Z-ДНК
Z-ДНК — одна из многих возможных двойных спиральных структур ДНК . Это левосторонняя двойная спиральная структура, в которой спираль зигзагообразно закручивается влево, а не вправо, как в более распространенной форме B-ДНК . Z-ДНК считается одной из трех биологически активных двухспиральных структур наряду с A-ДНК и B-ДНК .
История
[ редактировать ]Леворукая ДНК была впервые предложена Уэллсом и его коллегами как структура повторяющегося полимера инозин цитозин - Робертом . [1] Они наблюдали «обратный» спектр кругового дихроизма для таких ДНК и неправильно интерпретировали это как означающее, что нити закручиваются друг вокруг друга левосторонним образом. На связь между Z-ДНК и более известной B-ДНК указали работы Пола и Йовина. [2] который показал, что ультрафиолетовый круговой дихроизм поли(dG-dC) почти инвертируется в 4 М растворе хлорида натрия и что структура поли d(I-C)·poly d(I-C) на самом деле является правосторонней. Конформация D-ДНК. Подозрение, что это было результатом превращения B-ДНК в Z-ДНК, было подтверждено исследованием спектров комбинационного рассеяния этих растворов и кристаллов Z-ДНК. [3] Впоследствии была опубликована кристаллическая структура «Z-ДНК», которая оказалась первой монокристаллической рентгеновской структурой фрагмента ДНК (самомлементарного ДНК-гексамера d(CG) 3 ). Она была решена как левая двойная спираль с двумя антипараллельными цепями, которые удерживались вместе парами оснований Уотсона-Крика (см. Рентгеновскую кристаллографию ). Она была решена Эндрю Х. Дж. Вангом , Александром Ричем и его коллегами в 1979 году в Массачусетском технологическом институте . [4] Кристаллизация соединения B- и Z-ДНК в 2005 г. [5] обеспечили лучшее понимание потенциальной роли Z-ДНК в клетках. Всякий раз, когда формируется сегмент Z-ДНК, на его двух концах должны быть соединения B-Z, соединяющие его с B-формой ДНК, обнаруженной в остальной части генома .
В 2007 году РНК- версия Z-ДНК, Z-РНК , была описана как трансформированная версия двойной спирали А-РНК в левостороннюю спираль. [6] Однако переход от А-РНК к Z-РНК был описан уже в 1984 году. [7]
Структура
[ редактировать ]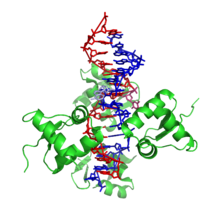
Z-ДНК сильно отличается от правосторонних форм. Фактически, Z-ДНК часто сравнивают с B-ДНК, чтобы проиллюстрировать основные различия. Спираль Z-ДНК левая и имеет структуру, повторяющую каждую вторую пару оснований. Большие и малые бороздки, в отличие от А- и В-ДНК, мало отличаются по ширине. Формирование этой структуры в целом неблагоприятно, хотя определенные условия могут этому способствовать; такие как чередующаяся пурин - пиримидиновая последовательность (особенно поли(dGC) 2 ), отрицательная сверхспирализация ДНК или высокое содержание соли и некоторые катионы (все при физиологической температуре, 37 ° C и pH 7,3–7,4). Z-ДНК может образовывать соединение с B-ДНК (называемое «соединительной коробкой B-Z») в структуре, которая включает экструзию пары оснований. [8] Конформацию Z-ДНК трудно изучать, поскольку она не существует как стабильная особенность двойной спирали. Напротив, это временная структура, которая иногда возникает в результате биологической активности, а затем быстро исчезает. [9]
Прогнозирование структуры Z-ДНК
[ редактировать ]Можно предсказать вероятность того, что последовательность ДНК образует структуру Z-ДНК. Алгоритм прогнозирования склонности ДНК к переходу из B-формы в Z-форму, ZHunt , был написан П. Шинг Хо в 1984 году в Массачусетском технологическом институте . [10] Этот алгоритм позже был разработан Трейси Кэмп , П. Кристофом Чампом , Сандором Морисом и Джеффри М. Варгасоном для полногеномного картирования Z-ДНК (с Хо в качестве главного исследователя). [11]
Путь образования Z-ДНК из B-ДНК
