Аспергилл фумигатус
| Аспергилл фумигатус | |
|---|---|
 | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | Грибы |
| Разделение: | Аскомикота |
| Сорт: | Евротиомицеты |
| Заказ: | Евротиалес |
| Семья: | Аспергилловые |
| Род: | Аспергилл |
| Разновидность: | А. фумигатус |
| Биномиальное имя | |
| Аспергилл фумигатус Фрезениус 1863 г. | |
| Синонимы | |
Неосартория фумигата | |
Aspergillus fumigatus — вид грибов рода Aspergillus , один из наиболее распространенных видов Aspergillus, вызывающих заболевания у людей с иммунодефицитом .
Aspergillus fumigatus , сапротроф , широко распространенный в природе, обычно встречается в почве и разлагающихся органических веществах, таких как компостные кучи, где он играет важную роль в углерода и азота . переработке [1] Колонии гриба образуются из конидиеносцев ; тысячи мельчайших серо-зеленых конидий (2–3 мкм), которые легко передаются по воздуху. В течение многих лет A. fumigatus считалось, что размножается только бесполым путем, поскольку ни спаривания, ни мейоза никогда не наблюдалось. В 2008 году было показано, что A. fumigatus обладает полностью функциональным половым репродуктивным циклом, спустя 145 лет после его первоначального описания Фрезениусом. [2] Хотя A. fumigatus встречается в районах с совершенно разным климатом и окружающей средой, он демонстрирует низкую генетическую изменчивость и отсутствие генетической дифференциации популяций в глобальном масштабе. [3] Таким образом, способность к сексу сохраняется, хотя генетические вариации возникают незначительно.
Гриб способен расти при температуре 37 °C или 99 °F ( нормальная температура человеческого тела ) и может расти при температуре до 50 °C или 122 °F, при этом конидии выживают при 70 °C или 158 °F — при определенных условиях. регулярно встречается в самонагревающихся компостных кучах. Его споры повсеместно распространены в атмосфере, и каждый вдыхает примерно несколько сотен спор каждый день; как правило, они быстро устраняются иммунной системой у здоровых людей. У людей с ослабленным иммунитетом , таких как реципиенты трансплантатов органов и люди со СПИДом или лейкемией , грибок с большей вероятностью станет патогенным , преодолевая ослабленную защиту хозяина и вызывая ряд заболеваний, обычно называемых аспергиллезом . В связи с недавним увеличением использования иммунодепрессантов для лечения заболеваний человека, по оценкам, A. fumigatus может быть причиной более 600 000 смертей ежегодно с уровнем смертности от 25 до 90%. [4] Было высказано предположение о нескольких факторах вирулентности , объясняющих такое оппортунистическое поведение. [5]
При скрининге ферментационного бульона A. fumigatus был обнаружен ряд индольных алкалоидов с антимитотическими свойствами. [6] Интересующие соединения относятся к классу, известному как трипростатин, причем спиротрипростатин B представляет особый интерес как противораковый препарат.
Aspergillus fumigatus, выращенный на некоторых строительных материалах, может продуцировать генотоксичные и цитотоксические микотоксины , такие как глиотоксин . [7]
Геном
[ редактировать ]Aspergillus fumigatus имеет стабильный гаплоидный геном из 29,4 миллионов пар оснований . Последовательности генома трех Aspergillus видов — Aspergillus fumigatus , Aspergillus nidulans и Aspergillus oryzae — были опубликованы в журнале Nature в декабре 2005 года. [8] [9] [10]
Патогенез
[ редактировать ]Aspergillus fumigatus является наиболее частой причиной инвазивной грибковой инфекции у лиц с иммуносупрессией, к которым относятся пациенты, получающие иммуносупрессивную терапию по поводу аутоиммунных или неопластических заболеваний, реципиенты трансплантатов органов и пациенты со СПИДом. [11] A. fumigatus в первую очередь вызывает инвазивную инфекцию легких и является основной причиной заболеваемости и смертности у этих людей. [12] Кроме того, A. fumigatus может вызывать хронические легочные инфекции, аллергический бронхолегочный аспергиллез или аллергические заболевания у иммунокомпетентных хозяев. [13]
Врожденный иммунный ответ
[ редактировать ]Ингаляционное воздействие конидий, передающихся по воздуху, является постоянным из-за их повсеместного распространения в окружающей среде. Однако у здоровых людей врожденная иммунная система является эффективным барьером на пути заражения A. fumigatus . [13] Большая часть вдыхаемых конидий очищается мукоцилиарным действием респираторного эпителия. [13] Из-за небольшого размера конидий многие из них откладываются в альвеолах , где взаимодействуют с эпителиальными и врожденными эффекторными клетками. [11] [13] Альвеолярные макрофаги фагоцитируют и разрушают конидии внутри своих фагосом . [11] [13] Эпителиальные клетки, особенно пневмоциты типа II, также интернализируют конидии, которые попадают в лизосому , где проглоченные конидии разрушаются. [11] [13] [14] Иммунные клетки первой линии также служат для рекрутирования нейтрофилов и других воспалительных клеток посредством высвобождения цитокинов и хемокинов, индуцированного лигированием специфических грибковых мотивов с рецепторами распознавания патогенов . [13] Нейтрофилы необходимы для устойчивости к аспергиллезу, как показано у лиц с нейтропенией, и способны изолировать как конидии, так и гифы посредством различных, нефагоцитарных механизмов. [11] [12] [13] Гифы слишком велики для клеточной интернализации, и, таким образом, повреждение, вызванное НАДФН-оксидазой, опосредованное нейтрофилами , представляет собой доминирующую защиту хозяина от гиф. [11] [13] В дополнение к этим клеточно-опосредованным механизмам элиминации антимикробные пептиды, секретируемые эпителием дыхательных путей, способствуют защите хозяина. [11] Гриб и его полисахариды обладают способностью регулировать функции дендритных клеток путем Сигнальный путь Wnt-β-катенин для индукции PD-L1 и стимулирования регуляторных Т-клеточных ответов [15] [16]
