Рабдовирусиды
Эта статья нуждается в дополнительных цитатах для проверки . ( декабрь 2012 г. ) |
| Рабдовирусиды | |
|---|---|
 | |
| Вирус везикулярного стоматита Индианы (ВСВ), прототипный рабдовирус | |
| Классификация вирусов | |
| (без рейтинга): | Вирус |
| Область : | Рибовирия |
| Королевство: | Орторнавиры |
| Тип: | Негарнавирикота |
| Сорт: | Монхивирицетес |
| Заказ: | Мононегавирусы |
| Семья: | Рабдовирусиды |
| Роды | |
Рабдовирусы — семейство РНК-вирусов с отрицательной цепью из отряда Mononegavirales . [1] позвоночные (включая млекопитающих и человека), беспозвоночные , растения , грибы и простейшие . Естественными хозяевами служат [2] [3] [4] Заболевания, связанные с вирусами-членами, включают бешенственный энцефалит, вызванный вирусом бешенства , и гриппоподобные симптомы у людей, вызванные везикуловирусами . Название происходит от древнегреческого слова «рабдос» , что означает «стержень», что связано с формой вирусных частиц. [5] В семействе 40 родов, большинство из которых относятся к трем подсемействам. [6]
Структура
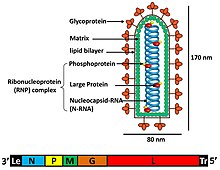
[ редактировать ]Отдельные вирусные частицы (вирионы) рабдовирусов состоят из РНК, белка, углеводов и липидов. Они имеют сложную палочковидную или пулевидную форму. Все эти вирусы имеют структурное сходство и отнесены к одному семейству. [7]
Вирионы имеют ширину около 75 нм и длину 180 нм. [2] Рабдовирусы имеют оболочку , спиральные нуклеокапсиды , а их геномы линейные, длиной около 11–15 т.п.н. [5] [2] Рабдовирусы несут свой генетический материал в форме с отрицательным смыслом одноцепочечной РНК . Обычно они несут гены пяти белков: большого белка (L), гликопротеина (G), нуклеопротеина (N), фосфопротеина (P) и матриксного белка (M). [8] Последовательность этих белковых генов от 3'-конца до 5'-конца в геноме имеет вид N–P–M–G–L. [9] Все рабдовирусы кодируют эти пять белков в своих геномах. Помимо этих белков, многие рабдовирусы кодируют один или несколько белков. [10] Первые четыре гена кодируют основные структурные белки, участвующие в строении оболочки вириона. [9]
Матриксный белок (М) образует слой между оболочкой вириона и ядром нуклеокапсида рабдовируса. [10] В дополнение к функциям, связанным со сборкой вируса, морфогенезом и отпочкованием от плазматической мембраны хозяина, были обнаружены дополнительные функции, такие как регуляция синтеза РНК, влияющая на баланс продуктов репликации и транскрипции, при проведении экспериментов по обратной генетике с вирусом бешенства. представитель семейства Rhabdoviridae. [11] Большой белок (L) выполняет несколько ферментативных функций в синтезе и процессинге вирусной РНК. [8] Ген L кодирует этот белок L, который содержит несколько доменов. Считается, что помимо синтеза РНК он участвует в активности метилкэпирования и полиаденилирования. [9]
Белок P играет важную и разнообразную роль во время транскрипции и репликации генома РНК. Многофункциональный белок P кодируется геном P. Белок P действует как некаталитический кофактор большой протеинполимеразы. Он связывается с белками N и L. Белок P имеет две независимые области связывания. Образуя комплексы NP, он может сохранять белок N в форме, подходящей для специфической инкапсуляции. Белок P вмешивается в врожденную иммунную систему хозяина посредством ингибирования активности регуляторного фактора интерферона 3 (IRF3) и преобразователя сигнала и активатора транскрипции 1 (STAT1), тем самым устраняя путь интерферона клеточного типа 1. Кроме того, белок P действует как антагонист противовирусной функции PML. [12] [13]
Рабдовирусы, поражающие позвоночных (особенно млекопитающих и рыб), растения и насекомых, обычно имеют пулевидную форму. [14] Однако в отличие от парамиксовирусов рабдовирусы не обладают гемагглютинирующей и нейраминидазной активностью. [14]
