РНК-вирус с отрицательной цепью
| Негарнавирикота | |
|---|---|
| |
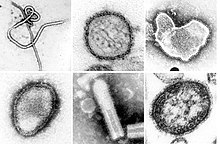
| Монтаж трансмиссионных электронных микрофотографий некоторых вирусов типа Negarnaviricota . Не в масштабе. Виды слева направо, сверху вниз: Заирский эболавирус , безымянный ортохантавирус , ортопневмовирус человека , генипавирус Хендра , неопознанный рабдовирус , морбилливирус кори . | |
| Классификация вирусов | |
| (без рейтинга): | Вирус |
| Область : | Рибовирия |
| Королевство: | Орторнавиры |
| Тип: | Негарнавирикота |
| Доплата | |
| Синонимы [ 1 ] [ 2 ] | |
| |
РНК-вирусы с отрицательной цепью ( -оцРНК-вирусы ) представляют собой группу родственных вирусов , которые имеют с отрицательным смыслом одноцепочечные геномы , состоящие из рибонуклеиновой кислоты (РНК). У них есть геномы, которые действуют как комплементарные цепи, из которых информационная РНК (мРНК) синтезируется вирусным ферментом РНК-зависимой РНК-полимеразой (RdRp). Во время репликации вирусного генома RdRp синтезирует антигеном с положительным смыслом, который использует в качестве матрицы для создания геномной РНК с отрицательным смыслом. Вирусы с отрицательной РНК также имеют ряд других характеристик: большинство из них содержат вирусную оболочку , окружающую капсид, в котором заключен вирусный геном, геномы вирусов -ssRNA обычно линейны, и их геном обычно сегментирован.
Вирусы с отрицательной цепью РНК составляют тип Negarnaviricota , в царстве Orthornavirae и царстве Riboviria . Они произошли от общего предка, который представлял собой вирус с двухцепочечной РНК (дцРНК) , и считаются сестринской кладой реовирусов , которые представляют собой вирусы с дцРНК. Внутри типа есть две основные ветви, которые образуют два подтипа: Haploviricotina , представители которого в основном несегментированы и кодируют RdRp, который синтезирует кэпы на мРНК, и Polyploviricotina , члены которого сегментированы и кодируют RdRp, который отрывает кэпы от мРНК. мРНК хозяина. Всего признано шесть классов этого типа.
РНК-вирусы с отрицательной цепью тесно связаны с членистоногими , и их можно неофициально разделить на те, передача которых зависит от членистоногих, и те, которые произошли от вирусов членистоногих, но теперь могут реплицироваться у позвоночных без помощи членистоногих. Известные вирусы -ssRNA, переносимые членистоногими, включают вирус лихорадки Рифт-Валли и вирус пятнистого увядания томатов . Известные вирусы -оцРНК позвоночных включают вирус Эбола , хантавирусы , вирусы гриппа , вирус лихорадки Ласса и вирус бешенства .
