Зигомикота
Эта статья нуждается в дополнительных цитатах для проверки . ( март 2019 г. ) |
| Зигомикота | |
|---|---|
 | |
| Спорангий Phycomyces sp. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | Грибы |
| Разделение: | Зигомикота К. Моро, 1954 г. (неофициально) |
| Сорт: | Зигомицеты Зима 1881 года |
| Заказы | |
Zygomycota , или зиготные грибы , является бывшим подразделением или типом царства Грибов . Члены теперь являются частью двух типов : Mucoromycota и Zoopagomycota . [1] Известно около 1060 видов. [2] В своей среде обитания они в основном ведут наземный образ жизни, живя в почве или на разлагающемся растительном или животном материале. Некоторые из них являются паразитами растений, насекомых и мелких животных, а другие образуют симбиотические отношения с растениями. [3] зигомицетов Гифы могут быть ценоцитарными , образуя перегородки только там, где образуются гаметы , или отгораживать мертвые гифы. Zygomycota больше не признается, поскольку не считалось действительно монофилетическим видом . [1]
Этимология
[ редактировать ]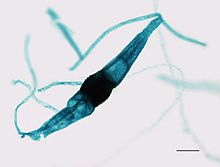
Название Zygomycota относится к зигоспорангиям, характерным для представителей этой клады, в которых устойчивые сферические споры образуются при половом размножении . Зигос по -гречески означает «соединение» или « ярмо », что означает слияние двух нитей гиф , в результате которого образуются эти споры, а -микота — суффикс, обозначающий разделение грибов. [4]
Споры
[ редактировать ]
Термин «спора» используется для описания структуры, связанной с распространением и распространением. Споры зигомицетов могут образовываться как половым, так и бесполым путем. До прорастания спора находится в состоянии покоя. В этот период скорость обмена веществ очень низкая и может длиться от нескольких часов до многих лет. Есть два типа покоя . Экзогенный покой контролируется факторами окружающей среды, такими как температура или доступность питательных веществ. Эндогенный или конститутивный покой зависит от характеристик самой споры; например, метаболические особенности. В этом типе покоя прорастание может быть предотвращено, даже если условия окружающей среды благоприятствуют росту.
Митоспоры
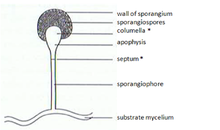
[ редактировать ]У зигомицетов митоспоры ( спорангиоспоры ) образуются бесполым путем. Они образуются в специализированных структурах — митоспорангиях (спорангиях), содержащих от нескольких до нескольких тысяч спор, в зависимости от вида. Митоспорангии переносят специализированные гифы — митоспорангиофоры (спорангиофоры). Эти специализированные гифы обычно демонстрируют отрицательный гравитропизм и положительный фототропизм, обеспечивающий хорошее распространение спор. Стенка спорангиев тонкая и легко разрушается под действием механических раздражителей (например, падающих капель дождя, проходящих мимо животных), что приводит к рассеиванию созревших митоспор. Стенки этих спор у некоторых видов содержат спорополленин . Спорополленин образуется из β-каротина и очень устойчив к биологическому и химическому разложению. Споры зигомицетов также можно классифицировать по их устойчивости:
Хламидоспоры
[ редактировать ]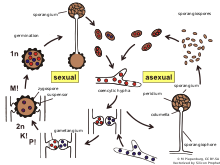
Хламидоспоры — это бесполые споры, отличные от спорангиоспор. Основная функция хламидоспор — сохранение мицелия, и они высвобождаются при разложении мицелия. Хламидоспоры не имеют механизма распространения. У зигомицетов образование хламидоспор обычно интеркалярное. Однако оно также может быть терминальным. В соответствии со своей функцией хламидоспоры имеют толстую клеточную стенку и пигментированы.