Транскрипция
[ редактировать ]
Транскриптаза рабдовируса состоит из белков 1 L и 3 P. Компоненты транскриптазы всегда присутствуют в полном вирионе, что позволяет рабдовирусам начать транскрипцию сразу после проникновения. [ нужна ссылка ]
Транскриптаза рабдовируса действует в направлении от 3' к 5' генома, и транскрипция заканчивается случайным образом на конце белковой последовательности. Например, если транскрипция заканчивается в конце последовательности M; лидерная РНК и мРНК N, P и M образуются отдельно друг от друга. [ нужна ссылка ]
Кроме того, мРНК накапливаются в соответствии с порядком белковых последовательностей в геноме, решая проблему логистики в клетке. Например, белок N необходим вирусу в больших количествах, поскольку он полностью покрывает внешнюю часть реплицируемых геномов. Поскольку последовательность N-белка расположена в начале генома (3'-конец) после лидерной последовательности РНК, мРНК N-белка всегда могут продуцироваться и накапливаться в больших количествах при каждом прекращении транскрипции. После процессов транскрипции все мРНК кэпируются на 5'-конце и полиаденилируются на 3'-конце с помощью L-белка.
Таким образом, этот механизм транскрипции обеспечивает мРНК в соответствии с потребностями вирусов. [10] : 173–184
Перевод
[ редактировать ]Белки вируса транслируются на свободных рибосомах, а белок G транслируется шероховатой эндоплазматической сетью. Это означает, что G-белок имеет сигнальный пептид в стартовых кодах своей мРНК. Фосфопротеины (P) и гликопротеин (G) подвергаются посттрансляционной модификации. Тримеры белка P образуются после фосфорилирования за счет киназной активности белка L. Белок G гликозилирован в шероховатой эндоплазматической сети и комплексе Гольджи. [10] : 180
Репликация
[ редактировать ]
Репликация вируса цитоплазматическая. Цикл репликации одинаков для большинства рабдовирусов. Все компоненты, необходимые для ранней транскрипции, и нуклеокапсид высвобождаются в цитоплазму инфицированной клетки после того, как происходят первые этапы связывания, проникновения и снятия покрытия. [9] Проникновение в клетку-хозяина достигается путем прикрепления вирусных G-гликопротеинов к рецепторам хозяина, что опосредует клатрин-опосредованный эндоцитоз. Репликация соответствует модели репликации вируса с отрицательной цепью РНК. Транскрипция вируса с отрицательной цепью РНК с использованием заикания полимеразы является методом транскрипции. Вирус покидает клетку-хозяина путем почкования и перемещения вируса по канальцам. Пути передачи – зооноз и укус. [5] [2]
Репликация многих рабдовирусов происходит в цитоплазме , хотя некоторые вирусы, инфицирующие растения, реплицируются в ядре. [15] Белок матрикса (М) рабдовируса очень мал (~ 20–25 кДа), однако играет ряд важных ролей во время цикла репликации вируса. Эти белки рабдовирусов составляют основные структурные компоненты вируса, они являются многофункциональными белками и необходимы для созревания вируса и процесса почкования вируса, а также регулируют баланс синтеза вирусной РНК путем переключения синтеза от транскрипции к репликации. [16] Для репликации должны быть экспрессированы белки L и P для регулирования транскрипции . [17] Фосфопротеин (P) также играет решающую роль во время репликации, поскольку для соответствующего и селективного инкапсидирования вирусной РНК необходимы комплексы NP, а не только N. Следовательно, репликация невозможна после заражения до тех пор, пока первичная транскрипция и трансляция не продуцируют достаточное количество N-белка. [18]

