Перевернутый повтор
Инвертированный повтор (или IR ) представляет собой одноцепочечную последовательность нуклеотидов , следовавшая ниже по течению из -за его обратного комплемента . [ 1 ] Промежуточная последовательность нуклеотидов между начальной последовательности и обратным комплементом может быть любой длиной, включая ноль. Например, 5 '--- ttacgnnnnnnnnnnnn CGTAA --- 3 ' -это перевернутая повторяющаяся последовательность. Когда промежуточная длина равен нулю, композитная последовательность представляет собой палиндромическую последовательность . [ 2 ]
Как перевернутые повтора, так и прямые повторения составляют типы нуклеотидных последовательностей , которые встречаются повторяющимися. Эти повторные последовательности ДНК часто варьируются от пары нуклеотидов до целого гена , в то время как близость повторяющихся последовательностей варьируется между широко рассеянными и простыми тандемными массивами . [ 3 ] Короткие тандемные повторные последовательности могут существовать, поскольку всего несколько копий в небольшой регионе на тысячи экземпляров, разбросанных по всему геному большинства эукариот . [ 4 ] Повторные последовательности с примерно 10–100 пар оснований известны как минисателлиты , в то время как более короткие повторные последовательности, имеющие в основном 2–4 пары оснований, известны как микросателлиты . [ 5 ] Наиболее распространенные повторения включают повторения динуклеотидов, которые имеют основы AC на одной пряди ДНК и GT на дополнительной цепи. [ 3 ] Некоторые элементы генома с уникальными последовательностями функционируют как экзоны , интроны и регуляторную ДНК. [ 6 ] Хотя наиболее знакомыми локусами повторяющихся последовательностей являются Centromere и Telomere , [ 6 ] Большая часть повторных последовательностей в геноме обнаружена среди некодирующей ДНК . [ 5 ]
Перевернутые повторения имеют ряд важных биологических функций. Они определяют границы в транспозонах и указывают области, способные к самообучению, спаривание оснований (области в пределах одной последовательности, которая может основать пару друг с другом). Эти свойства играют важную роль в нестабильности генома [ 7 ] и внести свой вклад не только в клеточную эволюцию и генетическое разнообразие [ 8 ] но также для мутации и болезней . [ 9 ] Чтобы подробно изучить эти эффекты, был разработан ряд программ и баз данных, чтобы помочь в обнаружении и аннотации инвертированных повторений в различных геномах.
Понимание перевернутых повторов
[ редактировать ]Пример перевернутого повторения
[ редактировать ]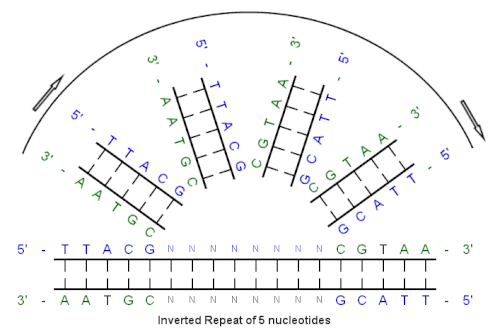
Начиная с этой начальной последовательности:
5'-ttacg-3 '
Дополнение, созданное базовым сопряжением, - это:
3'-Aatgc-5 '
Обратный дополнение:
5'-cgtaa-3 '
И инвертированная последовательность повторений составляет:
5 '--- ttacgnnnnnnnnnnnn Cgtaa --- 3 '
«Nnnnnn» представляет любое количество промежуточных нуклеотидов.
Против. прямой повтор
[ редактировать ]Прямой повтор возникает, когда последовательность повторяется с тем же рисунком вниз по течению. [ 1 ] Нет инверсии и нет обратного дополнения, связанного с прямым повторением. Нуклеотидная последовательность, написанная жирным шрифтом, означает повторную последовательность. Это может или не может иметь промежуточные нуклеотиды.
- 5´ ttacg nnnnnnnttacg 3´
- 3 ' aatgc nnnnnnaatgc 5'
Лингвистически, типичный прямой повтор сопоставим с рифмованным, как в «T ime on ad ime ».
Против. тандем повтор
[ редактировать ]Прямой повтор без промежуточных нуклеотидов между начальной последовательности и его нисходящей копией является тандемным повторением . Нуклеотидная последовательность, написанная жирным шрифтом, означает повторную последовательность.
