Желейный рулет складывается

Желейный рулет или складка швейцарского рулета представляет собой белковую складку или супервторичную структуру, состоящую из восьми бета-нитей, расположенных в двух четырехнитевых листах. Название структуры было введено Джейн С. Ричардсон в 1981 году, что отражает ее сходство с желе или рулетом . [ 2 ] Складка является развитием греческого ключевого мотива и иногда считается формой бета-барабана . Он очень распространен в вирусных белках , особенно в белках вирусного капсида . [ 3 ] [ 4 ] В совокупности желейный рулет и греческие ключевые структуры составляют около 30% всех бета-белков, аннотированных в базе данных Структурной классификации белков (SCOP). [ 5 ]
Структура
[ редактировать ]Базовая структура желеобразного рулона состоит из восьми бета-нитей, расположенных в двух четырехнитевых антипараллельных бета-листах, которые упаковываются вместе через гидрофобный интерфейс [где цитирование... uniprot]. Нити традиционно обозначаются от B до I по той исторической причине, что первая решенная структура капсидного белка желейного рулета из вируса кустистости томата имела дополнительную цепь A за пределами общего ядра складки. [ 6 ] [ 7 ] Листы состоят из нитей BIDG и CHEF, сложенных так, что нить B укладывается напротив нити C, I - напротив H и т. д. [ 4 ] [ 8 ]
Вирусные белки
[ редактировать ]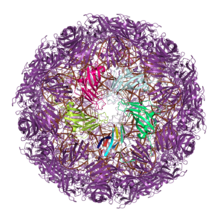
Большое количество вирусов строят свои внешние капсиды из белков, содержащих одинарную или двойную складку желейного рулета. Считается, что эта общая архитектура капсида отражает древние эволюционные отношения, возможно, возникшие еще до появления последнего универсального общего предка (LUCA) клеточной жизни. [ 8 ] [ 9 ] [ 10 ] Другие вирусные линии используют эволюционно неродственные белки для построения закрытых капсидов, которые, вероятно, эволюционировали независимо как минимум дважды. [ 9 ] [ 11 ] и, возможно, много раз, со связями с белками клеточного происхождения. [ 12 ]
Капсидные белки в виде одиночных желейных рулетов
[ редактировать ]Белки одиночного желейного капсида (JRC) обнаружены как минимум в шестнадцати различных вирусных семействах , в основном с икосаэдрическими структурами капсида и включая как РНК-вирусы, так и ДНК-вирусы . [ 13 ] Многие вирусы с одиночными желейными капсидами представляют собой вирусы с одноцепочечной РНК с положительным смыслом . Две группы вирусов с двухцепочечной ДНК с капсидами с одним JRC — это Papillomaviridae и Polyomaviridae , обе из которых имеют довольно маленькие капсиды; у этих вирусов архитектура собранного капсида ориентирует ось желейного валика параллельно или «горизонтально» относительно поверхности капсида. [ 11 ] Крупномасштабный анализ компонентов вирусного капсида показал, что одиночный горизонтальный желеобразный рулон является наиболее распространенной складкой среди белков капсида, на которую приходится около 28% известных примеров. [ 12 ]
Другая группа вирусов использует в своих капсидах отдельные желеобразные белки, но в вертикальной, а не горизонтальной ориентации. Эти вирусы эволюционно родственны большой группе вирусов с двойным желеобразным роллом, известной как , PRD1 вирусная линия аденовируса со схожей архитектурой капсида, реализуемой посредством сборки двух отдельных основных капсидных белков с одинарным желеобразным роллом, экспрессируемых из разных генов. [ 14 ] [ 15 ] Эти одиночные вертикальные желейные вирусы составляют таксон Helvetiavirae . [ 16 ] Известные вирусы с вертикальным одиночным желеобразным капсидом заражают экстремофильные прокариоты . [ 14 ] [ 12 ]
Белковые роллы с двойным желе
[ редактировать ]

Капсидные белки двойного желейного рулона состоят из двух одинарных складок желейного рулона, соединенных короткой линкерной областью. Они обнаружены как в вирусах с двухцепочечной ДНК, так и в вирусах с одноцепочечной ДНК , по крайней мере, десяти различных вирусных семейств, включая вирусы, которые заражают все сферы жизни и охватывают большой диапазон размеров капсида. [ 4 ] [ 11 ] [ 18 ] В архитектуре капсида с двумя желейными валками ось желейного валика ориентирована перпендикулярно или «вертикально» относительно поверхности капсида. [ 19 ]
