Гетероталлизм
Гетероталлические виды имеют полы , присущие разным особям. Этот термин применяется, в частности, для отличия гетероталличных грибов , которым для образования половых спор требуются два совместимых партнера, от гомоталличных , способных к половому размножению из одного организма.
У гетероталличных грибов два разных человека вносят ядра для образования зиготы. Примеры гетероталлизма включают Saccharomyces cerevisiae , Aspergillus fumigatus , Aspergillus flavus , Penicillium marneffei и Neurospora crassa . Гетероталлический жизненный цикл N. crassa приведен довольно подробно, поскольку сходные жизненные циклы присутствуют и у других гетероталличных грибов.
Жизненный цикл Saccharomyces cerevisiae
[ редактировать ]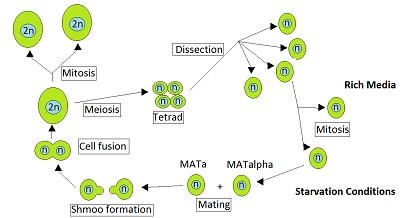
Дрожжи Saccharomyces cerevisiae гетероталличны. Это означает, что каждая дрожжевая клетка относится к определенному типу спаривания и может спариваться только с клеткой другого типа спаривания. Во время вегетативного роста, который обычно происходит при избытке питательных веществ, S. cerevisiae размножается путем митоза в виде гаплоидных или диплоидных клеток. Однако при голодании диплоидные клетки подвергаются мейозу с образованием гаплоидных спор. [1] Спаривание происходит, когда гаплоидные клетки противоположного типа спаривания, MATa и MATα, вступают в контакт. Рудерфер и др. [2] отметил, что такие контакты часто происходят между близкородственными дрожжевыми клетками по двум причинам. Во-первых, клетки противоположного типа спаривания присутствуют вместе в одной сумке , мешочке, который содержит тетраду клеток, непосредственно образующихся в результате одного мейоза , и эти клетки могут спариваться друг с другом. Вторая причина заключается в том, что гаплоидные клетки одного типа спаривания при делении клеток часто производят клетки противоположного типа спаривания, с которыми они могут спариваться.
Кац Эзов и др. [3] представили данные о том, что в природных популяциях S. cerevisiae преобладают клональное размножение и тип «самооплодотворения» (в форме интратетрадного спаривания). Рудерфер и др. [2] проанализировали происхождение природных штаммов S. cerevisiae и пришли к выводу, что ауткроссинг происходит только примерно один раз на каждые 50 000 клеточных делений. Таким образом, хотя S. cerevisiae является гетероталличным, оказывается, что в природе спаривание чаще всего происходит между близкородственными дрожжевыми клетками. Относительная редкость в природе мейотических событий, возникающих в результате ауткроссинга, позволяет предположить, что возможные долгосрочные преимущества ауткроссинга (например, создание генетического разнообразия ) вряд ли будут достаточными для общего сохранения пола от одного поколения к другому. [ нужна ссылка ] Скорее, краткосрочная польза, такая как мейотическая рекомбинационная репарация повреждений ДНК, вызванных стрессовыми условиями, такими как голодание, может быть ключом к поддержанию пола у S. cerevisiae . [4] [5]
Жизненный цикл Aspergillus fumigatus
[ редактировать ]Aspergillus fumigatus — гетероталличный гриб. [6] Это один из наиболее распространенных видов Aspergillus, вызывающих заболевания у людей с иммунодефицитом . A. fumigatus широко распространен в природе и обычно встречается в почве и разлагающихся органических веществах, таких как компостные кучи, где он играет важную роль в углерода и азота переработке . Колонии гриба производят из конидиеносцев тысячи мельчайших серо-зеленых конидий (2–3 мкм), которые легко передаются по воздуху. A. fumigatus обладает полностью функциональным половым репродуктивным циклом, который приводит к образованию клейстотеций и аскоспор . [7]
Хотя A. fumigatus встречается в районах с совершенно разным климатом и окружающей средой, он демонстрирует низкую генетическую изменчивость и отсутствие генетической дифференциации популяций в глобальном масштабе. [8] Таким образом, способность к гетероталличному сексу сохраняется, даже несмотря на небольшое генетическое разнообразие. Как и в случае с S. cereviae , описанном выше, краткосрочная выгода от мейоза может быть ключом к адаптивному поддержанию пола у этого вида.