Вторжение
[ редактировать ]
Лица с ослабленным иммунитетом восприимчивы к инвазивной инфекции A. fumigatus , которая чаще всего проявляется как инвазивный легочный аспергиллез. Вдыхаемые конидии, уклоняющиеся от разрушения иммунной системы хозяина, являются прародителями инвазивных заболеваний. Эти конидии выходят из состояния покоя и морфологически превращаются в гифы, прорастая в теплой, влажной, богатой питательными веществами среде легочных альвеол. [11] Прорастание происходит как внеклеточно, так и в эндосомах пневмоцитов II типа, содержащих конидии. [11] [14] После прорастания нитевидный рост гиф приводит к проникновению в эпителий и последующему проникновению в эндотелий сосудов. [11] [14] Процесс ангиоинвазии вызывает повреждение эндотелия и индуцирует провоспалительную реакцию, экспрессию тканевых факторов и активацию каскада свертывания крови . [11] Это приводит к внутрисосудистому тромбозу и локализованному инфаркту тканей , однако распространение фрагментов гиф обычно ограничено. [11] [14] Распространение через кровоток происходит только у лиц с тяжелым иммунодефицитом. [14]
Реакция на гипоксию
[ редактировать ]Как это часто бывает с опухолевыми клетками и другими патогенами, инвазивные гифы A. fumigatus сталкиваются с гипоксической (низкий уровень кислорода, ≤ 1%) микросредой в месте заражения в организме хозяина. [17] [18] [19] Текущие исследования показывают, что при инфекции некроз и воспаление вызывают повреждение тканей, что снижает концентрацию доступного кислорода из-за местного снижения перфузии , прохождения жидкости к органам. В частности, было обнаружено, что у A. fumigatus вторичные метаболиты ингибируют развитие новых кровеносных сосудов, что приводит к повреждению тканей, ингибированию восстановления тканей и, в конечном итоге, к возникновению локализованной гипоксической микросреды. [18] Точные последствия гипоксии для патогенеза грибков в настоящее время неизвестны, однако среда с низким содержанием кислорода уже давно связана с отрицательными клиническими исходами. Из-за выявленных значительных корреляций между гипоксией, грибковыми инфекциями и отрицательными клиническими исходами механизмы, с помощью которых A. fumigatus адаптируется к гипоксии, становятся все более объектом внимания для новых целевых лекарственных препаратов.
Было показано, что два хорошо изученных белка, связывающего стерол-регуляторные элементы, SrbA и SrbB, а также пути их процессинга влияют на приспособленность A. fumigatus в условиях гипоксии. Фактор транскрипции SrbA является главным регулятором реакции грибов на гипоксию in vivo и играет важную роль во многих биологических процессах, включая гомеостаз железа, устойчивость к противогрибковым азоловым препаратам и вирулентность. [20] Следовательно, потеря SrbA приводит к неспособности A. fumigatus расти в условиях с низким содержанием железа, более высокой чувствительности к противогрибковым азоловым препаратам и полной потере вирулентности на мышиных моделях IPA (инвазивный легочный аспергиллез). [21] Мутанты, нокаутные по SrbA, не проявляют каких-либо признаков роста in vitro в условиях низкого содержания кислорода, что, как полагают, связано с пониженной вирулентностью. Функциональность SrbA при гипоксии зависит от предшествующего процесса расщепления, осуществляемого белками RbdB, SppA и Dsc AE. [22] [23] [24] SrbA отщепляется от эндоплазматического ретикулума, содержащего белок-предшественник из 1015 аминокислот, до функциональной формы из 381 аминокислоты. Потеря любого из вышеуказанных процессинговых белков SrbA приводит к дисфункциональной копии SrbA и последующей потере роста in vitro при гипоксии, а также к ослаблению вирулентности. Исследования иммунопреципитации хроматина с белком SrbA привели к идентификации второго регулятора гипоксии - SrbB. [21] Хотя мало что известно о процессинге SrbB, было также показано, что этот фактор транскрипции играет ключевую роль в вирулентности и реакции грибковой гипоксии. [21] Подобно SrbA, нокаутный мутант SrbB приводил к потере вирулентности, однако не наблюдалось ни повышенной чувствительности к противогрибковым препаратам, ни полной потери роста в условиях гипоксии (снижение SrbB на 50% вместо снижения SrbA на 100%). [21] [20] Таким образом, было показано, что как SrbA, так и SrbB играют решающую роль в адаптации A. fumigatus к хозяину-млекопитающему.
