Мультикопийная одноцепочечная ДНК
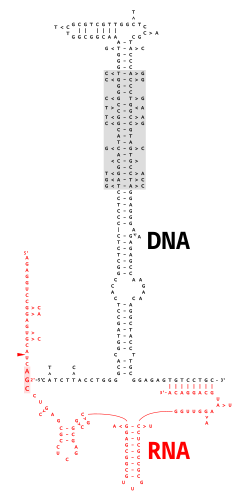
Мультикопийная одноцепочечная ДНК (мсДНК) представляет собой тип внехромосомной сателлитной ДНК одноцепочечной молекулы ДНК , ковалентно связанной через 2'-5' фосфодиэфирную связь с внутренним гуанозином молекулы РНК , которая состоит из . Полученная химера ДНК/РНК имеет две петли-стебли, соединенные ветвью, аналогичной ветвям, обнаруженным в промежуточных продуктах сплайсинга РНК . Кодирующая область мсДНК, называемая « ретрон », также кодирует тип обратной транскриптазы , которая необходима для синтеза мсДНК. [2]
Открытие
[ редактировать ]До открытия мсДНК миксобактерий у [3] [4] группы роящихся почвенных бактерий , считалось, что ферменты, известные как обратные транскриптазы (RT), существуют только у эукариот и вирусов . Это открытие привело к расширению исследований в этом районе. В результате было обнаружено, что мсДНК широко распространена среди бактерий, включая различные штаммы Escherichia coli и патогенные бактерии. [5] Дальнейшие исследования обнаружили сходство между ВИЧ обратной транскриптазой, кодируемой , и открытой рамкой считывания (ORF), обнаруженной в кодирующей области мсДНК. Тесты подтвердили наличие активности обратной транскриптазы в сырых лизатах ретрон-содержащих штаммов. [6] Хотя домен РНКазы H был предварительно идентифицирован в ORF ретрона, позже было обнаружено, что активность РНКазы H, необходимая для синтеза мсДНК, фактически обеспечивается хозяином. [7]
Ретроны
[ редактировать ]Открытие мсДНК привело к более широким вопросам относительно происхождения обратной транскриптазы, поскольку гены, кодирующие обратную транскриптазу (не обязательно связанные с мсДНК), были обнаружены у прокариот, эукариот, вирусов и даже архей . После того, как был обнаружен фрагмент ДНК, кодирующий продукцию мсДНК в E. coli , [8] было высказано предположение, что бактериофаги могли быть ответственны за внедрение гена RT в E. coli . [9] Эти открытия позволяют предположить, что обратная транскриптаза сыграла роль в эволюции вирусов из бактерий. Одна из гипотез утверждает, что с помощью обратной транскриптазы вирусы могли возникнуть как отколовшийся ген мсДНК, который приобрел белковую оболочку. Поскольку почти все гены RT участвуют в репликации ретровирусов и/или перемещении мобильных элементов , разумно предположить, что ретроны могут быть мобильными генетическими элементами, но существует мало подтверждающих доказательств такой гипотезы, за исключением наблюдаемого факта, что мсДНК широко, но спорадически рассеян среди видов бактерий, что позволяет предположить как горизонтальный, так и вертикальный перенос. [5] [10] [11] Поскольку неизвестно, представляют ли последовательности ретронов сами по себе мобильные элементы, ретроны функционально определяются их способностью продуцировать мсДНК, при этом намеренно избегая спекуляций о других возможных действиях.