- 5´ ttacg ttacg 3´
- 3 'aatgc aatgc 5'
Лингвистически, типичный тандемный повтор сопоставим с заиканием или намеренно повторяющимися словами, как в «пока».
Против. палиндром
[ редактировать ]Инвертированная последовательность повторений без промежуточных нуклеотидов между начальной последовательности и ее нижним обратным комплементом является палиндром . [ 1 ]
ПРИМЕР:
Шаг 1: Начните с перевернутого повторения: 5 'ttacgnnnnncgtaa 3'
Шаг 2: Снимите промежуточные нуклеотиды: 5 'ttacgcgtaa 3'
Эта полученная последовательность является палиндромной, потому что она является обратным комплемента. [ 1 ]
- 5 'ttacgcgtaa 3' тестовая последовательность (с шага 2 с удаленными промежуточными нуклеотидами)
- 3 'Aatgcgcatt 5' комплемент тестовой последовательности
- 5 'ttacgcgtaa 3' обратный комплемент. Это то же самое, что и тестовая последовательность выше, и, таким образом, это палиндром.
Биологические особенности и функциональность
[ редактировать ]Условия, которые способствуют синтезу
[ редактировать ]Разнообразные повтора по всему геному получены из транспонируемых элементов , которые теперь понимаются, чтобы «прыгать» на разные геномные местоположения, не передавая их первоначальные копии. [ 10 ] Последующее перекрытие одних и тех же последовательностей в течение многих поколений обеспечивает их множественность на протяжении всего генома. [ 10 ] Ограниченная рекомбинация последовательностей между двумя различными элементами последовательности, известными как консервативная сайт-специфическая рекомбинация (CSSR), приводит к инверсии сегмента ДНК, основанную на расположении последовательностей распознавания рекомбинации на ДНК доноров и ДНК реципиента. [ 10 ] Опять же, ориентация двух рекомбинирующих сайтов в молекуле донорской ДНК относительно асимметрии промежуточных последовательностей расщепления ДНК, известных как область кроссовера, является ключевой для образования либо перевернутых повторов, либо прямых повторений. [ 10 ] Таким образом, рекомбинация, возникающая в паре перевернутых участков, будет инвертировать последовательность ДНК между двумя сайтами. [ 10 ] Очень стабильные хромосомы наблюдались при сравнительно меньшем количестве инвертированных повторений, чем прямые повторения, что свидетельствует о взаимосвязи между стабильностью хромосом и количеством повторов. [ 11 ]
Регионы, где присутствие является обязательным
[ редактировать ]Терминальные перевернутые повторения наблюдались в ДНК различных эукариотических транспозонов, хотя их источник остается неизвестным. [ 12 ] Перевернутые повторения в основном обнаруживаются на происхождении репликации клеточного организма и органеллов, которые варьируются от фаговых плазмид, митохондрий и эукариотических вирусов до клеток млекопитающих. [ 13 ] Происхождение репликации фага G4 и других родственных фагов содержит сегмент почти 139 нуклеотидных оснований, которые включают три перевернутых повторения, которые необходимы для праймирования репликации. [ 13 ]
В геноме
[ редактировать ]В значительной степени части нуклеотидных повторов довольно часто наблюдаются в рамках редких комбинаций ДНК. [ 14 ] Три основных повторения, которые в основном обнаружены в конкретных ДНК-конструкциях, включают тесно точные перевернутые повторения гомопурин-гомопиримидина, которые в противном случае называются H palindromes, что является общим явлением в тройных спиральных конформациях H, которые могут содержать либо нуклеотидные триады TAT или CGC. Другие могут быть описаны такими же длинными перевернутыми повторами, имеющими тенденцию производить шпильки и крестообразные, и, наконец, прямые тандемные повторения, которые обычно существуют в структурах, описанных как проскальзывающая, крестообразная и левша Z-ДНК. [ 14 ]
Распространено в разных организмах
[ редактировать ]Прошлые исследования показывают, что повторения являются общей чертой эукариот в отличие от прокариот и археи . [ 14 ] Другие сообщения предполагают, что независимо от сравнительной нехватки повторяющихся элементов в прокариотических геномах, они, тем не менее, содержат сотни или даже тысячи больших повторов. [ 15 ] Текущий геномный анализ, по -видимому, предполагает существование большого избытка идеальных перевернутых повторений во многих прокариотических геномах по сравнению с эукариотическими геномами. [ 16 ]

Для количественной оценки и сравнения перевернутых повторений между несколькими видами, а именно на археи, см. [ 17 ]
Перевернутые повторения в псевдокнотах
[ редактировать ]Псевдокноты являются общими структурными мотивами, обнаруженными в РНК. Они образуются двумя вложенными петлями стеблей, так что стебель одной структуры образуется из петли другой. Есть несколько складных топологий среди псевдокнотов и большие различия в длине петли, что делает их структурно разнообразной группой. [ 18 ]
Перевернутые повторения являются ключевым компонентом псевдокнотов, как видно на иллюстрации естественного псевдокнота, обнаруженного в компоненте РНК теломеразы человека . [ 19 ] Четыре разных набора инвертированных повторений участвуют в этой структуре. Наборы 1 и 2 являются стеблем стебля-петли A и являются частью петли для петли STEM B. Аналогично, наборы 3 и 4 являются стеблем для петли STEM B и являются частью петли для петли STEM A.