Белок L обладает высокой ферментативной активностью, такой как репликация РНК, кэпирование мРНК, фосфорилирование белка P. L дает возможность репликации в цитоплазме. [17] пяти моноцистронных мРНК Транскрипция приводит к образованию , поскольку межгенные последовательности действуют как терминальные , так и промоторные последовательности для соседних генов . Этот тип механизма транскрипции объясняется моделью стоп-старт (заикающаяся транскрипция). Благодаря модели «стоп-старт» производится большое количество структурных белков. Согласно этой модели, вирус-ассоциированная РНК-полимераза начинает сначала синтез лидерной РНК, а затем пяти мРНК, которые будут продуцировать белки N, P, M, G, L соответственно. После образования лидерной РНК фермент полимераза повторно инициирует транскрипцию вириона на гене N и продолжает его синтез до тех пор, пока не закончится 3'-конец цепи. Затем синтез P-мРНК осуществляется тем же ферментом с новым стартовым синилалом. Эти шаги продолжаются до тех пор, пока фермент не достигнет конца гена L. Во время процесса транскрипции фермент полимераза может покинуть матрицу в любой точке, а затем связаться только с 3'-концом геномной РНК, чтобы снова начать синтез мРНК. Этот процесс приведет к градиенту концентрации количества мРНК в зависимости от ее места и диапазона от 3'-конца. В этих обстоятельствах количество видов мРНК меняется, и будут вырабатываться белки N>P>M>G>L. [19] Во время синтеза мРНК подвергаются процессингу для введения 5'-кэпа и 3'-полиаденилированного хвоста к каждой из молекул. Эта структура гомологична клеточным мРНК и, таким образом, может транслироваться клеточными рибосомами с образованием как структурных, так и неструктурных белков.
Репликация генома требует источника вновь синтезированного N-белка для инкапсида РНК. Это происходит во время его синтеза и приводит к созданию полноразмерной антигеномной копии. Это, в свою очередь, используется для производства большего количества геномной РНК с отрицательным смыслом. Для этого процесса необходима вирусная полимераза, но то, как полимераза участвует как в синтезе мРНК, так и в репликации генома, не совсем понятно.
Репликация обычно происходит во включении в цитоплазме, откуда они отпочковываются через различные цитоплазматические мембраны и внешнюю мембрану клетки. Этот процесс приводит к приобретению белков M+G, ответственных за характерную пулевидную морфологию вируса .
Классификация
[ редактировать ]Клады
[ редактировать ]Эти вирусы делятся на четыре группы в зависимости от гена РНК-полимеразы. [20] Базальная клада, по-видимому, представляет собой новирхабдовирусы , поражающие рыб. Циторхабдовирусы и нуклеорхабдовирусы , поражающие растения, являются сестринскими кладами. Лиссавирусы образуют собственную кладу, которая более тесно связана с кладами наземных позвоночных и насекомых, чем с вирусами растений. Остальные вирусы образуют ряд сильно разветвленных клад и поражают членистоногих и наземных позвоночных.
Анализ 99 видов рабдовирусов животных, проведенный в 2015 году, показал, что они делятся на 17 таксономических групп, восемь из которых — лиссавирусы , везикуловирусы , перхабдовирусы , сигмавирусы , эфемеровирусы , тибровирусы , тупавирусы и спрививирусы — которые были признаны ранее. [21] На основе полученных результатов авторы предложили семь новых таксонов: «Альмендравирус», «Бахиавирус», «Куриовирус», «Гапавирус», «Ледантевирус», «Савгравирус» и «Срипувирус». Семь видов не сгруппировались с остальными, что указывает на необходимость дополнительных таксонов.
Предлагаемые классификации
[ редактировать ]Неофициальная супергруппа – «Димархабдовирусы» – относится к родам Эфемеровирус и Везикуловирус . [22] К этому таксону относится и ряд других вирусов, не классифицированных по родам. В эту супергруппу входят роды, виды которых реплицируются как у позвоночных, так и у беспозвоночных-хозяев и имеют биологические циклы, включающие передачу гематофагами- двукрылыми (кровососущими мухами).
