Биологический жизненный цикл

В биологии биологический жизненный цикл (или просто жизненный цикл, если ясен биологический контекст) — это серия стадий жизни организма, которая начинается с зиготы, часто в яйце, и заканчивается взрослой особью, которая размножается. производя потомство в виде новой зиготы, которая затем сама проходит ту же серию стадий, причем процесс повторяется циклически.
«Эта концепция тесно связана с концепциями истории жизни, развития и онтогенеза , но отличается от них акцентом на обновлении». [ 1 ] [ 2 ] Переходы формы могут включать рост, бесполое размножение или половое размножение .
У некоторых организмов разные «поколения» видов сменяют друг друга в течение жизненного цикла. У растений и многих водорослей существуют две многоклеточные стадии, а жизненный цикл называется чередованием поколений . термин «история жизни» Часто используется , особенно для таких организмов, как красные водоросли , которые имеют три (или более) многоклеточные стадии, а не две. [ 3 ]
Жизненные циклы, включающие половое размножение, включают чередование гаплоидных ( n ) и диплоидных (2n ) изменение плоидности стадий, т.е. происходит . Чтобы вернуться из диплоидной стадии в гаплоидную, должен произойти мейоз . Что касается изменений плоидности , то различают три типа циклов:
- гаплоидный жизненный цикл — гаплоидная стадия — многоклеточная, а диплоидная — одноклеточная, мейоз «зиготический».
- диплоидный жизненный цикл — диплоидная стадия многоклеточная и гаплоидные гаметы , мейоз «гаметный». образуются
- гаплодиплонтный жизненный цикл (также называемый диплогаплонтным , дилобионтическим или дибионтным жизненным циклом) — встречаются многоклеточные диплоидные и гаплоидные стадии, мейоз «спорический».
Циклы различаются тем, когда происходит митоз (рост). Зиготический мейоз и гаметный мейоз имеют одну митотическую стадию: митоз происходит во время n- фазы зиготического мейоза и во время 2n - фазы гаметического мейоза. Поэтому зиготический и гаметный мейоз вместе называются «гаплобионтным» (одна митотическая фаза, не путать с гаплонтической). Споровый мейоз, с другой стороны, имеет митоз в две стадии: диплоидную и гаплоидную, что называется «диплобионтным» (не путать с диплоидным). [ нужна ссылка ]
Открытие
[ редактировать ]Изучением размножения и развития организмов занимались многие ботаники и зоологи.
Вильгельм Гофмейстер продемонстрировал, что чередование поколений является свойством, объединяющим растения, и опубликовал этот результат в 1851 году (см. сексуальность растений ).
Некоторые термины (гаплобионт и диплобионт), используемые для описания жизненных циклов, были первоначально предложены Нильсом Сведелиусом для водорослей, а затем стали использоваться и для других организмов. [ 4 ] [ 5 ] Другие термины (автогамия и гамонтогамия), используемые в жизненных циклах протистов, были введены Карлом Готлибом Греллем. [ 6 ] Описание сложных жизненных циклов различных организмов способствовало опровержению идей самозарождения в 1840-1850-х гг. [ 7 ]
Гаплонтический жизненный цикл
[ редактировать ]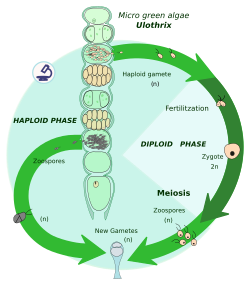
Зиготический мейоз — сразу после мейоз зиготы кариогамии , то есть слияния двух ядер клеток . Таким образом, организм завершает диплоидную фазу и производит несколько гаплоидных клеток. Эти клетки делятся митотически , образуя либо более крупные многоклеточные особи, либо более гаплоидные клетки. Два противоположных типа гамет (например, мужская и женская) этих особей или клеток сливаются, образуя зиготу.
За весь цикл зиготы — единственная диплоидная клетка; митоз происходит только в гаплоидной фазе.
