Генетическая история Италии
| История Италии |
|---|
 |
Генетическая история Италии включает в себя информацию о формировании, этногенезе и другую ДНК-специфическую информацию о жителях Италии. Современные итальянцы в основном происходят от древних народов Италии , в том числе от индоевропейцев ( римляне и другие латиняне , фалиски , пиценты , умбрийцы , самниты , оски , сикелы и адриатические венеты , а также греки в Великой Греции , цизальпийские галлы и япиги ). и доиндоевропейские носители ( этруски , лигуры , реты и камунны в материковой Италии, сиканы на Сицилии, нурагики на Сардинии и финикийцы как на Сицилии, так и на Сардинии). Другие группы мигрировали в Италию в результате Римской империи , когда итальянский полуостров привлекал людей из различных регионов империи (Северная Африка, Ближний Восток и остальная часть Европы). [2] и в средние века с приходом остготов , лонгобардов , сарацинов и норманнов , среди других. На основе анализа ДНК имеются свидетельства региональной генетической субструктуры и преемственности в современной Италии, восходящей к античности. [3] [4] [5] [6]
In their admixture ratios, Italians are similar to other Southern Europeans, and that is being of primarily Neolithic Early European Farmer ancestry, along with smaller, but still significant, amounts of Mesolithic Western Hunter-Gatherer, Bronze Age Steppe pastoralist (Indo-European speakers) and Chalcolithic or Bronze Age Iranian/Caucasus-related ancestry.[4][7][8][9] Southern Italians are closest to the modern Greeks,[10] while the Northern Italians are closest to the Spaniards and Southern French.[11][12][13][14] There is also Bronze/Iron Age West Asian and Middle Eastern admixture in Italy, with a much lower incidence in Northern Italy compared with Central Italy and Southern Italy.[15][8] North African admixture is also found in Southern Italy and the main islands, with the highest incidence being in Sicily and Sardinia.[15][8][4]
Overview
[edit]Latin samples from Latium in the Iron Age and early Roman Republican period were generally found to genetically cluster closest to modern Northern and Central Italians (four out of six were closest to Northern and Central Italians, while the other two were closest to Southern Italians).[16] DNA analysis demonstrates that ancient Greek colonization had a significant lasting effect on the local genetic landscape of Southern Italy and Sicily (Magna Graecia), with modern people from that region having significant Greek admixture.[17][18] Overall, the genetic differentiation between the Latins, Etruscans and the preceding proto-Villanovan population of Italy was found to be insignificant.[19] In 2019, aDNA analysis of Roman fossils detected substantial genetic ancestry shift towards central and northern European ancestry in the inhabitants of the city of Rome in late antiquity and the medieval era. The authors tentatively link the origin of this ancestry with Visigoths and Lombards. Previously, most citizens in the imperial era clustered with central and east Mediterranean peoples, such as central and south Italians, Greeks, Cypriots and Maltese, and to some extent, Levantine and Near Eastern peoples. This was caused by direct immigration and contact with Greek, Phoenician and Punic diasporas.[2][20] A 2020 analysis of maternal haplogroups from ancient and modern samples indicates a substantial genetic similarity and continuity between the modern inhabitants of Umbria in central Italy and ancient inhabitants of the region belonging to the Italic-speaking Umbrian culture.[6]
Multiple DNA studies confirmed that genetic variation in Italy is clinal, going from the Eastern to the Western Mediterranean. The Sardinians are the exception as genetic outliers in Italy and indeed in Europe, resulting from their predominantly Neolithic, Pre-Indo-European and non-Italic Nuragic ancestry.[21][7][8][9] Reflecting the history of Europe and the broader Mediterranean basin, the Italian populations have been found to be made up mostly of the same ancestral components, albeit in different proportions, from the Mesolithic, Neolithic and Bronze Age settlements of Europe.[15][12][22]
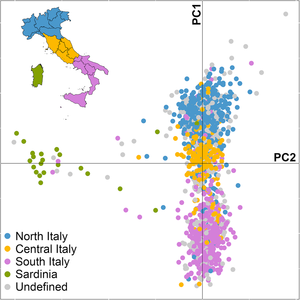
The genetic gap between northern and southern Italians is filled by an intermediate Central Italian cluster, creating a continuous cline of variation that mirrors geography.[23] The only exceptions are some minority populations (mostly Slovene minorities from the region of Friuli-Venezia Giulia) who cluster with the Slavic-speaking Central Europeans in Slovenia,[24] as well as the Sardinians, who are clearly differentiated from the populations of both mainland Italy and Sicily.[12][4] A study on some linguistic and isolated communities residing in Italy revealed that their genetic diversity at short (0–200 km) and intermediate distances (700–800 km) was greater than that observed throughout the entire European continent.[5]
The genetic distance between Northern and Southern Italians, although large for a single European nationality, is similar to that between the Northern and the Southern Germans.[25] Northern and Southern Italians began to diverge as early as the Late Glacial, and appear to encapsulate at a smaller scale the cline of genetic diversity observable across Europe.[26]
Historical populations of Italy
[edit]
Modern humans appeared during the Upper Paleolithic. Specimens of Aurignacian age were discovered in the cave of Fumane and date back about 34,000 years. During the Magdalenian period, the first humans from the Pyrenees populated Sardinia.[27]
During the Neolithic period, farming was introduced by people from the east and the first villages were built. Weapons became more sophisticated and the first objects in clay were produced. In the late Neolithic era the use of copper spread, and villages were built over piles near lakes. In Sardinia, Sicily and a part of Mainland Italy the Beaker culture spread from Western and Central Europe. Sicily also suffered the influences of the Aegean in the Mycenaean period.
During the Late Bronze Age the Urnfield Proto-Villanovan culture appeared in Central and Northern Italy. It is characterized by the rite of cremation of dead bodies, which originated from Central Europe. The use of iron began to spread.[28] In Sardinia, the Nuragic civilization flourished.
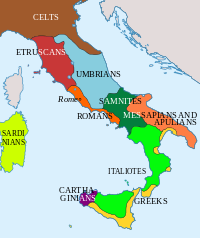
At the dawn of the Iron Age much of Italy was inhabited by Italic tribes such as the Latins, Sabines, Samnites, and Umbrians. The Northwest and Alpine territories were populated primarily by pre-Indo European speakers such as the Etruscans, Ligurians, Camunni and Raetians; while Iapygian tribes, possibly of Illyrian origin, populated Apulia.
From the 8th century BC, Greek colonists settled on the southern Italian coast and founded cities, forming what would be later called Magna Graecia. Around the same time, Phoenician colonists settled on the western side of Sicily. During the same period the Etruscan civilization developed on the coast of Southern Tuscany and Northern Latium. In the 4th century BC, Gauls settled in Northern Italy and in parts of Central Italy.[29] With the fall of the Western Roman Empire, different populations of Germanic origin invaded Italy, the most significant being the Lombards,[30] followed five centuries later by the Normans in Sicily.
Etruscans
[edit]
There have been a number of genetic studies of Etruscans and modern Tuscans compared with other populations, some of which indicate the local, European origin of Etruscans and others supportive of an origin from elsewhere. In general, the direct testing of ancient Etruscan DNA has supported a deep, local origin, while the testing of modern samples as a proxy for Etruscans is rather inconclusive and inconsistent.[31][32]
The very large mtDNA study from 2013 indicates, based on maternally-inherited DNA from 30 bone samples taken from tombs dating from the eight century to the first century BC from Tuscany and Lazio, that the Etruscans were a native population.[33][34] The study extracted and typed the hypervariable region of mitochondrial DNA of 14 individuals buried in two Etruscan necropoleis, analyzing them along with previously analyzed Etruscan mtDNA, other ancient European mtDNA, modern and Medieval samples from Tuscany, and 4,910 modern individuals from the Mediterranean basin. The ancient (30 Etruscans, 27 Medieval Tuscans) and modern DNA sequences (370 Tuscans) were subjected to several million computer simulation runs, showing that the Etruscans can be considered ancestral to Medieval and, especially in the subpopulations from Casentino and Volterra, of modern Tuscans; modern populations from Murlo and Florence, by contrast, were shown not to continue the Medieval population. By further considering two Anatolian samples (35 and 123 individuals), it was estimated that the genetic links between Tuscany and Anatolia date back to at least 5,000 years ago, and the "most likely separation time between Tuscany and Western Anatolia falls around 7,600 years ago", strongly suggesting that the Etruscan culture developed locally, and not as an immediate consequence of immigration from the Eastern Mediterranean shores. According to the study, ancient Etruscan mtDNA is closest among modern European populations and is not particularly close to Anatolian or other Eastern Mediterranean populations. Among ancient populations based on mtDNA, ancient Etruscans were found to be closest to LBK Neolithic farmers from Central Europe.[33][34]
This result is largely in line with previous mtDNA results from 2004 (in a smaller study also based on ancient DNA), and contradictory to results from 2007 (based on modern DNA). The 2004 study was based on mitochondrial DNA (mtDNA) from 80 bone samples, reduced to 28 bone samples in the analysis phase, taken from tombs dating from the seventh century to the third century BC from Veneto, Tuscany, Lazio and Campania.[35] This study found that the ancient DNA extracted from the Etruscan remains had some affinities with modern European populations including Germans, English people from Cornwall, and Tuscans in Italy. In addition the Etruscan samples possibly revealed more genetic inheritance from the eastern and southern Mediterranean than modern Italian samples contain. The study was marred by concerns that mtDNA sequences from the archeological samples represented severely damaged or contaminated DNA;[36] however, subsequent investigation showed that the samples passed the most stringent tests of DNA degradation available.[37]
A mtDNA study, published in 2018 in the journal American Journal of Physical Anthropology, compared both ancient and modern samples from Tuscany, from the Prehistory, Etruscan age, Roman age, Renaissance, and Present-day, and concluded that the Etruscans appear as a local population, intermediate between the prehistoric and the other samples, placing in the temporal network between the Eneolithic Age and the Roman Age.[38]
A 2019 genetic study published in the journal Science analyzed the remains of eleven Iron Age individuals from the areas around Rome, of which four were Etruscan individuals, one buried in Veio Grotta Gramiccia from the Villanovan period (900-800 BC) and three buried in La Mattonara Necropolis near Civitavecchia from the Orientalizing period (700-600 BC). The study concluded that Etruscans (900–600 BC) and the Latins (900–500 BC) from Latium vetus were genetically similar.,[39] genetic differences between the examined Etruscans and Latins were found to be insignificant.[40] The Etruscan individuals and contemporary Latins were distinguished from preceding populations of Italy by the presence of 30.7% steppe ancestry.[41] Their DNA was a mixture of two-thirds Copper Age ancestry (EEF + WHG; Etruscans ~66–72%, Latins ~62–75%) and one-third Steppe-related ancestry (Etruscans ~27–33%, Latins ~24–37%) (with the EEF component mainly deriving from Neolithic-era migrants to Europe from Anatolia and the WHG being local Western European hunter-gatherers, with both components, along with that from the steppe, being found in virtually all European populations).[39] The only sample of Y-DNA extracted belonged to haplogroup J-M12 (J2b-L283), found in an individual dated 700-600 BC, and carried exactly the M314 derived allele also found in a Middle Bronze Age individual from Croatia (1631-1531 calBCE). While the four samples of mtDNA extracted belonged to haplogroups U5a1, H, T2b32, K1a4.[42] Therefore, Etruscans had also Steppe-related ancestry despite speaking a pre-Indo-European language.