Жизненный цикл Aspergillus flavus
[ редактировать ]A. flavus является основным производителем канцерогенных афлатоксинов в сельскохозяйственных культурах во всем мире. Это также условно- патогенный микроорганизм человека и животных , вызывающий аспергиллез у лиц с ослабленным иммунитетом. В 2009 году было обнаружено, что половое состояние этого гетероталличного гриба возникает при совместном культивировании штаммов противоположного типа спаривания в соответствующих условиях. [9]
Сексуальность порождает разнообразие в кластере генов афлатоксина у A. flavus . [10] предполагая, что возникновение генетических вариаций может способствовать поддержанию гетероталлизма у этого вида.
Жизненный цикл Talaromyces marneffei
[ редактировать ]Хенк и др. [11] показали, что гены, необходимые для мейоза, присутствуют у T. marneffei и что у этого вида происходят спаривание и генетическая рекомбинация.
Хенк и др. [11] пришли к выводу, что T. marneffei размножается половым путем, но рекомбинация в естественных популяциях, скорее всего, будет происходить на пространственно и генетически ограниченных расстояниях, что приводит к сильно клональной структуре популяции. У этого вида сохраняется пол, хотя генетическая изменчивость очень незначительна . Пол может поддерживаться у T. marneffei за счет кратковременного эффекта мейоза, как у S. cerevisiae и A. fumigatus , обсуждавшихся выше.
Жизненный цикл Neurospora crassa
[ редактировать ]
Половой цикл N. crassa гетероталличный. Половые плодовые тела (перитеции) могут образоваться только при сближении двух мицелий разного типа спаривания. Как и другие аскомицеты , N. crassa имеет два типа спаривания, которые в данном случае обозначаются буквами «А» и «а». Между штаммами типов спаривания «А» и «А» нет очевидной морфологической разницы. Оба могут образовывать обильные протоперитеции — женскую репродуктивную структуру (см. рисунок вверху § ). Протоперитеции легче всего образуются в лаборатории, когда рост происходит на твердой (агаровой) синтетической среде с относительно низким содержанием азота. [12] Азотное голодание, по-видимому, необходимо для экспрессии генов, участвующих в половом развитии. [13] Протоперитеций состоит из аскогония — спиральной многоклеточной гифы, заключенной в узелковое скопление гиф. Разветвленная система тонких гиф, называемая трихогиной, простирается от кончика аскогония, выступая за пределы покрывающих гиф в воздух. Половой цикл инициируется (т. е. происходит оплодотворение), когда клетка (обычно конидия) противоположного типа спаривания контактирует с частью трихогины (см. рисунок вверху § ). За таким контактом может последовать слияние клеток, приводящее к тому, что одно или несколько ядер оплодотворяющей клетки мигрируют по трихогине в аскогоний. Поскольку штаммы «А» и «А» имеют одинаковую половую структуру, ни один из штаммов не может считаться исключительно мужским или женским. Однако в качестве реципиента протоперитеций штаммов «А» и «А» можно рассматривать как женскую структуру, а оплодотворяющий конидий можно рассматривать как участника мужского пола.
Последующие этапы после слияния гаплоидных клеток «А» и «а» были изложены Финчемом и Деем. [14] и Вагнер и Митчелл. [15] После слияния клеток дальнейшее слияние их ядер задерживается. Вместо этого ядро оплодотворяющей клетки и ядро аскогония связываются и начинают синхронно делиться. Продукты этих ядерных делений (еще парами разного типа спаривания, т. е. «А» / «а») мигрируют в многочисленные аскогенные гифы, которые затем начинают расти из аскогония. Каждая из этих аскогенных гиф изгибается, образуя крючок (или посох) на своем кончике, и пара гаплоидных ядер «А» и «а» внутри посоха делится синхронно. Далее формируются перегородки, разделяющие посох на три ячейки. Центральная ячейка изгиба крючка содержит одно ядро «А» и одно ядро «а» (см. рисунок вверху § ). Эта двуядерная клетка инициирует образование сумки и называется «инициальной клеткой аски». Затем две одноядерные клетки по обе стороны от первой клетки, образующей аск, сливаются друг с другом, образуя двуядерную клетку , которая может расти с образованием еще одной клетки, образующей аску, которая затем может сформировать свою собственную начальную клетку аска. Затем этот процесс можно повторить несколько раз.
