А-ДНК
А-ДНК является одной из возможных двойных спиральных структур, которые может принять ДНК . Считается, что А-ДНК является одной из трех биологически активных двойных спиральных структур наряду с B-ДНК и Z-ДНК . Это правосторонняя двойная спираль, очень похожая на более распространенную форму B-ДНК, но с более короткой и компактной спиральной структурой, пары оснований которой не перпендикулярны оси спирали, как в B-ДНК. Его открыла Розалинд Франклин , которая также назвала формы А и В. Она показала, что ДНК переходит в форму А в условиях обезвоживания. Такие условия обычно используются для образования кристаллов, и многие кристаллические структуры ДНК находятся в форме А. [1] Такая же спиральная конформация встречается в двухцепочечных РНК и в гибридных двойных спиралях ДНК-РНК.
Структура
[ редактировать ]Как и более распространенная B-ДНК, A-ДНК представляет собой правостороннюю двойную спираль с большими и малыми бороздками. Однако, как показано в сравнительной таблице ниже, наблюдается небольшое увеличение количества пар оснований (п.н.) за ход. Это приводит к меньшему углу поворота и меньшему подъему на пару оснований, так что A-ДНК на 20-25% короче, чем B-ДНК. Большая бороздка А-ДНК глубокая и узкая, а малая бороздка широкая и неглубокая. А-ДНК шире и более сжата вдоль своей оси, чем В-ДНК. [2] [3]
Отличительной характеристикой рентгеновской кристаллографии А-ДНК является отверстие в центре. [2] А-ДНК имеет С3'-эндо- складку, которая означает, что углерод С3' в фуранозном кольце находится ниже плоскости сахара.
Сравнительная геометрия наиболее распространенных форм ДНК
[ редактировать ]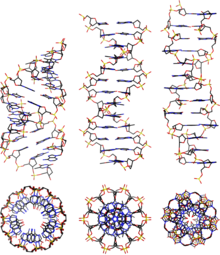

| Атрибут геометрии: | А-форма | B-форма | Z-форма |
|---|---|---|---|
| Чувство спирали | правша | правша | левша |
| Повторяющийся блок | 1 б.п. | 1 б.п. | 2 б.п. |
| Ротация/б.п. | 32.7° | 34.3° | 60°/2 |
| Среднее давление/оборот | 11 | 10 | 12 |
| Наклон б.п. к оси | +19° | −1.2° | −9° |
| Подъем/б.п. вдоль оси | 2,6 Å (0,26 нм) | 3,4 Å (0,34 нм) | 3,7 Å (0,37 нм) |
| Подъем/поворот спирали | 28,6 Å (2,86 нм) | 35,7 Å (3,57 нм) | 45,6 Å (4,56 нм) |
| Средний поворот пропеллера | +18° | +16° | 0° |
| Гликозильный угол | анти | анти | пиримидин: анти, пурин: син |
| Нуклеотидфосфат-фосфатное расстояние | 5,9 Å | 7,0 Å | С: 5,7 Å, Г: 6,1 Å |
| Сахарная складка | С3'-эндо | C2'-эндо | С: С2'-эндо, G: C3'-эндо |
| Диаметр | 23 Å (2,3 нм) | 20 Å (2,0 нм) | 18 Å (1,8 нм) |
Промежуточные A/B
[ редактировать ]Исследования также показывают, что ДНК А-формы может гибридизоваться с более распространенной B-ДНК. Эти промежуточные формы AB перенимают свойства сахарной складки и/или базовую конформацию обеих форм ДНК. В одном исследовании характерная C3'-эндо-складка обнаружена на первых трех сахарах цепи ДНК, тогда как последние три сахара имеют C2'-эндо-складку, как и B-ДНК. [2] Эти промежуточные соединения могут образовываться в водных растворах, когда основания цитозина метилируются или бромируются, изменяя конформацию. Альтернативно, было показано, что фрагменты, богатые гуанином и цитозином, легко превращаются из B в A-форму в водных растворах. [4]
Биологическая функция
[ редактировать ]А-ДНК может быть получена в результате нескольких процессов, включая дегидратацию и связывание белков. Обезвоживание ДНК переводит ее в форму А, которая, как было показано, защищает ДНК в таких условиях, как сильное высыхание бактерий. [5] [1] Связывание с белками также может удалять растворитель с ДНК и превращать ее в А-форму, о чем свидетельствует структура нескольких гипертермофильных вирусов архей. К таким вирусам относятся палочковидные рудивирусы SIRV2. [6] и ССРВ1, [7] оболочечные нитчатые липотриксвирусы AFV1, [8] СФВ1 [9] и БМП , [7] тристромавирус PFV2 [10] а также икосаэдрический портоглобовирус SPV1. [11] Считается, что А-форма ДНК является одной из адаптаций гипертермофильных вирусов архей к суровым условиям окружающей среды, в которых эти вирусы процветают.