Псевдокноты играют ряд различных ролей в биологии. Псевдокнот теломеразы на иллюстрации имеет решающее значение для активности этого фермента. [ 19 ] Рибозим . для дельта-вируса гепатита (HDV) складывается в структуру с двойной псевдоки и саморешивает его круговой геном, образуя РНК длины одного генома Псевдокноты также играют роль в запрограммированном рибосомном рамке, обнаруженном в некоторых вирусах и необходимых в репликации ретровирусов . [ 18 ]
В рибосвитчах
[ редактировать ]Перевернутые повторения играют важную роль в рибосвитчах , которые представляют собой регуляторные элементы РНК, которые контролируют экспрессию генов, которые продуцируют мРНК, из которых они являются. [ 10 ] Упрощенный пример рибосвитча флавина мононуклеотида (FMN) показан на иллюстрации. Этот рибосвит существует в транскрипте мРНК и имеет несколько структур петли стеблей вверх по течению от области кодирования . Тем не менее, на иллюстрации показаны только ключевые петли ствола, что было значительно упрощено, чтобы помочь показать роль перевернутых повторений. В этом рибосвитче есть несколько перевернутых повторений, как указано зеленым (желтый фон) и синий (оранжевый фон).
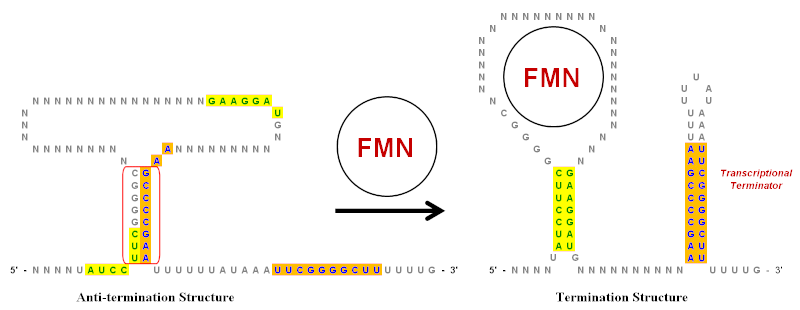
В отсутствие FMN структура анти-концов является предпочтительной конформацией для транскрипта мРНК. Он создан путем базовой пары инвертированной повторной области, окруженной красным. Когда FMN присутствует, он может связываться с петлей и предотвратить образование противоположной структуры. Это позволяет двум различным наборам инвертированных повторений в базовую паре и образует структуру завершения. [ 20 ] Стволовая петля на 3 'END является транскрипционным терминатором, потому что последовательность сразу же, следующая за ней, представляет собой серию урацилов (U). Если эта петля ствола образуется (из-за присутствия FMN), поскольку растущая РНК-цепа возникает из комплекса РНК-полимеразы , она создаст достаточное структурное напряжение, чтобы привести к диссоциации РНК-цепи и, таким образом, прекращает транскрипцию. Диссоциация происходит легко, потому что основание между U в РНК и А в границе шаблона является самым слабым из всех базовых пар. [ 10 ] Таким образом, при более высоких уровнях концентрации FMN снижает свою собственную транскрипцию, увеличивая образование структуры прекращения.