Этимология
[ редактировать ]Negarnaviricota берет первую часть своего названия от латинского nega , что означает отрицательный, средняя часть РНК относится к РНК, а последняя часть, viricota , является суффиксом, используемым для вирусных типов. Подтип Haploviricotina берет первую часть своего названия, Haplo , от древнегреческого ἁπλός, что означает простой, а «вирикотина» — это суффикс, используемый для подтипов вирусов. Подтип Polyploviricotina следует той же схеме: Polyplo происходит от древнегреческого πολύπλοκος, что означает комплекс. [ 1 ]
Характеристики
[ редактировать ]Геном
[ редактировать ]
Все вирусы Negarnaviricota представляют собой вирусы с одноцепочечной РНК (-оцРНК) с отрицательным смыслом. Их геномы состоят из РНК, которая является одноцепочечной, а не двухцепочечной. Их геномы имеют отрицательный смысл, что означает, что информационная РНК (мРНК) может быть синтезирована непосредственно из генома с помощью вирусного фермента РНК-зависимой РНК-полимеразы (RdRp), также называемой РНК-репликазой, которая кодируется всеми вирусами -оцРНК. За исключением вирусов рода Tenuivirus и некоторых вирусов семейства Chuviridae , все вирусы -ssRNA имеют линейные, а не кольцевые геномы, и геномы могут быть сегментированными или несегментированными. [ 1 ] [ 3 ] [ 4 ] Все геномы -оцРНК содержат терминальные инвертированные повторы , которые представляют собой палиндромные нуклеотидные последовательности на каждом конце генома. [ 5 ]
Репликация и транскрипция
[ редактировать ]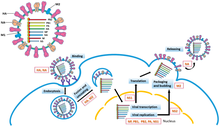
Репликация геномов -оцРНК осуществляется с помощью RdRp, который инициирует репликацию путем связывания с лидерной последовательностью на 3'-конце (обычно произносится как «три простых конца») генома. Затем RdRp использует геном с отрицательным смыслом в качестве матрицы для синтеза антигенома с положительным смыслом. При репликации антигенома RdRp сначала связывается с концевой последовательностью на 3'-конце антигенома. После этого RdRp игнорирует все сигналы транскрипции на антигеноме и синтезирует копию генома, используя антигеном в качестве матрицы. [ 6 ] Репликация выполняется, пока геном находится внутри нуклеокапсида, а RdRp раскрывает капсид и перемещается по геному во время репликации. По мере того как новые нуклеотидные последовательности синтезируются с помощью RdRp, капсидные белки собираются и инкапсидируют вновь реплицирующуюся вирусную РНК. [ 2 ]
Транскрипция мРНК из генома следует той же схеме, что и производство антигенома. В лидерной последовательности RdRp синтезирует 5'-конец (обычно произносится как «пять простых концов») трифосфат-лидерную РНК и либо, в случае подтипа Haploviricotina , блокирует 5'-конец, либо, в случае подтипа, Polyploviricotina отрывает колпачок от мРНК хозяина и прикрепляет его к вирусной мРНК, чтобы мРНК могла транслироваться клетки-хозяина рибосомами . [ 7 ] [ 8 ] [ 9 ]
После кэпирования мРНК RdRp инициирует транскрипцию по сигналу начала гена, а затем прекращает транскрипцию при достижении сигнала конца гена. [ 10 ] В конце транскрипции RdRp синтезирует полиаденилированный хвост (поли (А) хвост), состоящий из сотен аденинов на 3-конце мРНК, что можно сделать путем заикания последовательности урацилов . [ 11 ] [ 12 ] После построения поли(А)-хвоста мРНК высвобождается с помощью RdRp. В геномах, которые кодируют более одной транскрибируемой части, RdRp может продолжать сканирование до следующей стартовой последовательности, чтобы продолжить транскрипцию. [ 10 ] [ 7 ] [ 13 ]
Некоторые вирусы -ssRNA являются амбисенсными , что означает, что как отрицательная цепь генома, так и положительный антигеном по отдельности кодируют разные белки. Чтобы транскрибировать амбисенс-вирусы, выполняются два раунда транскрипции: сначала мРНК производится непосредственно из генома; во-вторых, мРНК создается из антигенома. Все амбисенс-вирусы содержат структуру шпильки, позволяющую остановить транскрипцию после транскрипции мРНК белка. [ 14 ]
Морфология
[ редактировать ]