Считается, что белки двойных желейных рулетов произошли от одиночных белков желейных рулетов в результате дупликации генов . [ 11 ] [ 19 ] Вполне вероятно, что вирусы с вертикальными одиночными желеобразными роллами представляют собой переходную форму и что белки капсида с вертикальными и горизонтальными желеобразными роллами имеют независимое эволюционное происхождение от предковых клеточных белков. [ 12 ] Степень структурного сходства капсидов вирусов с двойной желеобразной формой привела к выводу, что эти вирусы, вероятно, имеют общее эволюционное происхождение, несмотря на их разнообразие по размеру и кругу хозяев; это стало известно как линия PRD1 - аденовируса ( Bamfordvirae ). [ 19 ] [ 16 ] [ 20 ] [ 21 ] Многие члены этой группы были идентифицированы с помощью метагеномики , и в некоторых случаях они практически не имеют общих вирусных генов. [ 12 ] [ 22 ] Хотя большинство членов этой группы имеют икосаэдрическую геометрию капсида, некоторые семейства, такие как Poxviridae и Ascoviridae, имеют зрелые вирионы овальной или кирпичной формы; Поксвирусы, такие как Vaccinia, претерпевают резкие конформационные изменения, опосредованные высокопроизводными белками двойного желеобразного рулона, во время созревания и, вероятно, происходят от икосаэдрического предка. [ 11 ] [ 23 ] Общие капсидные белки с двойным желеобразным рулоном, наряду с другими гомологичными белками, также были упомянуты в поддержку предполагаемого порядка Megavirales, содержащего большие нуклеоцитоплазматические ДНК-вирусы (NCLDV). [ 24 ]
Первоначально считалось, что белки двойного желеобразного рулета уникальны для вирусов, поскольку они не наблюдались в клеточных белках. [ 11 ] Однако в 2022 году сравнение белковых структур выявило несколько семейств настоящих клеточных белков с двойной желейной складкой. [ 25 ]
Некапсидные белки
[ редактировать ]Одиночные желеобразные рулоны также встречаются в некапсидных вирусных белках, включая второстепенные компоненты собранного вириона , а также невирионные белки, такие как полиэдрин . [ 11 ] суперсемейства 30К, В вирусах растений белки движения ответственные за межклеточный транспорт вирусных геномов или целых капсидов через каналы плазмодесм, имеют одинарную желеобразную складку и произошли от капсидных белков небольших икосаэдрических вирусов. [ 26 ]
Клеточные белки
[ редактировать ]В белках клеточного происхождения встречаются как одинарные, так и двойные желейные складки. [ 11 ] [ 12 ] [ 25 ] Одним из классов клеточных белков с одинарной желеобразной складкой являются нуклеоплазмины , которые служат молекулярными белками-шаперонами для сборки гистонов в нуклеосомы . N -концевой домен нуклеоплазминов имеет одну желеобразную складку и собран в пентамер. [ 27 ] С тех пор аналогичные структуры были обнаружены в дополнительных группах белков, ремоделирующих хроматин . [ 28 ] Мотивы желеобразных рулонов с идентичными связями бета-листов также обнаружены в фактора некроза опухоли. лигандах [ 29 ] и белки бактерии Yersinia pseudotuberculosis , которые принадлежат к классу вирусных и бактериальных белков, известных как суперантигены . [ 30 ] [ 31 ]
В более широком смысле, членов чрезвычайно разнообразного суперсемейства купинов также часто называют рулетами с желе; хотя общее ядро доменной структуры купина содержит только шесть бета-цепей, многие купины имеют восемь. [ 32 ] Примеры включают негем - диоксигеназные ферменты. [ 33 ] [ 34 ] (включая альфа-кетоглутарат-зависимые гидроксилазы ) и JmjC семейства деметилазы гистонов . [ 35 ] [ 36 ]
Клеточные белки с двойной желеобразной складкой включают гликозидгидролазы семейства DUF2961, пептид:N-гликозидазу F (PNGases F) и пептидилглицин-альфа-амидирующую монооксигеназу . [ 25 ]