Прототипические рабдовирусы
[ редактировать ]Прототипическим и наиболее изученным рабдовирусом является вирус везикулярного стоматита Индианы . Это предпочтительная модельная система для изучения биологии рабдовирусов и мононегавирусов в целом. Заболевание млекопитающих бешенством . вызывается лиссавирусами, несколько из которых были идентифицированы
Рабдовирусы являются важными патогенами животных и растений. Рабдовирусы передаются хозяину членистоногими, такими как тли, цикадки, цикадки, мошки, москиты и комары.
В сентябре 2012 года исследователи, пишущие в журнале PLOS Pathogens, описали новый вид рабдовируса, названный вирусом Бас-Конго (BASV), который был обнаружен в образце крови пациента, перенесшего болезнь, напоминающую геморрагическую лихорадку. [20] С момента его открытия не было зарегистрировано ни одного случая BASV, и неясно, был ли BASV фактической причиной заболевания пациента. [23]
В 2015 году два новых рабдовируса, вирус Экпома 1 и вирус Экпома 2, были обнаружены в образцах крови двух здоровых женщин на юго-западе Нигерии. Вирус Экпома 1 и вирус Экпома 2, по-видимому, хорошо реплицируются у людей (вирусная нагрузка варьируется от ~45 000 до ~4,5 миллионов копий РНК/мл плазмы), но не вызывают каких-либо заметных симптомов заболевания. [24] Воздействие вируса Экпома 2, по-видимому, широко распространено в некоторых частях Нигерии, где уровень серологической распространенности приближается к 50%. [24]
Таксономия
[ редактировать ]В подсемействе Alpharhabdovirinae выделяют следующие роды: [6]
- Альмендравирус
- Альфанемравирус
- Альфапаправирус
- Альфарицинравирус
- Арурхавирус
- бархавирус
- Калигравирус
- Куриовирус
- Эфемеровирус
- Гапавирус
- Ледантевирус
- Лостравирус
- Лиссавирус
- Мерхавирус
- Мусравирус
- Олсравирус
- Перхабдовирус
- Согравирус
- Сигмавирус
- Спрививирус
- Шрипувирус
- Санравирус
- Тибровирус
- Тупавирус
- Везикуловирус
- Зархавирус
Роды остальных подсемейств следующие: [6]
- Бетархабдовиринае
- Альфануклеорхабдовирус (в настоящее время; см. нуклеорхабдовирус )
- Бетануклеорхабдовирус (в настоящее время; см. нуклеорхабдовирус )
- Циторхабдовирус
- Дихорхавирус
- Гаммануклеорхабдовирус (в настоящее время; см. нуклеорхабдовирус )
- Варикозавирус
- Гаммархабдовирины
Следующие роды не отнесены к подсемейству: [6]
- Альфакрустравирус
- Альфадросравирус
- Альфагимравирус
- бетагимравирус
- Бетанемравирус
- Бета-паправирус
- Бетарицинравирус
Помимо вышеперечисленных, существует большое количество рабдоподобных вирусов, которые еще не классифицированы официально ICTV . [5]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Уокер П.Дж., Бласделл К.Р., Калишер Ч. , Дицген Р.Г., Кондо Х., Курат Г. и др. (апрель 2018 г.). «Профиль таксономии вируса ICTV: Rhabdoviridae» . Журнал общей вирусологии . 99 (4): 447–448. дои : 10.1099/jgv.0.001020 . hdl : 10871/31776 . ПМИД 29465028 .
- ^ Перейти обратно: а б с д «Вирусная зона» . ЭксПАСи . Проверено 15 июня 2015 г.
- ^ Берд, Р.Г.; МакКол, TF (март 1976 г.). «Рабдовирусы Entamoeba histolytica и Entamoeba инвазий». Анналы тропической медицины и паразитологии . 70 (1): 81–93. дои : 10.1080/00034983.1976.11687098 . ПМИД 178279 .