Вирусы с отрицательной цепью РНК содержат рибонуклеопротеиновый комплекс, состоящий из генома и RdRp, прикрепленного к каждому сегменту генома, окруженного капсидом. [ 15 ] Капсид состоит из белков, складчатая структура которых содержит пять альфа-спиралей в N-концевой доле (мотив 5-H) и три альфа-спирали в C-концевой доле (мотив 3-H). Внутри капсида геном зажат между этими двумя мотивами. [ 2 ] За исключением семейства Aspiviridae , вирусы -ssRNA содержат внешнюю вирусную оболочку , тип липидной мембраны, окружающей капсид. Форма вирусной частицы, называемой вирионом, вирусов -ssRNA варьируется и может быть нитевидной, плеоморфной, сферической или трубчатой. [ 16 ]
Эволюция
[ редактировать ]Сегментация генома является характерной чертой многих вирусов -ssRNA, а вирусы -ssRNA варьируются от геномов с одним сегментом, типичных для представителей отряда Mononegavirales , до геномов с десятью сегментами, как в случае с Tilapia tilapinevirus . [ 5 ] [ 17 ] Не существует четкой тенденции с течением времени, определяющей количество сегментов, а сегментация генома среди -оцРНК-вирусов, по-видимому, является гибкой чертой, поскольку она неоднократно развивалась независимо. Большинство представителей подтипа Haploviricotina несегментированы, тогда как сегментация универсальна для Polyploviricotina . [ 2 ] [ 5 ]
Филогенетика
[ редактировать ]
Филогенетический анализ, основанный на RdRp, показывает, что вирусы -оцРНК произошли от общего предка и что они, вероятно, являются сестринской кладой реовирусов , которые представляют собой вирусы дцРНК. Внутри типа есть две четкие ветви, отнесенные к двум подтипам, в зависимости от того, синтезирует ли RdRp кэп на вирусной мРНК или отрывает кэп от мРНК хозяина и прикрепляет этот кэп к вирусной мРНК. [ 1 ] [ 3 ]
Внутри этого типа вирусы -оцРНК, поражающие членистоногих, по-видимому, являются базальными и предками всех других вирусов -оцРНК. Членистоногие часто живут вместе большими группами, что позволяет легко передавать вирусы. Со временем это привело к тому, что вирусы членистоногих -ssRNA приобрели высокий уровень разнообразия. Хотя членистоногие являются носителями большого количества вирусов, существуют разногласия относительно степени межвидовой передачи вирусов членистоногих -ssRNA среди членистоногих. [ 4 ] [ 5 ]
Вирусы -оцРНК растений и позвоночных, как правило, генетически связаны с вирусами, инфицированными членистоногими. Более того, большинство вирусов -ssRNA вне членистоногих обнаруживаются у видов, взаимодействующих с членистоногими. Таким образом, членистоногие служат как ключевыми хозяевами, так и векторами передачи вирусов -оцРНК. С точки зрения передачи вирусы -оцРНК, не относящиеся к членистоногим, можно разделить на те, передача которых зависит от членистоногих, и те, которые могут циркулировать среди позвоночных без помощи членистоногих. Последняя группа, вероятно, произошла от первой и адаптировалась к передаче инфекции только позвоночным. [ 5 ]
Классификация
[ редактировать ]Negarnaviricota принадлежит к царству Orthornavirae , которое включает в себя все РНК-вирусы, кодирующие RdRp, и к царству Riboviria , которое включает Orthornavirae, а также все вирусы, кодирующие обратную транскриптазу, в царстве Pararnavirae . Negarnaviricota содержит два подтипа, которые содержат в совокупности шесть классов, пять из которых монотипичны вплоть до низших таксонов: [ 2 ] [ 9 ] [ 18 ]
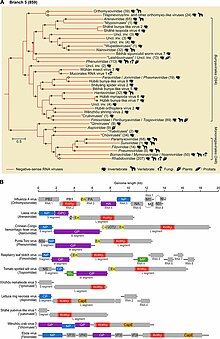
- Подтип: Haploviricotina , который содержит вирусы -оцРНК, кодирующие RdRp, который синтезирует структуру кэпа на вирусной мРНК и которые обычно имеют несегментированные геномы.
- Класс: Чунквивирицетес
- Отряд: Мувиралес
- Семейство: Qinviridae .
- Род: Yingvirus
- Семейство: Qinviridae .
- Отряд: Мувиралес
- Класс: Милневирицетес
- Отряд: Серпентовирусы.
- Семейство: Aspviridae.
- Род: Офиовирус.
- Семейство: Aspviridae.
- Отряд: Серпентовирусы.
- Класс: Монхивирицетес
- Класс: Юнчанвирицетес
- Отряд: Goujianvirales
- Семейство: Юэвириды.