Мутации и болезнь
[ редактировать ]Перевернутые повторения часто описываются как «горячие точки» эукариотической и прокариотической нестабильности генома. [ 7 ] Считается, что длинные перевернутые повторения значительно влияют на стабильность генома различных организмов. [ 21 ] Это иллюстрируется в E. coli , где геномные последовательности с длинными перевернутыми повторениями редко воспроизводятся, но скорее удаляются с помощью быстроты. [ 21 ] Опять же, длинные инвертированные повторения, наблюдаемые у дрожжей, значительно предпочитают рекомбинацию в тех же и прилегающих хромосомах, что приводит к одинаково очень высокой скорости удаления. [ 21 ] Наконец, очень высокая частота делеции и рекомбинации также наблюдалась в хромосомах млекопитающих с перевернутыми повторениями. [ 21 ] Сообщаемые различия в стабильности геномов взаимосвязанных организмов всегда являются признаком неравенства в перевернутых повторах. [ 11 ] Нестабильность возникает в результате тенденции инвертированных повторений складываться в структуры ДНК, подобные парикмахеру или крестообразным,. Эти специальные структуры могут препятствовать или запутать репликацию ДНК и другие геномные активности. [ 7 ] Таким образом, инвертированные повторения приводят к специальным конфигурациям как в РНК , так и в ДНК , которые в конечном итоге могут вызвать мутации и заболевания . [ 9 ]

Иллюстрация показывает перевернутое повторение, перенесенное крестообразное экструзию. ДНК в области инвертированного повторения раскручивается, а затем рекомбинирует, образуя четырехсторонний соединение с двумя стебля структурами . Структура крестообраза возникает из-за того, что инвертированные последовательности повторений самостоятельно самостоятельно на их собственную прядь. [ 22 ]
Экструдированные крестообразные могут привести к мутациям сдвига кадров , когда последовательность ДНК имеет инвертированные повторения в форме палиндрома в сочетании с областями прямых повторов с обеих сторон. Во время транскрипции проскальзывание и частичная диссоциация полимеразы из шаблонной цепи могут привести к мутациям удаления и вставки . [ 9 ] Удаление возникает, когда часть цепочки из пустого шаблона образует петли стебля, которая «пропущена» транскрипционной машиной. Вставка происходит, когда стволовая петля образуется в диссоциированной части зарождающейся (вновь синтезированной) цепи, вызывая часть нити шаблона, которая должна быть транскрибирована дважды. [ 9 ]
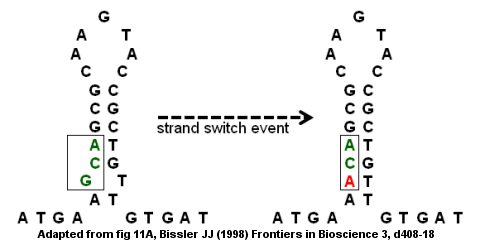
Дефицит антитробина из точечной мутации
[ редактировать ]Несовершенные перевернутые повторения могут привести к мутациям через интратрастр и переключение междрандов. [ 9 ] Кодирующая область гена антитробина III является примером несовершенного перевернутого повторения , как показано на рисунке справа. Структура ствола образуется с ударом внизу, потому что G и T не соединяются. Событие переключателя пряди может привести к тому, что G (в BUMP) будет заменен A, который удаляет «несовершенство» в перевернутом повторении и обеспечивает более сильную структуру ствола. Однако замена также создает точечную мутацию , преобразующую кодон GCA в ACA. Если за событием переключения пряди следует второй раунд репликации ДНК , мутация может быть зафиксирована в геноме и привести к заболеванию. В частности, миссенс -мутация приведет к дефектному гену и дефициту антитробина, что может привести к развитию венозной тромбоэмболии (сгустки крови в вене). [ 9 ]

Остеогенез несовершенного от мутации сдвига кадрика
[ редактировать ]Мутации в гене коллагена заболевания могут привести к несовершенству остеогенеза , который характеризуется хрупкими костями. [ 9 ] На рисунке стволовая петля, образованная из несовершенного перевернутого повторения, мутируется с помощью нуклеотидной вставки тимина (T) в результате меж- или внутри- или внутрипроизвольного переключателя. Добавление T создает «матч» базовой пары с аденином (а), который ранее был «ударом» на левой стороне стебля. Хотя это дополнение укрепляет ствол и совершенствует инвертированный повтор, оно также создает мутацию сдвига в нуклеотидной последовательности, которая изменяет рамку считывания и приведет к неправильной экспрессии гена. [ 9 ]
Программы и базы данных
[ редактировать ]Следующий список содержит информацию и внешние ссылки на различные программы и базы данных для перевернутых повторений:
- Бес-дБ база данных для интегрированных аннотаций и анализа мотивов формирования ДНК не B. [ 23 ] Эта база данных предоставляется Центром продвинутых биомедицинских вычислений (ABCC) в тогдашней Национальной лаборатории Фридриха по исследованиям рака (FNLCR). Он охватывает конформации A-ДНК и Z-ДНК, иначе известные как «не-B DNAS», потому что они не являются более распространенной формой B-ДНК правой ручной двойной спирали Уотсона-Крика . Эти «не-B DNA» включают Z-ДНК левшей, крестообразные , триплекс , тетраплекс и структуры шпильки . [ 23 ] Поиски могут быть выполнены на различных «повторных типах» (включая перевернутые повторения) и на нескольких видах.