[ редактировать ]С момента открытия и кристаллизации Z-ДНК в 1979 году эта конфигурация поставила ученых в тупик относительно пути и механизма перехода от конфигурации B-ДНК к конфигурации Z-ДНК. [12] Конформационное изменение структуры B-ДНК на структуру Z-ДНК было неизвестно на атомном уровне, но в 2010 году компьютерное моделирование, проведенное Lee et al. смогли вычислительно определить, что ступенчатое распространение перехода B-Z обеспечит более низкий энергетический барьер, чем ранее предполагаемый согласованный механизм. [13] Поскольку это было доказано вычислительно, этот путь все равно необходимо будет проверить экспериментально в лаборатории для дальнейшего подтверждения и достоверности, в чем Lee et al. В своей журнальной статье, в частности, говорится: «Текущий [вычислительный] результат может быть проверен с одиночной молекулой FRET (smFRET)». в будущем с помощью экспериментов [13] В 2018 году путь от B-ДНК к Z-ДНК был экспериментально доказан с помощью анализов smFRET. [14] Это было выполнено путем измерения значений интенсивности между донорными и акцепторными флуоресцентными красителями, также известными как флуорофоры , по отношению друг к другу, когда они обмениваются электронами, будучи прикрепленными к молекуле ДНК. [15] [16] Расстояния между флуорофорами можно использовать для количественного расчета изменений близости красителей и конформационных изменений ДНК. Высокоаффинный связывающий белок Z-ДНК , hZαADAR1, [17] использовали в различных концентрациях для индукции трансформации B-ДНК в Z-ДНК. [14] Анализ smFRET выявил переходное состояние B*, которое сформировалось в результате связывания hZαADAR1, накопленного на структуре B-ДНК, и стабилизировало ее. [14] Этот шаг происходит, чтобы избежать высокой энергии соединения, при которой структура B-ДНК может претерпевать конформационные изменения по сравнению со структурой Z-ДНК без серьезного разрушительного изменения энергии. Этот результат совпадает с результатами вычислений Lee et al. доказывая, что этот механизм является поэтапным и его цель состоит в том, что он обеспечивает более низкий энергетический барьер для конформационного изменения от конфигурации B-ДНК к конфигурации Z-ДНК. [13] Вопреки предыдущему представлению, связывающие белки фактически не стабилизируют конформацию Z-ДНК после ее формирования, а вместо этого фактически способствуют образованию Z-ДНК непосредственно из конформации B*, которая образуется B-ДНК. структура связывается белками с высоким сродством. [14]
Биологическое значение
[ редактировать ]Биологическая роль Z-ДНК в регуляции ответов интерферона I типа была подтверждена в исследованиях трех хорошо изученных редких менделевских заболеваний: симметричного наследственного дисхроматоза (OMIM: 127400), синдрома Айкарди-Гутьера (OMIM: 615010) и двустороннего полосатого тела. Некроз/Дистония. Семьи с гаплоидным транскриптомом ADAR позволили сопоставить варианты Zα непосредственно с заболеванием, показав, что генетическая информация кодируется в ДНК как по форме, так и последовательности. [18] Роль в регуляции реакции интерферона I типа при раке также подтверждается данными о том, что выживание 40% опухолей зависело от фермента ADAR. [19]
В предыдущих исследованиях Z-ДНК была связана как с болезнью Альцгеймера , так и с системной красной волчанкой . Чтобы продемонстрировать это, было проведено исследование ДНК, обнаруженной в гиппокампе головного мозга нормального человека, умеренно пораженного болезнью Альцгеймера и тяжело пораженного болезнью Альцгеймера. Благодаря использованию кругового дихроизма это исследование показало наличие Z-ДНК в ДНК тяжело пострадавших. [20] В этом исследовании также было обнаружено, что основные части умеренно пораженной ДНК находились в промежуточной конформации BZ. Это важно, поскольку на основе этих результатов был сделан вывод, что переход от B-ДНК к Z-ДНК зависит от прогрессирования болезни Альцгеймера. [20] Кроме того, Z-ДНК связана с системной красной волчанкой (СКВ) благодаря наличию естественных антител. Значительные количества антител против Z-ДНК были обнаружены у больных СКВ и отсутствовали при других ревматических заболеваниях. [21] Существует два типа этих антител. С помощью радиоиммуноанализа было обнаружено, что один взаимодействует с основаниями, экспонированными на поверхности Z-ДНК и денатурированной ДНК, а другой взаимодействует исключительно с зигзагообразным остовом только Z-ДНК. Подобно тому, как это происходит при болезни Альцгеймера, антитела варьируются в зависимости от стадии заболевания: максимальное количество антител наблюдается на наиболее активных стадиях СКВ.
