Кокколитовирус
| Кокколитовирус | |
|---|---|

| |
| Гигантский кокколитовирус, вирус Emiliania huxleyi 86 (стрелка), заражающий Emiliania huxleyi. кокколитофор | |
| Классификация вирусов | |
| (без рейтинга): | Вирус |
| Область : | Вариднавирия |
| Королевство: | Бэмфордвире |
| Тип: | Нуклеоцитовирикота |
| Сорт: | Мегавирицеты |
| Заказ: | Альгавиралес |
| Семья: | Фикоднавирусиды |
| Род: | Кокколитовирус |
| Штаммы | |
| |
Кокколитовирус — род гигантских двухцепочечных ДНК-вирусов семейства Phycodnaviridae . Водоросли, в частности Emiliania huxleyi , вид кокколитофоров . [1] служат естественными хозяевами. В этом роде описан только один вид: вирус Emiliania huxleyi 86 . [ 1 ] [ 2 ]
Структура
[ редактировать ]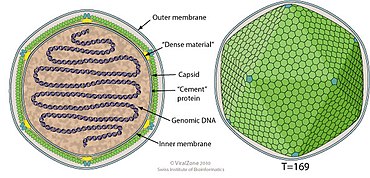
Кокколитовирусы имеют оболочку, икосаэдрическую форму и диаметр от 100 до 220 нм. Их геномы линейны, имеют длину от 410 до 415 КБ и, по прогнозам, кодируют примерно 472 белка. [ 1 ]
| Род | Структура | Симметрия | Капсид | Геномное расположение | Геномная сегментация |
|---|---|---|---|---|---|
| Кокколитовирус | икосаэдрический | Т=169 | обернутый | Линейный | Однодольный |
Жизненный цикл
[ редактировать ]Кокколитовирусы являются частью семейства Phycodnaviridae , одного из пяти семейств, принадлежащих к большой и филогенетически разнообразной группе вирусов, известных как нуклеоцитоплазматические большие дцДНК-вирусы ( NCLDV ). Эти вирусы либо реплицируются исключительно в цитоплазме клетки-хозяина, либо начинают свой жизненный цикл в ядре хозяина , но завершают его в цитоплазме. В случае EhV-86 стратегия заражения не до конца понятна, но Mackinder et al. (2009) [ 3 ] предложили следующую модель: вирус проникает в клетку-хозяина посредством эндоцитоза с последующим слиянием его липидной мембраны с мембраной вакуоли хозяина и высвобождением его нуклеопротеинового ядра в цитоплазму. Альтернативно мембрана вируса может сливаться непосредственно с плазматической мембраной хозяина. вируса Затем геном высвобождается из капсида в ядро, где он реплицируется вирусной ДНК-полимеразой . Реплицированный геном упаковывается в собранные капсиды в цитоплазме, и новообразованные (до 400–1000) вирионы, как полагают, транспортируются к плазматической мембране и высвобождаются посредством контролируемого механизма почкования, что приводит к клеточному распаду клетки-хозяина. . [ нужна ссылка ]
На стадиях жизненного цикла G2 и M коккосфера является неполной, и воздействие вируса на плазматическую мембрану увеличивается. Даже при интактной коккосфере заражение может произойти из-за естественных промежутков между кокколитами. [ 4 ]
E. huxleyi известен тем, что формирует сезонное цветение водорослей, площадь которого может достигать 250 000 км2. 2 , в течение которого плотность клеток в верхних 200 м увеличивается с 10 3 до 10 5 клеток на мл морской воды. [ 5 ] Цветение водорослей обычно прекращается через 5–8 дней, и несколько исследований показали, что прекращение цветения неразрывно связано с заражением кокколитовирусами. [ 6 ] Передача вирусов между водорослями-хозяевами происходит путем пассивной диффузии. Кроме того, ДНК EhV была также обнаружена у копепод , что привело к предположению, что вирусы далее распространяются через зоопланктон , несущий вирус . [ 7 ]
Геном
[ редактировать ]На сегодняшний день в период с 1999 по 2008 год было выделено 14 штаммов EhV, главным образом из Ла-Манша (EC), а также с норвежского и шотландского побережья. [ 8 ] [ 9 ] [ 10 ] [ 11 ] [ 12 ] Хотя доступны частичные последовательности всех этих 14 штаммов, EhV-86 является единственным штаммом, который был полностью секвенирован из-за высокой повторяемости генома. [ 13 ] Секвенирование EhV-86 выявило кольцевой геном длиной 407 339 п.н. с 472 предсказанными кодирующими последовательностями ( CDS ). Примечательно, что 80% этих предполагаемых генов на сегодняшний день не имеют гомологов в базе данных. Те, которым можно приписать функцию из-за сходства последовательностей или совпадений белковых доменов, включают субъединицы ДНК и РНК-полимеразы , восемь протеаз, а также по крайней мере четыре гена, которые кодируют белки, участвующие в биосинтезе сфинголипидов. Было показано, что они были получены от хозяина посредством горизонтального переноса генов . [ 14 ]