- Инвертированные повторные базы данных 2020-09-01 в архивировали Boston University. Эта база данных представляет собой веб -приложение, которое позволяет запросить и анализ повторений, проводимых в проекте общедоступной базы данных. Ученые также могут проанализировать свои собственные последовательности с помощью алгоритма инвертированных повторений. [ 24 ]
- P-MITE: база данных Plant Mite -эта база данных для миниатюрных перевернутых переосмысленных транспозируемых элементов (MITE) содержит последовательности из растений геномов . Последовательности могут быть поиском или загружены из базы данных. [ 25 ]
- Emboss -это «Европейская молекулярная биология открытая программная плата», которая работает в операционных системах UNIX и UNIX . [ 26 ] Документация и исходные файлы программы доступны на веб -сайте Emboss . Приложения, специально связанные с перевернутыми повторами, перечислены ниже:
- Emboss Einverted : находит инвертированные повторения в нуклеотидных последовательностях . Пороговые значения могут быть установлены для ограничения объема поиска. [ 26 ]
- Embess Palindrome : находит палиндромы, такие как области петли стеблей в нуклеотидных последовательностях. Программа найдет последовательности, которые включают разделы несоответствия и пробелы, которые могут соответствовать выпуклостям в петле STEM. [ 26 ]
Ссылки
[ редактировать ]- ^ Jump up to: а беременный в дюймовый USSERY, David W.; Вассенаар, Труди; Борини, Стефано (2008-12-22). «Частоты слов, повторения и связанные с повторными структурами в бактериальных геномах». Вычисление для сравнительной микробной геномики: биоинформатика для микробиологов . Вычислительная биология. Тол. 8 (1 изд.). Спрингер. С. 133–144. ISBN 978-1-84800-254-8 .
- ^ Вы, конггируя; Джи, Гули; Лян, Чун (2014). «Detectir: новая программа для обнаружения идеальных и несовершенных перевернутых повторений с использованием комплексных чисел и вычисления вектора» . Plos один . 9 (11): E113349. Bibcode : 2014ploso ... 9k3349y . doi : 10.1371/journal.pone.0113349 . PMC 4237412 . PMID 25409465 .
- ^ Jump up to: а беременный Ричардс, Гр; Ричардс, RI (25 апреля 1995 г.). «Простая тандемная ДНК повторяется и генетическое заболевание человека» . Труды Национальной академии наук Соединенных Штатов Америки . 92 (9): 3636–41. Bibcode : 1995pnas ... 92.3636S . doi : 10.1073/pnas.92.9.3636 . PMC 42017 . PMID 7731957 .
- ^ Ван Белкум, а; Scherer, S; Ван Альфен, L; Verbrugh, H (июнь 1998 г.). «Короткая последовательность ДНК повторяется в прокариотических геномах» . Микробиология и молекулярная биология обзоры . 62 (2): 275–93. doi : 10.1128/mmbr.62.2.275-293.1998 . PMC 98915 . PMID 9618442 .
- ^ Jump up to: а беременный Рамель, C (июнь 1997 г.). «Мини- и микросателлиты» . Перспективы здоровья окружающей среды . 105 (Suppl 4): 781–9. doi : 10.2307/3433284 . JSTOR 3433284 . PMC 1470042 . PMID 9255562 .