Было высказано предположение, что двигатели, упаковывающие двухцепочечную ДНК в бактериофагах, используют тот факт, что А-ДНК короче, чем B-ДНК, и что конформационные изменения в самой ДНК являются источником больших сил, генерируемых этими двигателями. [12] Экспериментальные доказательства того, что А-ДНК является промежуточным продуктом в упаковке вирусного биомотора, получены в результате измерений резонансного переноса энергии Фёрстера с двойным красителем , показавших, что B-ДНК укорачивается на 24% в застопорившейся («сжатой») промежуточной А-форме. [13] [14] В этой модели гидролиз АТФ используется для запуска конформационных изменений белка, которые попеременно обезвоживают и регидратируют ДНК, а цикл укорочения/удлинения ДНК связан с циклом захвата/высвобождения белок-ДНК для создания поступательного движения, которое перемещает ДНК в капсид. .
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б Розалинда, Франклин (1953). «Структура волокон тимонуклеата натрия. I. Влияние содержания воды» (PDF) . Акта Кристаллографика . 6 (8): 673–677. дои : 10.1107/s0365110x53001939 .
- ^ Перейти обратно: а б с Дикерсон, Ричард Э. (1992). "Структура ДНК из ткйко З" . Структуры ДНК. Часть A: Синтез и физический анализ ДНК . Методы энзимологии. Том. 211. стр. 67–111. дои : 10.1016/0076-6879(92)11007-6 . ISBN 9780121821128 . ПМИД 1406328 .
- ^ Кокс, Майкл М. (2015). Молекулярная биология: принципы и практика . Дженнифер А. Дудна, Майкл О'Доннелл (второе изд.). Нью-Йорк. ISBN 978-1-4641-2614-7 . OCLC 905380069 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Трантырек, Лукаш; Стефл, Ричард; Ворличкова, Микаэла; Коджа, Ярослав; Стекольщик Владимир; Кипр, Ярослав (07.04.2000). «Двойная спираль ДНК А-типа, имеющая складки дезоксирибозных колец В-типа 11. Под редакцией И. Тиноко» . Журнал молекулярной биологии . 297 (4): 907–922. дои : 10.1006/jmbi.2000.3592 . ISSN 0022-2836 . ПМИД 10736226 .
- ^ Уилан Д.Р. и др. (2014). «Обнаружение массового и обратимого конформационного перехода B- к A-ДНК у прокариот в ответ на высыхание» . Интерфейс JR Soc . 11 (97): 20140454. doi : 10.1098/rsif.2014.0454 . ПМК 4208382 . ПМИД 24898023 .
- ^ Ди Майо Ф., Эгельман Э.Х. и др. (2015). «Вирус, заражающий гипертермофила, инкапсидирует ДНК А-формы» . Наука . 348 (6237): 914–917. Бибкод : 2015Sci...348..914D . дои : 10.1126/science.aaa4181 . ПМК 5512286 . ПМИД 25999507 .