Приобретение питательных веществ
[ редактировать ]Aspergillus fumigatus должен получать питательные вещества из внешней среды, чтобы выжить и процветать внутри своего хозяина. Эксперименты с генетическими мутациями показали, что многие гены, участвующие в таких процессах, влияют на вирулентность. Примеры поглощения питательных веществ включают металлы, азот и макромолекулы, такие как пептиды. [12] [25]
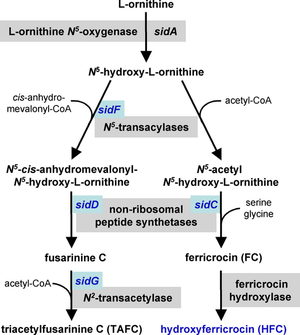
Приобретение железа
[ редактировать ]Железо является необходимым кофактором для многих ферментов и может действовать как катализатор в системе транспорта электронов. A. fumigatus имеет два механизма поглощения железа: восстановительное приобретение железа и сидерофором . опосредованное [27] [28] Приобретение восстановительного железа включает преобразование железа из трехвалентного железа (Fe +3 ) к черным (Fe +2 железа ) состояние и последующее поглощение через FtrA, пермеазу . Направленная мутация гена ftrA не вызывала снижения вирулентности на мышиной модели инвазии A. fumigatus . Напротив, целевая мутация sidA, первого гена в пути биосинтеза сидерофоров, доказала, что опосредованное сидерофорами поглощение железа имеет важное значение для вирулентности. [28] [29] Мутация нижестоящих генов биосинтеза сидерофоров sidC, sidD, sidF и sidG привела к появлению штаммов A. fumigatus с аналогичным снижением вирулентности. [26] Эти механизмы поглощения железа, по-видимому, работают параллельно, и оба активируются в ответ на железное голодание. [28]
Ассимиляция азота
[ редактировать ]Aspergillus fumigatus может выжить при использовании различных источников азота , и ассимиляция азота имеет клиническое значение, поскольку было показано, что он влияет на вирулентность. [25] [30] Белки, участвующие в ассимиляции азота, транскрипционно регулируются геном AfareA у A. fumigatus . Направленная мутация гена afareA показала снижение смертности на мышиной модели инвазии. [30] Белок RhbA, регулируемый Ras , также участвует в ассимиляции азота. Было обнаружено, что транскрипция RhbA активируется после контакта A. fumigatus с эндотелиальными клетками человека, а штаммы с целевой мутацией гена rhbA демонстрируют снижение роста на бедных источниках азота и снижение вирулентности in vivo . [31]
протеиназы
[ редактировать ]Легкие человека содержат большое количество коллагена и эластина — белков, которые обеспечивают гибкость тканей. [32] Aspergillus fumigatus производит и секретирует эластазы, протеазы , которые расщепляют эластин, чтобы расщепить эти макромолекулярные полимеры для поглощения. Существенная корреляция между количеством продукции эластазы и инвазией в ткани была впервые обнаружена в 1984 году. [33] Также было обнаружено, что клинические изоляты обладают большей эластазной активностью, чем штаммы A. fumigatus, находящиеся в окружающей среде . [34] Охарактеризован ряд эластаз, в том числе из семейств сериновых протеаз , аспарагиновых протеаз и металлопротеаз . [35] [36] [37] [38] Тем не менее, большая избыточность этих эластаз препятствует выявлению специфического воздействия на вирулентность. [12] [25]
Развернутый белковый ответ
[ редактировать ]Ряд исследований показал, что развернутая белковая реакция способствует вирулентности A. fumigatus . [39]
Вторичный метаболизм
[ редактировать ]Вторичные метаболиты в развитии грибов
[ редактировать ]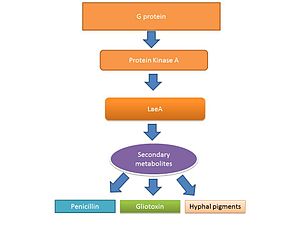
Жизненный цикл нитчатых грибов, включая Aspergillus spp. состоит из двух фаз: фазы роста гиф и репродуктивной фазы ( споруляции ). Переключение между фазами роста и размножения этих грибов частично регулируется уровнем продукции вторичных метаболитов. [41] [42] Считается, что вторичные метаболиты производятся для активации споруляции и пигментов, необходимых для структур споруляции. [43] Передача сигналов G-белка регулирует выработку вторичных метаболитов. [44] Секвенирование генома выявило 40 потенциальных генов, участвующих в производстве вторичных метаболитов, включая микотоксины, которые производятся во время споруляции. [9] [45]
Глиотоксин
[ редактировать ]Глиотоксин – это микотоксин, способный изменять защитные силы хозяина посредством иммуносупрессии. Нейтрофилы являются основной мишенью глиотоксина. [46] [47] Глиотоксин нарушает функцию лейкоцитов, ингибируя миграцию и выработку супероксида, и вызывает апоптоз макрофагов. [48] Глиотоксин нарушает провоспалительную реакцию посредством ингибирования NF-κB . [49]
Транскрипционная регуляция глиотоксина
