Западный охотник-собиратель
Генетическое происхождение охотников-собирателей в Европе между 14 и 9 тыс. лет назад, основная территория западных охотников-собирателей (WHG) выделена синим цветом. Отдельные номера соответствуют датам калиброванной выборки. [1] |

В археогенетике , западный охотник-собиратель ( WHG , также известный как западноевропейский охотник-собиратель . западноевропейский охотник-собиратель или кластер Оберкасселя ) ( ок. 15 000–5 000 лет назад ) является отчетливым наследственным компонентом современных европейцев, представляющим происхождение от населения мезолитических рассеявшихся охотников-собирателей, по западной , южной и центральной Европе , от Британских островов на западе до Карпат на востоке, после отступления ледникового покрова Последнего ледникового максимума . [2] Оно тесно связано и иногда считается синонимом концепции кластера Виллабруна , названного в честь пещеры Рипари Виллабруна в Италии, известной из конечного плейстоцена Европы, которая во многом является предком более поздних популяций WHG.
WHG имеют более близкое генетическое родство с древними и современными народами Ближнего Востока и Кавказа, чем более ранние европейские охотники-собиратели. Считается, что WHG мигрировали в Европу примерно в конце последнего ледникового максимума , распространившись по Западной Европе в конце плейстоцена, в значительной степени вытеснив более ранние кроманьонцев группы , такие как мадленцы .
Наряду со скандинавскими охотниками-собирателями (SHG) и восточными охотниками-собирателями (EHG), WHG составляли одну из трех основных генетических групп в послеледниковый период ранней голоценовой Европы. [3] Граница между WHG и EHG проходила примерно от нижнего Дуная на север вдоль западных лесов Днепра к западной части Балтийского моря . [2] EHG в основном состояли из смеси предков, связанных с WHG, и древней Северной Евразии (ANE), [4] в то время как ГСП представляли собой смесь WHG и EHG. [3] На Пиренейском полуострове охотники-собиратели раннего голоцена состояли из смеси WHG и мадленских кроманьонцев (GoyetQ2). [5]
Когда-то WHG составляли основную популяцию по всей Европе, они были в значительной степени заменены последовательными экспансиями ранних европейских фермеров (EEF) анатолийского происхождения во время раннего неолита , которые обычно несли незначительное количество предков WHG из-за смешения с группами WHG во время их европейской экспансии. Среди современного населения происхождение WHG наиболее распространено среди населения восточно-Балтийского региона. [6]
Исследовать
[ редактировать ]Западные охотники-собиратели (WHG) признаны отдельным предковым компонентом, вносящим вклад в происхождение большинства современных европейцев . [7] Большинство европейцев можно смоделировать как смесь WHG, EEF и WSH из Понтийско-Каспийской степи . [8] WHG также внесли свой вклад в происхождение других древних групп, таких как ранние европейские фермеры (EEF), которые, однако, были в основном выходцами из Анатолии. [7] С распространением неолита EEF стал доминировать в генофонде в большинстве частей Европы, хотя родословная WHG возродилась в Западной Европе от раннего неолита до среднего неолита. [9]
Происхождение и распространение в континентальную Европу
[ редактировать ]WHGs представляют собой крупный сдвиг населения в Европе в конце ледникового периода, вероятно, расширение населения в континентальную Европу из рефугиумов Юго-Восточной Европы или Западной Азии . [10] Считается, что их предки отделились от восточных евразийцев около 40 000 лет назад, а от древних северных евразийцев (ANE) - до 24 000 лет назад (предполагаемый возраст мальтийского мальчика ). Впоследствии эта дата была перенесена еще дальше во времени благодаря находкам на стоянке рогов носорога Яна , примерно на 38 тыс. лет назад, вскоре после расхождения западно-евразийских и восточно-евразийских линий. [7] [11] Валлини и др. 2022 утверждает, что рассредоточение и раскол западноевразийских линий произошли не ранее ок. 38 000 лет назад, причем более древние европейские экземпляры начального верхнего палеолита , такие как Златы Кун , Пештера ку Оасе и Бачо Киро , не были связаны с западными охотниками-собирателями, но были ближе к древневосточным евразийцам или были базальными для обоих. [12]
Генетические данные свидетельствуют о том, что предки популяции WHG генетически разошлись с охотниками-собирателями на Ближнем Востоке и на Кавказе около 26 000 лет назад, во время последнего ледникового максимума . [13] Геномы WHG демонстрируют более высокую близость к древним и современным популяциям Ближнего Востока по сравнению с европейцами раннего палеолита, такими как граветты . Близость к древним популяциям Ближнего Востока в Европе увеличилась после последнего ледникового максимума , что коррелирует с расширением происхождения WHG (Виллабруна или Оберкассель). Есть также свидетельства двунаправленного потока генов между популяциями WHG и Ближнего Востока еще 15 000 лет назад. Связанные с WHG останки принадлежали в первую очередь Y-хромосомы человеческим гаплогруппам I-M170 с более низкой частотой C-F3393 (в частности, кладе C-V20/C1a2 ), которые обычно обнаруживались среди более ранних палеолитических европейских останков, таких как Костенки-14 и Сунгирь . Отцовскую гаплогруппу C-V20 до сих пор можно обнаружить у мужчин, живущих в современной Испании , что свидетельствует о давнем присутствии этой линии в Западной Европе. Их митохондриальные хромосомы принадлежали преимущественно к гаплогруппе U5 . [4] [14]
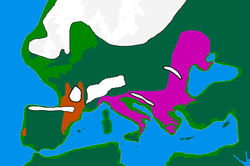