Зигофоры
[ редактировать ]Зигофоры представляют собой хемотропные воздушные гифы, являющиеся половыми органами зигомикот, за исключением Phycomyces, у которых они не воздушные, а находятся в субстрате. У них есть два разных типа спаривания (+) и (-). Противоположные типы спаривания растут навстречу друг другу за счет летучих феромонов, выделяемых противоположной цепью, в основном триспоровой кислоты и ее предшественников. Как только два противоположных типа спаривания вступают в первоначальный контакт, они в несколько этапов дают начало зигоспоре.
После контакта между зигофорами их стенки прилипают друг к другу, уплощаются, и тогда место контакта называется сращенной перегородкой. Кончики зигофора раздуваются и образуют так называемый прогаметангий. Перегородка развивается путем постепенного расширения внутрь, пока не отделит конечный гаметангий от прогаметангиального основания. В этот момент зигофор называется суспензором. Везикулы накапливаются у сращенной перегородки, после чего она начинает растворяться. Незадолго до полного растворения сращенной перегородки первичная наружная стенка начинает утолщаться. Это можно увидеть в виде темных пятен на первичной стенке по мере растворения сращенной перегородки. Эти темные пятна на стенке со временем разовьются в бородавчатые структуры, составляющие толщину стенки зигоспоры. По мере увеличения зигоспоры увеличиваются и бородавчатые структуры, пока они не станут прилегающими друг к другу вокруг всей клетки. В этот момент электронная микроскопия уже не может проникнуть через стену. Со временем бородавки прорастают через первичную стенку и темнеют, что, вероятно, вызвано меланин .
Мейоз обычно происходит до прорастания зигоспор, и существует несколько основных типов различимого ядерного поведения. Тип 1 — это когда ядра быстро, в течение нескольких дней, сливаются, в результате чего образуются зрелые зигоспоры с гаплоидными ядрами. При типе 2 некоторые ядра не спариваются и вместо этого дегенерируют, мейоз задерживается до прорастания. Тип 3 — это когда гаплоидные ядра продолжают делиться митотически, а затем некоторые объединяются в группы, а некоторые нет. В результате диплоидные обнаруживаются в зародышевых спорангиях и гаплоидные ядра .
Клеточная стенка
[ редактировать ]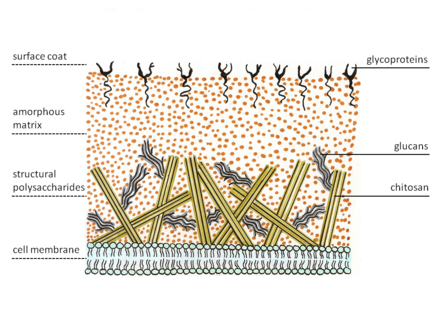
Зигомицеты обладают особым строением клеточной стенки. Большинство грибов имеют хитин в качестве структурного полисахарида , тогда как зигомицеты синтезируют хитозан , деацетилированный гомополимер хитина. Хитин построен из β-1,4-связанного N -ацетилглюкозамина. На кончике растут грибковые гифы. Таким образом, специализированные везикулы, хитосомы, выносят предшественники хитина и его синтезирующего фермента, хитинсинтетазы , наружу мембраны путем экзоцитоза . Фермент на мембране катализирует образование гликозидных связей из нуклеотидного сахарного субстрата, уридиндифосфо- N -ацетил-D-глюкозамина. Образовавшаяся полисахаридная цепь затем расщепляется ферментом хитиндеацетилазой . Фермент катализирует гидролитическое расщепление N -ацетамидной группы хитина. После этого полимерная цепь хитозана образует микрофибриллы . Эти волокна заключены в аморфную матрицу, состоящую из белков, глюканов (которые предположительно сшивают волокна хитозана), маннопротеинов , липидов и других соединений. [5] [6]
Триспоровая кислота
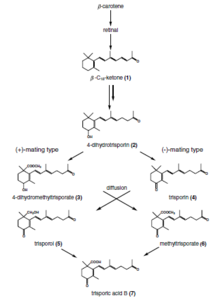
[ редактировать ]соединение C-18 Триспоровая кислота представляет собой терпеноидное , которое синтезируется β-каротина и ретинола в зигомицетах через пути . Это феромонное соединение, ответственное за половую дифференциацию этих видов грибов. [7]
История