[ редактировать ]LaeA и GliZ являются факторами транскрипции, которые, как известно, регулируют выработку глиотоксина. LaeA является универсальным регулятором продукции вторичных метаболитов у Aspergillus spp. [40] LaeA влияет на экспрессию 9,5% генома A. fumigatus , включая многие гены биосинтеза вторичных метаболитов, такие как нерибосомальные пептидсинтетазы . [50] Продукция многочисленных вторичных метаболитов, включая глиотоксин, была нарушена у мутантного штамма LaeA (ΔlaeA). [50] Мутант ΔlaeA показал повышенную восприимчивость к фагоцитозу макрофагов и снижение способности убивать нейтрофилы ex vivo . [47] Токсины, регулируемые LaeA, помимо глиотоксина, вероятно, играют роль в вирулентности, поскольку потеря продукции глиотоксина сама по себе не приводит к повторению гиповирулентного патотипа ∆laeA. [50]
Современные методы борьбы A. fumigatus с инфекциями
[ редактировать ]Современные неинвазивные методы лечения грибковых инфекций включают класс препаратов, известных как азолы. Азольные препараты, такие как вориконазол , итраконазол и имидазол, убивают грибы, ингибируя выработку эргостерина — важнейшего элемента мембран грибковых клеток. Механически эти препараты действуют путем ингибирования грибкового фермента цитохрома p450, известного как 14α-деметилаза . [51] Однако устойчивость A. fumigatus к азолам возрастает, возможно, из-за использования низких доз азолов в сельском хозяйстве. [52] [53] Основной способ резистентности – мутации гена cyp51a . [54] [55] Однако наблюдались другие формы резистентности, составляющие почти 40% резистентности клинических изолятов. [56] [57] [58] Наряду с азолами существуют и другие классы противогрибковых препаратов, такие как полиены и эхинокандины . [ нужна ссылка ]
Галерея
[ редактировать ]
 Конидии фиалоконидии A. fumigatus
Конидии фиалоконидии A. fumigatus Колония в чашке Петри
Колония в чашке Петри A. fumigatus, выделенный из лесной почвы.
A. fumigatus, выделенный из лесной почвы. Слайд мозга зараженной индейки
Слайд мозга зараженной индейки





См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Фанг В., Латге Дж.П. (август 2018 г.). «Микробный профиль: Aspergillus fumigatus: сапротрофный и условно-патогенный грибковый патоген» . Микробиология . 164 (8): 1009–1011. дои : 10.1099/mic.0.000651 . ПМК 6152418 . ПМИД 30066670 .
- ^ О'Горман СМ, Фуллер Х., Дайер П.С. (январь 2009 г.). «Открытие полового цикла у условно-патогенного грибка Aspergillus fumigatus». Природа . 457 (7228): 471–4. Бибкод : 2009Natur.457..471O . дои : 10.1038/nature07528 . ПМИД 19043401 . S2CID 4371721 .
- ^ Ридхольм К., Сакач Г., Лутзони Ф. (апрель 2006 г.). «Низкая генетическая изменчивость и отсутствие обнаруживаемой структуры популяции у aspergillus fumigatus по сравнению с близкородственными видами Neosartorya» . Эукариотическая клетка . 5 (4): 650–7. doi : 10.1128/EC.5.4.650-657.2006 . ПМК 1459663 . ПМИД 16607012 .
- ^ Дхингра С., Крамер Р.А. (2017). «Регуляция биосинтеза стерола у грибкового патогена человека Aspergillus fumigatus: возможности для терапевтического развития» . Границы микробиологии . 8 : 92. дои : 10.3389/fmicb.2017.00092 . ПМЦ 5285346 . ПМИД 28203225 .
- ^ Абад А., Фернандес-Молина Дж.В., Биканди Дж., Рамирес А., Маргарето Дж., Сендино Дж. и др. (декабрь 2010 г.). «Что делает Aspergillus fumigatus успешным патогеном? Гены и молекулы, участвующие в инвазивном аспергиллезе» (PDF) . Иберо-американский журнал микологии . 27 (4): 155–82. дои : 10.1016/j.riam.2010.10.003 . ПМИД 20974273 .
- ^ Цуй CB, Какея Х, Осада Х (август 1996 г.). «Спиротрипростатин B, новый ингибитор клеточного цикла млекопитающих, продуцируемый Aspergillus fumigatus» . Журнал антибиотиков . 49 (8): 832–5. дои : 10.7164/антибиотики.49.832 . ПМИД 8823522 .
- ^ Ниеминен С.М., Кярки Р., Ауриола С., Тойвола М., Лаатш Х., Лаатикайнен Р. и др. (октябрь 2002 г.). «Выделение и идентификация микотоксинов Aspergillus fumigatus на питательной среде и некоторых строительных материалах» . Прикладная и экологическая микробиология . 68 (10): 4871–5. Бибкод : 2002ApEnM..68.4871N . doi : 10.1128/aem.68.10.4871-4875.2002 . ПМК 126391 . ПМИД 12324333 .
- ^ Галаган Дж.Э., Кальво С.Е., Куомо С., Ма Л.Дж., Вортман Дж.Р., Бацоглу С. и др. (декабрь 2005 г.). «Секвенирование Aspergillus nidulans и сравнительный анализ с A. fumigatus и A. oryzae» . Природа . 438 (7071): 1105–15. Бибкод : 2005Natur.438.1105G . дои : 10.1038/nature04341 . ПМИД 16372000 .