Z-ДНК в транскрипции
[ редактировать ]Обычно считается, что Z-ДНК обеспечивает торсионного напряжения облегчение во время транскрипции , и это связано с отрицательной суперспирализацией . [5] [22] Однако, хотя сверхспирализация связана как с транскрипцией, так и с репликацией ДНК, образование Z-ДНК в первую очередь связано со скоростью транскрипции . [23]
Исследование 22-й хромосомы человека показало корреляцию между областями формирования Z-ДНК и областями промотора ядерного фактора I. Это предполагает, что транскрипция в некоторых генах человека может регулироваться образованием Z-ДНК и активацией ядерного фактора I. [11]
Было показано, что последовательности Z-ДНК, расположенные выше промоторных областей, стимулируют транскрипцию. Наибольшее увеличение активности наблюдается, когда последовательность Z-ДНК располагается на три витка спирали после последовательности промотора . Кроме того, используя метод сшивания микрококковой нуклеазой, [24] Z-ДНК вряд ли образует нуклеосомы , которые часто располагаются до и/или после последовательности, образующей Z-ДНК. Предполагается, что из-за этого свойства Z-ДНК кодирует границу позиционирования нуклеосом. Поскольку размещение нуклеосом влияет на связывание факторов транскрипции , считается, что Z-ДНК регулирует скорость транскрипции. [24]
возникшая на основе пути РНК-полимеразы Было показано, что Z-ДНК, образовавшаяся посредством активной транскрипции, посредством отрицательной суперспирализации, увеличивает генетическую нестабильность, создавая склонность к мутагенезу вблизи промоторов. [25] Исследование Escherichia coli генов показало, что делеции спонтанно происходят в областях плазмиды , содержащих последовательности, образующие Z-ДНК. [26] Было обнаружено, что в клетках млекопитающих присутствие таких последовательностей приводит к крупным делециям геномных фрагментов из-за хромосомных двухцепочечных разрывов . Обе эти генетические модификации связаны с транслокациями генов , обнаруженными при таких видах рака, как лейкемия и лимфома , поскольку области разрыва в опухолевых клетках нанесены вокруг последовательностей, образующих Z-ДНК. [25] Однако более мелкие делеции в бактериальных плазмидах связаны с проскальзыванием репликации , тогда как более крупные делеции, связанные с клетками млекопитающих, вызваны негомологичной репарацией соединения концов, которая, как известно, подвержена ошибкам. [25] [26]
Токсическое действие бромистого этидия (EtBr) на трипаносомы обусловлено сдвигом их кинетопластидной ДНК в Z-форму. Сдвиг вызван интеркаляцией EtBr и последующим разрыхлением структуры ДНК, что приводит к раскручиванию ДНК, переходу в Z-форму и ингибированию репликации ДНК. [27]
Открытие домена Zα
[ редактировать ]Первый домен, связывающий Z-ДНК с высоким сродством, был обнаружен в ADAR1 с использованием подхода, разработанного Аланом Гербертом. [28] [29] Кристаллографические и ЯМР- исследования подтвердили биохимические данные о том, что этот домен связывает Z-ДНК неспецифичным для последовательности образом. [30] [31] [32] Родственные домены были идентифицированы в ряде других белков посредством гомологии последовательностей . [29] Идентификация домена Zα послужила инструментом для других кристаллографических исследований, которые привели к характеристике Z-РНК и соединения B-Z. Биологические исследования показали, что Z-ДНК-связывающий домен ADAR1 может локализовать этот фермент, который модифицирует последовательность вновь образованной РНК в местах активной транскрипции. [33] [34] Роль Zα, Z-ДНК и Z-РНК в защите генома от вторжения ретро-элементов Alu у человека превратилась в механизм регуляции врожденных иммунных ответов на дцРНК. Мутации в Zα являются причиной интерферопатий человека, таких как синдром Менделя Айкарди-Гутьера. [35] [18] Кроме того, показано, что домены Zα локализуются в стрессовых гранулах из-за их врожденной способности связывать нуклеиновую кислоту. Более того, разные домены Zα по-разному связываются с Z-конформацией нуклеиновой кислоты, обеспечивая важные возможности для специфического нацеливания при разработке лекарств.
