Дайан; Комада, Томоко; Карпентер, Эдвард Дж.; Стиллман, Джонатон Х. (2013). «Emiliania huxleyi увеличивает кальцификацию, но не экспрессию генов, связанных с кальцификацией, при длительном воздействии повышенной температуры и р CO 2» . Философские труды Королевского общества B: Биологические науки . 368 (1627). дои : 10.1098/rstb.2013.0049 . ПМЦ 3758179 . ПМИД 23980248 .
- ^ Лобек, Аннетт; Титдженс, Майке; Бунд, Андреас (2014). «Физическая Я-концепция, ориентация на индивидуально предпочтительные эталонные нормы и целевая ориентация у детей младшего школьного возраста во втором и четвертом классах». Журнал спортивной психологии . 21 :1–12. дои : 10.1026/1612-5010/a000109 .
- ^ Перейти обратно: а б Эрба, Элизабетта; Тремолада, Фабрицио (2004). «Потоки нанноископаемых карбонатов в раннем меловом периоде: реакция фитопланктона на эпизоды нутрификации, атмосферный CO2 и аноксию» . Палеоокеанография . 19 (1): н/д. Бибкод : 2004PalOc..19.1008E . дои : 10.1029/2003PA000884 .
- ^ Перейти обратно: а б Эрба, Элизабетта (2006). «Первые 150 миллионов лет истории известкового наннопланктона: взаимодействие биосферы и геосферы». Палеогеография, Палеоклиматология, Палеоэкология . 232 (2–4): 237–250. Бибкод : 2006PPP...232..237E . дои : 10.1016/j.palaeo.2005.09.013 .
- ^ Перейти обратно: а б с д Бах, Леннарт Томас; Рибезель, Ульф; Гутовска, Магдалена А.; Федервиш, Луиза; Шульц, Кай Георг (2015). «Объединяющая концепция чувствительности кокколитофоров к изменению химического состава карбонатов, встроенная в экологическую структуру» . Прогресс в океанографии . 135 : 125–138. Бибкод : 2015Proce.135..125B . дои : 10.1016/j.pocean.2015.04.012 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Ван Де Ваал, Дедмер Б.; Джон, Уве; Зивери, Патрисия; Райхарт, Герт-Ян; Хойнс, Мирья; Слейс, Аппи; Рост, Бьёрн (2013). «Подкисление океана снижает рост и кальцификацию морских динофлагеллят» . ПЛОС ОДИН . 8 (6): e65987. Бибкод : 2013PLoSO...865987V . дои : 10.1371/journal.pone.0065987 . ПМК 3679017 . ПМИД 23776586 .
- ^ Тортелл, Филипп Д. (2000). «Эволюционные и экологические перспективы поглощения углерода фитопланктоном» . Лимнология и океанография . 45 (3): 744–750. Бибкод : 2000LimOc..45..744T . дои : 10.4319/lo.2000.45.3.0744 .
- ^ Фу, Фэй-Сюэ; Чжан, Яохун; Уорнер, Марк Э.; Фэн, Юаньюань; Вс, июнь; Хатчинс, Дэвид А. (2008). «Сравнение будущего увеличения CO2 и влияния температуры на симпатрический минимум Heterosigma akashiwo и Prorocentrum». Вредные водоросли . 7 : 76–90. дои : 10.1016/j.hal.2007.05.006 .
- ^ Рейнфелдер, Джон Р. (2011). «Механизмы концентрации углерода в эукариотическом морском фитопланктоне». Ежегодный обзор морской науки . 3 : 291–315. Бибкод : 2011ARMS....3..291R . doi : 10.1146/annurev-marine-120709-142720 . ПМИД 21329207 .