- ^ Перейти обратно: а б Нирман В.К., Пейн А., Андерсон М.Дж., Вортман Дж.Р., Ким Х.С., Арройо Дж. и др. (декабрь 2005 г.). «Геномная последовательность патогенного и аллергенного нитчатого гриба Aspergillus fumigatus» . Природа . 438 (7071): 1151–6. Бибкод : 2005Natur.438.1151N . дои : 10.1038/nature04332 . hdl : 10261/71531 . ПМИД 16372009 .
- ^ Мачида М., Асаи К., Сано М., Танака Т., Кумагай Т., Тераи Г. и др. (декабрь 2005 г.). «Секвенирование генома и анализ Aspergillus oryzae» . Природа . 438 (7071): 1157–61. Бибкод : 2005Natur.438.1157M . дои : 10.1038/nature04300 . ПМИД 16372010 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м Бен-Ами Р., Льюис Р.Э., Контойяннис Д.П. (август 2010 г.). «Враг (иммуносупрессированного) состояния: обновленная информация о патогенезе инфекции Aspergillus fumigatus» . Британский журнал гематологии . 150 (4): 406–17. дои : 10.1111/j.1365-2141.2010.08283.x . ПМИД 20618330 . S2CID 28216163 .
- ^ Перейти обратно: а б с д Холь Т.М., Фельдмессер М. (ноябрь 2007 г.). «Aspergillus fumigatus: принципы патогенеза и защиты хозяина» . Эукариотическая клетка . 6 (11): 1953–63. дои : 10.1128/EC.00274-07 . ПМК 2168400 . ПМИД 17890370 .
- ^ Перейти обратно: а б с д и ж г час я Сигал Б.Х. (апрель 2009 г.). «Аспергиллез». Медицинский журнал Новой Англии . 360 (18): 1870–84. дои : 10.1056/NEJMra0808853 . ПМИД 19403905 .
- ^ Перейти обратно: а б с д и ж Филлер С.Г., Шеппард, округ Колумбия (декабрь 2006 г.). «Грибковая инвазия в обычно нефагоцитарные клетки-хозяева» . ПЛОС Патогены . 2 (12): е129. дои : 10.1371/journal.ppat.0020129 . ПМК 1757199 . ПМИД 17196036 .
- ^ Карнам А., Бонам С.Р., Рамбабу Н., Вонг С.С., Айманианда В., Байри Дж. (16 ноября 2021 г.). «Передача сигналов Wnt-β-катенина в дендритных клетках человека опосредует регуляторные ответы Т-клеток на грибы через путь PD-L1» . мБио . 12 (6): e0282421. дои : 10.1128/mBio.02824-21 . ПМЦ 8593687 . ПМИД 34781737 .
- ^ Стивен-Виктор Э., Карнам А., Фонтен Т., Бове А., Дас М., Хегде П., Прахар П., Холла С., Баладжи К.Н., Кавери С.В., Латге Ж.П., Айманианда В., Байри Дж. (5 декабря 2017 г.). «А-(1,3)-глюкан клеточной стенки Aspergillus fumigatus стимулирует регуляторную поляризацию Т-клеток, индуцируя экспрессию PD-L1 на дендритных клетках человека» . J Заразить Дис . 216 (10): 1281–1294. дои : 10.1093/infdis/jix469 . ПМИД 28968869 .
- ^ Грал Н., Крамер Р.А. (февраль 2010 г.). «Регуляция адаптации к гипоксии: упущенный из виду атрибут вирулентности патогенных грибов?» . Медицинская микология . 48 (1): 1–15. дои : 10.3109/13693780902947342 . ПМЦ 2898717 . ПМИД 19462332 .
- ^ Перейти обратно: а б Грал Н., Шепардсон К.М., Чанг Д., Крамер Р.А. (май 2012 г.). «Гипоксия и грибковый патогенез: проветривать или не проветривать?» . Эукариотическая клетка . 11 (5): 560–70. дои : 10.1128/EC.00031-12 . ПМЦ 3346435 . ПМИД 22447924 .
- ^ Везенский С.Дж., Крамер Р.А. (апрель 2011 г.). «Последствия гипоксической микросреды при инвазивном аспергиллезе» . Медицинская микология . 49 (Приложение 1): S120–4. дои : 10.3109/13693786.2010.495139 . ПМЦ 2951492 . ПМИД 20560863 .
- ^ Перейти обратно: а б Виллгер С.Д., Путтикамонкул С., Ким К.Х., Берритт Дж.Б., Грал Н., Мецлер Л.Дж., Барбух Р., Бард М., Лоуренс CB, Крамер Р.А. (ноябрь 2008 г.). «Белок, связывающий стерол-регуляторный элемент, необходим для полярности клеток, адаптации к гипоксии, устойчивости к азолам и вирулентности у Aspergillus fumigatus» . ПЛОС Патогены . 4 (11): е1000200. дои : 10.1371/journal.ppat.1000200 . ПМК 2572145 . ПМИД 18989462 .
- ^ Перейти обратно: а б с д Чанг Д., Баркер Б.М., Кэри CC, Мерриман Б., Вернер Э.Р., Лехнер Б.Е., Дхингра С., Ченг С., Сюй В., Блоссер С.Дж., Морохаши К., Мазури А., Митчелл Т.К., Хаас Х., Митчелл А.П., Крамер Р.А. (ноябрь 2014 г.) ). «Анализ ChIP-seq и транскриптома in vivo Aspergillus fumigatus SREBP SrbA выявил новый регулятор реакции грибков на гипоксию и вирулентности» . ПЛОС Патогены . 10 (11): e1004487. дои : 10.1371/journal.ppat.1004487 . ПМК 4223079 . ПМИД 25375670 .