Особи или клетки в результате митоза представляют собой гаплонты, поэтому этот жизненный цикл также называют гаплонтическим жизненным циклом. Гаплонты – это:
- У архепластид : некоторые зеленые водоросли (например, Chlamydomonas , Zygnema , Chara ) [ 8 ] [ 9 ]
- В страменопилиях : некоторые золотистые водоросли. [ 8 ] [ 9 ]
- У альвелятов : многие динофлагелляты , например Ceratium, Gymnodinium, некоторые апикомплексаны (например, Plasmodium ). [ 10 ]
- У ризарий : некоторые эвглифиды , [ 11 ] аскетоспории
- В раскопках : некоторые парабасалиды. [ 12 ]
- У амебозойных животных : Dictyostelium. [ 8 ] [ 9 ]
- У опистоконтов : большинство грибов (некоторые хитриды , зигомицеты , некоторые аскомицеты , базидиомицеты ) [ 8 ] [ 9 ] [ 13 ]
Диплонтический жизненный цикл
[ редактировать ]
При гаметном мейозе вместо немедленного мейотического деления с образованием гаплоидных клеток зигота делится митотически с образованием многоклеточной диплоидной особи или группы более одноклеточных диплоидных клеток. Клетки диплоидных особей затем подвергаются мейозу с образованием гаплоидных клеток или гамет . Гаплоидные клетки могут снова делиться (путем митоза) с образованием большего количества гаплоидных клеток, как у многих дрожжей, но гаплоидная фаза не является преобладающей фазой жизненного цикла. У большинства диплодтов митоз происходит только в диплоидной фазе, т.е. гаметы обычно образуются быстро и сливаются, образуя диплоидные зиготы. [ 14 ]
Во всем цикле гаметы обычно являются единственными гаплоидными клетками, а митоз обычно происходит только в диплоидной фазе.
Диплоидная многоклеточная особь представляет собой диплонт, поэтому гаметный мейоз также называют диплодным жизненным циклом. Диплонты – это:
- У архепластид : некоторые зеленые водоросли (например, Cladophora glomerata , [ 15 ] Тарелки [ 8 ] [ 9 ] )
- У страменопилес : некоторые бурые водоросли (у Fucales , однако, их жизненный цикл можно интерпретировать и как сильно гетероморфно-диплогаплонтный, с сильно редуцированной фазой гаметофита, как у цветковых растений ), [ 16 ] некоторые ксантофиты (например, Vaucheria ), [ 17 ] большинство диатомей , [ 12 ] некоторые оомицеты (например, Saprolegnia , Plasmopara viticola ), [ 8 ] [ 9 ] опалины , [ 12 ] некоторые « солнечники » (например, Actinophrys , Actinosphaerium ) [ 12 ] [ 18 ]
- В альвеолятах : инфузории. [ 12 ]
- В раскопках : некоторые парабасалиды. [ 12 ]
- У опистоконтов : животные, некоторые грибы (например, некоторые аскомицеты ) [ 8 ] [ 9 ]
Гаплодиплонтический жизненный цикл
[ редактировать ]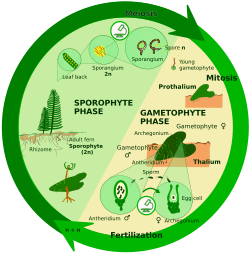
При споровом мейозе (также известном как промежуточный мейоз) зигота делится митотически с образованием многоклеточного диплоидного спорофита . Спорофит создает споры посредством мейоза, которые затем делятся митотически, образуя гаплоидные особи, называемые гаметофитами . Гаметофиты производят гаметы путем митоза. У некоторых растений гаметофит не только мелкий, но и недолговечный; у других растений и многих водорослей гаметофит является «доминирующей» стадией жизненного цикла. [ 19 ]
Гаплодиплонты – это:
- У архепластид : красные водоросли (имеющие два поколения спорофитов), некоторые зеленые водоросли (например, Ulva ), наземные растения. [ 8 ] [ 9 ]
- В страменопилях : большинство бурых водорослей. [ 8 ] [ 9 ]
- У ризарий : много фораминифер , [ 12 ] плазмодиофоромицеты [ 8 ] [ 9 ]
- У амеб : миксогастриды.
- У опистоконтов : некоторые грибы (некоторые хитриды , некоторые аскомицеты, такие как пивные дрожжи ). [ 8 ] [ 9 ]
- Другие эукариоты: гаптофиты. [ 12 ]
У некоторых животных есть система определения пола, называемая гаплодиплоидной , но она не связана с гаплодиплоидным жизненным циклом.