Последствия связывания Z-ДНК с белком E3L коровьей оспы
[ редактировать ]Поскольку Z-ДНК была исследована более тщательно, было обнаружено, что структура Z-ДНК может связываться с белками, связывающими Z-ДНК, посредством лондонной дисперсии и водородных связей . [36] Одним из примеров белка, связывающего Z-ДНК, является белок E3L коровьей оспы , который является продуктом гена E3L и имитирует белок млекопитающих, который связывает Z-ДНК. [37] [38] Белок E3L не только имеет сродство к Z-ДНК, но также было обнаружено, что он играет роль в уровне тяжести вирулентности у мышей, вызванной вирусом коровьей оспы, разновидностью поксвируса . Двумя критически важными компонентами белка E3L, определяющими вирулентность, являются N-конец и C-конец . N-конец состоит из последовательности, аналогичной последовательности домена Zα, также называемой доменом z-альфа аденозиндезаминазы , тогда как C-конец состоит из двухцепочечного мотива связывания РНК. [37] Благодаря исследованиям, проведенным Кимом, Ю. и др. в Массачусетском технологическом институте было показано, что замена N-конца белка E3L на последовательность домена Zα, содержащую 14 остатков связывания Z-ДНК, аналогичных E3L, практически не влияет на патогенность вируса у мышей. [37] Напротив, Ким Ю. и др. также обнаружили, что удаление всех 83 остатков N-конца E3L приводило к снижению вирулентности. Это подтверждает их утверждение о том, что N-конец, содержащий остатки связывания Z-ДНК, необходим для вирулентности. [37] В целом, эти результаты показывают, что сходные остатки связывания Z-ДНК на N-конце белка E3L и домене Zα являются наиболее важными структурными факторами, определяющими вирулентность, вызываемую вирусом осповакцины, в то время как аминокислотные остатки, не участвующие в Z-ДНК, привязка практически не имеет эффекта. Будущее значение этих результатов включает снижение связывания E3L с Z-ДНК в вакцинах, содержащих вирус коровьей оспы, чтобы можно было свести к минимуму негативные реакции на вирус у людей. [37]
Кроме того, Александр Рич и Джин-А Квон обнаружили, что E3L действует как трансактиватор человеческих генов IL-6, NF-AT и p53. Их результаты показывают, что клетки HeLa, содержащие E3L, имели повышенную экспрессию человеческих генов IL-6, NF-AT и p53, а точечные мутации или делеции определенных аминокислотных остатков, связывающих Z-ДНК, снижали эту экспрессию. [36] В частности, было обнаружено, что мутации в Tyr 48 и Pro 63 снижают трансактивацию ранее упомянутых генов в результате потери водородных связей и лондонских дисперсионных сил между E3L и Z-ДНК. [36] В целом, эти результаты показывают, что уменьшение связей и взаимодействий между Z-ДНК и белками, связывающими Z-ДНК, снижает как вирулентность, так и экспрессию генов, тем самым показывая важность наличия связей между Z-ДНК и связывающим белком E3L.
Сравнительная геометрия некоторых форм ДНК
[ редактировать ]

| А-форма | B-форма | Z-форма | |
|---|---|---|---|
| Чувство спирали | правша | правша | левша |
| Повторяющийся блок | 1 б.п. | 1 б.п. | 2 б.п. |
| Ротация/б.п. | 32.7° | 34.3° | 30° |
| б.п./оборот | 11 | 10 | 12 |
| Наклон б.п. к оси | +19° | −1.2° | −9° |
| Подъем/б.п. вдоль оси | 2,3 Å (0,23 нм) | 3,32 Å (0,332 нм) | 3,8 Å (0,38 нм) |
| Шаг/поворот спирали | 28,2 Å (2,82 нм) | 33,2 Å (3,32 нм) | 45,6 Å (4,56 нм) |
| Средний поворот пропеллера | +18° | +16° | 0° |
| Гликозильный угол | анти | анти | С: анти , G: syn |
| Сахарная складка | C3'- эндо | C2'- эндо | C: C2'- эндо , G: C3'- эндо |
| Диаметр | 23 Å (2,3 нм) | 20 Å (2,0 нм) | 18 Å (1,8 нм) |
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Мицуи, Ю.; Лэнгридж, Р.; Шортл, Бельгия; Кантор, ЧР; Грант, RC; Кодама, М.; Уэллс, Р.Д. (1970). «Физические и ферментативные исследования поли d (I – C) · поли d (I – C), необычной двойной спиральной ДНК». Природа . 228 (5277): 1166–1169. Бибкод : 1970Natur.228.1166M . дои : 10.1038/2281166a0 . ПМИД 4321098 . S2CID 4248932 .