Функция
[ редактировать ]Функция мсДНК остается неизвестной, хотя внутри клеток присутствует множество копий. Нокаутные мутации, которые не экспрессируют мсДНК, жизнеспособны, поэтому производство мсДНК не является необходимым для жизни в лабораторных условиях. Сверхэкспрессия мсДНК является мутагенной, по-видимому, в результате титрования репарационных белков несовпадающими парами оснований, типичными для их структуры. [10] Было высказано предположение, что мсДНК может играть определенную роль в патогенности или адаптации к стрессовым условиям. [12] Сравнение последовательностей мсДНК Myxococcus xanthus , Stigmatella aurantiaca , [1] и многие другие бактерии [5] [12] обнаруживают консервативные и гипервариабельные домены, напоминающие консервативные и гипервариабельные последовательности, обнаруженные в молекулах аллорепознания. [13] основные мсДНК M. xanthus и S. aurantiaca Например, имеют общую гомологию последовательностей на 94% , за исключением домена из 19 пар оснований, который имеет гомологию последовательностей только на 42%. [1] Наличие таких доменов важно, поскольку миксобактерии демонстрируют сложное кооперативное социальное поведение, включая роение и образование плодовых тел, в то время как E. coli и другие патогенные бактерии образуют биопленки , которые проявляют повышенную устойчивость к антибиотикам и детергентам. Устойчивость социальных собраний, требующих значительных индивидуальных затрат энергии, обычно зависит от развития механизмов аллеораспознавания , которые позволяют группам различать свое и чужое. [14]
Биосинтез
[ редактировать ]
Предполагается, что биосинтез мсДНК следует уникальному пути, который больше нигде не встречается в биохимии ДНК/РНК. Из-за сходства соединения 2'-5' ветви с соединениями ветвей, обнаруженными в промежуточных соединениях сплайсинга РНК, поначалу можно было ожидать, что образование ветвей будет происходить посредством лигирования, опосредованного сплайсосомами или рибозимами . Однако неожиданно эксперименты в бесклеточных системах с использованием очищенной обратной транскриптазы ретрона показали, что синтез кДНК осуществляется непосредственно с 2'-ОН-группы специфического внутреннего остатка G праймерной РНК. [15] RT распознает специфические структуры «стебель-петля» в РНК-предшественнике, делая синтез мсДНК с помощью RT высокоспецифичным к собственному ретрону. [16] Начало синтеза мсДНК представляет собой увлекательную задачу для нашего понимания синтеза ДНК. ДНК-полимеразы (включая RT) обладают высококонсервативными структурными особенностями, что означает, что их активные каталитические сайты мало различаются от вида к виду или даже между ДНК-полимеразами, использующими ДНК в качестве матрицы, по сравнению с ДНК-полимеразами, использующими РНК в качестве матрицы. Каталитическая область эукариотической обратной транскриптазы состоит из трех доменов, называемых «пальцы», «ладонь» и «большой палец», которые удерживают двухцепочечную матрицу праймера в правом захвате, при этом 3'-ОН праймера скрыт в активный центр полимеразы, [17] кластер высококонсервативных кислотных и полярных остатков, расположенный на ладони между указательным и средним пальцами. В эукариотических RT домен РНКазы H расположен на запястье ниже основания большого пальца, но ретронные RT лишены активности РНКазы H. Щель, связывающая нуклеиновую кислоту, простирающаяся от активного сайта полимеразы до активного сайта РНКазы H, имеет длину около 60 Å в эукариотических RT, что соответствует почти двум виткам спирали. [18] Когда эукариотическая RT удлиняет обычный праймер, растущая двойная спираль ДНК/РНК закручивается по спирали вдоль щели, и когда двойная спираль проходит через домен РНКазы H, матричная РНК переваривается с высвобождением образующейся цепи кДНК. Однако в случае удлинения праймера мсДНК длинная цепь РНК остается прикрепленной к 3'-ОН праймера G. Хотя можно смоделировать матричный комплекс RT-праймер, который сделал бы 2'-ОН доступным для реакция прайминга, [16] Дальнейшее удлинение цепи ДНК представляет собой проблему: по мере прогресса синтеза ДНК объемистая цепь РНК, отходящая от 3'-ОН, должна каким-то образом двигаться по спирали вниз по связывающей щели, не блокируясь стерическими препятствиями . Чтобы решить эту проблему, обратной транскриптазе мсДНК явно потребуются особые функции, которых нет у других RT. [10]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с Дхундейл А., Лэмпсон Б., Фуруичи Т., Иноуе М., Иноуе С. (декабрь 1987 г.). «Структура мсДНК Myxococcus xanthus: свидетельства существования длинного самоотжигающегося предшественника РНК для ковалентно связанной разветвленной РНК». Клетка . 51 (6): 1105–12. дои : 10.1016/0092-8674(87)90596-4 . ПМИД 2446773 . S2CID 21762469 .