Заметным различием между PNGазами F и другими белками двойного желеобразного рулона является отсутствие α-спиралей, которые следуют за нитями F и F' в капсидных белках и DUF2961. Эквивалентные области являются вариабельными в PNGases F и содержат либо длинные петли, либо вставки. Напротив, желеобразные домены белков DUF2961 содержат вставки коротких β-шпильок выше G- и G'-цепей двойной складки желейного рулона. Важно отметить, что белки семейства DUF2961 образуют тримеры, напоминающие вирусные капсомеры. [ 25 ]
Эволюция
[ редактировать ]Сравнительные исследования белков, классифицированных как структуры желейных рулетов и греческих ключевых структур, позволяют предположить, что ключевые греческие белки развились значительно раньше, чем их более топологически сложные аналоги желейных рулетов. [ 5 ] Исследования структурной биоинформатики, сравнивающие желейные белки капсида вируса с другими белками известной структуры, показывают, что белки капсида образуют хорошо разделенный кластер, что позволяет предположить, что они подвержены определенному набору эволюционных ограничений. [ 4 ] Одной из наиболее примечательных особенностей желейных белков вирусного капсида является их способность образовывать олигомеры с повторяющимся рисунком мозаики с образованием закрытой белковой оболочки; клеточные белки, наиболее сходные по складкам и топологии, в большинстве случаев также являются олигомерами. [ 4 ] Было высказано предположение, что вирусные желеобразные капсидные белки произошли из клеточных желейных белков, возможно, в нескольких независимых случаях, на самых ранних стадиях клеточной эволюции. [ 12 ]
История и номенклатура
[ редактировать ]Название «рулет с желе» было впервые использовано для структуры, состоящей из разработки греческого ключевого мотива Джейн С. Ричардсон в 1981 году, и было призвано отразить сходство структуры с желейным или швейцарским рулетом . [ 2 ] Структуре дали множество описательных названий, в том числе клин, бета-бочка и бета-ролл. Края двух листов не соприкасаются, образуя регулярные структуры водородных связей , поэтому его часто не считают настоящим бета-цилиндром . [ 3 ] хотя этот термин широко используется при описании архитектуры вирусного капсида. [ 14 ] [ 15 ] Клеточные белки, содержащие желеобразные структуры, могут быть описаны как купиновая складка, складка JmjC или двухцепочечная бета-спираль. [ 34 ]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б Ларсон С.Б., Дэй Дж.С., Макферсон А. (сентябрь 2014 г.). «Спутниковый вирус табачной мозаики, уточненный до разрешения 1,4 Å» . Акта Кристаллографика. Раздел D. Биологическая кристаллография . 70 (Часть 9): 2316–30. дои : 10.1107/S1399004714013789 . ПМЦ 4157444 . ПМИД 25195746 .
- ^ Перейти обратно: а б Ричардсон Дж.С. (1981). «Анатомия и систематика структуры белка». Достижения в области химии белков, том 34 . Том. 34. стр. 167–339. дои : 10.1016/S0065-3233(08)60520-3 . ISBN 9780120342341 . ПМИД 7020376 .
- ^ Перейти обратно: а б Челванаягам Г., Херинга Дж., Аргос П. (ноябрь 1992 г.). «Анатомия и эволюция белков, отображающих топологию желейного рулета вирусного капсида». Журнал молекулярной биологии . 228 (1): 220–42. дои : 10.1016/0022-2836(92)90502-Б . ПМИД 1447783 .
- ^ Перейти обратно: а б с д и Ченг С., Брукс CL (7 февраля 2013 г.). «Белки вирусного капсида разделены в пространстве структурных складок» . PLOS Вычислительная биология . 9 (2): e1002905. Бибкод : 2013PLSCB...9E2905C . дои : 10.1371/journal.pcbi.1002905 . ПМЦ 3567143 . ПМИД 23408879 .
- ^ Перейти обратно: а б Эдвардс Х., Абельн С., Дин С.М. (14 ноября 2013 г.). «Изучение предпочтений складчатого пространства у новорожденных и древних белковых суперсемейств» . PLOS Вычислительная биология . 9 (11): e1003325. Бибкод : 2013PLSCB...9E3325E . дои : 10.1371/journal.pcbi.1003325 . ПМЦ 3828129 . ПМИД 24244135 .