Вегетативный мейоз
[ редактировать ]Некоторые красные водоросли (например, Bonnemaisonia [ 20 ] и Lemanea ) и зеленые водоросли (такие как Prasiola ) имеют вегетативный мейоз, также называемый соматическим мейозом, что является редким явлением. [ 21 ] Вегетативный мейоз может происходить как в гаплодиплонтическом, так и в диплонтическом жизненном цикле. Гаметофиты остаются прикрепленными к спорофиту и являются его частью. Вегетативные (нерепродуктивные) диплоидные клетки подвергаются мейозу, образуя вегетативные гаплоидные клетки. Они претерпевают множество митозов и производят гаметы.
Другое явление, называемое вегетативной диплоидизацией, тип апомиксиса , встречается у некоторых бурых водорослей (например, Elachista stellaris ). [ 22 ] Клетки гаплоидной части растения спонтанно дублируют свои хромосомы с образованием диплоидной ткани.
Жизненный цикл паразита
[ редактировать ]Паразиты зависят от эксплуатации одного или нескольких хозяев . те, которые должны заразить более одного вида- Говорят, что Dirofilaria immitis , хозяина для завершения своего жизненного цикла, имеют сложные или непрямые жизненные циклы. Например, или сердечный червь, имеет непрямой жизненный цикл. Микрофилярии личиночную сначала должны быть проглочены самкой комара , где они развиваются в инфекционную . стадию Затем комар кусает животное и передает ему инфекционные личинки, где они мигрируют в легочную артерию и созревают во взрослых особей. [ 23 ]
Те паразиты, которые заражают один вид, имеют прямой жизненный цикл. Примером паразита с прямым жизненным циклом является Ancylostoma caninum , или собачий нематод. Они развиваются в окружающей среде до инфекционной личиночной стадии, затем проникают непосредственно в кожу собаки и созревают до взрослых особей в тонком кишечнике . [ 24 ] [ нужна проверка ]
Если паразиту необходимо заразить данного хозяина, чтобы завершить свой жизненный цикл, то говорят, что он является облигатным паразитом этого хозяина; иногда заражение носит факультативный характер — паразит может выжить и завершить свой жизненный цикл, не заражая конкретный вид хозяина. Паразиты иногда заражают хозяев, у которых они не могут завершить свой жизненный цикл; это случайные хозяева.
Хозяин, в котором паразиты размножаются половым путем, известен как окончательный, окончательный или первичный хозяин. У промежуточных хозяев паразиты либо не размножаются, либо размножаются бесполым путем, но у этого типа хозяев паразит всегда развивается до новой стадии. В некоторых случаях паразит заражает хозяина, но не развивается; такие хозяева называются паратеническими. [ 25 ] или транспортные узлы. Паратенический хозяин может быть полезен для повышения вероятности передачи паразита окончательному хозяину. Например, кошачий легочный червь ( Aelurostrongylus abstrusus ) использует в качестве промежуточного хозяина слизняка или улитку; личинка первой стадии проникает в моллюск и развивается в личинку третьей стадии, заразную для окончательного хозяина — кошки. Если мышь съедает слизняка, личинка третьей стадии проникнет в ткани мыши, но не будет развиваться. [ нужна ссылка ]

Эволюция
[ редактировать ]Примитивный тип жизненного цикла, вероятно, имел гаплоидные особи с бесполым размножением. [ 12 ] Бактерии и археи имеют такой жизненный цикл, и некоторые эукариоты, по-видимому, тоже (например, Cryptophyta , Choanoflagellata , многие Euglenozoa , многие Amoebozoa , некоторые красные водоросли, некоторые зеленые водоросли , несовершенные грибы , некоторые коловратки и многие другие группы, не обязательно гаплоидный). [ 26 ] Однако эти эукариоты, вероятно, не являются примитивно бесполыми, а утратили половое размножение или оно просто еще не наблюдалось. [ 27 ] [ 28 ] Многие эукариоты (включая животных и растения) демонстрируют бесполое размножение , которое может быть факультативным или обязательным в жизненном цикле, при этом половое размножение происходит более или менее часто. [ 29 ]