В исследовании 2021 года, проведенном Институтом Макса Планка , университетами Тюбингена , Флоренции и Гарварда , опубликованном в журнале Science Advances , были проанализированы Y-хромосома , митохондриальная ДНК и аутосомная ДНК 82 древних образцов из Этрурии (Тоскана и Лацио) и южная Италия (Базиликата), охватывающая период с 800 г. до н.э. по 1000 г. н.э., включая 48 человек железного века. Исследование подтвердило, что в образцах этрусков из Тосканы и Лацио предковый компонент Степь присутствовал в тех же процентных количествах, что и в ранее проанализированных образцах латинян железного века, и добавило, что в ДНК этрусков полностью отсутствовал, что является сигналом недавнего смешения с Анатолией или Восточным Средиземноморьем. Исследование пришло к выводу, что этруски были автохтонами и имели генетический профиль, аналогичный таковому у их латинских соседей раннего железного века. И этруски, и латиняне прочно принадлежали к европейскому кластеру: 75% выборок этрусских особей мужского пола оказались принадлежащими к гаплогруппе. R1b , особенно R1b-P312 и его производное R1b-L2, прямым предком которого является R1b-U152. Что касается гаплогрупп митохондриальной ДНК, то наиболее распространенной была в основном H, за которой следовали J и T. Данные однородительских маркеров и данные аутосомной ДНК из образцов этрусских людей железного века позволяют предположить, что Этрурия получила миграции с большим наследственным степным компонентом во 2-м тысячелетии до нашей эры , связанные с гаплогруппами митохондриальной ДНК. к распространению индоевропейских языков , начиная с культуры колокольных кубков , и что эти миграции слились с популяциями древнейшего доиндоевропейского слоя, существовавшего, по крайней мере, со времен неолита, но выжил именно язык последнего, Ситуация аналогична той, что произошла в баскском регионе на севере Испании . Исследование также пришло к выводу, что проанализированные образцы показывают, что этруски сохраняли свой генетический профиль неизменным в течение почти 1000 лет, что указывает на редкое присутствие в Этрурии иностранцев, и что демографические изменения в Этрурии произошли только с периода Римской империи. , в котором происходит примесь к местному населению предковых компонентов из Восточного Средиземноморья. Анализ образцов людей, живших в период Римской империи и в эпоху Средневековья, также позволяет предположить, что генетический ландшафт современной центральной Италии сформировался в основном около 1000 лет назад после вторжений варваров и что прибытие германских лангобардов в Италии способствовали формированию генофонда современного населения Тосканы и северного Лацио. [43]
В 2024 году были изучены 6 экземпляров этрусских останков из Тарквинии , Лацио , датированные 9-7 веками до нашей эры, которые подтвердили предыдущие находки. Модель примеси показала, что они на 84-92% были итальянскими колокольчиками и на 8-26% дополнительными предками Ямной Самары (связанными со Степью), но при этом один человек был более похож на популяции железного века из Скандинавии и северо-западной Европы. Два человека мужского пола, изучавшие Y-хромосому, принадлежали к линии J2b/J-M12, а пять изученных митохондриальных гаплогрупп были типичными для постнеолитической Европы. Фенотипические черты включали голубые глаза, светло-/темно-каштановые волосы и бледный или средний оттенок кожи. [44]
Исследование мтДНК, проведенное в 2007 году, напротив, ранее предполагало ближневосточное происхождение. [45] Ахилли и др . (2007) обнаружили в современной выборке из 86 человек из Мурло, небольшого городка на юге Тосканы, необычно высокую частоту (17,5%) предполагаемых ближневосточных гаплогрупп мтДНК, в то время как другие тосканские популяции не демонстрируют такой же поразительной особенности. Основываясь на этом результате, Ахилли пришел к выводу, что «их данные подтверждают сценарий постнеолитического генетического вклада с Ближнего Востока в современное население Тосканы, сценарий, согласующийся с лидийским происхождением этрусков». Это исследование подверглось резкой критике со стороны археологов, этрускологов и классиков. [46] В отсутствие каких-либо свидетельств датировки нет прямой связи между этим генетическим вкладом, обнаруженным у Мурло, и этрусков. Более того, нет никаких доказательств того, что эти гаплогруппы мтДНК, обнаруженные в Мурло, могут быть доказательством восточного происхождения этрусков, поскольку некоторые из этих гаплогрупп мтДНК были обнаружены в других исследованиях еще в эпоху неолита и энеолита в Италии и Германии. [38] Все гаплогруппы мтДНК, обнаруженные в современной выборке из Мурло и классифицированные Achilli et al. Ближневосточного происхождения на самом деле широко распространены в современных образцах из других областей Италии и Европы, не связанных с этрусками. [47]
Недавнее исследование Y-ДНК, проведенное в 2018 году на современной выборке из 113 человек из Вольтерры , города этрусского происхождения, Grugni et al. оставляет все возможности открытыми, хотя автохтонный сценарий наиболее подкреплен цифрами, и приходит к выводу, что «наличие J2a-M67* (2,7%) предполагает контакты по морю с анатолийскими народами, обнаружение центральноевропейской линии G2a-L497 (7,1%) со значительной частотой скорее поддерживают североевропейское происхождение этрусков, тогда как высокая распространенность европейских линий R1b (R1b 49,8%, R1b-U152 24,5%) не может исключить сценарий автохтонного процесса формирования этрусков. цивилизации от предшествующего общества Вилланова, как предполагает Дионисий Галикарнасский». [48] В Италии Y-ДНК J2a-M67*, пока не обнаруженная в этрусских образцах, более распространена на побережье Адриатического моря между Марке и Абруццо , а не в тех, где когда-то жили этруски, и в исследовании имеет свой пик в Ионическом море. сторона Калабрии . [49] [50] позднего бронзового века было обнаружено, что образец культуры Кьятице В 2014 году в Венгрии представляет собой J2a1-M67, [51] пара J2a1b была обнаружена в образцах позднего неолита из культуры LBK в Австрии , [52] J2a1a был обнаружен в образце культуры Сопота среднего неолита из Хорватии . [52] J2a был обнаружен в образце культуры Лендьель позднего неолита из Венгрии . [53] В 2019 году в Стэнфордском исследовании, опубликованном в журнале Science, было обнаружено, что два древних образца из неолитического поселения Рипабьянка-ди-Монтерадо в провинции Анкона , в регионе Марке в Италии, представляют собой Y-ДНК J-L26 и J-M304. [39] В 2021 году еще в двух древних образцах из поселения Халколитич Гротта-ла-Сасса в провинции Латина на юге Лацио было обнаружено Y-ДНК J2a7-Z2397. [54] Следовательно, Y-ДНК J2a-M67, вероятно, находится в Италии со времен неолита и не может быть доказательством недавних контактов с Анатолией.