- ^ Иноуе С., Герцер П.Дж., Иноуе М. (февраль 1990 г.). «Два независимых ретрона с весьма разнообразными обратными транскриптазами у Myxococcus xanthus» . Труды Национальной академии наук Соединенных Штатов Америки . 87 (3): 942–5. Бибкод : 1990PNAS...87..942I . дои : 10.1073/pnas.87.3.942 . ПМК 53385 . ПМИД 1689062 .
- ^ Йи Т., Фуруичи Т., Иноуе С., Иноуе М. (август 1984 г.). «Мультикопийная одноцепочечная ДНК, выделенная из грамотрицательной бактерии Myxococcus xanthus». Клетка . 38 (1): 203–9. дои : 10.1016/0092-8674(84)90541-5 . ПМИД 6088065 . S2CID 41165293 .
- ^ Фуруичи Т., Иноуе С., Иноуе М. (январь 1987 г.). «Биосинтез и структура стабильной разветвленной РНК, ковалентно связанной с 5'-концом мультикопийной одноцепочечной ДНК Stigmatella aurantiaca». Клетка . 48 (1): 55–62. дои : 10.1016/0092-8674(87)90355-2 . ПМИД 2431795 . S2CID 32376617 .
- ^ Перейти обратно: а б с Дас Р., Симамото Т., Хосен С.М., Арифуззаман М. (2011). «Сравнительное исследование различных структур мсДНК (многокопийная одноцепочечная ДНК) и филогенетическое сравнение обратных транскриптаз (ОТ): доказательства вертикального наследования» (PDF) . Биоинформация . 7 (4): 176–9. дои : 10.6026/97320630007176 . ПМЦ 3218519 . ПМИД 22102774 .
- ^ Лэмпсон BC, Сунь Дж., Сюй М.Ю., Вальехо-Рамирес Дж., Иноуе С., Иноуе М. (февраль 1989 г.). «Обратная транскриптаза в клиническом штамме Escherichia coli: производство разветвленной РНК-связанной мсДНК» (PDF) . Наука . 243 (4894, ч. 1): 1033–8. Бибкод : 1989Sci...243.1033L . дои : 10.1126/science.2466332 . ПМИД 2466332 . Архивировано из оригинала (PDF) 22 декабря 2014 г. Проверено 08 февраля 2012 г.
- ^ Лима ТМ, Лим Д (май 1995 г.). «Выделение и характеристика мутантов-хозяев с дефектом синтеза мсДНК: роль рибонуклеазы H в синтезе мсДНК» . Плазмида . 33 (3): 235–8. дои : 10.1006/plas.1995.1026 . PMID 7568472 .
- ^ Сюй М.Ю., Иноуе М., Иноуе С. (декабрь 1990 г.). «Ретрон для мультикопийной одноцепочечной ДНК из 67 оснований из Escherichia coli: потенциальный мобильный элемент, кодирующий как функции обратной транскриптазы, так и функции Dam-метилазы» . Труды Национальной академии наук Соединенных Штатов Америки . 87 (23): 9454–8. Бибкод : 1990PNAS...87.9454H . дои : 10.1073/pnas.87.23.9454 . ПМК 55184 . ПМИД 1701261 .
- ^ Иноуе С.; Иноуэ М. (1993). «Бактериальная обратная транскриптаза». В Гоффе, Стивене и Анне М. Скалка (ред.). Обратная транскриптаза . Серия монографий Колд-Спринг-Харбор. Том. 23. Плейнвью, Нью-Йорк: Лабораторное издательство Колд-Спринг-Харбор. ISBN 978-0-87969-382-4 .
- ^ Перейти обратно: а б с д Лэмпсон BC, Иноуе М, Иноуе С (2005). «Ретроны, мсДНК и бактериальный геном» (PDF) . Цитогенетические и геномные исследования . 110 (1–4): 491–9. дои : 10.1159/000084982 . ПМИД 16093702 . S2CID 24854188 . Архивировано из оригинала (PDF) 5 марта 2016 г. Проверено 08 февраля 2012 г.
- ^ Саймон Д.М., Циммерли С. (декабрь 2008 г.). «Разнообразие нехарактерных обратных транскриптаз у бактерий» . Исследования нуклеиновых кислот . 36 (22): 7219–29. дои : 10.1093/нар/gkn867 . ПМК 2602772 . ПМИД 19004871 .