- Род: Yuyuevirus.
- Семейство: Юэвириды.
- Отряд: Goujianvirales
- Класс: Чунквивирицетес
- Подтип: Polyploviricotina , который содержит вирусы -оцРНК, кодирующие RdRp, который берет кэп из мРНК хозяина для использования в качестве кэпа на вирусной мРНК и имеет сегментированные геномы.
- Класс: Эллиовирицетес
- Отряд: Буньявирусы.
- Класс: Insthoviricetes
- Отряд: Articulavirales.
- Класс: Эллиовирицетес
Вирусы с отрицательной цепью РНК классифицируются как Группа V в Балтиморской классификационной системе, которая объединяет вирусы в зависимости от способа производства мРНК и которая часто используется наряду со стандартной таксономией вирусов, основанной на истории эволюции. Таким образом, группа V и Negarnaviricota являются синонимами. [ 1 ]
Болезнь
[ редактировать ]Вирусы с отрицательной цепью РНК вызывают многие широко известные заболевания. Многие из них передаются членистоногими, в том числе вирус лихорадки Рифт-Валли и вирус пятнистого увядания томатов . [ 19 ] [ 20 ] Среди позвоночных летучие мыши и грызуны являются распространенными переносчиками многих вирусов, включая вирус Эбола и вирус бешенства , передающихся летучими мышами и другими позвоночными животными. [ 21 ] [ 22 ] вирус лихорадки Ласса и хантавирусы , передаваемые грызунами. [ 23 ] [ 24 ] Вирусы гриппа распространены среди птиц и млекопитающих. [ 25 ] Специфические для человека вирусы -ssRNA включают вирус кори и вирус эпидемического паротита . [ 26 ] [ 27 ]
История
[ редактировать ]Многие заболевания, вызываемые вирусами -ssRNA, были известны на протяжении всей истории, включая хантавирусную инфекцию, корь и бешенство. [ 28 ] [ 29 ] [ 30 ] В современной истории некоторые из них, такие как Эбола и грипп, вызывали вспышки смертельных заболеваний. [ 31 ] [ 32 ] Вирус везикулярного стоматита , впервые выделенный в 1925 году и один из первых вирусов животных, подлежащих изучению, поскольку его можно было хорошо изучить в культурах клеток , был идентифицирован как вирус -оцРНК, который был уникальным в то время, поскольку другие РНК-вирусы, которые были обнаружили положительный смысл. [ 33 ] [ 34 ] В начале XXI века чума крупного рогатого скота , вызываемая -ssRNA вирусом чумы крупного рогатого скота, стала второй болезнью, которую удалось искоренить, после оспы , вызываемой ДНК-вирусом. [ 35 ]
В 21 веке вирусная метагеномика стала обычным явлением для идентификации вирусов в окружающей среде. Для вирусов -ssRNA это позволило идентифицировать большое количество вирусов беспозвоночных, и особенно членистоногих, что помогло пролить свет на историю эволюции вирусов -ssRNA. На основании филогенетического анализа RdRp, показавшего, что вирусы -ssRNA произошли от общего предка, Negarnaviricota и два его подтипа были созданы в 2018 году, и они были помещены в недавно созданную область Riboviria . [ 1 ] [ 36 ]
Галерея
[ редактировать ]-

-

-

-

-

-

-
 Корь ( Парамиксовирусы )
Корь ( Парамиксовирусы ) -

-

-

-

-













Примечания
[ редактировать ]- ^ Вирус гепатита D часто называют вирусом, но более конкретно его можно описать как вирусоидоподобную патогенную цепь -оцРНК. Он исключен из Negarnaviricota, поскольку, хотя он и является -оцРНК, он не кодирует RdRp, который является объединяющим признаком вирусов Orthornavirae .
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и ж Вольф Ю., Крупович М., Чжан Ю.З., Маес П., Должи В., Кунин Е.В. (21 августа 2017 г.). «Мегатаксономия РНК-вирусов с отрицательным смыслом» (docx) . Международный комитет по таксономии вирусов (ICTV) . Проверено 6 августа 2020 г.