Недавние исследования популяционной структуры современных итальянцев показали, что в Италии существует линия с севера на юг по линиям Y-хромосомы и аутосомным локусам с четкой дифференциацией полуостровных итальянцев от сардинцев и что современные тосканцы представляют собой популяцию Центральная Италия генетически наиболее близка к жителям северной Италии . [55] Исследование 2019 года, основанное на аутосомной ДНК 1616 человек из всех 20 итальянских административных регионов, пришло к выводу, что тосканцы присоединяются к северному итальянскому кластеру, близкому к жителям Лигурии и Эмилии-Романьи . [56] Исследование 2013 года, основанное на однородительских маркерах 884 неродственных людей из 23 итальянских мест, показало, что структура, наблюдаемая для отцовских линий в континентальной Италии и Сицилии, предполагает общий генетический фон между людьми из Тосканы и Северной Италии, с одной стороны, и людьми из Южной Италии и Адриатического побережья с другой стороны. Наиболее частыми гаплогруппами Y-ДНК в группе, представленной популяциями Северо-Запада Италии, включая Тоскану и большую часть равнины Падана, являются четыре R1b-линии (R-U152*, R-M269*, R-P312* и R -L2*). [50]
В коллективном сборнике «Этрускология», опубликованном в 2017 году, британский археолог Фил Перкинс представляет анализ состояния исследований ДНК и пишет, что «ни одно из исследований ДНК на сегодняшний день убедительно не доказывает, что этруски были интрузивным населением в Италии, возникшим в Восточном Средиземноморье или Анатолия» и «есть признаки того, что данные ДНК могут подтвердить теорию о том, что этрусские народы являются автохтонными в центральной Италии». [32]
В своей книге «Краткая история человечества» , опубликованной в 2021 году, немецкий генетик Йоханнес Краузе , содиректор Института эволюционной антропологии Макса Планка в Йене , приходит к выводу, что вполне вероятно, что этрусский язык (а также баскский , палео-сардинский и минойский ) «развился на континенте в ходе неолитической революции ». [57]Генетическое разнообразие Y-ДНК
[ редактировать ]
Многие итальянцы , особенно в Северной Италии и Центральной Италии, принадлежат к гаплогруппе R1b , распространенной в Западной и Центральной Европе. Самая высокая частота R1b встречается в Гарфаньяне (76,2%) в Тоскане и в долинах Бергамо (80,8%) в Ломбардии . [58] [59] Этот процент ниже на юге Италии, в Калабрии (33,2%). [58] С другой стороны, 39% сардинцев принадлежат к мезолитической европейской гаплогруппе I2a1a . [60] [61]
Исследование, проведенное в Университете Каттолики дель Сакро Куоре, показало, что, хотя греческая колонизация оставила мало значимого генетического вклада, анализ данных, взятых из 12 участков на итальянском полуострове, подтвердил модель диффузии мужских демов и неолитическую примесь с мезолитическим населением. [62] Результаты подтвердили распределение генетических вариаций вдоль оси север-юг и подтвердили демическую диффузию. Образцы из Южной Италии сгруппированы с образцами из Юго-Восточной и Юго-Центральной Европы, а северные группы - с образцами из Западной Европы. [62] [63]
Исследование 2004 года, проведенное Semino et al. показало, что итальянцы из северо-центральных регионов имели около 26,9% J2; у апулийцев, калабрийцев и сицилийцев было 29,1%, 21,5% и 16,7% J2 соответственно; было у сардинцев 9,7% J2. [64]
Генетическое исследование 2018 года, посвященное линиям Y-хромосомы и гаплогрупп, их разнообразию и распространению с участием около 817 репрезентативных субъектов, отдает должное традиционному разделению населения на север и юг, делая вывод, что из-за неолитических миграций южные итальянцы «показывают более высокое сходство с популяциями Ближнего Востока и Южных Балкан , чем с северными, и наоборот, северные образцы генетически ближе к группам Северо-Западной Европы и Северных Балкан». Положение Вольтерры в центральной Тоскане оставляет дискуссию о происхождении этрусков открытой, хотя цифры решительно говорят в пользу автохтонного тезиса: низкое присутствие J2a-M67* (2,7%) предполагает контакты по морю с анатолийскими народами; присутствие центральноевропейской линии G2a-L497 (7,1%) со значительной частотой скорее подтверждает центральноевропейское происхождение этрусков; и, наконец, высокая распространенность европейских линий R1b (R1b примерно 50%, R1b-U152 24,5%) — особенно гаплогруппы R1b-U152 — может указывать на автохтонное происхождение, обусловленное процессом формирования этрусской цивилизации из предшествующей Вилланованская культура , следуя теориям Дионисия Галикарнасского , [58] что уже подтверждено археологией, антропологией и лингвистикой. [65] [66] [67] [68] В 2019 году в Стэнфордском исследовании, опубликованном в журнале Science, было обнаружено, что два древних образца из неолитического поселения Рипабьянка-ди-Монтерадо в провинции Анкона , в регионе Марке в Италии, представляют собой Y-ДНК J-L26 и J-M304. [2] Следовательно, Y-ДНК J2a-M67, следующая за Y-ДНК J-L26 и J-M304, скорее всего, находится в Италии со времен неолита и не может быть доказательством недавних контактов с Анатолией.
Y-ДНК, занесенная исторической иммиграцией
[ редактировать ]В двух деревнях Лацио и Абруццо ( Каппадокия и Валлепьетра ) I1 является наиболее распространенной Y-ДНК, зарегистрированной на уровнях 35% и 28%. [69] На Сицилии дальнейшие миграции вандалов и сарацин лишь незначительно повлияли на этнический состав сицилийского народа. Однако конкретно греческое генетическое наследие на Сицилии оценивается в 37%. [17]
Норманнское завоевание южной Италии привело к созданию Норманнского королевства Сицилия в 1130 году со столицей в Палермо, через 70 лет после первоначального нормандского вторжения и через 40 лет после завоевания последнего города, Ното в 1091 году, и просуществовало до 1198 года. В настоящее время нормандская Y-ДНК распространена в центральной и западной Сицилии: от 15% до 20% линий принадлежат к гаплогруппе I , в восточной части острова этот процент падает до 8%. Вклад североафриканских мужчин в Сицилию оценивается от 0% до 7,5%. [70] [18] [71] В целом, предполагаемый отцовский вклад Южных Балкан и Западной Европы на Сицилии составляет около 63% и 26% соответственно. [71]
Генетическое исследование, проведенное в 2015 году в шести небольших горных деревнях на востоке Лацио и одной горной общине в близлежащем западном Абруццо, выявило некоторые генетические сходства между этими общинами и популяциями Ближнего Востока , в основном в мужском генетическом пуле. Гаплогруппа Y Q , распространенная в Западной и Центральной Азии , также была обнаружена среди этой выборки населения, что позволяет предположить, что в прошлом здесь могло находиться поселение из Анатолии. [69] Также он составляет около 0,6% в континентальной Италии , но возрастает до 2,5% (6/236) на Сицилии , где достигает 16,7% (3/18) в регионе Мадзара-дель-Валло , за которым следует 7,1% (2/28). в Рагузе , 3,6% в Шакке , [17] и 3,7% в Бельведере-Мариттимо . [72]
Генетический состав итальянской мтДНК
[ редактировать ]большинство линий мтДНК принадлежат к гаплогруппе H. В Италии, как и везде в Европе , Несколько независимых исследований пришли к выводу, что гаплогруппа H, вероятно, возникла в Западной Азии c. 25 000 лет назад. Он был занесен в Европу миграциями c. 20–25 000 лет назад и распространился вместе с населением юго-запада континента. [73] [74] Его появление примерно совпало с возникновением Граветтской культуры. Распространение субкладов H1, H3 и сестринской гаплогруппы V отражает вторую внутриевропейскую экспансию из Франко-Кантабрийского региона после последнего ледникового максимума , ок. 13 000 лет назад. [73] [75]
Линии африканской гаплогруппы L встречаются относительно нечасто (менее 1%) по всей Италии, за исключением Лациума , Вольтерры , Базиликаты и Сицилии, где частота встречаемости составляет от 2 до 3%. [76]
Исследование Brisighelli et al., проведенное в 2012 году. заявил, что анализ наследственных информативных маркеров , «проведенный в настоящем исследовании, показал, что в Италии наблюдается очень незначительный компонент Африки к югу от Сахары, который, однако, немного выше, чем в несредиземноморской Европе». Обсуждая африканские мтДНК, в исследовании говорится, что они указывают на то, что значительная часть этих линий могла попасть в Италию более 10 000 лет назад; следовательно, их присутствие в Италии не обязательно относится ко времени Римской империи, атлантической работорговли или современной миграции». [22] Эти мтДНК, полученные Brisighelli et al. были представлены с такими результатами как «Гаплотипы митохондриальной ДНК африканского происхождения в основном представлены гаплогруппами M1 (0,3%), U6 (0,8%) и L (1,2%) для 583 протестированных образцов. [22] Гаплогруппы M1 и U6 можно считать североафриканским происхождением и, следовательно, их можно использовать для обозначения документально подтвержденного исторического вклада Африки. Гаплогруппа M1 наблюдалась только у двух носителей из Трапани (Западная Сицилия), тогда как U6 наблюдалась только в Лучере, Южная Апулия, и еще один на оконечности полуострова (Калабрия). [22]
Исследование 2013 года, проведенное Алессио Боаттини и соавт. нашел 0 африканских гаплогрупп L во всей Италии из 865 образцов. Проценты берберских гаплогрупп M1 и U6 составляли 0,46% и 0,35% соответственно. [77]
Исследование Стефании Сарно и соавт., проведенное в 2014 году. обнаружил 0 африканских гаплогрупп L и M1 на материковой части Южной Италии из 115 образцов. Из 115 образцов были найдены только два берберских U6: один из Лечче и один из Козенцы. [71]
Близкое генетическое сходство между ашкенази и итальянцами было отмечено в генетических исследованиях, возможно, из-за того, что евреи-ашкенази имеют значительную европейскую примесь (30–60%), в основном южноевропейскую, многие из которых прибыли из Италии, когда были евреями. мужчины диаспоры ближневосточного происхождения мигрировали в Рим и нашли жен среди местных женщин, которые затем обратились в иудаизм . [78] [79] [80] [81] [82] [13] [83] Более конкретно, евреи-ашкенази можно смоделировать как 50% левантийцев и 50% европейцев, при этом средняя доля южноевропейской примеси, по оценкам, составляет 37,5%. Большая часть (30,5%), по-видимому, имеет итальянское происхождение. [84] [85]
Исследование еврейской генеалогии 2010 года показало, что что касается нееврейских европейских групп, населением, которое наиболее тесно связано с евреями-ашкенази, являются современные итальянцы, за которыми следуют французы и сардинцы. [86] [87]
Недавние исследования показали, что Италия сыграла важную роль в восстановлении « Западной Европы » в конце Последнего ледникового периода . Исследование, которое было сосредоточено на митохондриальной гаплогруппе U5b3, обнаружило, что эта женская линия на самом деле зародилась в Италии и около 10 000 лет назад распространилась от полуострова в сторону Прованса и Балкан . В Провансе, вероятно, между 9000 и 7000 лет назад, он дал начало субкладу гаплогруппы U5b3a1. Этот субклад U5b3a1 позже попал из Прованса на остров Сардиния через торговцев обсидианом, поскольку, по оценкам, 80% обсидиана, находящегося во Франции, поступает из Монте-Арчи на Сардинии, что отражает тесную связь, которая когда-то существовала между этими двумя регионы. Еще около 4% женского населения Сардинии принадлежит к этому гаплотипу. [88]
Исследование мтДНК, опубликованное в 2018 году в журнале American Journal of Physical Anthropology , сравнило как древние, так и современные образцы из Тосканы, из предыстории , этрусской эпохи, римской эпохи , эпохи Возрождения и наших дней, и пришло к выводу, что этруски появляются как местное население, промежуточное между доисторическими и другими образцами, помещающееся во временной сети между энеолитом и римским веком. [89]
Анализ материнских гаплогрупп 2020 года из древних и современных образцов в центральном итальянском регионе Умбрия обнаруживает существенное генетическое сходство между современными умбрийцами и доримскими жителями этого региона, а также доказательства существенной генетической преемственности в регионе с доримских времен до римских времен. подарок. Было обнаружено, что как современные, так и древние умбрийцы имеют высокий уровень гаплогрупп мтДНК U4 и U5a, а также чрезмерное представительство J (примерно 30%). Исследование также показало, что «локальная генетическая преемственность дополнительно подтверждается шестью терминальными ветвями (H1e1, J1c3, J2b1, U2e2a, U8b1b1 и K1a4a)», которые также характерны для древних и современных умбрийцев. [6]
Аутосомный
[ редактировать ]
Южная/Западная Европа Северная/Восточная Европа Кавказ
Западная Азия Южная Азия Восточная Азия Северная Африка/Страны Африки к югу от Сахары [90]
Уэйд и др. (2008) определили, что Италия является одним из двух последних оставшихся генетических островов в Европе, вторым является Финляндия . Частично это связано с наличием альпийской горной цепи, которая на протяжении веков предотвращала крупные миграционные потоки. [91]
Недавние полногеномные исследования позволили обнаружить и количественно оценить примесь, как никогда раньше. Ли и др. (2008), используя более 600 000 аутосомных SNP, идентифицируют семь глобальных групп населения, включая Европу, Ближний Восток и Центральную/Южную Азию. Все итальянские образцы относятся к центрально-западной группе с незначительными влияниями, относящимися к периоду неолита. [92]
Лопес Эрраес и др. (2009) типировали те же образцы, насчитывающие около 1 миллиона SNP, и проанализировали их в контексте Западной Евразии, выявив ряд подкластеров. На этот раз во всех европейских образцах присутствуют незначительные примеси. Среди итальянцев по-прежнему больше всего в Тоскане, немного и на Сардинии, а также в Ломбардии (Бергамо), которая находится еще дальше на север. [93]
Исследование Мурджани и соавт., проведенное в 2011 году. обнаружили, что многие южные европейцы унаследовали 1–3% предков, живущих к югу от Сахары, хотя при повторном анализе с помощью статистической модели «СТРУКТУРА» эти проценты были ниже (0,2–2,1%). Была также рассчитана средняя дата примеси около 55 поколений/1100 лет назад, что «согласуется с потоком генов в Северной Африке в конце Римской империи и последующими арабскими миграциями». [94]
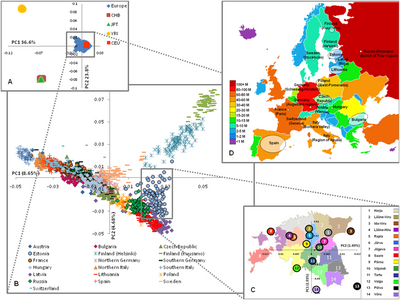
Исследование 2012 года, проведенное Ди Гаэтано и соавт. использовали 1014 итальянцев с широким географическим охватом. Оно показало, что нынешнее население Сардинии может быть четко дифференцировано генетически от материковой Италии и Сицилии, и что определенная степень генетической дифференциации обнаруживается внутри нынешнего населения итальянского полуострова.