Отдельные организмы, участвующие в биологическом жизненном цикле, обычно стареют и умирают, тогда как клетки этих организмов, соединяющие последовательные поколения жизненного цикла (клетки зародышевой линии и их потомки), потенциально бессмертны. В основе этого различия лежит фундаментальная проблема биологии. Российский биолог и историк Жорес Александрович Медведев. [ 30 ] считали, что точность репликации генома и других синтетических систем сама по себе не может объяснить бессмертие зародышевых линий . Скорее Медведев считал, что известные особенности биохимии и генетики полового размножения указывают на наличие уникальных процессов поддержания и восстановления информации на этапе гаметогенеза биологического жизненного цикла. В частности, Медведев считал, что важнейшие возможности информационного обеспечения половых клеток создаются за счет рекомбинации при мейозе и репарации ДНК ; он рассматривал их как процессы внутри клеток зародышевой линии, которые были способны восстанавливать целостность ДНК и хромосом от тех типов повреждений, которые вызывают необратимое старение в клетках незародышевой линии, например, в соматических клетках . [ 30 ]
Предки каждой современной клетки, по-видимому, ведут непрерывную линию на протяжении более 3 миллиардов лет к зарождению жизни . являются не клетки, На самом деле бессмертными а клеточные линии, состоящие из нескольких поколений. [ 31 ] Бессмертие клеточной линии зависит от сохранения потенциала клеточного деления . Этот потенциал может быть утерян в любой конкретной линии из-за повреждения клеток, терминальной дифференцировки , как это происходит в нервных клетках, или запрограммированной гибели клеток ( апоптоза ) во время развития. Поддержание потенциала клеточного деления биологического жизненного цикла на протяжении последующих поколений зависит от предотвращения и точного восстановления клеточных повреждений, особенно повреждений ДНК . В половых организмах непрерывность зародышевой линии на протяжении последующих поколений клеточного цикла зависит от эффективности процессов предотвращения повреждений ДНК и восстановления тех повреждений ДНК , которые действительно происходят. Половые процессы у эукариот предоставляют возможность эффективной репарации повреждений ДНК в зародышевой линии путем гомологичной рекомбинации . [ 31 ] [ 32 ]
См. также
[ редактировать ]- Смена поколений – Репродуктивный цикл растений и водорослей
- Апомиксис - замена нормального полового размножения бесполым без оплодотворения.
- Гаплодиплоидия - биологическая система, в которой пол определяется количеством наборов хромосом.
- Парасексуальный цикл - неполовой механизм передачи генетического материала без мейоза.
- Партеногенез – бесполое размножение без оплодотворения.
- Метаморфоза - Глубокие изменения структуры тела в постэмбриональном развитии организма.
- Репродуктивная биология - раздел биологии, изучающий репродукцию.
- Митотическая рекомбинация - тип генетической рекомбинации.
Ссылки
[ редактировать ]- ^ Белл, Грэм; Куфопану, Василики (1991). «Архитектура жизненного цикла малых организмов». Философские труды: Биологические науки . 332 (1262): 81–89. Бибкод : 1991РСТБ.332...81Б . дои : 10.1098/rstb.1991.0035 . JSTOR 55494 .
- ^ Родригес, Хулиани Кола Фернандес; Годиньо, Жозеан Лима Прадо; Де Соуза, Вандерли (2014). «Биология патогенных трипаносоматид человека: эпидемиология, жизненный цикл и ультраструктура». Белки и протеомика лейшмании и трипаносомы . Субклеточная биохимия. Том. 74. стр. 1–42. дои : 10.1007/978-94-007-7305-9_1 . ISBN 978-94-007-7304-2 . ПМИД 24264239 .
- ^ Диксон, PS 1973. Биология Rhodophyta. Оливер и Бойд. ISBN 0 05 002485 Х [ нужна страница ]
- ^ К. Скоттсберг (1961), «Нильс Эберхард Сведелиус. 1873–1960», Биографические мемуары членов Королевского общества , 7 : 294–312, doi : 10.1098/rsbm.1961.0023
- ^ Сведелиус, Н. 1931. Ядерные фазы и чередование у Rhodophyceae. Архивировано 5 октября 2013 г. в Wayback Machine в: Дополнения к Botanical Centralblatt. Том 48/1: 38–59.