- ^ Пол, FM; Джовин, ТМ (1972). «Совместное конформационное изменение синтетической ДНК, индуцированное солью: равновесные и кинетические исследования с поли(dG-dC)». Журнал молекулярной биологии . 67 (3): 375–396. дои : 10.1016/0022-2836(72)90457-3 . ПМИД 5045303 .
- ^ Таманн, Ти Джей; Лорд, RC; Ван, АХ; Рич, А. (1981). «Высокосолевая форма поли(dG–dC) · поли(dG–dC) представляет собой левостороннюю Z-ДНК: спектры комбинационного рассеяния света кристаллов и растворов» . Исследования нуклеиновых кислот . 9 (20): 5443–5457. дои : 10.1093/нар/9.20.5443 . ПМЦ 327531 . ПМИД 7301594 .
- ^ Ван, АХ; Куигли, Дж.Дж.; Колпак, Ф.Дж.; Кроуфорд, Дж. Л.; ван Бум, Дж. Х.; ван дер Марель, Г.; Рич, А. (1979). «Молекулярная структура левостороннего фрагмента двойной спирали ДНК при атомном разрешении». Природа . 282 (5740): 680–686. Бибкод : 1979Natur.282..680W . дои : 10.1038/282680a0 . ПМИД 514347 . S2CID 4337955 .
- ^ Перейти обратно: а б Ха, СК; Ловенхаупт, К.; Рич, А.; Ким, Ю.Г.; Ким, К.К. (2005). «Кристаллическая структура соединения между B-ДНК и Z-ДНК обнаруживает два экструдированных основания». Природа . 437 (7062): 1183–1186. Бибкод : 2005Natur.437.1183H . дои : 10.1038/nature04088 . ПМИД 16237447 . S2CID 2539819 .
- ^ Плачидо, Д.; Браун, бакалавр II; Ловенхаупт, К.; Рич, А.; Афанасиадис, А. (2007). «Левая двойная спираль РНК, связанная доменом Zalpha фермента ADAR1, редактирующего РНК» . Структура . 15 (4): 395–404. дои : 10.1016/j.str.2007.03.001 . ПМК 2082211 . ПМИД 17437712 .
- ^ Холл, К.; Круз, П.; Тиноко, И. младший; Джовин, ТМ; ван де Санде, JH (октябрь 1984 г.). « Z-РНК — левосторонняя двойная спираль РНК». Природа . 311 (5986): 584–586. Бибкод : 1984Natur.311..584H . дои : 10.1038/311584a0 . ПМИД 6482970 . S2CID 4316862 .
- ^ де Роза, М.; де Санктис, Д.; Росарио, Алабама; Арчер, М.; Рич, А.; Атанасиадис, А.; Каррондо, Массачусетс (май 2010 г.). «Кристаллическая структура соединения двух спиралей Z-ДНК» . Труды Национальной академии наук . 107 (20): 9088–9092. Бибкод : 2010PNAS..107.9088D . дои : 10.1073/pnas.1003182107 . ПМК 2889044 . ПМИД 20439751 .
- ^ Чжан, Х.; Ю, Х.; Рен, Дж.; Цюй, X. (2006). «Обратимый переход B/Z-ДНК в условиях низкого содержания соли и селективность поли(dA)поли(dT) не-B-формы за счет кубаноподобного комплекса европия- L -аспарагиновой кислоты» . Биофизический журнал . 90 (9): 3203–3207. Бибкод : 2006BpJ....90.3203Z . дои : 10.1529/biophysj.105.078402 . ПМЦ 1432110 . ПМИД 16473901 .
- ^ Хо, PS; Эллисон, MJ; Куигли, Дж.Дж.; Рич, А. (1986). «Компьютерный термодинамический подход для прогнозирования образования Z-ДНК в природных последовательностях» . Журнал ЭМБО . 5 (10): 2737–2744. дои : 10.1002/j.1460-2075.1986.tb04558.x . ПМЦ 1167176 . ПМИД 3780676 .
- ^ Перейти обратно: а б Чемп, ПК; Морис, С.; Варгасон, Дж. М.; Кэмп, Т.; Хо, PS (2004). «Распределение Z-ДНК и ядерного фактора I в 22 хромосоме человека: модель сопряженной регуляции транскрипции» . Исследования нуклеиновых кислот . 32 (22): 6501–6510. дои : 10.1093/nar/gkh988 . ПМК 545456 . ПМИД 15598822 .