[ редактировать ]Триспоровая кислота была обнаружена в 1964 году как метаболит, вызывающий повышенное производство каротина у Blakeslea trispora . Позже было показано, что именно этот гормон вызывает образование зигофоров у Mucor mucedo . [8] Американский миколог и генетик Альберт Фрэнсис Блейксли обнаружил, что некоторые виды Mucorales являются самостерильными ( гетероталлическими ), у которых для начала половой активности необходимы взаимодействия двух штаммов, обозначенных (+) и (-). Ганс Бургефф из Геттингенского университета обнаружил, что это взаимодействие происходит за счет обмена низкомолекулярными веществами, которые диффундируют через субстрат и атмосферу. Эта работа стала первой демонстрацией активности половых гормонов у любого гриба. Выяснение гормонального контроля полового взаимодействия у Mucorales продолжается уже более 60 лет, в нем участвуют микологи и биохимики из Германии, Италии, Нидерландов, Великобритании и США. [8]
Функции триспоровой кислоты в Mucorales
[ редактировать ]Распознавание совместимых половых партнеров у zygomycota основано на кооперативном пути биосинтеза триспоровой кислоты. Ранние производные триспороидов и триспоровая кислота вызывают набухание двух потенциальных гиф, поэтому называемых зигофорами, а химический градиент этих молекул-индукторов приводит к росту друг к другу. Эти прогаметангии соприкасаются друг с другом и создают прочную связь. На следующем этапе устанавливаются перегородки, ограничивающие развитие зигоспор из вегетативного мицелия, и таким образом зигофоры становятся суспензорными гифами и образуются гаметангии. После растворения стенки слияния цитоплазма и большое количество ядер обоих гаметангиев смешиваются. Процесс отбора (неизученный) приводит к редукции ядер и происходит мейоз (также неизученный до сих пор). несколько модификаций клеточной стенки, а также включение спорополленина Происходит (ответственного за темный цвет спор), что приводит к образованию зрелой зигоспоры.
Триспоровая кислота, как конечная точка этого пути распознавания, может вырабатываться только в присутствии обоих совместимых партнеров, которые ферментативно продуцируют предшественники триспороидов для дальнейшего использования потенциальным сексуальным партнером. Видовая специфичность этих реакций достигается, среди прочего, за счет пространственной сегрегации, физико-химических особенностей производных (летучесть и светочувствительность), химических модификаций триспороидов и регуляции транскрипции/посттранскрипции.
Парасексуализм
[ редактировать ]Триспороиды также используются в качестве посредника в распознавании между паразитом и хозяином. Примером может служить паразитарно-хозяинное взаимодействие парасексуальной природы, наблюдаемое между Parasitella parasitica , факультативным микопаразитом зигомицетов, и Absidia glauca . Это взаимодействие является примером паразитизма биотрофного слияния, поскольку генетическая информация передается хозяину. Видно много морфологических сходств по сравнению с образованием зигоспор, но зрелая спора называется сикиоспорой и является паразитической. производит галлоподобные структуры Во время этого процесса хозяин Absidia glauca .Это, в сочетании с дополнительными данными, привело к предположению, что триспороиды не являются строго видоспецифичными, но могут представлять собой общий принцип распознавания спаривания у Mucorales. [9]
Фототропизм
[ редактировать ]Регуляция света была исследована у зигомицетов Phycomyces blakesleeanus , Mucor circinelloides и Pilobolus Crystallinus . Например, у Pilobolus Crystallinus за механизм распространения отвечает свет, а спорангиофоры Phycomyces blakesleeanus растут в направлении света. Когда свет, особенно синий свет, участвует в регуляции развития грибов, он направляет рост грибковых структур и активирует метаболические пути. Например, зигомикоты используют свет как сигнал, способствующий вегетативному размножению и росту воздушных гиф, способствующих распространению спор.