- ^ Маргулис, Л. (6 февраля 1996 г.). «Архейно-эубактериальные слияния в происхождении эукариев: филогенетическая классификация жизни» . Труды Национальной академии наук Соединенных Штатов Америки . 93 (3): 1071–1076. Бибкод : 1996PNAS...93.1071M . дои : 10.1073/pnas.93.3.1071 . ПМЦ 40032 . ПМИД 8577716 .
- ^ Моселио Шехтер (2009). Энциклопедия микробиологии . Академическая пресса. Том 4, с. 85.
- ^ Перейти обратно: а б с д и ж г час я дж к Диас, TE; Фернандес-Карвахаль, К.; Фернандес, JA (2004). Курс ботаники . Хихон: Треа.
- ^ Перейти обратно: а б с д и ж г час я дж к Диас Гонсалес, Томас; Фернандес-Карвахаль Альварес, Мю дель Кармен; Фернандес Прието, Хосе Антонио. «Ботаника: Биологические циклы растений» . Кафедра биологии организмов и систем Университета Овьедо (на испанском языке). Архивировано из оригинала 14 мая 2020 года.
- ^ Синден, Р.Э.; Хартли, Р.Х. (ноябрь 1985 г.). «Идентификация мейотического отдела малярийных паразитов». Журнал протозоологии . 32 (4): 742–744. дои : 10.1111/j.1550-7408.1985.tb03113.x . ПМИД 3906103 .
- ^ Лар, Дэниел Дж.Г.; Парфри, Лаура Вегенер; Митчелл, Эдвард А.Д.; Кац, Лаура А.; Лара, Энрике (22 июля 2011 г.). «Целомудрие амеб: переоценка доказательств пола у амебоидных организмов» . Труды Королевского общества B: Биологические науки . 278 (1715): 2081–2090. дои : 10.1098/rspb.2011.0289 . ПМК 3107637 . ПМИД 21429931 .
- ^ Перейти обратно: а б с д и ж г час я Руперт, Эдвард Э.; Фокс, Ричард С.; Барнс, Роберт Д. (2004). Зоология беспозвоночных: функционально-эволюционный подход . Томсон-Брукс/Коул. п. 26. ISBN 978-0-03-025982-1 .
- ^ ван ден Хук, Манн и Янс 1995 , стр. 15.
- ^ Смит, Гилберт М. (1938). «Ядерные фазы и чередование поколений у Chlorophyceae» . Ботанический обзор . 4 (3): 132–139. Бибкод : 1938BotRv...4..132S . дои : 10.1007/BF02872350 . ISSN 0006-8101 . JSTOR 4353174 .
- ^ ОП Шарма. Учебник по Водорослям, с. 189
- ^ ван ден Хук, Манн и Янс 1995 , стр. 207.
- ^ ван ден Хук, Манн и Янс 1995 , стр. 124.
- ^ Белл, Грэм (1988). Секс и смерть у простейших: история одержимости . Издательство Кембриджского университета. п. 11. ISBN 978-0-521-36141-5 .
- ^ Бенничи, Андреа (2008). «Происхождение и ранняя эволюция наземных растений: проблемы и соображения» . Коммуникативная и интегративная биология . 1 (2): 212–218. дои : 10.4161/cib.1.2.6987 . ISSN 1942-0889 . ПМК 2686025 . ПМИД 19513262 .
- ^ Сальвадор Солер, Ноэми; Гомес Гаррета, Амелия; Антония Рибера Сигуан, М. (август 2009 г.). «Соматический мейоз в истории жизни Bonnemaisonia asparagoides и Bonnemaisonia clavata (Bonnemaisoniales, Rhodophyta) с Пиренейского полуострова». Европейский журнал психологии . 44 (3): 381–393. Бибкод : 2009EJPhy..44..381S . дои : 10.1080/09670260902780782 . S2CID 217511084 .
- ^ ван ден Хук, Манн и Янс 1995 , стр. 82.
- ^ Льюис, Рэймонд Дж. (январь 1996 г.). «Хромосомы бурых водорослей». Психология . 35 (1): 19–40. Бибкод : 1996Phyco..35...19L . дои : 10.2216/i0031-8884-35-1-19.1 .