- ^ Ван, Эндрю Х.-Дж.; Куигли, Гэри Дж.; Колпак, Фрэнсис Дж.; Кроуфорд, Джеймс Л.; ван Бум, Жак Х.; ван дер Марель, Гийс; Рич, Александр (декабрь 1979 г.). «Молекулярная структура левостороннего фрагмента двойной спирали ДНК при атомном разрешении». Природа . 282 (5740): 680–686. Бибкод : 1979Natur.282..680W . дои : 10.1038/282680a0 . ISSN 0028-0836 . ПМИД 514347 . S2CID 4337955 .
- ^ Перейти обратно: а б с Ли, Джуйонг; Ким, Ян-Гюн; Ким, Кён Гю; Сок, Чаок (5 августа 2010 г.). «Переход между B-ДНК и Z-ДНК: ландшафт свободной энергии для распространения B-Z-перехода». Журнал физической химии Б. 114 (30): 9872–9881. CiteSeerX 10.1.1.610.1717 . дои : 10.1021/jp103419t . ISSN 1520-6106 . ПМИД 20666528 .
- ^ Перейти обратно: а б с д Ким, Сук Хо; Лим, Со Хи; Ли, Аэ-Ри; Квон, До Хун; Сон, Хён Гю; Ли, Джун-Хва; Чо, Минхэн; Джонер, Альберт; Ли, Нам Гён (23 марта 2018 г.). «Открытие пути к Z-ДНК при индуцированном белком переходе B – Z» . Исследования нуклеиновых кислот . 46 (8): 4129–4137. дои : 10.1093/nar/gky200 . ISSN 0305-1048 . ПМЦ 5934635 . ПМИД 29584891 .
- ^ Купер, Дэвид; Хм, Хеуи; Таузин, Лоуренс Дж.; Поддар, Нитеш; Ландес, Кристи Ф. (3 июня 2013 г.). «Время жизни цианиновых флуорофоров, используемых для фотообесцвечивания для одиночной молекулярной резонансной передачи энергии Фёрстера в присутствии различных систем фотозащиты» . ХимБиоХим . 14 (9): 1075–1080. дои : 10.1002/cbic.201300030 . ISSN 1439-4227 . ПМЦ 3871170 . ПМИД 23733413 .
- ^ Диденко, Владимир В. (ноябрь 2001 г.). «ДНК-зонды, использующие резонансный перенос энергии флуоресценции (FRET): конструкции и применение» . БиоТехники . 31 (5): 1106–1121. дои : 10.2144/01315rv02 . ISSN 0736-6205 . ЧВК 1941713 . ПМИД 11730017 .
- ^ Герберт, А.; Альфкен, Дж.; Ким, Ю.-Г.; Миан, И.С.; Нисикура, К.; Рич, А. (5 августа 1997 г.). «Домен, связывающий Z-ДНК, присутствует в редактирующем ферменте человека, аденозиндезаминазе двухцепочечной РНК» . Труды Национальной академии наук . 94 (16): 8421–8426. Бибкод : 1997PNAS...94.8421H . дои : 10.1073/pnas.94.16.8421 . ISSN 0027-8424 . ПМК 22942 . ПМИД 9237992 .
- ^ Перейти обратно: а б Герберт, А. (2019). «Менделевская болезнь, вызванная вариантами, влияющими на распознавание Z-ДНК и Z-РНК доменом Zα фермента редактирования двухцепочечной РНК ADAR» . Европейский журнал генетики человека . 8 (1): 114–117. дои : 10.1038/s41431-019-0458-6 . ПМК 6906422 . ПМИД 31320745 .
- ^ Герберт, А. (2019). «ADAR и подавление иммунитета при раке». Тенденции рака . 5 (5): 272–282. дои : 10.1016/j.trecan.2019.03.004 . ПМИД 31174840 . S2CID 155209484 .
- ^ Перейти обратно: а б Сурам, Анита; Рао, Джаганнатха К.С.; С., Лата К.; А., Вишвамитра М. (2002). «Первые доказательства, показывающие топологическое изменение конформации ДНК от B-ДНК до Z-ДНК в гиппокампе мозга при болезни Альцгеймера». Нейромолекулярная медицина . 2 (3): 289–298. дои : 10.1385/нмм:2:3:289 . ISSN 1535-1084 . ПМИД 12622407 . S2CID 29059186 .