- ^ Jump up to: а беременный Eichler, EE (август 1998 г.). «Маскирующие повторения: паралогические ловушки человеческого генома» . Исследование генома . 8 (8): 758–62. doi : 10.1101/gr.8.8.758 . PMID 9724321 .
- ^ Jump up to: а беременный в Миркин, я; Нараянан, V; Лобачев, KS; Миркин, С.М. (22 июля 2008 г.). «Репликация остановка при нестабильных перевернутых повторах: взаимодействие между шпильками ДНК и стабилизирующими вилками белков» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (29): 9936–41. Bibcode : 2008pnas..105.9936v . doi : 10.1073/pnas.0804510105 . PMC 2481305 . PMID 18632578 .
- ^ Lin, Ct; Лин, ч; Lyu, Yl; Whang-Peng, J (1 сентября 2001 г.). «Перевернутые повторения как генетические элементы для стимулирования инвертированного дублирования ДНК: последствия для амплификации генов» . Исследование нуклеиновых кислот . 29 (17): 3529–38. doi : 10.1093/nar/29.17.3529 . PMC 55881 . PMID 11522822 .
- ^ Jump up to: а беременный в дюймовый и фон глин час Бисслер, JJ (27 марта 1998 г.). «ДНК -инвертированные повтора и болезнь человека» (PDF) . Границы в биологии . 3 (4): D408–18. doi : 10.2741/a284 . PMID 9516381 . S2CID 12982 . Архивировано из оригинала (PDF) 3 марта 2019 года.
- ^ Jump up to: а беременный в дюймовый и фон глин Школа, Джеймс Д. Уотсон, Лаборатория Колд -Спринг -Харбор, Таня А. Бейкер, Массачусетский технологический институт, Стивен П. Белл, Массачусетский технологический институт, Александр Ганн, Лаборатория Колд -Спринг -Харбор, Майкл Левин, Калифорнийский университет, Беркли, Ричард Лосик, Гарвардский университет; со Стивеном С. Харрисоном, Гарвард Медицинский (2014). Молекулярная биология гена (седьмое изд.). Бостон: издательская компания Бенджамин-Каммингс. ISBN 9780321762436 .
{{cite book}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а беременный Ахаз, G; Coissac, E; Netter, p; Роча, EP (август 2003 г.). «Ассоциации между перевернутыми повторами и структурной эволюцией бактериальных геномов» . Генетика . 164 (4): 1279–89. doi : 10.1093/Genetics/164.4.1279 . PMC 1462642 . PMID 12930739 .
- ^ Чжан, HH; Сюй, он; Шен, да; Хан, MJ; Zhang, Z (январь 2013 г.). «Происхождение и эволюция шести миниатюрных перевернутых переосмысленных транспонируемых элементов в Bombyx Mori и Rhodnius prolixus» . Биология и эволюция генома . 5 (11): 2020–31. doi : 10.1093/gbe/evt153 . PMC 3845634 . PMID 24115603 .
- ^ Jump up to: а беременный Пирсон, CE; Зорбас, ч; Цена, ГБ; Zannis-Hadjopoulos, M (октябрь 1996 г.). «Перевернутые повторения, петли стеблей и крестообразные: значение для инициации репликации ДНК». Журнал сотовой биохимии . 63 (1): 1–22. doi : 10.1002/(SICI) 1097-4644 (199610) 63: 1 <1 :: AID-JCB1> 3.0.CO; 2-3 . PMID 8891900 . S2CID 22204780 .
- ^ Jump up to: а беременный в Heringa, J (июнь 1998 г.). «Обнаружение внутренних повторений: насколько они распространены?». Современное мнение в структурной биологии . 8 (3): 338–45. doi : 10.1016/s0959-440x (98) 80068-7 . PMID 9666330 .
- ^ Треанген, TJ; Авраам, Ал; Touckon, M; Роча, EP (май 2009 г.). «Бытие, эффекты и судьбы повторений в прокариотических геномах» (PDF) . Обзоры микробиологии FEMS . 33 (3): 539–71. doi : 10.1111/j.1574-6976.2009.00169.x . PMID 19396957 .
- ^ Ladoukakis, ed; Эйр-Уокер, А (сентябрь 2008 г.). «Избыток небольших перевернутых повторов в прокариотах» (PDF) . Журнал молекулярной эволюции . 67 (3): 291–300. Bibcode : 2008jmole..67..291l . Citeseerx 10.1.1.578.7466 . doi : 10.1007/s00239-008-9151-z . PMID 18696026 . S2CID 29953202 .