- ^ Фань Му, Бо Ли, Шуфэнь Ченг, Цзичун Цзя, Даохун Цзян, Яньпин Фу, Цзясен Ченг, Ян Линь, Тао Чен, Цзятао Се (2021). Девять вирусов из восьми линий, демонстрирующих новые эволюционные пути, которые одновременно заражают гиповирулентные фитопатогены. грибок Плос Патогены.
- ^ Перейти обратно: а б с д «Семейство: Rhabdoviridae | ICTV» . www.ictv.global .
- ^ Перейти обратно: а б с д «Таксономия вирусов: выпуск 2020 г.» . Международный комитет по таксономии вирусов (ICTV). Март 2021 года . Проверено 18 мая 2021 г.
- ^ Джексон, АО; Франки, РИБ; Зуйдема, Доуве (1987). «Биология, структура и репликация рабдовирусов растений». Рабдовирусы . стр. 427–508. дои : 10.1007/978-1-4684-7032-1_10 . ISBN 978-1-4684-7034-5 .
- ^ Перейти обратно: а б Огино М., Ито Н., Сугияма М., Огино Т. (май 2016 г.). «Белок L вируса бешенства катализирует блокирование мРНК с помощью полирибонуклеотидилтрансферазы GDP» . Вирусы . 8 (5): 144. дои : 10.3390/v8050144 . ПМЦ 4885099 . ПМИД 27213429 .
- ^ Перейти обратно: а б с д Ассенберг, Р.; Дельмас, О.; Морен, Б.; Грэм, Южная Каролина; Де Ламбаллери, X.; Лауберт, К.; Кутард, Б.; Граймс, Дж. М.; Нейтс, Дж.; Оуэнс, Р.Дж.; Брандт, BW; Горбаленя А.; Такер, П.; Стюарт, Д.И.; Канард, Б.; Бури, Х. (август 2010 г.). «Геномика и исследования структуры/функции белков Rhabdoviridae, участвующих в репликации и транскрипции» (PDF) . Противовирусные исследования . 87 (2): 149–161. дои : 10.1016/j.antiviral.2010.02.322 . ПМИД 20188763 . S2CID 8840157 .
- ^ Перейти обратно: а б с д Картер Дж.Б., Сондерс В.А. (2007). Вирусология: принципы и приложения . Чичестер, Англия: Джон Уайли и сыновья. ISBN 978-0-470-02386-0 . OCLC 124160564 .
- ^ Финке С., Конзельманн К.К. (ноябрь 2003 г.). «Диссоциация функций белков матрикса вируса бешенства в регуляции синтеза вирусной РНК и сборки вируса» . Журнал вирусологии . 77 (22): 12074–12082. doi : 10.1128/JVI.77.22.12074-12082.2003 . ПМК 254266 . ПМИД 14581544 .
- ^ Ван Л, Ву Х, Тао Икс, Ли Х, Рейнер С, Лян Г, Тан К (январь 2013 г.). «Генетическая и эволюционная характеристика RABV из Китая с использованием гена фосфопротеина» . Вирусологический журнал . 10 (1): 14. дои : 10.1186/1743-422X-10-14 . ПМЦ 3548735 . ПМИД 23294868 .
- ^ Окада К., Ито Н., Ямаока С., Масатани Т., Эбихара Х., Гото Х. и др. (сентябрь 2016 г.). Лайлс Д.С. (ред.). «Роль изоформ фосфопротеина вируса бешенства в патогенезе» . Журнал вирусологии . 90 (18): 8226–37. дои : 10.1128/JVI.00809-16 . ПМК 5008078 . ПМИД 27384657 .
- ^ Перейти обратно: а б Николас Х (2007). Основы молекулярной вирусологии . Англия: Уайли. стр. 175–187.
- ^ «Род: Alphanucleorhabdovirus - Rhabdoviridae - РНК-вирусы с отрицательным смыслом - ICTV» . talk.ictvonline.org . Архивировано из оригинала 23 июня 2021 года . Проверено 24 июня 2021 г.