- ^ Дхингра С., Ковальски CH, Таммахонг А., Битти С.Р., Балтман К.М., Крамер Р.А. (2016). «RbdB, ромбовидная протеаза, имеющая решающее значение для активации и вирулентности SREBP в Aspergillus fumigatus» . мСфера . 1 (2). дои : 10.1128/mSphere.00035-16 . ПМЦ 4863583 . ПМИД 27303716 .
- ^ Бат-Очир С., Квак Дж.Ю., Ко С.К., Чон М.Х., Чунг Д., Ли Ю.В., Че С.К. (май 2016 г.). «Сигнальная пептидаза пептидаза SppA участвует в расщеплении белка, связывающего регуляторные элементы стерола, и адаптации к гипоксии у Aspergillus nidulans» . Молекулярная микробиология . 100 (4): 635–55. дои : 10.1111/mmi.13341 . ПМИД 26822492 .
- ^ Виллгер С.Д., Корниш Э.Дж., Чанг Д., Флеминг Б.А., Леманн М.М., Путтикамонкул С., Крамер Р.А. (декабрь 2012 г.). «Ортологи Dsc необходимы для адаптации к гипоксии, реакции на препараты триазола и вирулентности грибов у Aspergillus fumigatus» . Эукариотическая клетка . 11 (12): 1557–67. дои : 10.1128/EC.00252-12 . ПМЦ 3536281 . ПМИД 23104569 .
- ^ Перейти обратно: а б с Дагенайс Т.Р., Келлер Н.П. (июль 2009 г.). «Патогенез Aspergillus fumigatus при инвазивном аспергиллезе» . Обзоры клинической микробиологии . 22 (3): 447–65. дои : 10.1128/CMR.00055-08 . ПМК 2708386 . ПМИД 19597008 .
- ^ Перейти обратно: а б Шреттл М., Бигнелл Э., Крагл С., Сабиха Ю., Лосс О., Эйзендл М. и др. (сентябрь 2007 г.). «Различные роли внутри- и внеклеточных сидерофоров при инфекции Aspergillus fumigatus» . ПЛОС Патогены . 3 (9): 1195–207. дои : 10.1371/journal.ppat.0030128 . ЧВК 1971116 . ПМИД 17845073 .
- ^ Хаас Х (сентябрь 2003 г.). «Молекулярная генетика биосинтеза и поглощения сидерофоров грибов: роль сидерофоров в поглощении и хранении железа». Прикладная микробиология и биотехнология . 62 (4): 316–30. дои : 10.1007/s00253-003-1335-2 . ПМИД 12759789 . S2CID 10989925 .
- ^ Перейти обратно: а б с Шреттл М., Бигнелл Э., Крагл С., Джохл С., Роджерс Т., Арст Х.Н. и др. (ноябрь 2004 г.). «Для вирулентности Aspergillus fumigatus важен биосинтез сидерофоров, а не восстановительная ассимиляция железа» . Журнал экспериментальной медицины . 200 (9): 1213–9. дои : 10.1084/jem.20041242 . ПМК 2211866 . ПМИД 15504822 .
- ^ Хиссен А.Х., Ван А.Н., Варвас М.Л., Пинто Л.Дж., Мур М.М. (сентябрь 2005 г.). «Для вирулентности необходим ген биосинтеза сидерофора Aspergillus fumigatus sidA, кодирующий L-орнитин-N5-оксигеназу» . Инфекция и иммунитет . 73 (9): 5493–503. дои : 10.1128/IAI.73.9.5493-5503.2005 . ПМЦ 1231119 . ПМИД 16113265 .
- ^ Перейти обратно: а б Хенсель М., Арст Х.Н., Офовр-Браун А., Холден Д.В. (июнь 1998 г.). «Роль гена Aspergillus fumigatus areA в инвазивном легочном аспергиллезе». Молекулярная и общая генетика . 258 (5): 553–7. дои : 10.1007/s004380050767 . ПМИД 9669338 . S2CID 27753283 .
- ^ Панепинто Дж.К., Оливер Б.Г., Амлунг Т.В., Аскью Д.С., Роудс Дж.К. (август 2002 г.). «Экспрессия гомолога реба Aspergillus fumigatus, rhbA, индуцируется азотным голоданием». Грибковая генетика и биология . 36 (3): 207–14. дои : 10.1016/S1087-1845(02)00022-1 . ПМИД 12135576 .
- ^ Розенблум Дж. (декабрь 1984 г.). «Эластин: связь структуры белка и генов с болезнями». Лабораторные исследования; Журнал технических методов и патологии . 51 (6): 605–23. ПМИД 6150137 .
- ^ Котари М.Х., Чейз Т., Макмиллан Дж.Д. (январь 1984 г.). «Корреляция продукции эластазы некоторыми штаммами Aspergillus fumigatus со способностью вызывать инвазивный аспергиллез легких у мышей» . Инфекция и иммунитет . 43 (1): 320–5. дои : 10.1128/IAI.43.1.320-325.1984 . ПМК 263429 . ПМИД 6360904 .