- ^ Хоссейни, м; Пратас, D; Pinho, AJ (2017). «О роли инвертированных повторов в сходстве последовательности ДНК». 11 -я Международная конференция по практическим применению вычислительной биологии и биоинформатики . Достижения в интеллектуальных системах и вычислениях. Тол. 616. Springer. С. 228–236. doi : 10.1007/978-3-319-60816-7_28 . ISBN 978-3-319-60815-0 .
- ^ Jump up to: а беременный Основной, DW; Мясник, SE (июнь 2005 г.). «Псевдокноты: РНК -структуры с разнообразными функциями» . PLOS Биология . 3 (6): E213. doi : 10.1371/journal.pbio.0030213 . PMC 1149493 . PMID 15941360 .

- ^ Jump up to: а беременный Чен, JL; Greider, CW (7 июня 2005 г.). «Функциональный анализ структуры псевдокнот в РНК теломеразы человека» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (23): 8080–5, обсуждение 8077–9. Bibcode : 2005pnas..102.8080c . doi : 10.1073/pnas.0502259102 . PMC 1149427 . PMID 15849264 .
- ^ Винклер, WC; Cohen-Chalamish, S; Breaker, RR (10 декабря 2002 г.). «Структура мРНК, которая контролирует экспрессию генов путем связывания FMN» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (25): 15908–13. Bibcode : 2002pnas ... 9915908W . doi : 10.1073/pnas.212628899 . PMC 138538 . PMID 12456892 .
- ^ Jump up to: а беременный в дюймовый Stormo, g; Чанг, Кентукки; Варли, К; Stormo, GD (28 февраля 2007 г.). Холл, Нил (ред.). «Доказательства активного поддержания инвертированных повторных структур, идентифицированных с помощью сравнительного геномного подхода» . Plos один . 2 (2): E262. Bibcode : 2007ploso ... 2..262Z . doi : 10.1371/journal.pone.0000262 . PMC 1803023 . PMID 17327921 .
- ^ Рамредди, т; Sachidanandam, R; Стрик, ТР (май 2011 г.). «В реальном времени обнаружение крестообразной экструзии с помощью одномолекулярной наноманипуляции ДНК» . Исследование нуклеиновых кислот . 39 (10): 4275–83. doi : 10.1093/nar/gkr008 . PMC 3105387 . PMID 21266478 .
- ^ Jump up to: а беременный Cer, rz; Donohue, de; Мудунури, США; Темиз, На; Потеря, Ма; Starner, NJ; Halusa, GN; Вольфвский, n; Yi, m; Люк, Бт; Bacolla, A; Коллинз, младший; Стивенс, RM (январь 2013 г.). «Не-B DB v2.0: база данных прогнозируемых не-образных ДНК-мотивов не B и связанных с ним инструментов» . Исследование нуклеиновых кислот . 41 (проблема базы данных): D94 - D100. doi : 10.1093/nar/gks955 . PMC 3531222 . PMID 23125372 .
- ^ Гелфанд, y; Родригес, а; Бенсон, G (январь 2007 г.). «TRDB-тандем повторяет базу данных» . Исследование нуклеиновых кислот . 35 (проблема базы данных): D80–7. doi : 10.1093/nar/gkl1013 . PMC 1781109 . PMID 17175540 .
- ^ Чен, J; Ху, Q; Чжан, y; Lu, c; Куанг, ч (29 октября 2013 г.). «P-MITE: база данных для миниатюрных перевернутых перевернутых элементов» . Исследование нуклеиновых кислот . 42 (1): D1176–81. doi : 10.1093/nar/gkt1000 . PMC 3964958 . PMID 24174541 .
- ^ Jump up to: а беременный в Рис, р; Лонгден, я; Bleasby, A (июнь 2000 г.). «Emboss: Европейская молекулярная биология открытая программная набор» . Тенденции в генетике . 16 (6): 276–7. doi : 10.1016/s0168-9525 (00) 02024-2 . PMID 10827456 .
Внешние ссылки
[ редактировать ]- Перевернутый+повторный+последовательность библиотеки США в Национальной медицинской библиотеке Медицинской (Mesh)