- ^ Лафер, Э.М.; Валле, РП; Мёллер, А; Нордхейм, А; Шур, PH; Рич, А; Столлар, Б.Д. (1 февраля 1983 г.). «З-ДНК-специфические антитела при системной красной волчанке человека» . Журнал клинических исследований . 71 (2): 314–321. дои : 10.1172/jci110771 . ISSN 0021-9738 . ПМК 436869 . ПМИД 6822666 .
- ^ Рич, А; Чжан, С (2003). «Хронология: Z-ДНК: долгий путь к биологической функции». Обзоры природы Генетика . 4 (7): 566–572. дои : 10.1038/nrg1115 . ПМИД 12838348 . S2CID 835548 .
- ^ Виттиг, Б.; Дорбич, Т.; Рич, А. (1991). «Транкрипция связана с образованием Z-ДНК в метаболически активных пермеабилизированных ядрах клеток млекопитающих» . Труды Национальной академии наук . 88 (6): 2259–2263. Бибкод : 1991PNAS...88.2259W . дои : 10.1073/pnas.88.6.2259 . ПМК 51210 . ПМИД 2006166 .
- ^ Перейти обратно: а б Вонг, Б.; Чен, С.; Квон, Ж.-А.; Рич, А. (2007). «Характеристика Z-ДНК как нуклеосомно-граничного элемента в дрожжах Saccharomyces cerevisiae » . Труды Национальной академии наук . 104 (7): 2229–2234. Бибкод : 2007PNAS..104.2229W . дои : 10.1073/pnas.0611447104 . ЧВК 1892989 . ПМИД 17284586 .
- ^ Перейти обратно: а б с Ван, Г.; Кристенсен, Луизиана; Васкес, К.М. (2006). «Последовательности, образующие Z-ДНК, вызывают крупномасштабные делеции в клетках млекопитающих» . Труды Национальной академии наук . 108 (8): 2677–2682. Бибкод : 2006PNAS..103.2677W . дои : 10.1073/pnas.0511084103 . ПМЦ 1413824 . ПМИД 16473937 .
- ^ Перейти обратно: а б Фройнд, AM; Бичара, М.; Фукс, Р.П. (1989). «Последовательности, образующие Z-ДНК, представляют собой горячие точки спонтанных делеций» . Труды Национальной академии наук . 86 (19): 7465–7469. Бибкод : 1989PNAS...86.7465F . дои : 10.1073/pnas.86.19.7465 . ПМК 298085 . ПМИД 2552445 .
- ^ Рой Чоудхури, А.; Бакши, Р.; Ван, Дж.; Йылдырыр, Г.; Лю, Б.; Паппас-Браун, В.; Толун, Г.; Гриффит, доктор медицинских наук; Шапиро, Т.А.; Дженсен, Р.Э.; Энглунд, ПТ (декабрь 2010 г.). «Уничтожение африканских трипаносом бромидом этидия» . ПЛОС Патогены . 6 (12): e1001226. дои : 10.1371/journal.ppat.1001226 . ПМК 3002999 . ПМИД 21187912 .
- ^ Герберт, А.; Рич, А. (1993). «Метод идентификации и характеристики белков, связывающих Z-ДНК, с использованием линейного олигодезоксинуклеотида» . Исследования нуклеиновых кислот . 21 (11): 2669–2672. дои : 10.1093/нар/21.11.2669 . ПМК 309597 . ПМИД 8332463 .
- ^ Перейти обратно: а б Герберт, А.; Альфкен, Дж.; Ким, Ю.Г.; Миан, И.С.; Нисикура, К.; Рич, А. (1997). «Домен, связывающий Z-ДНК, присутствует в редактирующем ферменте человека, аденозиндезаминазе двухцепочечной РНК» . Труды Национальной академии наук . 94 (16): 8421–8426. Бибкод : 1997PNAS...94.8421H . дои : 10.1073/pnas.94.16.8421 . ПМК 22942 . ПМИД 9237992 .
- ^ Герберт, А.; Шаде, М.; Ловенхаупт, К.; Альфкен, Дж; Шварц, Т.; Шляхтенко Л.С.; Любченко Ю.Л.; Рич, А. (1998). «Домен Zα человеческого ADAR1 связывается с конформером Z-ДНК множества различных последовательностей» . Исследования нуклеиновых кислот . 26 (15): 2669–2672. дои : 10.1093/нар/26.15.3486 . ПМЦ 147729 . ПМИД 9671809 .
- ^ Шварц, Т.; Роулд, Массачусетс; Ловенхаупт, К.; Герберт, А.; Рич, А. (1999). «Кристаллическая структура домена Zα человеческого фермента редактирования ADAR1, связанного с левой Z-ДНК». Наука . 284 (5421): 1841–1845. дои : 10.1126/science.284.5421.1841 . ПМИД 10364558 .