После образования аск-инициальной клетки ядра «А» и «а» сливаются друг с другом, образуя диплоидное ядро (см . рисунок вверху § ). Это ядро — единственное диплоидное ядро за весь жизненный цикл N. crassa . Диплоидное ядро состоит из 14 хромосом, образовавшихся из двух сросшихся гаплоидных ядер, каждое из которых содержало по 7 хромосом. За образованием диплоидного ядра сразу же следует мейоз . Два последовательных деления мейоза приводят к образованию четырех гаплоидных ядер: двух типа спаривания «А» и двух типа спаривания «а». Еще одно митотическое деление приводит к образованию четырех ядер «А» и четырех ядер «А» в каждой сумке . Мейоз является неотъемлемой частью жизненного цикла всех организмов, размножающихся половым путем, и по своим основным признакам мейоз у N. crassa кажется типичным для мейоза в целом.
При возникновении вышеописанных событий мицелиальная оболочка, окружавшая аскогоний, развивается в стенку перитеция, пропитывается меланином и чернеет. Зрелый перитеций имеет колбовидное строение.
Зрелый перитеций может содержать до 300 аск, каждая из которых происходит из идентичных слитых диплоидных ядер. Обычно в природе при созревании перитеций аскоспоры довольно сильно выбрасываются в воздух. Эти аскоспоры термостойки, и в лаборатории для их прорастания требуется нагревание при температуре 60 °C в течение 30 минут. Для нормальных штаммов весь половой цикл занимает от 10 до 15 дней. У зрелой аски, содержащей 8 аскоспор, пары соседних спор идентичны по генетическому строению, так как последнее деление митотическое, а аскоспоры содержатся в мешке аскоспоры, удерживающем их в определенном порядке, определяемом направлением ядерных сегрегаций во время мейоз. Поскольку четыре первичных продукта также расположены последовательно, структуру генетических маркеров сегрегации первого деления можно отличить от маркеров модели сегрегации второго деления.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Герсковиц I (декабрь 1988 г.). «Жизненный цикл почкующихся дрожжей Saccharomyces cerevisiae» . Микробиол. Преподобный . 52 (4): 536–53. дои : 10.1128/MMBR.52.4.536-553.1988 . ПМЦ 373162 . ПМИД 3070323 .
- ^ Jump up to: а б Рудерфер Д.М., Пратт С.С., Зайдель Х.С., Кругляк Л. (сентябрь 2006 г.). «Популяционно-геномный анализ ауткроссинга и рекомбинации у дрожжей». Нат. Жене . 38 (9): 1077–81. дои : 10.1038/ng1859 . ПМИД 16892060 . S2CID 783720 .
- ^ Кац Эзов Т., Чанг С.Л., Френкель З., Сегре А.В., Бахалул М., Мюррей А.В., Леу Дж.Ю., Король А., Каши Ю. (январь 2010 г.). «Гететаллизм у изолятов Saccharomyces cerevisiae из природы: влияние локуса HO на способ размножения» . Мол. Экол . 19 (1): 121–31. дои : 10.1111/j.1365-294X.2009.04436.x . ПМЦ 3892377 . ПМИД 20002587 .