Используя программное обеспечение ADMIXTURE, авторы получили при K = 4 наименьшую ошибку перекрестной проверки. Лица HapMap CEU показали среднее происхождение из Северной Европы (NE) 83%. Аналогичная картина наблюдается во французском, североитальянском и центральноитальянском населении с северо-восточным происхождением 70%, 56% и 52% соответственно. Согласно графику PCA, также в анализе ADMIXTURE имеются относительно небольшие различия в происхождении между северными итальянцами и центральными итальянцами, в то время как южные итальянцы показали более низкую среднюю долю примеси NE (44%), чем северная и центральная Италия, и более высокое европеоидное происхождение итальянцев. 28%. Образцы Сардинии демонстрируют образец малинового цвета, общий для других европейских популяций, но с более высокой частотой (70%).Средняя доля примесей североевропейского происхождения в нынешнем сардинском населении составляет 14,3%, при этом некоторые люди имеют очень низкую североевропейскую родословную (менее 5% у 36 человек из 268, что составляет 13% выборки). [12]
Исследование Peristera Paschou et al., проведенное в 2013 году. подтверждает, что Средиземное море стало сильным барьером для потока генов из-за географической изоляции после первоначальных поселений. Образцы из (Северной) Италии, Тосканы, Сицилии и Сардинии наиболее близки к другим южным европейцам из Иберии, Балкан и Греции, которые, в свою очередь, наиболее близки к неолитическим мигрантам, распространившим земледелие по всей Европе, представленным здесь каппадокийским образцом из Анатолии. Но с тех пор не было никакого значительного примеси из Ближнего Востока или Северной Африки в Италию и остальную часть Южной Европы. [14]
Анализ древней ДНК показывает, что Эци Ледяной человек группируется с современными южными европейцами и наиболее близок к итальянцам (оранжевые точки «Европа S» на графиках ниже), особенно с острова Сардиния. Другие итальянцы отодвигаются в сторону Юго-Восточной и Центральной Европы в соответствии с географией и некоторым постнеолитическим потоком генов из этих областей (например, италийцы, греки, этруски, кельты), но, несмотря на это и многовековую историю, они все еще очень похожи на своих доисторических предок. [95]
Исследование Botigué et al., проведенное в 2013 году. В 2013 году применили алгоритм неконтролируемой кластеризации ADMIXTURE для оценки общего распределения аллелей между африканцами и европейцами. Что касается итальянцев, то североафриканское происхождение не превышает 2% их геномов. В среднем 1% еврейского происхождения встречается у тосканского населения HapMap и итальянских швейцарцев, а также у греков и киприотов. Вопреки предыдущим наблюдениям, происхождение жителей стран южнее Сахары выявляется у <1% в Европе, за исключением Канарских островов. [71]
Хаак и др. (2015) провели полногеномное исследование 94 древних скелетов из Европы и России. В исследовании утверждается, что степные скотоводы бронзового века из ямной культуры распространили индоевропейские языки в Европе. Аутосомные тесты показывают, что ямные люди произошли в результате смешения двух разных популяций охотников-собирателей: восточных охотников-собирателей из русской степи и либо кавказских охотников-собирателей, либо иранцев энеолита (которые очень похожи). Вольфганг Хаак оценил 27% наследственный вклад ямной в ДНК современных тосканцев , 25% наследственный вклад ямной в ДНК современных северных итальянцев из Бергамо , исключая сардинцев (7%) и в меньшей степени сицилийцев ( 12%). [11]
Исследование Sazzini et al., проведенное в 2016 году, подтверждает результаты предыдущих исследований Di Gaetano et al. (2012) и Фиорито и др. (2015), но имеет гораздо лучший географический охват образцов: тестирование прошли 737 человек из 20 мест в 15 различных регионах. Исследование также впервые включает формальный тест на примесь, который моделирует происхождение итальянцев путем вывода о событиях примеси с использованием всех образцов Западной Евразии. Результаты очень интересны в свете древних доказательств ДНК, которые появились за последние пару лет:
В дополнение к закономерности, описанной в основном тексте, выборка SARD, по-видимому, сыграла важную роль в качестве источника примеси для большинства исследованных популяций, особенно итальянских, а не как реципиента миграционных процессов. Фактически, наиболее значимые значения f3 для трио, включающих SARD, указывают на то, что полуостровные итальянцы являются правдоподобным результатом смешения SARD и популяций из Ирана , Кавказа и России . Этот сценарий можно интерпретировать как еще одно свидетельство того, что сардинцы сохраняют значительную часть предполагаемого предкового геномного фона, который был широко распространен по всей Европе, по крайней мере, до неолита и который впоследствии был стерт или замаскирован у большинства современных европейских популяций. [23]
Сарно и др. (2017) концентрируются на генетическом воздействии исторических миграций вокруг Средиземноморья на Южную Италию и Сицилию и приходят к выводу, что «результаты демонстрируют, что генетическая изменчивость современных популяций Южной Италии характеризуется общей генетической преемственностью, простирающейся до большие части центральных и восточных побережий Средиземного моря», показывая при этом, что «Южная Италия больше похожа на грекоязычные острова Средиземного моря, простирающиеся на восток до Кипра , чем на образцы из континентальной Греции, что предполагает возможную родовую связь, которая возможно, сохранились на островах в менее смешанной форме», также уточняется, что «помимо преобладающего неолитического компонента, наш анализ обнаруживает значительное влияние постнеолитических кавказских и левантийских предков». [18] Новостная статья, связанная с Обществом Макса Планка , в которой рассматриваются результаты, хотя она начинается с заявления о том, что «популяции вдоль восточного побережья Средиземного моря имеют общее генетическое наследие, выходящее за пределы национальности», также указывает, насколько это исследование интересно для дебатов, касающихся распространения индоевропейской языковой семьи в Европе, поскольку, несмотря на влияние Кавказа , не существует генетического маркера, связанного с Понтийско-Каспийской степью , «очень характерный генетический сигнал, хорошо представленный в Северо-Центральной и Восточной Европе, который предыдущие исследования связано с появлением на континенте индоевропейских языков». [18]
Равеан и др. (2019) в ходе полногеномного исследования современных итальянцев обнаружили вклад кавказских охотников-собирателей из третьего тысячелетия анатолийского бронзового века, преимущественно в Южной Италии. Кроме того, в соответствии с предыдущими исследованиями модели региональных различий показали географическую структуру в Южной Италии, Северной Италии и Сардинии. Еще более детальная структура наблюдалась между субрегиональными кластерами, вызванная географией и расстоянием, а также исторической примесью, возможно, связанной с событиями конца Римской империи и в последующие периоды. [4]
Антонио и др. и Рима в разные периоды времени (2019) изучали исторические популяции Лациума . Они обнаружили, что, несмотря на лингвистические различия, у латинян и этрусков не было выявлено существенных генетических различий. Их аутосомная ДНК представляла собой смесь в одинаковых пропорциях западных охотников-собирателей (мезолит), ранних европейских земледельцев (неолит) и западных степных скотоводов (бронзовый век). [2]
Полногеномное исследование более 700 человек из Южного Средиземноморья (102 из Южной Италии) в 2022 году в сочетании с древней ДНК из соседних регионов выявило высокое сходство юго-восточных итальянцев с современными восточными пелопоннесцами , а также более близкое родство древних Греческие геномы совпадают с геномами из определенных регионов Южной Италии, чем геномы современной Греции. [96]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Пароло С., Лиза А., Джентилини Д., Ди Блазио А.М., Барлера С., Николис Э.Б. и др. (ноябрь 2015 г.). «Характеристика биологических процессов, формирующих генетическую структуру итальянского населения» . БМК Генетика . 16 : 132. дои : 10.1186/s12863-015-0293-x . ПМК 4640365 . ПМИД 26553317 .