Фототропизм грибов был подробно исследован на плодового тела спорангиофора Phycomyces модели . Phycomyces имеет сложную систему фоторецепторов. Он способен реагировать на разную интенсивность света и разные длины волн. В отличие от положительной реакции на синий свет, существует и отрицательная реакция на УФ-свет. Также наблюдались реакции на красный свет.
Активация биосинтеза бета-каротина светом
[ редактировать ]Два гена ферментов фитоендесатуразы (carB) и бифункционального фитоенасинтаза/каротинциклаза (carRA у Phycomyces , carRP у Mucor ) ответственны за синтез бета-каротина. Продукт гена crgA, обнаруженный у Мукора, подавляет образование каротина, ингибируя накопление мРНК carB и carRP.
Влияние света на споруляцию и половое развитие
[ редактировать ]Зигомицет P. blakesleeanus строит два типа спорангиофоров: макрофоры и микрофоры, которые различаются по размеру. Формирование этих спорангиофоров происходит при разных флюенсах света и, следовательно, с использованием специфических фоторецепторов. Свет также регулирует бесполое спорообразование. В Мукоре активатором выступает продукт гена crgA. Напротив, половое развитие Phycomyces тормозится светом из-за специализированной системы фоторецепторов.
Гравитропизм
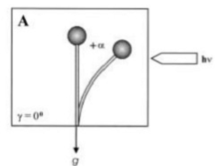
[ редактировать ]Гравитропизм — это поворотное или ростовое движение растения или гриба в ответ на силу тяжести. Он одинаково широко распространен в обоих королевствах. Статолиты необходимы как грибам, так и растениям для механизма определения гравитации. Спорангиефоры Zygomycota происходят из специализированных «базальных гиф» и проходят несколько различных стадий развития, пока не высвободятся зрелые бесполые споры. Помимо положительного фототропизма, спорангиофоры направляются за счет отрицательной гравитропной реакции в положение, подходящее для рассеивания и распределения спор. Оба ответа представляют собой реакции роста, т.е. изгиб вызван дифференциальным ростом на соответствующих противоположных сторонах спорангиеносца и влияют друг на друга. Единственная модель механизма гравитропной реакции Phycomyces основана на плавучести вакуоли внутри окружающей цитоплазмы. [10] Предполагается, что возникающее в результате асимметричное распределение цитоплазмы приводит к усиленному росту стенок на нижней стороне горизонтально расположенных спорангиофоров, поскольку в формирующемся там более толстом цитоплазматическом слое количество везикул, секретирующих материал клеточной стенки, будет выше, чем на верхней стороне. Гравитропное изгибание начинается примерно через 15–30 мин у горизонтально расположенных спорангиеносцев и продолжается до тех пор, пока примерно через 12–14 ч кончик спорангиеносца не вернется в исходное вертикальное положение. Обычно гравитропный ответ более слабый по сравнению с фототрофным. Однако в определенных условиях равновесие может быть установлено, и реакции сопоставимы. У растений и грибов фототропизм и гравитропизм взаимодействуют сложным образом. При непрерывном облучении односторонним светом спорангиофор (плодовое тело) зигомицета гриба Phycomyces blakesleeanus достигает угла изгиба фотогравитропного равновесия, при котором гравитропные и фототропные стимулы уравновешивают друг друга (рис. 1, угол изгиба +α, за счет света облучение [11] ).