- ^ Бердселл Дж. А., Уиллс С. (2003). Эволюционное происхождение и поддержание половой рекомбинации: обзор современных моделей. Серия «Эволюционная биология» >> Эволюционная биология, Vol. 33 стр. 27–137. Макинтайр, Росс Дж.; Клегг, Майкл, Т. (ред.), Спрингер. ISBN 978-0306472619
- ^ Эльвира Хёрандл (2013). Мейоз и парадокс пола в природе, Мейоз, ISBN 978-953-51-1197-9 , ИнТех, DOI: 10.5772/56542
- ^ Суги Дж.А., Лосада Л., Ван В., Варга Дж., Нгамскулрунгрой П., Абу-Асаб М., Чанг Ю.К., О'Горман С.М., Уикс Б.Л., Нирман В.К., Дайер П.С., Квон-Чунг К.Дж. (2011). «Идентификация и характеристика пары «суперматер» Aspergillus fumigatus» . мБио . 2 (6): e00234–11. дои : 10.1128/mBio.00234-11 . ПМК 3225970 . ПМИД 22108383 .
- ^ О'Горман СМ, Фуллер Х., Дайер П.С. (январь 2009 г.). «Открытие полового цикла у условно-патогенного грибкового возбудителя Aspergillus fumigatus». Природа . 457 (7228): 471–4. Бибкод : 2009Natur.457..471O . дои : 10.1038/nature07528 . ПМИД 19043401 . S2CID 4371721 .
- ^ Ридхольм К., Сакач Г., Лутзони Ф. (апрель 2006 г.). «Низкая генетическая изменчивость и отсутствие обнаруживаемой структуры популяции у aspergillus fumigatus по сравнению с близкородственными видами Neosartorya» . Эукариотическая клетка . 5 (4): 650–7. doi : 10.1128/EC.5.4.650-657.2006 . ПМК 1459663 . ПМИД 16607012 .
- ^ Хорн Б.В., Мур Г.Г., Карбоне I (2009). «Половое размножение Aspergillus flavus» . Микология . 101 (3): 423–9. дои : 10.3852/09-011 . ПМИД 19537215 . S2CID 20648447 .
- ^ Мур Г.Г., Эллиотт Дж.Л., Сингх Р., Хорн Б.В., Дорнер Дж.В., Стоун Е.А., Чулз С.Н., Баррос Г.Г., Найк МК, Райт Г.К., Хелл К., Карбоне I (2013). «Сексуальность порождает разнообразие в кластере генов афлатоксина: доказательства в глобальном масштабе» . ПЛОС Патог . 9 (8): e1003574. дои : 10.1371/journal.ppat.1003574 . ПМЦ 3757046 . ПМИД 24009506 .
- ^ Jump up to: а б Хенк Д.А., Шахар-Голан Р., Деви К.Р., Бойс К.Дж., Жан Н., Федорова Н.Д., Нирман В.К., Сюэ П.Р., Юэн К.Ю., Сьеу Т.П., Кинь Н.В., Вертхайм Х., Бейкер С.Г., Дэй Дж.Н., Ваниттанаком Н., Бигнелл Э.М., Андрианопулос А., Фишер MC (2012). «Клональность независимо от пола: эволюция связанных с хозяином сексуальных соседств у патогенного гриба Penicillium marneffei» . ПЛОС Патог . 8 (10): e1002851. дои : 10.1371/journal.ppat.1002851 . ПМЦ 3464222 . ПМИД 23055919 .
- ^ Вестергаард М, Митчелл Х.К. (1947). «Нейроспора. Часть V. Синтетическая среда, способствующая половому размножению». Американский журнал ботаники . 34 : 573–577. дои : 10.1002/j.1537-2197.1947.tb13032.x .
- ^ Нельсон М.А., Метценберг Р.Л. (сентябрь 1992 г.). «Гены полового развития Neurospora crassa» . Генетика . 132 (1): 149–162. дои : 10.1093/генетика/132.1.149 . ПМК 1205113 . ПМИД 1356883 .
- ^ Финчем Дж. Р. С., День PR (1963). Грибковая генетика . Оксфорд, Великобритания: Научные публикации Блэквелла. АСИН B000W851KO .
- ^ Вагнер Р.П., Митчелл Х.К. (1964). Генетика и обмен веществ . Нью-Йорк, штат Нью-Йорк: Джон Уайли и сыновья. ASIN B00BXTC5BO .