- ^ «ВетФолио» . www.vetfolio.com . Проверено 18 мая 2021 г.
- ^ Дац, Крейг (2011). «Паразитарные и протозойные болезни». Педиатрия мелких животных . стр. 154–160. дои : 10.1016/B978-1-4160-4889-3.00019-X . ISBN 978-1-4160-4889-3 .
- ^ Шмидт и Робертс. 1985. Основы паразитологии, 3-е изд. Times Mirror / Издательство колледжа Мосби [ нужна страница ]
- ^ Хейвуд, П.; Маги, ПТ (1976). «Мейоз у простейших. Некоторые структурные и физиологические аспекты мейоза у водорослей, грибов и простейших» . Бактериологические обзоры . 40 (1): 190–240. дои : 10.1128/ммбр.40.1.190-240.1976 . ПМК 413949 . ПМИД 773364 .
- ^ Шере-Бану Малик; Артур В. Пайтлинг; Лорен М. Стефаниак; Эндрю М. Шурко и Джон М. Логсдон-младший (2008). «Расширенный перечень консервативных мейотических генов доказывает наличие пола у Trichomonas vaginalis » . ПЛОС ОДИН . 3 (8): e2879. Бибкод : 2008PLoSO...3.2879M . дои : 10.1371/journal.pone.0002879 . ПМЦ 2488364 . ПМИД 18663385 .
- ^ Спейер, Дэйв; Лукеш, Юлиус; Элиаш, Марек (21 июля 2015 г.). «Секс — это вездесущий, древний и неотъемлемый атрибут эукариотической жизни» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (29): 8827–8834. Бибкод : 2015PNAS..112.8827S . дои : 10.1073/pnas.1501725112 . ПМЦ 4517231 . ПМИД 26195746 .
- ^ Шен, Иса; Мартенс, Коэн; Дейк, Питер ван (2009). Потерянный пол: эволюционная биология партеногенеза . Springer Science & Business Media. ISBN 978-90-481-2770-2 . [ нужна страница ]
- ^ Перейти обратно: а б Медведев, Жорес А. (1981). «О бессмертии зародышевой линии: генетические и биохимические механизмы. Обзор». Механизмы старения и развития . 17 (4): 331–359. дои : 10.1016/0047-6374(81)90052-X . ПМИД 6173551 . S2CID 35719466 .
- ^ Перейти обратно: а б Бернштейн, К.; Бернштейн, Х.; Пейн, К. (1999). «Бессмертие клеток: поддержание потенциала деления клеток». Иммортализация клеток . Прогресс молекулярной и субклеточной биологии. Том. 24. С. 23–50. дои : 10.1007/978-3-662-06227-2_2 . ISBN 978-3-642-08491-1 . ПМИД 10547857 .
- ^ Авис, Джон К. (октябрь 1993 г.). «Перспектива: эволюционная биология старения, полового размножения и восстановления ДНК» . Эволюция . 47 (5): 1293–1301. дои : 10.1111/j.1558-5646.1993.tb02155.x . ПМИД 28564887 . S2CID 29262885 .
Источники
[ редактировать ]- ван ден Хук, К.; Ноль; Янс, Х.М. (1995). Водоросли: введение в психологию . Издательство Кембриджского университета. ISBN 978-0-521-31687-3 .
Дальнейшее чтение
[ редактировать ]- Боннер, Джон Тайлер (1995). Жизненные циклы: размышления биолога-эволюциониста . Издательство Принстонского университета. ISBN 978-0-691-00151-7 .
- Валеро, Мириам; Ричерд, Софи; Перро, Вероника; Дестомб, Кристоф (январь 1992 г.). «Эволюция чередования гаплоидных и диплоидных фаз в жизненных циклах». Тенденции в экологии и эволюции . 7 (1): 25–29. дои : 10.1016/0169-5347(92)90195-H . ПМИД 21235940 .
- Мейбл, Барбара К.; Отто, Сара П. (1998). «Эволюция жизненных циклов с гаплоидными и диплоидными фазами». Биоэссе . 20 (6): 453–462. doi : 10.1002/(sici)1521-1878(199806)20:6<453::aid-bies3>3.0.co;2-n . S2CID 11841044 .
| Базы данных органов управления : Национальные |
|---|