Белковые кристаллы, участвующие в гравиперцепции
[ редактировать ]У Phycomyces blakesleeanus спорангиофоры дикого типа содержат крупные, хорошо различимые октаэдрические паракристаллы размером до 5×5×5 мкм. Обычно они встречаются вблизи главной вакуоли в скоплениях, состоящих более чем из десяти кристаллов. Они часто связаны с вакуолярными трансептами. Седиментацию со скоростью около 100 мкм/с можно наблюдать при наклоне спорангиофоров. Скольжение во время седиментации или растяжение вакуолярных мембран и трансептов служит межклеточным сигналом вероятной реакции цитоскелета и активирует рецепторы, расположенные в клеточной мембране. Эти рецепторы, в свою очередь, запускают цепочку событий, которая в конечном итоге приводит к асимметричному росту клеточной стенки. Исследования угла изгиба роста спорангиофоров дикого типа и мутантных штаммов показали, что мутантные штаммы, не имеющие кристаллов, проявляют сниженную гравитропную реакцию. [11]
Липидные капли, участвующие в гравиперцепции
[ редактировать ]Комплекс апикальных липидных глобул также участвует в гравиперцепции. Эти липиды группируются в клеточных структурах в виде комплекса липидных глобул примерно на 0,1 мм ниже самого кончика верхушки. (Рис. 2) При образовании спорангия глобулы мигрируют в колумеллу. Считается, что на зрелой стадии этот комплекс действует как гравирецептор из-за своей плавучести. Мутанты, у которых отсутствует этот липидный комплекс, демонстрируют значительно сниженную гравитропную реакцию. [10]
Филогения
[ редактировать ]Исторически все грибы, образующие зигоспоры, считались родственными и помещались в группу Zygomycota. Использование молекулярной филогенетики все чаще показывает, что эта группа является парафилетической . [1] Однако ранг (т. е. тип или подтип) этих клад является спорным. Далее следует филогения грибов с подтипом зигомицетов, полученная из Spatafora et al. (2016) [1] с обоими возможными названиями типов.
| |||||||||||||||||||
| |||||||||||||||||||
Промышленное использование
[ редактировать ]Многие виды зигомицетов могут использоваться в важных промышленных процессах. Их резюме представлено в таблице.
| Разновидность | Продукт | Использование |
|---|---|---|
| Несколько видов Mucor и Rhizopus . | Липазы и протеазы | Кожаная, моющая и медицинская промышленность (трансформация стероидов) |
| Ризопус | Целлюлазы | Производство продуктов питания (например, темпе) |
| R. oryzae , другие виды Rhizopus . | Фумаровая кислота | Разнообразный |
| виды Rhizopus . | Молочная кислота | Разнообразный |
| Р. Делемар | Биотин | Разнообразный |
| Mortierella romanniana , Mortierella vinacea и Mucor indicus. | Линоленовая кислота | Разнообразный |
| Альпийская Мортьерелла | Арахидоновая кислота | Разнообразный |
| Блейксли триспора | β-каротин | Разнообразный |
Условия культивирования
[ редактировать ]
Зигомицеты способны расти в самых разных средах. Большинство из них мезофильны (растут при 10–40 °C с оптимумом 20–35 °C), но некоторые, например Mucor miehei или Mucor pusillus , термофильны с минимальной температурой роста около 20 °C и максимальной до 20 °C. 60 °С. Другие, такие как Mucor hiemalis, могут расти при температуре ниже 0 °C.
Некоторые виды отряда Mucorales способны расти в анаэробных условиях, тогда как большинству из них необходимы аэробные условия. Более того, хотя большинство зигомицетов растут только при высокой активности воды, некоторые из них способны расти при концентрации соли не менее 15%. Большинство видов Mucor быстро растут на агаре при комнатной температуре, заполняя чашку Петри за 2–3 дня грубым воздушным мицелием. При инкубации в жидкой культуре в полуанаэробных условиях некоторые виды растут в дрожжеподобном состоянии. Образование зигоспор может стимулироваться при более высоких температурах инкубации (30–40 °C).

Рост Zygomycota в твердом агаре может привести к образованию колоний с низким или очень высоким содержанием волокон, которые быстро заполняют всю чашку Петри. Его цвет может варьироваться от чисто белого до оттенков серого или коричневого. В старых культурах наблюдаются темнопигментированные спорангии. Все зависит от вида и используемой среды. В жидкой культуре Zygomycota обычно образует мягкую массу и не образует спор. Это потому, что они не могут выращивать воздушные гифы.
