Митотическая рекомбинация
Митотическая рекомбинация — это тип генетической рекомбинации , которая может происходить в соматических клетках во время их подготовки к митозу как у половых, так и у бесполых организмов. У бесполых организмов изучение митотической рекомбинации является одним из способов понять генетическую связь , поскольку это единственный источник рекомбинации внутри человека. [1] Кроме того, митотическая рекомбинация может привести к экспрессии рецессивных аллелей у гетерозиготного человека. Эта экспрессия имеет важное значение для изучения онкогенеза и летальных рецессивных аллелей. [1] [2] Митотическая гомологичная рекомбинация происходит главным образом между сестринскими хроматидами после репликации (но до деления клеток). Межсестринская гомологичная рекомбинация обычно генетически молчалива. Во время митоза частота рекомбинации между несестринскими гомологичными хроматидами составляет лишь около 1% от частоты рекомбинации между сестринскими хроматидами. [3]
Открытие
[ редактировать ]Открытие митотической рекомбинации произошло в результате наблюдения двойных пятен у Drosophila melanogaster . Эта двойная пятнистость, или мозаичная пятнистость, наблюдалась у D. melanogaster еще в 1925 г., но только в 1936 г. Курт Стерн объяснил ее результатом митотической рекомбинации. До работы Стерна предполагалось, что близнецовая мазня произошла потому, что определенные гены обладали способностью уничтожать хромосому, на которой они располагались. [4] Более поздние эксперименты выявили, когда в клеточном цикле происходит митотическая рекомбинация, и механизмы, лежащие в основе рекомбинации.
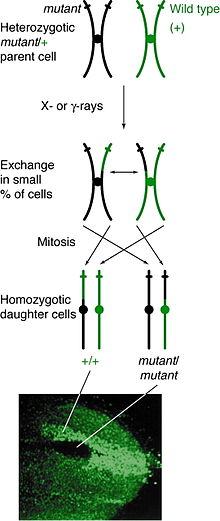
Возникновение
[ редактировать ]Митотическая рекомбинация может произойти в любом локусе, но наблюдается у людей, гетерозиготных по данному локусу. Если событие кроссовера между несестринскими хроматидами затрагивает этот локус, то обе гомологичные хромосомы будут иметь по одной хроматиде, содержащей каждый генотип. Результирующий фенотип дочерних клеток зависит от того, как хромосомы выстраиваются в метафазной пластинке . Если хроматиды, содержащие разные аллели, выстраиваются на одной стороне пластинки, то полученные дочерние клетки будут выглядеть гетерозиготными и необнаружимыми, несмотря на событие кроссинговера. Однако если хроматиды, содержащие одни и те же аллели, выстраиваются на одной стороне, дочерние клетки будут гомозиготными в этом локусе. Это приводит к пятнистости близнецов , когда одна клетка представляет гомозиготный рецессивный фенотип, а другая клетка имеет гомозиготный фенотип дикого типа. Если эти дочерние клетки продолжат реплицироваться и делиться, пятна-близнецы будут продолжать расти и отражать дифференциальный фенотип.
Митотическая рекомбинация происходит во время интерфазы . Было высказано предположение, что рекомбинация происходит во время G1, когда ДНК находится в двухцепочечной фазе, и реплицируется во время синтеза ДНК. [5] Также возможно, что разрыв ДНК, приводящий к митотической рекомбинации, произойдет во время G1, но восстановление произойдет после репликации. [6] [7]
Реакция на повреждение ДНК
[ редактировать ]У почкующихся дрожжей Saccharomyces cerevisiae мутации в нескольких генах, необходимых для митотической (и мейотической) рекомбинации, вызывают повышенную чувствительность к инактивации радиацией и/или генотоксичными химическими веществами . [8] Например, ген rad52 необходим для митотической рекомбинации. [9] а также мейотическая рекомбинация. [10] Мутантные дрожжевые клетки Rad52 обладают повышенной чувствительностью к уничтожению рентгеновскими лучами , метилметансульфонатом и сшивания ДНК агентом 8-метоксипсораленом плюс УФ-свет, что позволяет предположить, что митотическая рекомбинационная репарация необходима для устранения различных повреждений ДНК, вызванных этими агентами.