- ^ Бланко Х.Л., Хонтесильяс Р., Буза Э., Бланко И., Пелаес Т., Муньос П. и др. (май 2002 г.). «Корреляция между индексом активности эластазы и инвазивностью клинических изолятов Aspergillus fumigatus» . Журнал клинической микробиологии . 40 (5): 1811–3. doi : 10.1128/JCM.40.5.1811-1813.2002 . ПМЦ 130931 . ПМИД 11980964 .
- ^ Райхард У, Бюттнер С, Эйфферт Х, Стаиб Ф, Рюхель Р (декабрь 1990 г.). «Очистка и характеристика внеклеточной сериновой протеиназы Aspergillus fumigatus и ее обнаружение в тканях» . Журнал медицинской микробиологии . 33 (4): 243–51. дои : 10.1099/00222615-33-4-243 . ПМИД 2258912 .
- ^ Маркарян А., Морозова И., Ю.Х., Колаттукуди Ч.Е. (июнь 1994 г.). «Очистка и характеристика эластинолитической металлопротеазы из Aspergillus fumigatus и иммуноэлектронно-микроскопические доказательства секреции этого фермента грибком, проникающим в легкие мыши» . Инфекция и иммунитет . 62 (6): 2149–57. дои : 10.1128/IAI.62.6.2149-2157.1994 . ЧВК 186491 . ПМИД 8188335 .
- ^ Ли Дж.Д., Колаттукуди П.Е. (октябрь 1995 г.). «Молекулярное клонирование кДНК и гена эластинолитической аспарагиновой протеиназы из Aspergillus fumigatus и доказательства ее секреции грибом во время инвазии в легкие хозяина» . Инфекция и иммунитет . 63 (10): 3796–803. дои : 10.1128/IAI.63.10.3796-3803.1995 . ПМЦ 173533 . ПМИД 7558282 .
- ^ Ядарола П., Лунгарелла Г., Марторана П.А., Вильо С., Гульельминетти М., Корзус Е. и др. (1998). «Повреждение легких и деградация компонентов внеклеточного матрикса сериновой протеиназой Aspergillus fumigatus». Экспериментальное исследование легких . 24 (3): 233–51. дои : 10.3109/01902149809041532 . ПМИД 9635248 .
- ^ Фэн Икс, Кришнан К., Ричи Д.Л., Айманианда В., Хартл Л., Грал Н. и др. (октябрь 2011 г.). «Независимые от HacA функции датчика стресса ER IreA взаимодействуют с каноническим UPR, влияя на признаки вирулентности Aspergillus fumigatus» . ПЛОС Патогены . 7 (10): e1002330. дои : 10.1371/journal.ppat.1002330 . ПМК 3197630 . ПМИД 22028661 .
- ^ Перейти обратно: а б Бок Дж.В., Келлер Н.П. (апрель 2004 г.). «LaeA, регулятор вторичного метаболизма Aspergillus spp» . Эукариотическая клетка . 3 (2): 527–35. doi : 10.1128/EC.3.2.527-535.2004 . ПМЦ 387652 . ПМИД 15075281 .
- ^ Кальво А.М., Уилсон Р.А., Бок Дж.В., Келлер Н.П. (сентябрь 2002 г.). «Связь между вторичным метаболизмом и развитием грибков» . Обзоры микробиологии и молекулярной биологии . 66 (3): 447–59, оглавление. дои : 10.1128/MMBR.66.3.447-459.2002 . ПМК 120793 . ПМИД 12208999 .
- ^ Тао Л., Ю Дж. Х. (февраль 2011 г.). «AbaA и WetA управляют различными стадиями развития Aspergillus fumigatus» . Микробиология . 157 (Часть 2): 313–26. дои : 10.1099/mic.0.044271-0 . ПМИД 20966095 .
- ^ Адамс Т.Х., Ю Дж.Х. (декабрь 1998 г.). «Координационный контроль производства вторичных метаболитов и бесполого спорообразования у Aspergillus nidulans». Современное мнение в микробиологии . 1 (6): 674–7. дои : 10.1016/S1369-5274(98)80114-8 . ПМИД 10066549 .
- ^ Бродхаген М., Келлер Н.П. (июль 2006 г.). «Сигнальные пути, связывающие продукцию микотоксинов и споруляцию» . Молекулярная патология растений . 7 (4): 285–301. дои : 10.1111/j.1364-3703.2006.00338.x . ПМИД 20507448 .
- ^ Трейл F, Маханти Н., Линц Дж. (апрель 1995 г.). «Молекулярная биология биосинтеза афлатоксина» . Микробиология . 141 (4): 755–65. дои : 10.1099/13500872-141-4-755 . ПМИД 7773383 .
- ^ Спайкс С., Сюй Р., Нгуен К.К., Чамилос Г., Контойяннис Д.П., Джейкобсон Р.Х. и др. (февраль 2008 г.). «Продуцирование глиотоксина Aspergillus fumigatus способствует специфичным различиям в вирулентности в зависимости от хозяина» . Журнал инфекционных болезней . 197 (3): 479–86. дои : 10.1086/525044 . ПМИД 18199036 .
- ^ Перейти обратно: а б Бок Дж.В., Чунг Д., Баладжи С.А., Марр К.А., Андес Д., Нильсен К.Ф. и др. (декабрь 2006 г.). «GliZ, транскрипционный регулятор биосинтеза глиотоксина, способствует вирулентности Aspergillus fumigatus» . Инфекция и иммунитет . 74 (12): 6761–8. дои : 10.1128/IAI.00780-06 . ПМК 1698057 . ПМИД 17030582 .