- ^ Jump up to: а б с д Антонио М.Л., Гао З., Мутс Х.М., Луччи М., Кандилио Ф., Сойер С. и др. (ноябрь 2019 г.). «Древний Рим: генетический перекресток Европы и Средиземноморья» . Наука . 366 (6466). Американская ассоциация содействия развитию науки (опубликовано 8 ноября 2019 г.): 708–714. Бибкод : 2019Sci...366..708A . дои : 10.1126/science.aay6826 . hdl : 2318/1715466 . ПМК 7093155 . ПМИД 31699931 .
Интересно, что, хотя люди железного века были отобраны как из этрусского (n=3), так и из латинского (n=6) контекстов, мы не обнаружили каких-либо существенных различий между двумя группами со статистикой f4 в форме f4(RMPR_Etruscan, RMPR_Latin; test популяция, Онге), что позволяет предположить общее происхождение или обширный генетический обмен между ними. ... В средневековый и ранний современный периоды (n = 28 особей) мы наблюдаем сдвиг предков в сторону Центральной и Северной Европы в PCA (рис. 3E), а также дальнейшее увеличение европейского кластера (C7) и утрату кластеров Ближнего Востока и Восточного Средиземноморья (C4 и C5) в ChromoPainter (рис. 4C). Средневековое население примерно состояло из современных жителей Центральной Италии (рис. 3F). Его можно смоделировать как двустороннюю комбинацию позднеантичного населения Рима и европейского населения-донора, с потенциальными источниками, включающими многие древние и современные популяции в Центральной и Северной Европе: лангобардов из Венгрии, саксов из Англии и викингов из Швеции, среди другие (таблица S26).
- ^ Ральф П., Куп Дж. (2013). «География недавнего генетического происхождения в Европе» . ПЛОС Биология . 11 (5): e1001555. дои : 10.1371/journal.pbio.1001555 . ПМЦ 3646727 . ПМИД 23667324 .
- ^ Jump up to: а б с д и Равеан А., Анели С., Монтинаро Ф., Афанасиадис Г., Барлера С., Бироло Г. и др. (сентябрь 2019 г.). «Структура населения современных итальянцев раскрывает образцы древнего и архаичного происхождения в Южной Европе» . Достижения науки . 5 (9): eaaw3492. Бибкод : 2019SciA....5.3492R . дои : 10.1126/sciadv.aaw3492 . ПМК 6726452 . ПМИД 31517044 .
- ^ Jump up to: а б Капокаса М., Анагносту П., Бачис В., Баттаджа С., Бертончини С., Бионди Г. и др. (2014). «Лингвистическая, географическая и генетическая изоляция: совместное исследование итальянского населения» . Журнал антропологических наук . 92 (92): 201–31. doi : 10.4436/JASS.92001 (неактивен 26 апреля 2024 г.). ПМИД 24607994 .
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на апрель 2024 г. ( ссылка ) - ^ Jump up to: а б с Моди А., Лансиони Х., Кардинали И., Каподиферро М.Р., Рамбальди Мильоре Н., Хусейн А. и др. (июль 2020 г.). «Митогеномный портрет Умбрии в Центральной Италии, изображенный современными жителями и доримскими руинами» . Научные отчеты . 10 (1): 10700. Бибкод : 2020НатСР..1010700М . дои : 10.1038/s41598-020-67445-0 . ПМЦ 7329865 . ПМИД 32612271 .
- ^ Jump up to: а б Чианг К.В., Маркус Дж.Х., Сидоре С., Бидданда А., Аль-Асади Х., Золедзевска М. и др. (октябрь 2018 г.). «Геномная история населения Сардинии» . Природная генетика . 50 (10): 1426–1434. дои : 10.1038/s41588-018-0215-8 . ПМК 6168346 . ПМИД 30224645 .
- ^ Jump up to: а б с д Маркус Дж. Х., Пост С., Рингбауэр Х., Лай Л., Скитс Р., Сидоре С. и др. (февраль 2020 г.). «Генетическая история от среднего неолита до представления на средиземноморском острове Сардиния» . Природные коммуникации . 11 (1): 939. Бибкод : 2020NatCo..11..939M . дои : 10.1038/s41467-020-14523-6 . ПМК 7039977 . ПМИД 32094358 .
- ^ Jump up to: а б Фернандес Д.М., Миттник А., Олальде И., Лазаридис И., Черонет О., Роланд Н. и др. (март 2020 г.). «Распространение степного и иранского происхождения на островах западного Средиземноморья» . Экология и эволюция природы . 4 (3): 334–345. дои : 10.1038/s41559-020-1102-0 . ПМК 7080320 . ПМИД 32094539 .
- ^ «Сицилия и Южная Италия были сильно колонизированы греками, начиная с восьмого по девятый век до нашей эры. Демографическое развитие греческих колоний в Южной Италии было замечательным, и в классические времена этот регион назывался Magna Graecia (Великая Греция), потому что он, вероятно, превосходил по численности греческое население родины». Кавалли-Сфорца Л., Меноцци П., Пьяцца А (1994). История и география человеческих генов . Издательство Принстонского университета. п. 278. ИСБН 978-0-691-08750-4 .
- ^ Jump up to: а б Хаак В., Лазаридис И., Паттерсон Н., Роланд Н., Маллик С., Лламас Б. и др. (июнь 2015 г.). «Массовая миграция из степи явилась источником индоевропейских языков в Европе» . Природа . 522 (7555): 207–11. arXiv : 1502.02783 . Бибкод : 2015Natur.522..207H . дои : 10.1038/nature14317 . ПМК 5048219 . ПМИД 25731166 .
- ^ Jump up to: а б с д Ди Гаэтано С., Воглино Ф., Гуаррера С., Фиорито Дж., Роза Ф., Ди Блазио А.М. и др. (2012). «Обзор генетической структуры итальянской популяции на основе полногеномных данных» . ПЛОС ОДИН . 7 (9): е43759. Бибкод : 2012PLoSO...743759D . дои : 10.1371/journal.pone.0043759 . ПМЦ 3440425 . ПМИД 22984441 .
- ^ Jump up to: а б Прайс А.Л., Батлер Дж., Паттерсон Н., Капелли С., Паскали В.Л., Скарниччи Ф. и др. (январь 2008 г.). «Выявление происхождения американцев европейского происхождения в исследованиях генетических ассоциаций» . ПЛОС Генетика . 4 (1): е236. дои : 10.1371/journal.pgen.0030236 . ПМК 2211542 . ПМИД 18208327 .
- ^ Jump up to: а б Пашу П., Дринеас П., Яннаки Э., Разу А., Канаки К., Цецос Ф. и др. (июнь 2014 г.). «Морской путь колонизации Европы» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (25): 9211–6. Бибкод : 2014PNAS..111.9211P . дои : 10.1073/pnas.1320811111 . ПМК 4078858 . ПМИД 24927591 .
- ^ Jump up to: а б с Фиорито Дж., Ди Гаэтано С., Гуаррера С., Роза Ф., Фельдман М.В., Пьяцца А., Матулло Дж. (июль 2016 г.). «Итальянский геном отражает историю Европы и Средиземноморского бассейна» . Европейский журнал генетики человека . 24 (7): 1056–62. дои : 10.1038/ejhg.2015.233 . ПМК 5070887 . ПМИД 26554880 .
- ^ Антонио и др. 2019 , с. 2.
- ^ Jump up to: а б с Ди Гаэтано С., Черутти Н., Кробу Ф., Робино С., Интурри С., Джино С. и др. (январь 2009 г.). «Дифференциальная миграция греков и северной Африки на Сицилию подтверждается генетическими данными Y-хромосомы» . Европейский журнал генетики человека . 17 (1): 91–9. дои : 10.1038/ejhg.2008.120 . ПМЦ 2985948 . ПМИД 18685561 .
Генетический вклад греческих хромосом в генофонд Сицилии оценивается примерно в 37%, тогда как вклад популяций Северной Африки оценивается примерно в 6%.
- ^ Jump up to: а б с д Сарно С., Боаттини А., Пагани Л., Саццини М., Де Фанти С., Квальариелло А. и др. (май 2017 г.). «Древние и недавние слои примесей на Сицилии и Южной Италии прослеживают многочисленные пути миграции вдоль Средиземноморья» . Научные отчеты . 7 (1): 1984. Бибкод : 2017НацСР...7.1984С . дои : 10.1038/s41598-017-01802-4 . ПМК 5434004 . ПМИД 28512355 .
- «Население восточного побережья Средиземного моря имеет общее генетическое наследие, выходящее за рамки национальности» . Физика.орг . 17 мая 2017 г.
- ^ Антонио и др. 2019 , с. 3.
- ^ Уэйд, Лиззи (2019). «Иммигранты с Ближнего Востока сформировали Рим» . Наука . 366 (6466): 673. Бибкод : 2019Sci...366..673W . дои : 10.1126/science.366.6466.673 . ПМИД 31699914 . S2CID 207965960 . Архивировано из оригинала 24 февраля 2021 г. Проверено 25 января 2021 г. «Торговые маршруты отправляли людей и товары в новую столицу, а эпидемии и вторжения сократили население Рима примерно до 100 000 человек. Вторгающиеся варвары принесли с собой больше европейских предков. Рим постепенно утратил свою прочную генетическую связь с Восточным Средиземноморьем и Ближним Востоком. К средневековым временам , жители города снова генетически напоминали европейское население».
- ^ ..."Отделение Сардинии от остального континента, да и вообще от всех других европейских популяций, что, вероятно, указывает на более древнее происхождение ее населения, независимое от италийского населения и имеющее происхождение на Ближнем Востоке. Средиземноморье». Альберто Пьяцца , Генетические профили итальянцев , Академия наук Турина.
- ^ Jump up to: а б с д Бризигелли Ф., Альварес-Иглесиас В., Фондевила М., Бланко-Вереа А., Карраседо А., Паскали В.Л. и др. (2012). «Однородительские маркеры современного итальянского населения раскрывают подробности его доримского наследия» . ПЛОС ОДИН . 7 (12): e50794. Бибкод : 2012PLoSO...750794B . дои : 10.1371/journal.pone.0050794 . ПМК 3519480 . ПМИД 23251386 .