- ^ Шаде, М.; Тернер, CJ; Кюне, Р.; Шмидер, П.; Ловенхаупт, К.; Герберт, А.; Рич, А.; Ошкинат, Х (1999). «Структура раствора домена Zα человеческого фермента редактирования РНК ADAR1 обнаруживает предварительно расположенную поверхность связывания для Z-ДНК» . Труды Национальной академии наук . 96 (22): 2465–2470. Бибкод : 1999PNAS...9612465S . дои : 10.1073/pnas.96.22.12465 . ПМК 22950 . ПМИД 10535945 .
- ^ Герберт, А.; Рич, А. (2001). «Роль связывающих доменов дсРНК и Z-ДНК в in vivo редактировании минимальных субстратов с помощью ADAR1» . Труды Национальной академии наук . 98 (21): 12132–12137. Бибкод : 2001PNAS...9812132H . дои : 10.1073/pnas.211419898 . ПМК 59780 . ПМИД 11593027 .
- ^ Хальбер, Д. (11 сентября 1999 г.). «Ученые наблюдают биологическую активность «левой» ДНК» . Пресс-служба Массачусетского технологического института . Проверено 29 сентября 2008 г.
- ^ Герберт, А. (2019). «Z-ДНК и Z-РНК при заболеваниях человека» . Коммуникационная биология . 2 :7. doi : 10.1038/s42003-018-0237-x . ПМК 6323056 . ПМИД 30729177 .
- ^ Перейти обратно: а б с Квон, Ж.-А.; Рич, А. (26 августа 2005 г.). «Биологическая функция Z-ДНК-связывающего белка E3L вируса коровьей оспы: трансактивация генов и антиапоптотическая активность в клетках HeLa» . Труды Национальной академии наук . 102 (36): 12759–12764. Бибкод : 2005PNAS..10212759K . дои : 10.1073/pnas.0506011102 . ISSN 0027-8424 . ПМК 1200295 . ПМИД 16126896 .
- ^ Перейти обратно: а б с д и Ким, Ю.-Г.; Муралинат, М.; Брандт, Т.; Пирси, М.; Хаунс, К.; Ловенхаупт, К.; Джейкобс, БЛ; Рич, А. (30 мая 2003 г.). «Роль связывания Z-ДНК в патогенезе вируса коровьей оспы» . Труды Национальной академии наук . 100 (12): 6974–6979. Бибкод : 2003PNAS..100.6974K . дои : 10.1073/pnas.0431131100 . ISSN 0027-8424 . ПМК 165815 . ПМИД 12777633 .
- ^ Ким, Ю.-Г.; Ловенхаупт, К.; Ой, Д.-Б.; Ким, К.К.; Рич, А. (2 февраля 2004 г.). «Доказательства того, что фактор вирулентности коровьей оспы E3L связывается с Z-ДНК in vivo: значение для разработки терапии поксвирусной инфекции» . Труды Национальной академии наук . 101 (6): 1514–1518. Бибкод : 2004PNAS..101.1514K . дои : 10.1073/pnas.0308260100 . ISSN 0027-8424 . ПМК 341766 . ПМИД 14757814 .
- ^ Синден, Ричард Р. (1994). Структура и функция ДНК (1-е изд.). Академическая пресса. п. 398. ИСБН 978-0-126-45750-6 .
- ^ Рич, А.; Норхейм, А.; Ван, А.Х. (1984). «Химия и биология левосторонней Z-ДНК». Ежегодный обзор биохимии . 53 (1): 791–846. дои : 10.1146/annurev.bi.53.070184.004043 . ПМИД 6383204 .
- ^ Хо, PS (27 сентября 1994 г.). «Структура d(CA/TG) n, не относящаяся к B-ДНК , не отличается от структуры Z-ДНК» . Труды Национальной академии наук . 91 (20): 9549–9553. Бибкод : 1994PNAS...91.9549H . дои : 10.1073/pnas.91.20.9549 . ПМК 44850 . ПМИД 7937803 .
Виды нуклеиновых кислот | |||||||
|---|---|---|---|---|---|---|---|
| Составляющие | |||||||
| Рибонуклеиновые кислоты (кодирование, некодирование ) |
| ||||||
| Дезоксирибонуклеиновая кислоты | |||||||
| Аналоги | |||||||
| Клонирование векторов | |||||||