- ^ Грэм С.С., Ассенберг Р., Дельмас О., Верма А., Голами А., Талби С. и др. (декабрь 2008 г.). «Белковые структуры матрикса рабдовируса обнаруживают новый способ самоассоциации» . ПЛОС Патогены . 4 (12): e1000251. дои : 10.1371/journal.ppat.1000251 . ПМК 2603668 . ПМИД 19112510 .
- ^ Перейти обратно: а б Ачесон, Нью-Хэмпшир (2011). Основы молекулярной вирусологии (2-е изд.). Джон Уайли и сыновья, Inc. ISBN 978-0470900598 .
- ^ Финке С., Конзельманн К.К. (ноябрь 2003 г.). «Диссоциация функций белков матрикса вируса бешенства в регуляции синтеза вирусной РНК и сборки вируса» . Журнал вирусологии . 77 (22): 12074–82. doi : 10.1128/JVI.77.22.12074-12082.2003 . ПМК 254266 . ПМИД 14581544 .
- ^ Маклахлан, Нью-Джерси, Дубови Э.Дж., ред. (2011). «Рабдовирусиды» . Ветеринарная вирусология Феннера . стр. 327–41. дои : 10.1016/B978-0-12-375158-4.00018-3 . ISBN 978-0-12-375158-4 .
- ^ Перейти обратно: а б Грард Дж., Фэйр Дж.Н., Ли Д., Сликас Э., Стеффен И., Муйембе Дж.Дж. и др. (сентябрь 2012 г.). «Новый рабдовирус, связанный с острой геморрагической лихорадкой в Центральной Африке» . ПЛОС Патогены . 8 (9): e1002924. дои : 10.1371/journal.ppat.1002924 . ПМЦ 3460624 . ПМИД 23028323 .
- ^ Уокер П.Дж., Ферт С., Уайден С.Г., Бласделл К.Р., Гузман Х., Вуд Т.Г. и др. (февраль 2015 г.). «Эволюция размера и сложности генома рабдовирусов» . ПЛОС Патогены . 11 (2): e1004664. дои : 10.1371/journal.ppat.1004664 . ПМЦ 4334499 . ПМИД 25679389 .
- ^ Бури Х., Коули Дж.А., Ларроуз Ф., Холмс Э.К., Уокер П.Дж. (октябрь 2005 г.). «Филогенетические взаимоотношения между рабдовирусами, выявленные с использованием гена L-полимеразы» . Журнал общей вирусологии . 86 (Часть 10): 2849–2858. дои : 10.1099/vir.0.81128-0 . ПМИД 16186241 .
- ^ Бранко Л.М., Гарри РФ (3 декабря 2018 г.). «Вирус Нижнего Конго – не установленный патоген» . Архивировано из оригинала 30 января 2020 года . Проверено 30 января 2020 г.
- ^ Перейти обратно: а б Стремлау М.Х., Андерсен К.Г., Фоларин О.А., Гроув Дж.Н., Одиа И., Эхиан П.Е. и др. (март 2015 г.). Рупрехт CE (ред.). «Обнаружение новых рабдовирусов в крови здоровых людей из Западной Африки» . PLOS Забытые тропические болезни . 9 (3): e0003631. дои : 10.1371/journal.pntd.0003631 . ПМЦ 4363514 . ПМИД 25781465 .
Дальнейшее чтение
[ редактировать ]- Роуз Дж.К., Уитт М.А. (2001). «Рабдовирусы: вирусы и их репликация». В Knipe DM, Howley PM (ред.). Вирусология Филда . Том. 1 (4-е изд.). Филадельфия: Липпинкотт Уильямс и Уилкинс. стр. 1221–44. ISBN 978-0781718325 .
- Вагнер Р.Р., изд. (1987). Рабдовирусы . Пленум Пресс. ISBN 978-0-306-42453-3 .