- ^ Jump up to: а б Саццини М., Гнекки Русконе Г.А., Джулиани С., Сарно С., Квальариелло А., Де Фанти С. и др. (сентябрь 2016 г.). «Сложное взаимодействие между нейтральной и адаптивной эволюцией сформировало дифференциальный геномный фон и восприимчивость к болезням на итальянском полуострове» . Научные отчеты . 6 : 32513. Бибкод : 2016NatSR...632513S . дои : 10.1038/srep32513 . ПМК 5007512 . ПМИД 27582244 . Зендо: 165505 .
- ^ Эско Т., Меззавилла М., Нелис М., Борель С., Дебняк Т., Яккула Е. и др. (июнь 2013 г.). «Генетическая характеристика изолятов северо-восточной Италии в контексте более широкого европейского генетического разнообразия» . Европейский журнал генетики человека . 21 (6): 659–65. дои : 10.1038/ejhg.2012.229 . ПМЦ 3658181 . ПМИД 23249956 .
- ^ Jump up to: а б Нелис М., Эско Т., Мяги Р., Зимприч Ф., Зимприч А., Тончева Д. и др. (2009). «Генетическая структура европейцев: взгляд с Северо-Востока» . ПЛОС ОДИН . 4 (5): е5472. Бибкод : 2009PLoSO...4.5472N . дои : 10.1371/journal.pone.0005472 . ПМК 2675054 . ПМИД 19424496 .
- ^ Саццини М., Абондио П., Сарно С., Гнекки-Русконе Г.А., Рагно М., Джулиани С. и др. (май 2020 г.). «Геномная история итальянского населения повторяет ключевую эволюционную динамику как континентальных, так и южных европейцев» . БМК Биология . 18 (1): 51. дои : 10.1186/s12915-020-00778-4 . ПМЦ 7243322 . ПМИД 32438927 .
- ^ Руци С. (декабрь 2006 г.). «Поток доисторических генов гаплогруппы I Y-хромосомы в Европе» (PDF) . Документа Преисторика . 33 : 17–20. дои : 10.4312/dp.33.3 . Архивировано из оригинала (PDF) 6 марта 2009 г.
- ^ «Недавние культуры бронзового века в северной Италии и их взаимоотношения с «культурой урнового поля» » . Археологическая группа Монтекларансе. Архивировано из оригинала 10 мая 2006 года.
- ^ Хейвуд Дж (2014). Кельты: от бронзового века до нового века . Рутледж. п. 21. ISBN 978-1-317-87017-3 .
- ^ «Ломбардский народ» . Британская энциклопедия .
- ^ Крон, Джефф (2013). «Конкретизация демографии Этрурии». В Macintosh Turfa, Джин (ред.). Этрусский мир . Лондон; Нью-Йорк: Рутледж. стр. 56–78. ISBN 978-0-415-67308-2 .
- ^ Jump up to: а б Перкинс, Фил (2017). «ДНК и этрусская идентичность». В Насо, Алессандро (ред.). Этрускология . Берлин: Де Грюйтер. стр. 109–18. ISBN 978-1-934078-49-5 .
- ^ Jump up to: а б Гиротто, Сильвия; Тасси, Франческа; Фумагалли, Эрика; Колонна, Винченца; Сандиониги, Анна; Лари, Мартина; и др. (2013). «Происхождение и эволюция мтДНК этрусков» . ПЛОС ОДИН . 8 (2): e55519. Бибкод : 2013PLoSO...855519G . дои : 10.1371/journal.pone.0055519 . ПМК 3566088 . ПМИД 23405165 .
- ^ Jump up to: а б Тасси, Франческа; Гиротто, Сильвия; Карамелли, Дэвид; Барбуджани, Гвидо (2013). «Генетические данные не подтверждают этрусское происхождение в Анатолии». Американский журнал физической антропологии . 152 (1): 11–18. дои : 10.1002/ajpa.22319 . ПМИД 23900768 .
- ^ Вернеси С., Карамелли Д., Дюпанлуп И. и др. (апрель 2004 г.). «Этруски: популяционно-генетическое исследование» . Являюсь. Дж. Хум. Жене . 74 (4): 694–704. дои : 10.1086/383284 . ПМК 1181945 . ПМИД 15015132 .
- ^ Бандельт HJ (ноябрь 2004 г.). «Этрусские артефакты» . Являюсь. Дж. Хум. Жене . 75 (5): 919–20, ответ автора 923–7. дои : 10.1086/425180 . ПМЦ 1182123 . ПМИД 15457405 .
- ^ Матейу Л.М., Раннала Б.Х. (2008). «Байесовский вывод об ошибках в древней ДНК, вызванных посмертной деградацией» . Мол Биол Эвол . 25 (7): 1503–11. дои : 10.1093/molbev/msn095 . ПМИД 18420593 .
- ^ Jump up to: а б Леонарди, Микела; Сандиониги, Анна; Конзато, Анналиса; Вай, Стефания; Лари, Мартина (2018). «Рассказ предка женского пола: длительная материнская преемственность в неизолированном регионе Тосканы» . Американский журнал физической антропологии . 167 (3). Нью-Йорк: John Wiley & Sons (опубликовано 6 сентября 2018 г.): 497–506. дои : 10.1002/ajpa.23679 . ПМИД 30187463 . S2CID 52161000 .
- ^ Jump up to: а б с Антонио, Маргарет Л.; Гао, Зиюэ; Мутс, Ханна М.; и др. (ноябрь 2019 г.). «Древний Рим: генетический перекресток Европы и Средиземноморья» . Наука . 366 (6466). Вашингтон, округ Колумбия: Американская ассоциация содействия развитию науки (опубликовано 8 ноября 2019 г.): 708–714. Бибкод : 2019Sci...366..708A . дои : 10.1126/science.aay6826 . hdl : 2318/1715466 . ПМК 7093155 . ПМИД 31699931 .
Интересно, что, хотя люди железного века были отобраны как из этрусского (n=3), так и из латинского (n=6) контекстов, мы не обнаружили каких-либо существенных различий между двумя группами со статистикой f4 в форме f4(RMPR_Etruscan, RMPR_Latin; test популяция, Онге), что позволяет предположить общее происхождение или обширный генетический обмен между ними.
- ^ Антонио и др. 2019 , с. 3.
- ^ Антонио и др. 2019 , с. 2.
- ^ Антонио и др. 2019 г. , Таблица 2. Пример информации, строки 33–35.
- ^ Пост, Козимо; Заро, Валентина; Спиру, Мария А. (24 сентября 2021 г.). «Происхождение и наследие этрусков через 2000-летний археогеномный разрез времени» . Достижения науки . 7 (39). Вашингтон, округ Колумбия: Американская ассоциация содействия развитию науки: eabi7673. Бибкод : 2021SciA....7.7673P . дои : 10.1126/sciadv.abi7673 . ПМЦ 8462907 . ПМИД 34559560 .
- ^ Баньяско, Г.; Марзулло, М.; Каттанео, К.; Билер-Гомез, Л.; Маццарелли, Д.; Риккарди, В.; Мюллер, В.; Коппа, А.; Маклафлин, Р.; Мотта, Л.; Прато, О.; Шмидт, Ф.; Гаверио, Ф.; Маррас, Великобритания; Миллет, Массачусетс (28 мая 2024 г.). «Биоархеология помогает культурному пониманию шести персонажей, ищущих свою деятельность (Тарквиния, девятый-седьмой век до нашей эры, центральная Италия)» . Научные отчеты . 14 (1): 11895. Бибкод : 2024NatSR..1411895B . дои : 10.1038/s41598-024-61052-z . ISSN 2045-2322 . ПМЦ 11133411 . ПМИД 38806487 .
- ^ Ахилли А., Оливьери А., Пала М. и др. (апрель 2007 г.). «Вариации митохондриальной ДНК современных тосканцев подтверждают ближневосточное происхождение этрусков» . Являюсь. Дж. Хум. Жене . 80 (4): 759–68. дои : 10.1086/512822 . ПМЦ 1852723 . ПМИД 17357081 .
- ^ Уайтхед, Джейн К. (2007). «ДНК и этническое происхождение: возможное и невероятное». Этрусские новости (8). Нью-Йорк: Американское отделение Института этрусских и курсивных исследований.
- ^ Гандини, Франческа (2016). «Картирование расселения людей на Африканском Роге из рефугиумов Аравийского ледникового периода с использованием митогеномов» . Научные отчеты . 6 (25472): 25472. Бибкод : 2016NatSR...625472G . дои : 10.1038/srep25472 . ПМЦ 4857117 . ПМИД 27146119 .
- ^ Груни, Виола (2018). «Реконструкция генетической истории итальянцев: новые идеи с точки зрения мужчины (Y-хромосомы)». Анналы биологии человека . 45 (1) (опубликовано 30 января 2018 г.): 44–56. дои : 10.1080/03014460.2017.1409801 . ПМИД 29382284 . S2CID 43501209 .
Фактически, хотя наличие J2a-M67* предполагает контакты по морю с анатолийцами, в соответствии с гипотезой Геродота о внешнем анатолийском источнике этрусков, обнаружение центральноевропейской линии G2a-L497 со значительной частотой может скорее поддерживают североевропейское происхождение этрусков. С другой стороны, высокая распространенность европейских линий R1b не может исключить сценарий автохтонного процесса формирования этрусской цивилизации из предшествующего виллановского общества, как впервые предположил Дионисий Галикарнасский; подробный анализ гаплогруппы R1b-U152 может оказаться очень информативным в этом отношении.
- ^ Бризигелли, Франческа (2012). «Опородительные маркеры современного итальянского населения раскрывают подробности его доримского наследия» . ПЛОС ОДИН . 7 (12). Публичная научная библиотека (опубликовано 10 декабря 2012 г.): e50794. Бибкод : 2012PLoSO...750794B . дои : 10.1371/journal.pone.0050794 . ПМК 3519480 . ПМИД 23251386 .
- ^ Jump up to: а б Боаттини, Алессио (2013). «Маркеры однородительского поведения в Италии выявляют генетическую структуру с предвзятостью по признаку пола и различные исторические слои» . ПЛОС ОДИН . 8 (5). Публичная научная библиотека (опубликовано 29 мая 2013 г.): e65441. Бибкод : 2013PLoSO...865441B . дои : 10.1371/journal.pone.0065441 . ПМК 3666984 . ПМИД 23734255 .