- ^ Харрисон С.К., Олсон А.Дж., Шутт С.Э., Винклер Ф.К., Бриконь Дж. (ноябрь 1978 г.). «Вирус густого куста томата с разрешением 2,9 А». Природа . 276 (5686): 368–73. Бибкод : 1978Natur.276..368H . дои : 10.1038/276368a0 . ПМИД 19711552 . S2CID 4341051 .
- ^ Россманн М.Г., Абад-Сапатеро С., Мурти М.Р., Лильяс Л., Джонс Т.А., Страндберг Б. (апрель 1983 г.). «Структурные сравнения некоторых небольших сферических вирусов растений». Журнал молекулярной биологии . 165 (4): 711–36. дои : 10.1016/S0022-2836(83)80276-9 . ПМИД 6854630 .
- ^ Перейти обратно: а б Бенсон С.Д., Бэмфорд Дж.К., Бэмфорд Д.Х., Бернетт Р.М. (декабрь 2004 г.). «Выявляет ли общая архитектура вирусную линию, охватывающую все три сферы жизни?» . Молекулярная клетка . 16 (5): 673–85. doi : 10.1016/j.molcel.2004.11.016 . ПМИД 15574324 .
- ^ Перейти обратно: а б Фортерре П., Прангишвили Д. (сентябрь 2009 г.). «Происхождение вирусов». Исследования в области микробиологии . 160 (7): 466–72. дои : 10.1016/j.resmic.2009.07.008 . ПМИД 19647075 . S2CID 2767388 .
- ^ Холмс ЕС (июнь 2011 г.). «Что эволюция вирусов говорит нам о происхождении вирусов?» . Журнал вирусологии . 85 (11): 5247–51. дои : 10.1128/JVI.02203-10 . ПМК 3094976 . ПМИД 21450811 .
- ^ Перейти обратно: а б с д и ж г час Крупович М., Бэмфорд Д.Х. (август 2011 г.). «Двухцепочечные ДНК-вирусы: 20 семейств и всего пять различных архитектурных принципов сборки вирионов». Современное мнение в вирусологии . 1 (2): 118–24. дои : 10.1016/j.coviro.2011.06.001 . ПМИД 22440622 .
- ^ Перейти обратно: а б с д и ж г Крупович М., Кунин Е.В. (март 2017 г.). «Множественное происхождение белков вирусного капсида от клеточных предков» . Труды Национальной академии наук Соединенных Штатов Америки . 114 (12): Е2401–Е2410. Бибкод : 2017PNAS..114E2401K . дои : 10.1073/pnas.1621061114 . ПМЦ 5373398 . ПМИД 28265094 .
- ^ Крупович М. (октябрь 2013 г.). «Сети эволюционных взаимодействий, лежащие в основе полифилетического происхождения вирусов оцДНК». Современное мнение в вирусологии . 3 (5): 578–86. дои : 10.1016/j.coviro.2013.06.010 . ПМИД 23850154 .
- ^ Перейти обратно: а б с Гил-Картон Д., Яаккола С.Т., Чарро Д., Перальта Б., Кастаньо-Диес Д., Оксанен Х.М. и др. (октябрь 2015 г.). «Изучение сборки вирусов с вертикальными основными капсидными белками с одним β-цилиндром» . Структура . 23 (10): 1866–1877. дои : 10.1016/j.str.2015.07.015 . ПМИД 26320579 .
- ^ Перейти обратно: а б Сантос-Перес И., Чарро Д., Гил-Картон Д., Азкаргорта М., Элорца Ф., Бэмфорд Д.Х. и др. (март 2019 г.). «Структурная основа сборки вертикальных одноствольных вирусов β-ствола» . Природные коммуникации . 10 (1): 1184. Бибкод : 2019NatCo..10.1184S . дои : 10.1038/s41467-019-08927-2 . ПМК 6414509 . ПМИД 30862777 .
- ^ Перейти обратно: а б Кунин Е.В., Доля В.В., Крупович М., Варсани А., Вольф Ю.И., Ютин Н., Зербини М., Кун Дж.Х. (октябрь 2019 г.). «Создать мегатаксономическую структуру, заполнив все основные таксономические ранги, для ДНК-вирусов, кодирующих основные капсидные белки типа вертикального желеобразного рулона» . Предложение ICTV (Taxoprop) : 2019.003G. дои : 10.13140/RG.2.2.14886.47684 .