- ^ Сюй, К.; Хатчинс, Д.; Гао, К. (9 апреля 2018 г.). «Расположение кокколита соответствует эйлеровой математике в кокколитофоре Emiliania huxleyi» . ПерДж . 6 : е4608. дои : 10.7717/PEERJ.4608 . ISSN 2167-8359 . ПМК 5896503 . ПМИД 29666762 . Викиданные Q5
- ^ Файлман, Э.С.; Каммингс, Д.Г.; Ллевеллин, Калифорния (2002). «Структура сообщества микропланктона и влияние выпаса микрозоопланктона во время цветения Emiliania huxleyi у побережья Девона». Журнал Морской биологической ассоциации Соединенного Королевства . 82 (3): 359–368. Бибкод : 2002JMBUK..82..359F . дои : 10.1017/S0025315402005593 . S2CID 85890446 .
- ^ Олсон, М.Брэйди; Стром, Сюзанна Л. (2002). «Рост фитопланктона, растительноядность микрозоопланктона и структура сообщества на юго-востоке Берингова моря: понимание формирования и временной устойчивости цветения Emiliania huxleyi». Глубоководные исследования, часть II: Актуальные исследования в океанографии . 49 (26): 5969–5990. Бибкод : 2002DSRII..49.5969O . дои : 10.1016/S0967-0645(02)00329-6 .
- ^ Майерс, KMJ; Поултон, Эй Джей; Дэниелс, CJ; Уэллс, СР; Вудворд, служба скорой помощи; Тарран, Джорджия; Виддикомб, CE; Мэр, диджей; Аткинсон, А.; Гиринг, SLC (2019). «Рост и смертность кокколитофоров весной в шельфовом море с умеренным климатом (Кельтское море, апрель 2015 г.)» . Прогресс в океанографии . 177 : 101928. Бибкод : 2019Proce.17701928M . дои : 10.1016/j.pocean.2018.02.024 . S2CID 135347218 .
- ^ Майерс, Кайл М.Дж.; Поултон, Алекс Дж.; Бидл, Кей; Таматракольн, Кимберли; Шилер, Бретань; Гиринг, Сара Л.К.; Уэллс, Сеона Р.; Тарран, Глен А.; Мэр Дэн; Джонсон, Мэтью; Рибезель, Ульф; Ларсен, Ауд; Варди, Асаф; Харви, Элизабет Л. (2020). «Обладание кокколитами не может отпугнуть поедающих микрозоопланктон» . Границы морской науки . 7 . дои : 10.3389/fmars.2020.569896 . hdl : 1912/26802 .
- ^ Хансен, ФК; Витте, HJ; Пассарж, Дж. (1996). «Выпас гетеротрофной динофлагелляты Oxyris marina: селективность по размеру и предпочтение кальцинированных клеток Emiliania huxleyi» . Водная микробная экология . 10 : 307–313. дои : 10.3354/ame010307 .
- ^ Перейти обратно: а б Харви, Элизабет Л.; Бидл, Кей Д.; Джонсон, Мэтью Д. (2015). «Последствия изменчивости штаммов и кальцификации при Emiliania выпасе микрозоопланктона huxleyion» . Журнал исследований планктона : fbv081. дои : 10.1093/plankt/fbv081 . hdl : 1912/7739 .
- ^ Перейти обратно: а б Стром, Сюзанна Л.; Брайт, Келли Дж.; Фредриксон, Керри А.; Куни, Элизабет К. (2018). «Защита фитопланктона: защищают ли кокколиты Emiliania huxleyi от хищников микрозоопланктона?» . Лимнология и океанография . 63 (2): 617–627. Бибкод : 2018LimOc..63..617S . дои : 10.1002/lno.10655 . S2CID 90415703 .