- ^ Гамба, Кристина; Джонс, Эппи Р.; Тисдейл, Мэтью Д. (2014). «Поток и застой генома в пятитысячелетнем разрезе европейской предыстории» . Природные коммуникации . 5 (5257). Лондон: Nature Publishing Group (опубликовано 21 октября 2014 г.): 5257. Бибкод : 2014NatCo...5.5257G . дои : 10.1038/ncomms6257 . ПМК 4218962 . ПМИД 25334030 .
- ^ Jump up to: а б Мэтисон, Иэн (2018). «Геномная история Юго-Восточной Европы» . Природа . 555 (7695). Лондон: Nature Publishing Group (опубликовано 21 февраля 2018 г.): 197–203. Бибкод : 2018Natur.555..197M . дои : 10.1038/nature25778 . ПМК 6091220 . ПМИД 29466330 .
- ^ Липсон, Марк (2017). «Параллельные палеогеномные разрезы раскрывают сложную генетическую историю ранних европейских земледельцев» . Природа . 551 (7680). Лондон: Nature Publishing Group (опубликовано 8 ноября 2017 г.): 368–372. Бибкод : 2017Natur.551..368L . дои : 10.1038/nature24476 . ПМК 5973800 . ПМИД 29144465 .
- ^ Саупе, Тина; Монтинаро, Франческо; Скэджион, Чинция; Каррара, Никола; Кивисилд, Тоомас; Д'Атанасио, Евгения; Хуэй, Жоюнь; Сольник, Ану; Лебрассер, Офелия; Ларсон, Грегер; Алессандри, Лука (21 июня 2021 г.). «Древние геномы обнаруживают структурные сдвиги после прибытия степных предков на Итальянский полуостров» . Современная биология . 31 (12): 2576–2591.e12. Бибкод : 2021CBio...31E2576S . дои : 10.1016/j.cub.2021.04.022 . hdl : 11585/827581 . ISSN 0960-9822 . ПМИД 33974848 . S2CID 234471370 .
Гротта Ла Сасса (Национальный кадастр пещер, идентификатор: LA 2001) был обнаружен в 2015 году во время исследования природных пещер гор Аузони, проведенного двумя спелеологическими группами: Gruppo Grotte Castelli Romani и Speleo Club Roma. (...) В Ла-Сасса два самца LSC002/004 и LSC011 имеют идентичный гаплотип Ychr (J2a-M410/J2a7-Z2397; Таблица 1; Данные S1B и S1F).
- ^ Саццини, М.; Гнекки Русконе, Г.; Джулиани, К. (2016). «Сложное взаимодействие между нейтральной и адаптивной эволюцией сформировало дифференциальный геномный фон и восприимчивость к болезням на итальянском полуострове» . Научные отчеты . 6 (32513): 32513. Бибкод : 2016NatSR...632513S . дои : 10.1038/srep32513 . ПМК 5007512 . ПМИД 27582244 .
- ^ Равеан, Марко (2019). «Структура населения современных итальянцев раскрывает образцы древнего и архаичного происхождения в Южной Европе» . Достижения науки . 5 (9): eaaw3492. Бибкод : 2019SciA....5.3492R . дои : 10.1126/sciadv.aaw3492 . ПМК 6726452 . ПМИД 31517044 .
- ^ Краузе, Джон ; Траппе, Томас (2021) [2019]. Краткая история человечества: Новая история старой Европы [ Путешествие наших генов: история о нас и наших предках ]. Перевод Уэйт, Кэролайн (я ред.). Нью-Йорк: Рэндом Хаус. п. 217. ИСБН 9780593229422 .
Вполне вероятно, что баскский, палео-сардинский, минойский и этрусский языки развились на континенте в ходе неолитической революции. К сожалению, истинное разнообразие языков, когда-то существовавших в Европе, никогда не будет известно.
- ^ Jump up to: а б с Гругни В., Равеан А., Маттиоли Ф., Батталья В., Сала С., Тониоло Д. и др. (февраль 2018 г.). «Реконструкция генетической истории итальянцев: новые идеи с точки зрения мужчины (Y-хромосомы)». Анналы биологии человека . 45 (1): 44–56. дои : 10.1080/03014460.2017.1409801 . ПМИД 29382284 . S2CID 43501209 .
- ^ Ди Джакомо Ф., Лука Ф., Ананью Н., Чаварелла Дж., Корбо Р.М., Креста М. и др. (сентябрь 2003 г.). «Клинические закономерности разнообразия Y-хромосомы человека в континентальной Италии и Греции определяются эффектами дрейфа и основателя» (PDF) . Молекулярная филогенетика и эволюция . 28 (3): 387–95. дои : 10.1016/s1055-7903(03)00016-2 . ПМИД 12927125 . Архивировано из оригинала (PDF) 20 января 2017 г.
- ^ Франкалаччи, П.; Морелли, Л.; Андерхилл, Пенсильвания; Лилли, А.С.; Пассарино, Г.; Усели, А.; Мадедду, Р.; Паоли, Г.; Тофанелли, С.; Кало, CM; Гиани, Мэн; Вареси, Л.; Мемми, М.; Вона, Г.; Лин, А.А. (июль 2003 г.). «Население трех средиземноморских островов (Корсика, Сардиния и Сицилия) на основе биаллельной изменчивости Y-хромосомы» . Американский журнал физической антропологии . 121 (3): 270–279. дои : 10.1002/ajpa.10265 . ISSN 0002-9483 . ПМИД 12772214 .
- ^ Франкалаччи, Паоло; Морелли, Лаура; Ангиус, Андреа; Берутти, Риккардо; Рейнье, Фредерик; Ацени, Россано; Пилу, Розелла; Бусонеро, Фабио; Мале, Андреа; Зара, Иления; Санна, Дарья; Усели, Антонелла; Урру, Мария Франческа; Марчелли, Марко; Кусано, Роберто (2 августа 2013 г.). «Низкопроходное секвенирование ДНК 1200 сардинцев реконструирует европейскую филогению Y-хромосомы» . Наука . 341 (6145): 565–569. Бибкод : 2013Sci...341..565F . дои : 10.1126/science.1237947 . ISSN 0036-8075 . ПМК 5500864 . ПМИД 23908240 .
- ^ Jump up to: а б Капелли С., Бризигелли Ф., Скарниччи Ф., Арреди Б., Калья А., Ветруньо Г. и др. (июль 2007 г.). «Генетическая вариация Y-хромосомы на итальянском полуострове является клинальной и подтверждает модель примеси для встречи мезолита и неолита». Молекулярная филогенетика и эволюция . 44 (1): 228–39. дои : 10.1016/j.ympev.2006.11.030 . ПМИД 17275346 .
- ^ Кавалли-Сфорца Л., Меноцци П., Пьяцца А (1994). История и география человеческих генов . Издательство Принстонского университета. ISBN 978-0-691-08750-4 . > : 295
- ^ Семино О., Магри С., Бенуцци Г., Лин А.А., Аль-Захери Н., Батталья В. и др. (май 2004 г.). «Происхождение, распространение и дифференциация гаплогрупп E и J Y-хромосомы: выводы о неолитизации Европы и более поздних миграционных событиях в Средиземноморье» . Американский журнал генетики человека . 74 (5): 1023–34. дои : 10.1086/386295 . ПМК 1181965 . ПМИД 15069642 .
- ^ Баркер, Грэм ; Расмуссен, Том (2000). Этруски . Народы Европы. Оксфорд: Издательство Блэквелл. п. 44. ИСБН 978-0-631-22038-1 .
- ^ Турфа, Джин Макинтош (2017). «Этруски». В Фарни, Гэри Д.; Брэдли, Гэри (ред.). Народы Древней Италии . Берлин: Де Грюйтер. стр. 637–672. дои : 10.1515/9781614513001 . ISBN 978-1-61451-520-3 .
- ^ Де Граммонд, Нэнси Т. (2014). «Этнос и этруски». В Макинерни, Джереми (ред.). Спутник этнической принадлежности в Древнем Средиземноморье . Чичестер, Великобритания: John Wiley & Sons, Inc., стр. 405–422. дои : 10.1002/9781118834312 . ISBN 9781444337341 .
- ^ Шипли, Люси (2017). «Где дом?». Этруски: утраченные цивилизации . Лондон: Книги реакции. стр. 28–46. ISBN 9781780238623 .
- ^ Jump up to: а б Мессина Ф., Финоккио А., Рольфо М.Ф., Де Анжелис Ф., Рапоне С., Колетта М. и др. (2015). «Следы забытых исторических событий в горных общинах Центральной Италии: генетическое понимание» . Американский журнал биологии человека . 27 (4): 508–19. дои : 10.1002/ajhb.22677 . ПМИД 25728801 . S2CID 30111156 .
- ^ Капелли С., Онофри В., Бризигелли Ф., Боски И., Скарниччи Ф., Масулло М. и др. (июнь 2009 г.). «Мавры и сарацины в Европе: оценка средневекового мужского наследия Северной Африки в южной Европе» . Европейский журнал генетики человека . 17 (6): 848–52. дои : 10.1038/ejhg.2008.258 . ПМЦ 2947089 . ПМИД 19156170 .
- ^ Jump up to: а б с д Сарно С., Боаттини А., Карта М., Ферри Г., Алу М., Яо Д.Ю. и др. (2014). «Древний средиземноморский плавильный котел: исследование однородительской генетической структуры и истории населения Сицилии и южной Италии» . ПЛОС ОДИН . 9 (4): е96074. Бибкод : 2014PLoSO...996074S . дои : 10.1371/journal.pone.0096074 . ПМЦ 4005757 . ПМИД 24788788 .
 Эта статья содержит цитаты из этого источника, который доступен по лицензии Creative Commons Attribution 4.0 International (CC BY 4.0) .
Эта статья содержит цитаты из этого источника, который доступен по лицензии Creative Commons Attribution 4.0 International (CC BY 4.0) . - ^ Бризигелли, Ф (2012). «Однородительские маркеры современного итальянского населения раскрывают подробности его доримского наследия» . ПЛОС ОДИН . 7 (12): e50794. Бибкод : 2012PLoSO...750794B . дои : 10.1371/journal.pone.0050794 . ПМК 3519480 . ПМИД 23251386 . В таблице S4 гаплотип #BEL50 оценивается как Q-M378, хотя он показан как P* (xR1).