- ^ Перейти обратно: а б Абресия Н.Г., Граймс Дж.М., Кивеля Х.М., Ассенберг Р., Саттон Г.К., Батчер С.Дж. и др. (сентябрь 2008 г.). «Понимание эволюции вируса и мембранного биогенеза на основе структуры морского липидсодержащего бактериофага PM2» . Молекулярная клетка . 31 (5): 749–61. doi : 10.1016/j.molcel.2008.06.026 . ПМИД 18775333 .
- ^ Лаанто Э., Мянтюнен С., Де Колибус Л., Марьякангас Дж., Гиллум А., Стюарт Д.И. и др. (август 2017 г.). «Вирус, обнаруженный в бореальном озере, связывает вирусы оцДНК и дцДНК» . Труды Национальной академии наук Соединенных Штатов Америки . 114 (31): 8378–8383. Бибкод : 2017PNAS..114.8378L . дои : 10.1073/pnas.1703834114 . ПМЦ 5547622 . ПМИД 28716906 .
- ^ Перейти обратно: а б с Крупович М., Бэмфорд Д.Х. (декабрь 2008 г.). «Эволюция вируса: насколько далеко простирается вирусная линия с двойным бета-цилиндром?». Обзоры природы. Микробиология . 6 (12): 941–8. дои : 10.1038/nrmicro2033 . ПМИД 19008892 . S2CID 31542714 .
- ^ Кунин Е.В., Доля В.В., Крупович М., Варсани А., Вольф Ю.И., Ютин Н. и др. (май 2020 г.). «Глобальная организация и предлагаемая мегатаксономия мира вирусов» . Обзоры микробиологии и молекулярной биологии . 84 (2): e00061–19, /mmbr/84/2/MMBR.00061–19.atom. дои : 10.1128/MMBR.00061-19 . ПМК 7062200 . ПМИД 32132243 .
- ^ Уокер П.Дж., Сидделл С.Г., Лефковиц Э.Дж., Мушегян А.Р., Адриансенс Э.М., Демпси Д.М. и др. (ноябрь 2020 г.). «Изменения в таксономии вирусов и Уставе, ратифицированные Международным комитетом по таксономии вирусов (2020 г.)» . Архив вирусологии . 165 (11): 2737–2748. дои : 10.1007/s00705-020-04752-x . ПМИД 32816125 . S2CID 221182789 .
- ^ Ютин Н., Бэкстрем Д., Эттема Т.Дж., Крупович М., Кунин Е.В. (апрель 2018 г.). «Огромное разнообразие геномов прокариотических вирусов, кодирующих основные капсидные белки двойного желе, обнаруженные с помощью анализа геномных и метагеномных последовательностей» . Вирусологический журнал . 15 (1): 67. дои : 10.1186/s12985-018-0974-y . ПМК 5894146 . ПМИД 29636073 .
- ^ Бахар М.В., Грэм С.С., Стюарт Д.И., Граймс Дж.М. (июль 2011 г.). «Понимание эволюции сложного вируса на основе кристаллической структуры вируса коровьей оспы D13» . Структура . 19 (7): 1011–20. дои : 10.1016/j.str.2011.03.023 . ПМК 3136756 . ПМИД 21742267 .
- ^ Колсон П., Де Ламбаллери X, Ютин Н., Асгари С., Биго Ю., Бидеши Д.К. и др. (декабрь 2013 г.). « Мегавирусы», предлагаемый новый порядок эукариотических нуклеоцитоплазматических крупных ДНК-вирусов» . Архив вирусологии . 158 (12): 2517–21. дои : 10.1007/s00705-013-1768-6 . ПМК 4066373 . ПМИД 23812617 .
- ^ Перейти обратно: а б с д Крупович, М; Макарова К.С.; Кунин Е.В. (1 февраля 2022 г.). «Клеточные гомологи основных белков капсида с двойным желеобразным роллом проясняют происхождение древнего вирусного царства» . Труды Национальной академии наук Соединенных Штатов Америки . 119 (5): e2120620119. Бибкод : 2022PNAS..11920620K . дои : 10.1073/pnas.2120620119 . ПМЦ 8812541 . ПМИД 35078938 .