Более того, в геноме EhV-86 обнаружено три различных семейства (A, B, C) повторяющихся областей внутри генома. [ 15 ] Семейство C состоит из некодирующих повторов, богатых АТ, которые, вероятно, являются частью начала репликации ( ORF ). Семейство B представляет собой богатые GC повторы, которые обнаружены в белковых продуктах восьми предсказанных CDS. Гомологические области семейства A различаются по размеру от 30 до 300 п.н. и обнаруживаются в участке генома размером 104 кб (200–304 т.п.н.), который не содержит гомологов генов с известной функцией в текущих базах данных. Повторяющиеся единицы семейства А некодируют и характеризуются наномером (GTTCCC(T/C)AA), который в общей сложности появляется в 106 местах в этой области. Эта последовательность находится непосредственно перед 86 CDS и, вероятно, играет роль в контроле экспрессии связанных CDS. [ нужна ссылка ]
Во время инфекции был описан отчетливый характер экспрессии вирусных генов, который можно разделить на три фазы в зависимости от экспрессии CDS. [ 16 ] Через час после заражения было транскрибировано 39 вирусных генов, затем еще 194 гена через 2 часа и 71 ген через 4 часа. Все 39 генов, которые экспрессируются через 1 час после инфицирования, расположены в области размером 104 т.п.н. и имеют наномер непосредственно перед инициирующим кодоном. Поскольку экспрессия вирусной РНК-полимеразы не была обнаружена через 1 ч после заражения, еще предстоит установить, распознается ли промотор упакованной вирусной РНК-полимеразой или РНК-полимеразой хозяина. Однако протеомный анализ вириона EhV-86 не выявил каких-либо основных субъединиц РНК-полимеразы. [ 17 ]
История
[ редактировать ]Уильям Уилсон и его команда из Ассоциации морской биологии (MBA), Университета Восточной Англии и Плимутской морской лаборатории (PML) впервые наблюдали вирус в 1999 году. Позже, летом 2005 года, исследователи из Плимутской морской лаборатории (Уилли Уилсон и др.) .) и в Институте Сэнгера (Холден и др.) секвенировали геном штамма EhV-86, обнаружив, что он содержит 472 гена, кодирующих белок, что делает его « гигантским вирусом». ", и самый крупный из известных морских вирусов по геному [2] .
В ходе первоначального исследования генома кокколитовирусов последовательность генов, ответственных за выработку церамидов . была обнаружена [ 18 ] Керамид является контролирующим фактором гибели клеток, и в настоящее время считается, что кокколитовирус использует его для продления жизни Emiliania huxleyi , одновременно используя клетку-хозяина для репликации. Это уникальная способность, невиданная на сегодняшний день ни в одном другом вирусном геноме. [ нужна ссылка ]
См. также
[ редактировать ]- Мимивирус - крупнейший зарегистрированный гигантский вирус по геному.
- Mycoplasmagentium , Pelagibacter ubique – одни из самых маленьких известных бактерий.
- Наноархей - самый маленький известный архей
- Наименьшие организмы
- Парвовирус - наименьшее известное семейство вирусов.
- Phycodnaviridae - вирусы, поражающие водоросли.
Примечания
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б «Вирусная зона» . ЭксПАСи . Проверено 15 июня 2015 г.
- ^ «Таксономия вирусов: выпуск 2020 г.» . Международный комитет по таксономии вирусов (ICTV). Март 2021 года . Проверено 22 мая 2021 г.
- ^ Маккиндер LCM, Уорти Калифорния, Бигги Дж., Холл М, Райан КП, Варсани А, Харпер GM, Уилсон WH, Браунли С, Шредер, округ Колумбия. (2009) Одноклеточный вирус водорослей, Emilienia huxleyivirus 86, использует стратегию заражения, подобную животной. Журнал общей вирусологии 90:2306–2316.
- ^ Пааше Э. (2001) Обзор кокколитофорида Emiliania huxleyi (Prymnesiophyceae) с особым упором на рост, образование кокколитов и взаимодействие кальцификации и фотосинтеза. Психология 40 (6): 503–52.
- ^ Шредер, округ Колумбия, Оке Дж., Холл М., Малин Дж., Уилсон WH. (2003)Вирусная последовательность наблюдалась во время цветения Emilinaia huxleyi. Прикладная и экологическая микробиология 69:2484–2490.
- ^ Уилсон В.Х., Тарран Г.А., Шредер Д., Кокс М., Оук Дж., Малин Г. (2002) Выделение вирусов, ответственных за гибель цветения Emiliania huxleyi в Ла-Манше. Журнал Морской биологической ассоциации Соединенного Королевства 82: 369–377.