- ^ Jump up to: а б Перейра Л., Ричардс М., Гойос А., Алонсо А., Альбарран С., Гарсия О. и др. (январь 2005 г.). «Свидетельства мтДНК высокого разрешения о позднеледниковом переселении Европы из иберийского рефугиума» . Геномные исследования . 15 (1): 19–24. дои : 10.1101/гр.3182305 . ПМК 540273 . ПМИД 15632086 .
- ^ Ричардс М., Маколей В., Хики Е., Вега Е., Сайкс Б., Гуида В. и др. (ноябрь 2000 г.). «Отслеживание линий европейских основателей в пуле мтДНК Ближнего Востока» . Американский журнал генетики человека . 67 (5): 1251–76. дои : 10.1016/S0002-9297(07)62954-1 . ПМК 1288566 . ПМИД 11032788 .
- ^ Ахилли А., Ренго С., Магри С., Батталья В., Оливьери А., Скоццари Р. и др. (ноябрь 2004 г.). «Молекулярное исследование гаплогруппы H мтДНК подтверждает, что франко-кантабрийское ледниковое убежище было основным источником европейского генофонда» . Американский журнал генетики человека . 75 (5): 910–8. дои : 10.1086/425590 . ПМЦ 1182122 . ПМИД 15382008 .
- ^ Ахилли А., Оливьери А., Пала М., Метспалу Э., Форнарино С., Батталья В. и др. (апрель 2007 г.). «Вариации митохондриальной ДНК современных тосканцев подтверждают ближневосточное происхождение этрусков» . Американский журнал генетики человека . 80 (4): 759–68. дои : 10.1086/512822 . ПМЦ 1852723 . ПМИД 17357081 .
4/138=2,9% в Лациуме; 3/114=2,6% в Вольтерре; 2/92=2,2% в Базиликате и 3/154=2,0% на Сицилии.
- ^ Боаттини А., Мартинес-Круз Б., Сарно С., Хармант С., Усели А., Санс П. и др. (2013). «Маркеры однородителей в Италии выявляют генетическую структуру с предвзятостью по признаку пола и различные исторические слои» . ПЛОС ОДИН . 8 (5): e65441. Бибкод : 2013PLoSO...865441B . дои : 10.1371/journal.pone.0065441 . ПМК 3666984 . ПМИД 23734255 .
- ^ Балтер М. (3 июня 2010 г.). «В поисках корней еврейства» . Научный журнал . Американская ассоциация содействия развитию науки.
- ^ Зооссманн-Дискин А (октябрь 2010 г.). «Происхождение восточноевропейских евреев выявлено по аутосомному, половому хромосомному полиморфизму и полиморфизму мтДНК» . Биология Директ . 5 (57): 57. дои : 10.1186/1745-6150-5-57 . ПМЦ 2964539 . ПМИД 20925954 .
- ^ Балтер М. (8 октября 2013 г.). «Происходили ли современные евреи в Италии?» . Наука СЕЙЧАС .
- ^ Янделл К. (2013). «Генетические корни евреев-ашкенази» . Ученый .
- ^ Розенберг Н.А., Притчард Дж.К., Вебер Дж.Л., Канн Х.М., Кидд К.К., Животовский Л.А., Фельдман М.В. (декабрь 2002 г.). «Генетическая структура человеческих популяций». Наука . 298 (5602): 2381–5. Бибкод : 2002Sci...298.2381R . дои : 10.1126/science.1078311 . ПМИД 12493913 . S2CID 8127224 .
- ^ Коста М.Д., Перейра Ж.Б., Пала М., Фернандес В., Оливьери А., Ахилли А. и др. (2013). «Существенное доисторическое европейское происхождение среди материнских линий ашкенази» . Природные коммуникации . 4 : 2543. Бибкод : 2013NatCo...4.2543C . дои : 10.1038/ncomms3543 . ПМЦ 3806353 . ПМИД 24104924 .
- ^ Банда И., Квале М., Хоффманн Т., Хессельсон С., Тан Х., Ранатунга Д. и др. (2013). «Оценка примеси в популяции основателей» . Am Soc Hum Genet . Архивировано из оригинала 11 августа 2019 г. Проверено 31 октября 2016 г.
- ^ Брей С.М., Мулле Дж.Г., Додд А.Ф., Пулвер А.Е., Вудинг С., Уоррен С.Т. (сентябрь 2010 г.). «Признаки влияния основателей, примеси и отбора в еврейском населении ашкенази» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (37): 16222–7. Бибкод : 2010PNAS..10716222B . дои : 10.1073/pnas.1004381107 . ПМЦ 2941333 . ПМИД 20798349 .
- ^ Ацмон Г., Хао Л., Пеер И., Велес С., Перлман А., Паламара П.Ф., Морроу Б., Фридман Е., Одду С., Бернс Е., Острер Х. (июнь 2010 г.). «Дети Авраама в эпоху генома: основные популяции еврейской диаспоры составляют отдельные генетические кластеры с общим ближневосточным происхождением» . Американский журнал генетики человека . 86 (6): 850–9. дои : 10.1016/j.ajhg.2010.04.015 . ПМК 3032072 . ПМИД 20560205 .
- ^ «Гены отличают евреев, результаты исследования» . Американский учёный . Проверено 8 ноября 2013 г.
- «Исследование обнаруживает генетические связи среди еврейского народа» . ЭврекАлерт! (Пресс-релиз). 3 июня 2010 г.
- ^ Пала М., Ахилли А., Оливьери А., Хушиар Кашани Б., Перего У.А., Санна Д. и др. (июнь 2009 г.). «Митохондриальная гаплогруппа U5b3: отдаленное эхо эпипалеолита в Италии и наследие ранних сардинцев» (PDF) . Американский журнал генетики человека . 84 (6): 814–21. дои : 10.1016/j.ajhg.2009.05.004 . ПМК 2694970 . ПМИД 19500771 . Архивировано из оригинала (PDF) 22 июля 2018 г. Проверено 15 июля 2010 г.
- ^ Леонарди, Микела; Сандиониги, Анна; Конзато, Анналиса; Вай, Стефания; Лари, Мартина (2018). «Рассказ предка женского пола: длительная материнская преемственность в неизолированном регионе Тосканы» . Американский журнал физической антропологии . 167 (3). Нью-Йорк: John Wiley & Sons (опубликовано 6 сентября 2018 г.): 497–506. дои : 10.1002/ajpa.23679 . ПМИД 30187463 . S2CID 52161000 .
- ^ Ковачевич Л., Тамбетс К., Илумяэ А.М., Кушниаревич А., Юнусбаев Б., Сольник А. и др. (2014). «Стоим у ворот в Европу: генетическая структура населения Западных Балкан, основанная на аутосомных и гаплоидных маркерах» . ПЛОС ОДИН . 9 (8): e105090. Бибкод : 2014PLoSO...9j5090K . дои : 10.1371/journal.pone.0105090 . ПМК 4141785 . ПМИД 25148043 .
- ^ Уэйд Н. (13 августа 2008 г.). «Генетическая карта Европы» . Нью-Йорк Таймс . Проверено 25 февраля 2014 г.
- ^ Ли Дж. З., Абшер Д. М., Тан Х, Саутвик А. М., Касто А. М., Рамачандран С., Канн Х. М., Барш Г. С., Фельдман М., Кавалли-Сфорца Л. Л., Майерс Р. М. (февраль 2008 г.). «Человеческие отношения во всем мире, выведенные на основе общегеномных вариаций» (PDF) . Наука . 319 (5866). Нью-Йорк, штат Нью-Йорк: 11:00–4. Бибкод : 2008Sci...319.1100L . дои : 10.1126/science.1153717 . ПМИД 18292342 . S2CID 53541133 . Архивировано из оригинала (PDF) 27 марта 2009 г.
- ^ Лопес Эрраес Д., Боше М., Тан К., Теунерт С., Пугач И., Ли Дж. и др. (ноябрь 2009 г.). «Генетическая изменчивость и недавний положительный отбор в человеческих популяциях по всему миру: данные почти 1 миллиона SNP» . ПЛОС ОДИН . 4 (11): е7888. Бибкод : 2009PLoSO...4.7888L . дои : 10.1371/journal.pone.0007888 . ПМЦ 2775638 . ПМИД 19924308 .
- ^ Мурджани П., Паттерсон Н., Хиршхорн Дж.Н., Кейнан А., Хао Л., Ацмон Г., Бернс Э., Острер Х., Прайс А.Л., Райх Д. (апрель 2011 г.). «История потока африканских генов в южных европейцев, левантийцев и евреев» . ПЛОС Генетика . 7 (4): e1001373. дои : 10.1371/journal.pgen.1001373 . ПМК 3080861 . ПМИД 21533020 .
- ^ Келлер А., Грефен А., Болл М., Мацас М., Буагерен В., Майкснер Ф. и др. (февраль 2012 г.). «Новое понимание происхождения и фенотипа тирольского ледяного человека, полученное с помощью полногеномного секвенирования» . Природные коммуникации . 3 : 698. Бибкод : 2012NatCo...3..698K . дои : 10.1038/ncomms1701 . ПМИД 22426219 .
- ^ Равеан, Александр; Молинаро, Людовика; Анели, Серена; Каподиферро, Марко Росарио; Онгаро, Линда; Лучший, Никола Рамбальди; Взорвите себя, Сара; Скарано, Теодоро; Торрони, Антонио; Ахилли, Алессандро; Вентура, Марио; Пагани, Лука; Волосы, Кристиан; Оливьери, Анна; Бертолини, Франческо (01 марта 2022 г.). «Оценка временных и географических контактов через Адриатическое море посредством анализа полногеномных данных из Южной Италии» : 2022.02.26.482072. дои : 10.1101/2022.02.26.482072 . S2CID 247231413 .
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь )
Дальнейшее чтение
[ редактировать ]- Саупе, Тина и др. «Древние геномы обнаруживают структурные сдвиги после прибытия степных предков на Итальянский полуостров». В: Current Biology , том 31, выпуск 12, 21 июня 2021 г., страницы 2576–2591.e12. DOI: https://doi.org/10.1016/j.cub.2021.04.022.