- ^ Буткович, А; Доля, В.В.; Кунин Е.В.; Крупович, М (2023). «Белки движения растительных вирусов произошли из белков желейного капсида» . ПЛОС Биология . 21 (6): e3002157. дои : 10.1371/journal.pbio.3002157 . ПМЦ 10306228 . ПМИД 37319262 .
- ^ Дутта С., Эйки И.В., Дингуолл С., Хартман К.Л., Лауэ Т., Нолте Р.Т. и др. (октябрь 2001 г.). «Кристаллическая структура ядра нуклеоплазмина: значение для связывания гистонов и сборки нуклеосом» . Молекулярная клетка . 8 (4): 841–53. дои : 10.1016/S1097-2765(01)00354-9 . ПМИД 11684019 .
- ^ Эдлих-Мут С., Артеро Дж.Б., Кэллоу П., Пшевлока М.Р., Уотсон А.А., Чжан В. и др. (май 2015 г.). «Пентамерная складка нуклеоплазмина присутствует у Drosophila FKBP39 и большого количества белков, связанных с хроматином» . Журнал молекулярной биологии . 427 (10): 1949–63. дои : 10.1016/j.jmb.2015.03.010 . ПМЦ 4414354 . ПМИД 25813344 .
- ^ Бодмер Дж.Л., Шнайдер П., Чопп Дж. (январь 2002 г.). «Молекулярная архитектура суперсемейства TNF» . Тенденции биохимических наук . 27 (1): 19–26. дои : 10.1016/S0968-0004(01)01995-8 . ПМИД 11796220 .
- ^ Донадини Р., Лью К.В., Кван А.Х., Маккей Дж.П., Филдс Б.А. (январь 2004 г.). «Структуры кристаллов и раствора суперантигена Yersinia pseudotuberculosis обнаруживают желеобразную складку» . Структура . 12 (1): 145–56. дои : 10.1016/j.str.2003.12.002 . ПМИД 14725774 .
- ^ Фрейзер Дж. Д., Профт Т (октябрь 2008 г.). «Бактериальный суперантиген и суперантигеноподобные белки». Иммунологические обзоры . 225 (1): 226–43. дои : 10.1111/j.1600-065X.2008.00681.x . ПМИД 18837785 . S2CID 39174409 .
- ^ Хури С., Баккер Ф.Т., Данвелл Дж.М. (апрель 2001 г.). «Филогения, функции и эволюция купинов, структурно консервативного, функционально разнообразного суперсемейства белков» . Молекулярная биология и эволюция . 18 (4): 593–605. doi : 10.1093/oxfordjournals.molbev.a003840 . ПМИД 11264412 .
- ^ Озер А., Брюик Р.К. (март 2007 г.). «Негемовые диоксигеназы: клеточные сенсоры и регуляторы в одном лице?». Химическая биология природы . 3 (3): 144–53. дои : 10.1038/nchembio863 . ПМИД 17301803 .
- ^ Перейти обратно: а б Айк В., Макдонаф М.А., Талхаммер А., Чоудхури Р., Шофилд С.Дж. (декабрь 2012 г.). «Роль желейной складки в связывании субстрата 2-оксоглутаратоксигеназой». Современное мнение в области структурной биологии . 22 (6): 691–700. дои : 10.1016/j.sbi.2012.10.001 . ПМИД 23142576 .
- ^ Чен З., Занг Дж., Уэтстин Дж., Хонг Х., Давразу Ф., Кутателадзе Т.Г. и др. (май 2006 г.). «Структурные данные о деметилировании гистонов членами семейства JMJD2» . Клетка . 125 (4): 691–702. дои : 10.1016/j.cell.2006.04.024 . ПМИД 16677698 . S2CID 15273763 .
- ^ Клозе Р.Дж., Чжан Ю (апрель 2007 г.). «Регуляция метилирования гистонов путем деметилиминирования и деметилирования». Обзоры природы. Молекулярно-клеточная биология . 8 (4): 307–18. дои : 10.1038/nrm2143 . ПМИД 17342184 . S2CID 2616900 .
Внешние ссылки
[ редактировать ]- Антипараллельные β-домены , раздел из «Анатомия и таксономия структуры белка». книги Джейн С. Ричардсон
- «Желеобразный рулет жизни» Жаклин Хамфрис из блога «Учтенные мелочи» , спонсируемого Американским обществом микробиологии.