- ^ Камей К., Ватанабэ А. (май 2005 г.). «Микотоксины аспергилл и их влияние на хозяина» . Медицинская микология . 43 (Приложение 1): С95-9. дои : 10.1080/13693780500051547 . ПМИД 16110799 .
- ^ Гардинер Д.М., Уоринг П., Хоулетт Б.Дж. (апрель 2005 г.). «Класс грибковых токсинов эпиполитиодиоксопиперазина (ЭТП): распределение, механизм действия, функции и биосинтез» . Микробиология . 151 (Часть 4): 1021–1032. дои : 10.1099/mic.0.27847-0 . ПМИД 15817772 .
- ^ Перейти обратно: а б с Перрин Р.М., Федорова Н.Д., Бок Дж.В., Крамер Р.А., Вортман Дж.Р., Ким Х.С. и др. (апрель 2007 г.). «Транкрипционная регуляция химического разнообразия Aspergillus fumigatus с помощью LaeA» . ПЛОС Патогены . 3 (4): е50. doi : 10.1371/journal.ppat.0030050 . ПМК 1851976 . ПМИД 17432932 .
- ^ Панакал А.А., Беннетт Дж.Э., Уильямсон П.Р. (сентябрь 2014 г.). «Варианты лечения инвазивного аспергиллеза» . Современные варианты лечения инфекционных заболеваний . 6 (3): 309–325. дои : 10.1007/s40506-014-0016-2 . ПМК 4200583 . ПМИД 25328449 .
- ^ Бергер С., Эль Чазли Ю., Бабу А.Ф., Косте А.Т. (07.06.2017). «Aspergillus fumigatus: последствия использования противогрибковых средств в сельском хозяйстве?» . Границы микробиологии . 8 : 1024. дои : 10.3389/fmicb.2017.01024 . ПМЦ 5461301 . ПМИД 28638374 .
- ^ Буейд А., Ховард С.Дж., Мур CB, Ричардсон М.Д., Харрисон Э., Бойер П., Деннинг Д.В. (октябрь 2010 г.). «Противогрибковая резистентность к азолу у Aspergillus fumigatus: 2008 и 2009 годы» . Журнал антимикробной химиотерапии . 65 (10): 2116–8. дои : 10.1093/jac/dkq279 . ПМИД 20729241 .
- ^ Нэш А., Роудс Дж. (апрель 2018 г.). «Моделирование CYP51A Aspergillus fumigatus в модельном бислое дает представление о лекарственной устойчивости к триазолам» . Медицинская микология . 56 (3): 361–373. дои : 10.1093/mmy/myx056 . ПМК 5895076 . ПМИД 28992260 .
- ^ Снелдерс Э., Каравайчик А., Шафтенаар Г., Вервей П.Е., Мельчерс В.Дж. (июнь 2010 г.). «Профиль устойчивости к азолу аминокислотных изменений в CYP51A Aspergillus fumigatus на основе моделирования гомологии белков» . Антимикробные средства и химиотерапия . 54 (6): 2425–30. дои : 10.1128/AAC.01599-09 . ПМЦ 2876375 . ПМИД 20385860 .
- ^ Рыбак Дж.М., Ге В., Видерхольд Н.П., Паркер Дж.Э., Келли С.Л., Роджерс П.Д., Фортвендел Дж.Р. (апрель 2019 г.). Олспо Дж.А. (ред.). «hmg1, бросая вызов парадигме клинической резистентности к триазолам у Aspergillus fumigatus» . мБио . 10 (2): e00437–19, /mbio/10/2/mBio.00437–19.atom. дои : 10.1128/mBio.00437-19 . ПМК 6445940 . ПМИД 30940706 .
- ^ Кэмпс С.М., Дутиль Б.Е., Арендруп М.К., Рейс А.Дж., Снелдерс Э., Хуйнен М.А. и др. (30 ноября 2012 г.). «Обнаружение мутации HapE, вызывающей устойчивость к азолам у Aspergillus fumigatus, посредством полногеномного секвенирования и полового скрещивания» . ПЛОС ОДИН . 7 (11): e50034. Бибкод : 2012PLoSO...750034C . дои : 10.1371/journal.pone.0050034 . ПМЦ 3511431 . ПМИД 23226235 .
- ^ Фурукава Т., ван Рейн Н., Фрачек М., Гсаллер Ф., Дэвис Э., Карр П. и др. (январь 2020 г.). «Комплекс негативного кофактора 2 является ключевым регулятором лекарственной устойчивости Aspergillus fumigatus» . Природные коммуникации . 11 (1): 427. Бибкод : 2020NatCo..11..427F . дои : 10.1038/s41467-019-14191-1 . ПМК 7194077 . ПМИД 31969561 .
Внешние ссылки
[ редактировать ]- Возникновение устойчивости к азолу у Aspergillus fumigatus и распространение единого механизма устойчивости. в SciVee
- The Aspergillus Trust Зарегистрированная в Великобритании благотворительная организация, занимающаяся поддержкой людей с аспергиллезом во всем мире и исследованием методов лечения.
- Фонд исследований грибков
- об аспергиллах Информация с сайта DoctorFungus.org.
| Базы данных органов управления : Национальные |
|---|