Механизмы
[ редактировать ]Механизмы митотической рекомбинации аналогичны механизмам мейотической рекомбинации. К ним относятся обмен сестринских хроматид и механизмы, связанные с восстановлением двухцепочечного разрыва ДНК путем гомологичной рекомбинации, такой как одноцепочечный отжиг , синтез-зависимый отжиг цепи ( SDSA ) и конверсия генов через промежуточное соединение двойного соединения Холлидея или SDSA. Кроме того, возможна негомологичная митотическая рекомбинация, которую часто можно объяснить негомологичным соединением концов . [6] [7] [11] [12]
Метод
[ редактировать ]Существует несколько теорий о том, как происходит митотический кроссинговер. В модели простого кроссовера две гомологичные хромосомы перекрываются на общем хромосомном хрупком участке (CFS) или рядом с ним. Это приводит к двухцепочечному разрыву . [13] который затем восстанавливается с использованием одной из двух нитей. Это может привести к тому, что две хроматиды поменяются местами. В другой модели две перекрывающиеся сестринские хроматиды образуют двойное соединение Холлидея в общем сайте повтора, а затем разрезаются таким образом, что меняются местами. В любой модели хромосомы не гарантированно торгуются равномерно или даже воссоединяются на противоположных сторонах, поэтому большинство моделей расщепления не приводят к какому-либо событию кроссинговера. Неравномерная торговля приводит ко многим пагубным последствиям митотического кроссинговера.
Альтернативно, кроссинговер может произойти во время репарации ДНК. [14] если из-за обширного повреждения гомологичная хромосома выбрана в качестве матрицы вместо сестринской хроматиды. Это приводит к синтезу гена , поскольку одна копия аллеля копируется из гомологичной хромосомы, а затем синтезируется в месте повреждения поврежденной хромосомы. Конечным результатом этого будет появление одной гетерозиготной хромосомы и одной гомозиготной хромосомы.
Преимущества и недостатки
[ редактировать ]Известно, что митотический кроссинговер происходит у D. melanogaster , некоторых грибов, размножающихся бесполым путем, и в нормальных клетках человека, где это событие может способствовать экспрессии обычно рецессивных аллелей, вызывающих рак, и, таким образом, предрасполагать клетку, в которой это происходит, к развитию рака . С другой стороны, клетка может стать гомозиготным мутантом по гену, подавляющему опухоль, что приведет к тому же результату. [2] Например, синдром Блума вызван мутацией хеликазы RecQ , которая играет роль в репликации и репарации ДНК. Эта мутация приводит к высокой скорости митотической рекомбинации у мышей, а эта скорость рекомбинации, в свою очередь, ответственна за предрасположенность к опухолям у этих мышей. [15] В то же время митотическая рекомбинация может быть полезной: она может играть важную роль в восстановлении двухцепочечных разрывов и может быть полезна для организма, если наличие гомозиготных доминантных аллелей более функционально, чем гетерозиготное состояние. [2] Для использования в экспериментах с геномами модельных организмов, таких как Drosophila melanogaster , митотическую рекомбинацию можно индуцировать с помощью рентгеновских лучей и системы рекомбинации FLP-FRT . [16]
Ссылки
[ редактировать ]- ^ Jump up to: а б Хартл, Дэниел Л. и Мэриеллен Руволо (2012). Генетика: анализ генетики и геномов . Берлингтон: Джонс и Бартлетт.
- ^ Jump up to: а б с Тишфилд, Джей А. (ноябрь 1997 г.). «Утрата гетерозиготности, или: Как я научился не волноваться и полюбил митотическую рекомбинацию» . Американский журнал генетики человека . 61 (5): 995–999. дои : 10.1086/301617 . ПМК 1716040 . ПМИД 9345110 .
- ^ Мойнахан М.Э., Джасин М. (2010). «Митотическая гомологичная рекомбинация поддерживает стабильность генома и подавляет онкогенез» . Нат. Преподобный мол. Клеточная Биол . 11 (3): 196–207. дои : 10.1038/nrm2851 . ПМК 3261768 . ПМИД 20177395 .
- ^ Стерн, Курт (1936). «Соматический кроссинговер и сегрегация у Drosophila Melanogaster» . Генетика . 21 (6): 625–730. дои : 10.1093/генетика/21.6.625 . ПМЦ 1208727 . ПМИД 17246815 .