- ^ Перейти обратно: а б с д и Луо М., Террел-младший, Макманус С.А. (30 июля 2020 г.). «Нуклеокапсидная структура вируса с отрицательной цепью РНК» . Вирусы . 12 (8): 835. дои : 10.3390/v12080835 . ПМЦ 7472042 . ПМИД 32751700 .
- ^ Перейти обратно: а б Вольф Й.И., Казлаускас Д., Иранзо Дж., Люсия-Санс А., Кун Дж.Х., Крупович М., Доля В.В., Кунинг Е.В. (27 ноября 2018 г.). «Происхождение и эволюция глобального РНК-вирома» . мБио 9 (6): e02329-1 дои : 10.1128/mBio.02329-18 . ПМК 6282212 . ПМИД 30482837 .
- ^ Перейти обратно: а б Кефер С., Параскевопулу С., Циркель Ф., Визеке Н., Донат А., Петерсен М., Джонс Т.К., Лю С., Чжоу Х, Миддендорф М., Юнглен С., Мисоф Б., Дростен С. (12 декабря 2019 г.). «Повторная оценка разнообразия РНК-вирусов с отрицательной цепью у насекомых» . ПЛОС Патог . 15 (12): e1008224. дои : 10.1371/journal.ppat.1008224 . ПМЦ 6932829 . ПМИД 31830128 .
- ^ Перейти обратно: а б с д и Ли CX, Ши М, Тянь Дж. Х., Линь XD, Кан YJ, Чен LJ, Цинь XC, Сюй Дж, Холмс EC, Чжан YZ (29 января 2015 г.). «Беспрецедентное геномное разнообразие РНК-вирусов у членистоногих указывает на происхождение РНК-вирусов с отрицательным смыслом» . электронная жизнь . 4 (4): e05378. doi : 10.7554/eLife.05378 . ПМЦ 4384744 . ПМИД 25633976 .
- ^ «Репликация вируса с отрицательной цепью РНК» . ВиралЗона . Швейцарский институт биоинформатики . Проверено 6 августа 2020 г.
- ^ Перейти обратно: а б «Транскрипция вируса с отрицательной цепью РНК» . ВиралЗона . Швейцарский институт биоинформатики . Проверено 6 августа 2020 г.
- ^ «Похищение кепки» . ВиралЗона . Швейцарский институт биоинформатики . Проверено 6 августа 2020 г.
- ^ Перейти обратно: а б Кун Дж.Х., Вольф Ю.И., Крупович М., Чжан Ю.З., Маес П., Доля В.В., Кунин Е.В. (февраль 2019 г.). «Классифицируйте вирусы – выгода стоит затраченных усилий» . Природа . 566 (7744): 318–320. Бибкод : 2019Natur.566..318K . дои : 10.1038/d41586-019-00599-8 . ПМЦ 11165669 . ПМИД 30787460 . S2CID 67769904 . Проверено 6 августа 2020 г.
- ^ Перейти обратно: а б Фернс, Рэйчел; Племпер, Ричард К. (15 апреля 2017 г.). «Полимеразы парамиксовирусов и пневмовирусов» . Вирусные исследования . 234 : 87–102. doi : 10.1016/j.virusres.2017.01.008 . ISSN 0168-1702 . ПМЦ 5476513 . ПМИД 28104450 .
- ^ Хармон, Шон Б.; Мего, А. Джордж; Вертц, Гейл В. (1 января 2001 г.). «Последовательности РНК, участвующие в терминации транскрипции респираторно-синцитиального вируса» . Журнал вирусологии . 75 (1): 36–44. doi : 10.1128/JVI.75.1.36-44.2001 . ПМЦ 113895 . ПМИД 11119571 .
- ^ Жак, JP; Колакофски, Д. (1 мая 1991 г.). «Псевдошаблонная транскрипция в прокариотических и эукариотических организмах» . Гены и развитие . 5 (5): 707–713. дои : 10.1101/gad.5.5.707 . ISSN 0890-9369 . ПМИД 2026325 .