- ^ Фрада М.Дж., Шац Д., Фарстей В., Оссолински Дж.Э., Сабанай Х., Бен-Дор С., Корен И., Варди А. (2014) Зоопланктон может служить векторами передачи вирусов, заражающих цветение водорослей в океане. Текущая биология 24: 2592–2597.
- ^ Нисимов Дж.И., Напирб Дж.А., Кимманс С.А., Аллен М.Дж. (2014)Постоянные проекты геномов четырех новых кокколитовирусов: EhV-18, EhV-145, EhV-156 и EhV-164. Морская геномика 15:7–8.
- ^ Нисимов Дж.И., Уорти К.А., Рукс П., Нэпьер Дж.А., Кимманс С.А., Хенн М.Р., Огата Х., Аллен М.Дж. (2011)Проект последовательности генома кокколитовируса EhV-84. Стандарты геномной науки 5:1–11.
- ^ Нисимов Дж.И., Уорти К.А., Рукс П., Нэпьер Дж.А., Кимманс С.А., Хенн М.Р., Огата Х., Аллен М.Дж. (2012)Проект геномной последовательности четырех кокколитопвирусов: вирус Emiliania huxleyi EhV-88, EhV-201, EhV-207 и EhV-208. Журнал вирусологии 86(5):2896–2897.
- ^ Пагарете и др. 2012, Аллен и др. 2006d, Нисимов Дж.И., Уорти К.А., Рукс П., Нэпьер Дж.А., Кимманс С.А., Хенн М.Р., Огата Х., Аллен М.Дж. (2012) Проект последовательности генома вируса кокколитопвируса Эмилиании Хаксли 202. Журнал вирусологии 86 (4): 380–2381.
- ^ Нисимов Дж.И., Уорти К.А., Рукс П., Нэпьер Дж.А., Кимманс С.А., Хенн М.Р., Огата Х., Аллен М.Дж. (2011) Проект геномной последовательности кокколитовируса Emiliania Huxley Virus 203. Journal of Virology 85(24):13468–13469.
- ^ Уилсон WH, Шредер, округ Колумбия, Аллен М.Дж., Холден МТГ, Паркхилл Дж., Баррелл Б.Г., Черчер С., Хэмлин Н., Мангалл К., Норбертчак Х., Куэйл М.А., Прайс С., Раббинович Э., Уокер Д., Крейгон М., Рой Д., Газаль П. (2005)Полная последовательность генома и профиль транскрипции литической фазы кокколитовируса. Наука 309: 1090–1092.
- ^ Монье А., Пагарете А., Де Варгас С., Аллен М.Дж., Рид Б., Клавери Дж., Огата Х., Де Варгас С. (2009) Горизонтальный перенос генов всего метаболического пути между эукариотической водорослью и ее ДНК-вирусом. Исследования генома 19: 1441–1449.
- ^ Аллен М.Дж., Шредер, округ Колумбия, Уилсон WH. (2006)Предварительная характеристика семейств повторов в геноме EhV-86, вируса гигантских водорослей, который заражает морские микроводоросли Emiliania huxleyi. Архивы вирусологии 151: 525–535.
- ^ Аллен М.Дж., Форстер Т., Шредер, округ Колумбия, Холл М., Рой Д., Газаль П., Уилсон WH. (2006) Характер экспрессии локус-специфических генов предполагает уникальную стратегию размножения вируса гигантских водорослей. Журнал вирусологии 80: 7699–7705.
- ^ Аллен М.Дж., Ховард Дж.А., Лилли К.С., Уилсон WH. (2008)Протеомный анализ вириона EhV-86. Протеомная наука 6 (11).
- ^ Уилсон WH, Шредер, округ Колумбия, Аллен М.Дж., Холден МТГ, Паркхилл Дж., Баррелл Б.Г., Черчер С., Хэмлин Н., Мангалл К., Норбертчак Х., Куэйл М.А., Прайс С., Раббинович Э., Уокер Д., Крейгон М., Рой Д., Газаль П. (2005)Полная последовательность генома и профиль транскрипции литической фазы кокколитовируса. Наука 309: 1090–1092.
Дальнейшее чтение
[ редактировать ]- Уилсон, У.Х., Шредер, О.К., Аллен, М.Дж., Холден, М.Т.Г., Паркхилл, Дж., Баррелл, Б.Г., Черчер, К., Хэмлин, Н., Мангалл, К., Норбертчак, Х., Куэйл, Массачусетс, Прайс К., Раббинович Э., Уокер Д., Крейгон М., Рой Д. и Газаль П. (2005) Полный геном Профиль транскрипции последовательности и литической фазы кокколитовируса . Наука 5737 , 1090–1092
- Аллен М.Дж., Шредер Д.К., Холден М.Т. и Уилсон У.Х. (2006) Эволюционная история кокколитовирусов . Мол Биол Эвол. 23 , 86–92 ( требуется вход в Афины )