- ^ Перейти обратно: а б Дас Р., Симамото Т., Арифуззаман М. (2011). «Новый штамм мсДНК (мультикопийная одноцепочечная ДНК), присутствующий в энтеропатогенных бактериях Yersinia frederiksenii ATCC 33641 Contig01029, с геномным анализом его ретрона» . Журнал патогенов . 2011 (693769): 693769. doi : 10.4061/2011/693769 . ПМЦ 3335539 . ПМИД 22567337 .
- ^ Шерман Л.А., Чаттопадьяй С. (1993). «Молекулярные основы аллорепознания». Ежегодный обзор иммунологии . 11 : 385–402. дои : 10.1146/annurev.iy.11.040193.002125 . ПМИД 8476567 .
- ^ Басс, Лео (2006). Эволюция индивидуальности . Издательство Принстонского университета. ISBN 978-0-691-08469-5 .
- ^ Симамото Т., Каваниси Х., Цучия Т., Иноуе С., Иноуе М. (июнь 1998 г.). «Синтез in vitro мультикопийной одноцепочечной ДНК с использованием отдельных праймеров и матричных РНК с помощью обратной транскриптазы Escherichia coli» . Журнал бактериологии . 180 (11): 2999–3002. дои : 10.1128/JB.180.11.2999-3002.1998 . ПМЦ 107272 . ПМИД 9603895 .
- ^ Перейти обратно: а б Иноуе С., Сюй М.Ю., Сюй А., Иноуе М. (октябрь 1999 г.). «Высокоспецифическое распознавание структур праймерной РНК для реакции прайминга 2'-ОН бактериальными обратными транскриптазами» . Журнал биологической химии . 274 (44): 31236–44. дои : 10.1074/jbc.274.44.31236 . ПМИД 10531319 .
- ^ Хакобо-Молина А., Дин Дж., Нанни Р.Г., Кларк А.Д., Лу Х, Тантилло С., Уильямс Р.Л., Камер Г., Феррис А.Л., Кларк П. (июль 1993 г.). «Кристаллическая структура обратной транскриптазы вируса иммунодефицита человека типа 1 в комплексе с двухцепочечной ДНК при разрешении 3,0 А демонстрирует изогнутую ДНК» . Труды Национальной академии наук Соединенных Штатов Америки . 90 (13): 6320–4. Бибкод : 1993PNAS...90.6320J . дои : 10.1073/pnas.90.13.6320 . ПМК 46920 . ПМИД 7687065 .
- ^ Сарафианос С.Г., Дас К., Тантилло С., Кларк А.Д., Дин Дж., Уиткомб Дж.М., Бойер П.Л., Хьюз Ш.Х., Арнольд Э. (март 2001 г.). «Кристаллическая структура обратной транскриптазы ВИЧ-1 в комплексе с полипуриновым трактом РНК: ДНК» . Журнал ЭМБО . 20 (6): 1449–61. дои : 10.1093/emboj/20.6.1449 . ПМЦ 145536 . ПМИД 11250910 .
Дальнейшее чтение
[ редактировать ]- Лэмпсон Б., Иноуе М., Иноуе С. (2001). мсДНК бактерий . Прогресс в исследованиях нуклеиновых кислот и молекулярной биологии. Том. 67. С. 65–91. дои : 10.1016/S0079-6603(01)67025-9 . ISBN 9780125400671 . ПМИД 11525386 .
- Циммерли, Стивен (2005). «Мобильные интроны и ретроэлементы у бактерий». В Маллани, Питер (ред.). Динамический бактериальный геном . Достижения молекулярной и клеточной микробиологии. Том. 8. Издательство Кембриджского университета. стр. 121–148. дои : 10.1017/CBO9780511541544.004 . ISBN 978-0-511-54154-4 .
Виды нуклеиновых кислот | |||||||
|---|---|---|---|---|---|---|---|
| Составляющие | |||||||
| Рибонуклеиновые кислоты (кодирование, некодирование ) |
| ||||||
| Дезоксирибонуклеиновая кислоты | |||||||
| Аналоги | |||||||
| Клонирование векторов | |||||||