- ^ «Заикание полимеразы отрицательноцепочечной РНК-вируса» . ВиралЗона . Швейцарский институт биоинформатики . Проверено 6 августа 2020 г.
- ^ «Амбисенс-транскрипция в вирусах с отрицательной цепью РНК» . ВиралЗона . Швейцарский институт биоинформатики . Проверено 6 августа 2020 г.
- ^ Чжоу Х, Сунь Ю, Го Ю, Лу Цзы (сентябрь 2013 г.). «Структурный взгляд на образование рибонуклеопротеинового комплекса в вирусах с одноцепочечной РНК с отрицательным смыслом». Тенденции Микробиол . 21 (9): 475–484. дои : 10.1016/j.tim.2013.07.006 . ПМИД 23953596 .
- ^ Фермин, Г. (2018). Вирусы: молекулярная биология, взаимодействия с хозяином и приложения к биотехнологии . Эльзевир. стр. 19–27, 43. doi : 10.1016/B978-0-12-811257-1.00002-4 . ISBN 9780128112571 . S2CID 89706800 .
- ^ Бахарах Э , Мишра Н , Бриз Т , Зоди MC , Кембу Цофак Дж. Е , Замостиано Р , Берковиц А , Нг Дж , Нитидо А , Корвело А , Туссен NC , Абель Нильсен СК , Хорниг М , Дель Позо Дж , Блум Т , Фергюсон Х , Эльдар А., Липкин В.И. (5 апреля 2016 г.). «Характеристика нового ортомиксоподобного вируса, вызывающего массовую гибель тилапии» . мБио 7 (2):e00431-16. дои : 10.1128/mBio.00431-16 . ПМЦ 4959514 . ПМИД 27048802 .
- ^ «Таксономия вирусов: выпуск 2019 г.» . talk.ictvonline.org . Международный комитет по таксономии вирусов . Проверено 6 августа 2020 г.
- ^ Хартман А. (июнь 2017 г.). «Лихорадка Рифт-Валли» . Клин Лаб Мед . 37 (2): 285–301. дои : 10.1016/j.cll.2017.01.004 . ПМЦ 5458783 . ПМИД 28457351 .
- ^ Шольтхоф К.Б., Адкинс С., Чоснек Х., Палукайтис П., Жако Э., Хон Т., Хон Б., Сондерс К., Кандресс Т., Алквист П., Хеменуэй С., Фостер Г.Д. (декабрь 2011 г.). «10 главных растительных вирусов в молекулярной патологии растений» . Мол Плант Патол . 12 (9): 938–954. дои : 10.1111/j.1364-3703.2011.00752.x . ПМК 6640423 . ПМИД 22017770 .
- ^ Муньос-Фонтела С., МакЭлрой АК (2017). «Болезнь, вызванная вирусом Эбола, у человека: патофизиология и иммунитет» . Curr Top Microbiol Immunol . Актуальные темы микробиологии и иммунологии. 411 : 141–169. дои : 10.1007/82_2017_11 . ISBN 978-3-319-68946-3 . ПМК 7122202 . ПМИД 28653186 .
- ^ Фишер Ч.Р., Штрейкер Д.Г., Шнелл М.Дж. (апрель 2018 г.). «Распространение и эволюция вируса бешенства: покорение новых рубежей» . Nat Rev Микробиол . 16 (4): 241–255. дои : 10.1038/nrmicro.2018.11 . ПМК 6899062 . ПМИД 29479072 .
- ^ Юн Н.Э., Уокер Д.Х. (9 октября 2012 г.). «Патогенез лихорадки Ласса» . Вирусы . 4 (10): 2031–2048. дои : 10.3390/v4102031 . ПМК 3497040 . ПМИД 23202452 .
- ^ Авшич-Жупанц Т., Саксида А., Корва М. (апрель 2019 г.). «Хантавирусные инфекции» . Клин Микробиол Инфект . 21С : е6–е16. дои : 10.1111/1469-0691.12291 . ПМИД 24750436 . Проверено 6 августа 2020 г.