- ^ Перейти обратно: а б Ван, Ф; Бакеро, ДП; Бельтран, ЖК; Су, З; Осинский, Т; Чжэн, Вт; Прангишвили, Д; Крупович, М; Эгельман, Э.Г. (5 августа 2020 г.). «Структуры нитчатых вирусов, заражающих гипертермофильные археи, объясняют стабилизацию ДНК в экстремальных условиях» . Труды Национальной академии наук Соединенных Штатов Америки . 117 (33): 19643–19652. Бибкод : 2020PNAS..11719643W . дои : 10.1073/pnas.2011125117 . ПМЦ 7443925 . ПМИД 32759221 .
- ^ Кассон, П; ДиМайо, Ф; Ю, Х; Лукас-Стаат, С; Крупович, М; Схоутен, С; Прангишвили, Д; Эгельман, Э.Х. (2017). «Модель новой мембранной оболочки нитчатого гипертермофильного вируса» . электронная жизнь . 6 : е26268. дои : 10.7554/eLife.26268 . ПМК 5517147 . ПМИД 28639939 .
- ^ Лю, Ю; Осинский, Т; Ван, Ф; Крупович, М; Схаутен, С; Кассон, П; Прангишвили, Д; Эгельман, Э.Х. (2018). «Структурная консервативность нитчатого вируса с мембранной оболочкой, инфицирующего гипертермофильный ацидофил» . Природные коммуникации . 9 (1): 3360. Бибкод : 2018NatCo...9.3360L . дои : 10.1038/s41467-018-05684-6 . ПМК 6105669 . ПМИД 30135568 .
- ^ Ван, Ф; Бакеро, ДП; Су, З; Осинский, Т; Прангишвили, Д; Эгельман, Э.Х.; Крупович, М (2020). «Структура нитчатого вируса раскрывает семейные связи внутри виросферы архей» . Эволюция вирусов . 6 (1): veaa023. дои : 10.1093/ve/veaa023 . ПМЦ 7189273 . ПМИД 32368353 .
- ^ Ван, Ф; Лю, Ю; Су, З; Осинский, Т; де Оливейра, GAP; Конвей, Дж. Ф.; Схоутен, С; Крупович, М; Прангишвили, Д; Эгельман, Э.Х. (2019). «Упаковка А-формы ДНК в икосаэдрическом вирусе» . Труды Национальной академии наук Соединенных Штатов Америки . 116 (45): 22591–22597. Бибкод : 2019PNAS..11622591W . дои : 10.1073/pnas.1908242116 . ПМК 6842630 . ПМИД 31636205 .
- ^ Харви, Южная Каролина (2015). «Гипотеза скранччервя: переходы между А-ДНК и В-ДНК обеспечивают движущую силу упаковки генома в бактериофагах с двухцепочечной ДНК» . Журнал структурной биологии . 189 (1): 1–8. дои : 10.1016/j.jsb.2014.11.012 . ПМЦ 4357361 . ПМИД 25486612 .
- ^ Орам, М. (2008). «Модуляция реакции упаковки терминазы бактериофага Т4 структурой ДНК» . Дж Мол Биол . 381 (1): 61–72. дои : 10.1016/j.jmb.2008.05.074 . ПМК 2528301 . ПМИД 18586272 .
- ^ Рэй, К. (2010). «Разрушение ДНК с помощью двигателя вирусной упаковки: сжатие субстрата Y-ДНК, остановленного прокапсидным порталом» . Вирусология . 398 (2): 224–232. дои : 10.1016/j.virol.2009.11.047 . ПМК 2824061 . ПМИД 20060554 .
Внешние ссылки
[ редактировать ]Виды нуклеиновых кислот | |||||||
|---|---|---|---|---|---|---|---|
| Составляющие | |||||||
| Рибонуклеиновые кислоты (кодирование, некодирование ) |
| ||||||
| Дезоксирибонуклеиновая кислоты | |||||||
| Аналоги | |||||||
| Клонирование векторов | |||||||