- ^ Мехия, Р. (2011), «Помогут ли ионные каналы кокколитофорам адаптироваться к закислению океана?», PLOS Biology , 9 (6): e1001087, doi : 10.1371/journal.pbio.1001087 , PMC 3119655 , PMID 21713029
- ^ Маккиндер; Уилер, Глен; Шредер, Деклан; Рибезель, Ульф; Браунли, Колин; и др. (2010), «Молекулярные механизмы, лежащие в основе кальцификации кокколитофоров», Geomicrobiology Journal , 27 (6–7): 585–595, Бибкод : 2010GmbJ...27..585M , doi : 10.1080/01490451003703014 , S2CID 85403507
- ^ Бейтс; Майклс, Энтони Ф.; Кнап, Энтони Х.; и др. (1996), «Изменения щелочности в Саргассовом море; геохимические свидетельства кальфикации?», Marine Chemistry , 51 (4): 347–358, Бибкод : 1996MarCh..51..347B , doi : 10.1016/0304-4203(95) )00068-2
- ^ Перейти обратно: а б Марш, Мэн (2003), «Регуляция образования CaCO3 в кокколитофорах», Сравнительная биохимия и физиология B , 136 (4): 743–754, doi : 10.1016/s1096-4959(03)00180-5 , PMID 14662299
- ^ Перейти обратно: а б Бофорт, Л.; и др. (2011), «Чувствительность кокколитофоров к карбонатной химии и закислению океана», Nature , 476 (7358): 80–3, doi : 10.1038/nature10295 , PMID 21814280 , S2CID 4417285
- ^ Тайрелл, Т.; и др. (1999), «Оптическое воздействие цветения океанических кокколитофор», Journal of Geophysical Research , 104 (C2): 3223–3241, Бибкод : 1999JGR...104.3223T , doi : 10.1029/1998jc900052
- ^ «Могут ли ракушки спасти мир?» . Independent.co.uk . 22 апреля 2008 г.
- ^ "кал.мар.о" . Архивировано из оригинала 30 декабря 2020 г. Проверено 24 апреля 2021 г.
- ^ Тейлор, Арканзас; Храчри, А.; Уилер, Г.; Годдард, Х.; Браунли, К. (2011). «Напряженно-зависимый H + канал, лежащий в основе гомеостаза pH при кальцификации кокколитофоров» . PLOS Biology . 9 (6): e1001085. doi : 10.1371/ . PMC 3119654. . PMID 21713028 journal.pbio.1001085
- ^ Селф-Трейл, JM; и др. (2012), «Изменения комплекса известковых наннофоссилий в палеоцен-эоценовом термическом максимуме: данные из условий шельфа» , Морская микропалеонтология , 92–93: 61–80, Бибкод : 2012MarMP..92...61S , doi : 10.1016/ j.marmicro.2012.05.003
- ^ Ллойд, GT; и др. (2011), «Количественная оценка систематической ошибки глубоководных пород и окаменелостей с использованием кокколитофоров», Геологическое общество, Лондон, Специальные публикации , 358 (1): 167–177, Бибкод : 2011GSLSP.358..167L , doi : 10.1144/sp358 .11 , S2CID 129049029
- ^ Чарльсон, Роберт Дж.; Лавлок, Джеймс Э.; Андреэ, Мейнрат О.; Уоррен, Стивен Г. (1987). «Океанический фитопланктон, атмосферная сера, альбедо облаков и климат». Природа . 326 (6114): 655–661. Бибкод : 1987Natur.326..655C . дои : 10.1038/326655a0 . S2CID 4321239 .
- ^ Лавлок, Джеймс (2007). Месть Геи . Пингвин. ISBN 978-0-14-102597-1 .
- ^ Ларсен, С.Х. (2005). «Изменчивость Солнца, диметилсульфид, облака и климат». Глобальные биогеохимические циклы . 19 (1): GB1014. Бибкод : 2005GBioC..19.1014L . дои : 10.1029/2004GB002333 . S2CID 128504924 .
Внешние ссылки
[ редактировать ]Источники подробной информации
- Nannotax3 - иллюстрированное руководство по систематике кокколитофоров и других нанноокаменелостей.
- INA — Международная ассоциация наннопланктона
- Эмилиании Хаксли Домашняя страница
Знакомство с кокколитофорами
- Калифорнийский университет, Беркли. Музей палеонтологии: «Знакомство с Prymnesiophyta».
- Палеонтологический портал: Известковый нанопланктон
- РадиоЛаб - подкаст о кокколитофорах

| Базы данных органов управления : Национальные |
|---|