Культурные СМИ
[ редактировать ]Зигомицеты хорошо растут на большинстве стандартных сред для культивирования грибов , таких как декстрозный агар Сабуро. Они также могут расти как на селективных, так и на неселективных средах. Также можно использовать минимальные среды, дополнительные среды и индукционные среды. Большинство зигомицетов чувствительны к циклогексимиду (актидиону), и этот агент не следует использовать в культуральных средах.
Воспроизведение
[ редактировать ]Распространенным примером зигомицета является черная хлебная плесень ( Rhizopus stolonifer ), представитель рода Mucorales . Он распространяется по поверхности хлеба и других источников пищи, направляя гифы внутрь для поглощения питательных веществ. В бесполой развиваются луковицеобразные черные спорангии фазе на кончиках прямостоячих гиф , каждая из которых содержит сотни гаплоидных спор .
Как и у большинства зигомицетов, бесполое размножение является наиболее распространенной формой размножения. Половое размножение у Rhizopus stolonifer , как и у других зигомицетов, происходит при нахождении гаплоидных гиф разных типов спаривания в непосредственной близости друг к другу. Рост гаметангиев начинается после соприкосновения гаметангиев и происходит плазмогамия , или слияние цитоплазмы. Сразу за этим следует кариогамия , то есть слияние ядер. Зигоспорангии тогда диплоидны . Зигоспорангии обычно толстостенные, очень устойчивы к неблагоприятным воздействиям окружающей среды и метаболически инертны. Однако когда условия улучшаются, они прорастают, образуя спорангий или вегетативные гифы . Мейоз происходит во время прорастания зигоспорангия, поэтому образующиеся споры или гифы гаплоидны. Растет в теплых и влажных условиях.
Некоторые зигомицеты рассеивают свои споры более точным образом, чем просто позволяя им бесцельно дрейфовать в потоках воздуха. Pilobolus , гриб, который растет на навозе животных, изгибает свои спорангиофоры в сторону света с помощью светочувствительного пигмента ( бета-каротина ), а затем «выстреливает» их взрывной струей цитоплазмы под высоким давлением . Спорангии можно запускать на расстояние до 2 м, располагая их подальше от навоза и, возможно, на растительности, которая будет съедена травоядным животным и в конечном итоге отложена вместе с навозом в другом месте. Среди представителей отряда зигомицетов Entomophthorales развились различные механизмы принудительного выброса спор .
Эволюция конидий
[ редактировать ]Эволюция конидия из спорангиоспоры является основным определяющим различием между зигомицетами и аскомицетами . [12] Эволюцию спорангиоспор, типичных для зигомицетов, в конидии, подобные тем, что встречаются у аскомицетов, можно смоделировать рядом форм, наблюдаемых у зигомицетов. Многие зигомицеты образуют несколько спорангиоспор внутри одного спорангия. У некоторых развились множественные мелкие спорангиолы, содержащие мало спорангиоспор. В некоторых случаях в каждом спорангиоле может быть от нескольких до трех спор, а у некоторых видов есть спорангиолы, содержащие только одну спору. Хоанефора , зигомицет, имеет спорангиол, содержащий одну спору со стенкой спорангия, видимой у основания спорангия. Эта структура похожа на конидий, который имеет две сросшиеся клеточные стенки: внутреннюю стенку спор и внешнюю стенку спорангия.