- ^ Боркенхаген Л.К., Салман, доктор медицинских наук, Ма М.Дж., Грей Г.К. (ноябрь 2019 г.). «Инфекции человека вирусом гриппа животных: комментарий» . Int J Заразить Дис . 88 : 113–119. дои : 10.1016/j.ijid.2019.08.002 . ПМИД 31401200 . Проверено 6 августа 2020 г.
- ^ «Передача кори» . cdc.gov . Центры по контролю и профилактике заболеваний (CDC). 5 февраля 2018 года . Проверено 6 августа 2020 г.
- ^ Рубин С., Экхаус М., Ренник Л.Дж., Бэмфорд К.Г., Дюпрекс В.П. (январь 2015 г.). «Молекулярная биология, патогенез и патология вируса паротита» . Дж. Патол . 235 (2): 242–252. дои : 10.1002/путь.4445 . ПМК 4268314 . ПМИД 25229387 .
- ^ Цзян Х, Чжэн Икс, Ван Л, Ду Х, Ван П, Бай Икс (2017). «Хантавирусная инфекция: глобальная зоонозная проблема» . Вирол Син . 32 (1): 32–43. дои : 10.1007/s12250-016-3899-x . ПМК 6598904 . ПМИД 28120221 .
- ^ «История кори» . cdc.gov . Центры по контролю и профилактике заболеваний (CDC). 5 февраля 2018 года . Проверено 6 августа 2020 г.
- ^ Веласко-Вилла А, Молдин М.Р., Ши М., Эскобар Л.Е., Галлардо-Ромеро Н.Ф., Дэймон И., Олсон В.А., Штрейкер Д.Г., Эмерсон Г. (октябрь 2017 г.). «История бешенства в Западном полушарии» . Противовирусный Рес . 146 : 221–232. дои : 10.1016/j.antiviral.2017.03.013 . ПМК 5620125 . ПМИД 28365457 .
- ^ Завилинска Б, Кош-Вненчак М (2014). «Общее введение в биологию и заболевание вируса Эбола» (PDF ) Фолиа Мед Краков . 54 (3): 57–65. ПМИД 25694096 . Получено 6 августа.
- ^ Краммер Ф., Смит Г., Фушье Р., Пейрис М., Кедзерска К., Доэрти П.С., Палезе П., Шоу М.Л., Треанор Дж., Вебстер Р.Г., Гарсиа-Састре А. (28 июня 2018 г.). «Грипп» . Праймеры Nat Rev Dis . 4 (1): 3. doi : 10.1038/s41572-018-0002-y . ПМК 7097467 . ПМИД 29955068 .
- ^ «Вирус везикулярного стоматита» (PDF) . Информационный центр здоровья свиней . Центр продовольственной безопасности и общественного здравоохранения Колледжа ветеринарной медицины Университета штата Айова. Ноябрь 2015 года . Проверено 6 августа 2020 г.
- ^ Колакофский Д. (апрель 2015 г.). «Краткая предвзятая история РНК-вирусов» . РНК . 21 (4): 667–669. дои : 10.1261/rna.049916.115 . ПМЦ 4371325 . ПМИД 25780183 . Проверено 6 августа 2020 г.
- ^ Гринвуд Б (12 мая 2014 г.). «Вклад вакцинации в глобальное здравоохранение: прошлое, настоящее и будущее» . Philos Trans R Soc Lond B Biol Sci . 369 (1645): 20130433. doi : 10.1098/rstb.2013.0433 . ПМК 4024226 . ПМИД 24821919 .
- ^ «История таксономии ICTV: Negarnaviricota» . Международный комитет по таксономии вирусов (ICTV) . Проверено 6 августа 2020 г.
Дальнейшее чтение
[ редактировать ]- Уорд, CW (1993). «Прогресс к более высокой систематике вирусов» . Исследования в области вирусологии . 144 (6): 419–53. дои : 10.1016/S0923-2516(06)80059-2 . ПМЦ 7135741 . ПМИД 8140287 .