- ^ Эспозито, Майкл С. (сентябрь 1978 г.). «Доказательства того, что спонтанная митотическая рекомбинация происходит на двухцепочечной стадии» . Труды Национальной академии наук США . 75 (9): 4436–4440. Бибкод : 1978PNAS...75.4436E . дои : 10.1073/pnas.75.9.4436 . ПМК 336130 . ПМИД 360220 .
- ^ Jump up to: а б Ли, Фиби С.; Гринвелл, Патрисия В.; Доминска, Маргарет; Гавель, Малгожата; Гамильтон, Моника; Питс, Томас Д. (2009). «Карта тонкой структуры спонтанных митотических кроссинговеров у дрожжей Saccharomyces cerevisiae» . ПЛОС Генет . 5 (3): e1000410. дои : 10.1371/journal.pgen.1000410 . ПМЦ 2646836 . ПМИД 19282969 .
- ^ Jump up to: а б ЛаФэйв, MC; Дж. Секельский (2009). «Митотическая рекомбинация: почему? Когда? Как? Где?» . ПЛОС Генет . 5 (3): e1000411. дои : 10.1371/journal.pgen.1000411 . ПМЦ 2648873 . ПМИД 19282976 .
- ^ Хейнс, Р. Х. и Кунц, бакалавр (1981). Репарация ДНК и мутагенез у дрожжей. В: Стратерн, Дж; Джонс, Э; Редакторы Броуча Дж. Молекулярная биология дрожжей Saccharomyces. Жизненный цикл и наследование. Колд-Спринг-Харбор, Нью-Йорк, Лаборатория Колд-Спринг-Харбор, 371-414.
- ^ Мэлоун Р.Э., Эспозито Р.Э. (1980). « Ген RAD5 2 необходим для гомоталличной взаимной конверсии типов спаривания и спонтанной митотической рекомбинации у дрожжей» . Учеб. Натл. акад. наук. США . 77 (1): 503–7. Бибкод : 1980ПНАС...77..503М . дои : 10.1073/pnas.77.1.503 . ПМК 348300 . ПМИД 6987653 .
- ^ Game JC, Замб Т.Дж., Браун Р.Дж., Резник М., Рот Р.М. (1980). «Роль генов радиации (rad) в мейотической рекомбинации у дрожжей» . Генетика . 94 (1): 51–68. дои : 10.1093/генетика/94.1.51 . ПМЦ 1214137 . ПМИД 17248996 .
- ^ Хелледей, Томас (2003). «Пути митотической гомологичной рекомбинации в клетках млекопитающих». Мутационные исследования . 532 (1–2): 103–115. дои : 10.1016/j.mrfmmm.2003.08.013 . ПМИД 14643432 .
- ^ Пакес, Фредерик; Джеймс Э. Хабер (1999). «Множественные пути рекомбинации, индуцированные двухцепочечными разрывами в Saccharomyces cerevisiae » . Обзоры микробиологии и молекулярной биологии . 63 (2): 349–404. дои : 10.1128/MMBR.63.2.349-404.1999 . ПМК 98970 . ПМИД 10357855 .
- ^ Хелледей, Т. «Ремонт двухцепочечного разрыва с помощью двойных соединений Холлидея (модель Шостака)» . Анимация . Массачусетский технологический институт.
- ^ Хелледей, Томас (27 ноября 2003 г.). «Пути митотической гомологичной рекомбинации в клетках млекопитающих» (PDF) . Мутационные исследования . 532 (1–2): 103–115. дои : 10.1016/j.mrfmmm.2003.08.013 . ПМИД 14643432 . Архивировано из оригинала (PDF) 24 мая 2014 года . Проверено 26 декабря 2012 г.
- ^ Ло, Гуанбинь; и др. (2000). «Предрасположенность к раку, вызванная повышенной митотической рекомбинацией у мышей Блума». Природная генетика . 26 (4): 424–429. дои : 10.1038/82548 . ПМИД 11101838 . S2CID 21218975 .
- ^ Сюй, Т; ГМ Рубин (апрель 1993 г.). «Анализ генетической мозаики в тканях развивающихся и взрослых дрозофил». Разработка . 117 (4): 1223–12237. дои : 10.1242/dev.117.4.1223 . ПМИД 8404527 .
- Гриффитс и др. 1999. Современный генетический анализ . WH Фриман и компания.