Ссылки
[ редактировать ]- ^ Jump up to: а б с д Спатафора, Джозеф В.; Чанг, Ин; Бенни, Джеральд Л.; Лазарь, Кэти; Смит, Мэтью Э.; Берби, Мэри Л.; Бонито, Грегори; Корради, Николас; Григорьев Игорь; Грыганский Андрей; Джеймс, Тимоти Ю.; О'Доннелл, Керри; Роберсон, Роберт В.; Тейлор, Томас Н.; Юлинг, Джесси; Вилгалис, Ритас; Уайт, Мерлин М.; Стаич, Джейсон Э. (2016). «Филогенетическая классификация зигомицетных грибов на уровне типа, основанная на данных масштаба генома» . Микология . 108 (5): 1028–1046. дои : 10.3852/16-042 . ISSN 0027-5514 . ПМК 6078412 . ПМИД 27738200 .
- ^ Крог, Дэвид (2010). Биология: Путеводитель по миру природы . Бенджамин Каммингс . п. 409. ИСБН 978-0-321-61655-5 .
- ^ Рэйвен, штат Пенсильвания; Эверт, РФ; Эйххорн, SE (2005). «Грибки». Биология растений (7-е изд.). У. Х. Фриман. стр. 268–9 . ISBN 978-0716762843 .
- ^ Дэвид Мур; Джеффри Д. Робсон; Энтони Пи Джей Тринчи (14 июля 2011 г.). Путеводитель по грибам XXI века . Издательство Кембриджского университета. п. 52. ИСБН 978-1-107-00676-8 .
- ^ Гоу, Нил А.Р.; Гэдд, Джеффри М. , ред. (1995). Выращивание гриба . Спрингер. ISBN 978-0-412-46600-7 .
- ^ Уоткинсон, Сара С.; Бодди, Линн; Деньги, Николас (2015). Грибы (3-е изд.). Академическая пресса. ISBN 978-0-12-382035-8 .
- ^ Добрый день, Грэм В.; Карлайл, Майкл Дж. (август 1997 г.). «Открытие грибковых половых гормонов: III. Триспоровая кислота и ее предшественники». Миколог . 11 (3): 126–130. дои : 10.1016/S0269-915X(97)80017-1 .
- ^ Jump up to: а б Шульце, Корнелия; Шимек, Кристина; Вёстемайер, Йоханнес; Бурместер, Анке (2005). «Сексуальность и паразитизм имеют общие пути регуляции у гриба Parasitella parasitica ». Джин . 348 : 33–44. дои : 10.1016/j.gene.2005.01.007 . ПМИД 15777660 .
- ^ Шимек, Кристина; Клеппе, Катрин; Салим, Абдель-Рахман; Фойгт, Керстин; Бурместер, Анке; Вёстемайер, Йоханнес (2003). «Сексуальные реакции у Mortierellales опосредуются системой триспоровой кислоты». Микологические исследования . 107 (6): 736–747. дои : 10.1017/S0953756203007949 . ПМИД 12951800 .
- ^ Jump up to: а б Гролиг Ф., Херкенрат Х., Пумм Т., Гросс А., Галланд П. (февраль 2004 г.). «Гравитационная подвеска за счет плавучести: плавающие липидные глобулы в спорангиофоре фикомицетов». Планта . 218 (4): 658–667. дои : 10.1007/s00425-003-1145-x . ПМИД 14605883 . S2CID 21460919 .
- ^ Jump up to: а б Шимек С., Эйбе П., Хориэль Т., Галланд П., Оотаки Т. (1999). «Белковые кристаллы в спорангиофорах фикомицетов участвуют в гравиперцепции». Достижения в космических исследованиях . 24 (6): 687–696. Бибкод : 1999AdSpR..24..687S . дои : 10.1016/S0273-1177(99)00400-7 . ПМИД 11542610 .
- ^ Каин, РФ (1972). «Эволюция грибов». Микология . 64 (1): 1–14. дои : 10.2307/3758010 . JSTOR 3758010 .
Внешние ссылки
[ редактировать ]- Zygomycota в веб-проекте «Древо жизни»
- Zygomycetes.org
- Список всех видов зигомицетов из базы данных зигомицетов П.М. Кирка в Каталоге жизни 2008 г.
- Мукоралес в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
| Базы данных органов управления : Национальные |
|---|