Самые ранние известные в Европе особи преимущественно WHG/виллабрунского происхождения известны из Италии и датируются примерно 17 000 лет назад. [15] хотя человек из пещеры Эль-Мирон на севере Испании, имеющий 43% предков Виллабруна, известен 19 000 лет назад. [4] Ранние популяции WHG/Виллабруна связаны с эпиграветтской археологической культурой, которая в значительной степени заменила популяции, связанные с мадленской культурой, около 14 000 лет назад (предками мадленских особей были популяции, связанные с западными граветтскими , солютрейскими и ориньякскими культурами). [4] [16] Родословная WHG впервые обнаружена к северу от Альп у двух особей возрастом 14 000 лет в Оберкасселе , Германия, которых можно смоделировать как смесь предков Виллабруна (которая сама моделируется как смесь линии, связанной с кластером Вестонице , и линией, предковой для Костенки-14 и особи Goyet Q116-1 ), а происхождение Goyet-Q2 связано с особями, обнаруженными в Европе до последнего ледникового максимума. В исследовании говорится, что всех особей кластера Оберкассель можно смоделировать как c. 75% Виллабруна и 25% происхождение Гойе-Q2 или, альтернативно, как c. 90% предков Виллабруна и 10% предков Фурнола, недавно идентифицированного кластера, описанного как сестринская линия предков Гойета Q116-1, обнаруженного у людей, связанных с граветтской культурой юго-западной Европы. [4] Исследование предполагает, что родословная Оберкасселя в основном уже сформировалась до того, как распространилась, возможно, вокруг западной стороны Альп, в Западную и Центральную Европу и Великобританию, где отобранные особи WHG генетически однородны. Это контрастирует с прибытием предков Виллабруны и Оберкасселя в Иберию, которое, по-видимому, включало в себя повторяющиеся события примеси с местным населением, несущим высокий уровень предков Goyet-Q2. Это, а также выживание специфических клад гаплогруппы C1 Y-ДНК, ранее наблюдавшееся среди ранних европейских охотников-собирателей, предполагает относительно более высокую генетическую преемственность на юго-западе Европы в этот период. [4]
Есть признаки того, что WHG несет в себе «аллели риска диабета и болезни Альцгеймера ». [17]
Взаимодействие с другими популяциями
[ редактировать ]
Также было обнаружено, что WHG внесли свой вклад в происхождение популяций на границах Европы, таких как ранние анатолийские фермеры и древние африканцы Северо-Запада, [19] а также другие европейские группы, такие как восточные охотники-собиратели . [20] Отношения WHG с EHG остаются неубедительными. [20] Смоделировано, что EHG имеют различную степень происхождения от линии, связанной с WHG, в диапазоне от всего лишь 25% до 91%, а остальная часть связана с потоком генов от палеолитических сибиряков (ANE) и, возможно, кавказских охотников-собирателей . Другая линия, известная как скандинавские охотники-собиратели (SHG), представляет собой смесь EHG и WHG. [а] [22] [23]
Люди мезолитической культуры Кунда и нарвской культуры восточной Балтики представляли собой смесь WHG и EHG, [24] демонстрируя наибольшую близость к WHG. Было обнаружено, что образцы украинского мезолита и неолита плотно сгруппированы между WHG и EHG, что позволяет предположить генетическую преемственность в Днепровских порогах в течение 4000 лет. Украинские образцы принадлежали исключительно к материнской гаплогруппе U , которая встречается примерно в 80% всех европейских образцов охотников-собирателей. [25]
С ЭХГ были тесно связаны представители культуры ямочно-гребенчатой керамики (КСС) Восточной Балтики. [26] В отличие от большинства WHG, WHG восточной Балтики не получили примеси европейских земледельцев в эпоху неолита. Таким образом, современное население восточной Балтики имеет большее количество предков WHG, чем любое другое население Европы. [24]
Было обнаружено, что SHG содержат смесь компонентов WHG, которые, вероятно, мигрировали в Скандинавию с юга, и EHG, которые позже мигрировали в Скандинавию с северо-востока вдоль норвежского побережья. Эта гипотеза подтверждается данными о том, что у ГСП из западной и северной Скандинавии было меньше предков WHG (около 51%), чем у людей из восточной Скандинавии (около 62%). Считается, что WHG, проникшие в Скандинавию, принадлежали к аренсбургской культуре . EHG и WHG показали более низкие частоты аллелей SLC45A2 и SLC24A5 , которые вызывают депигментацию, и OCA/Herc2 , который вызывает светлый цвет глаз , чем SHG. [27]

ДНК одиннадцати WHG из верхнего палеолита и мезолита в Западной Европе , Центральной Европе и на Балканах была проанализирована на предмет их гаплогрупп Y-ДНК и гаплогрупп мтДНК . Анализ показал, что когда-то WHG были широко распространены от побережья Атлантического океана на западе до Сицилии на юге и до Балкан на юго-востоке на протяжении более шести тысяч лет. [28] Исследование также включало анализ большого количества особей доисторической Восточной Европы. Тридцать семь образцов собрано с территории мезолитической и неолитической Украины (9500-6000 гг. до н.э.). Было установлено, что они являются промежуточными между EHG и SHG, хотя происхождение WHG в этой популяции увеличилось в эпоху неолита. Образцы Y-ДНК, выделенные от этих людей, принадлежали исключительно гаплотипам R (особенно субкладам R1b1 ) и гаплотипам I (особенно субкладам I2 ). мтДНК принадлежала почти исключительно U (особенно субкладам U5 и U4 ). [28] большое количество особей из могильника Звейниеки Проанализировано , которые в основном принадлежали кундинской культуре и нарвской культуре в восточной Прибалтике. На более ранних стадиях эти люди в основном были потомками WHG, но со временем происхождение EHG стало преобладающим. Y-ДНК этого сайта принадлежала почти исключительно гаплотипам гаплогруппы R1b1a1a и I2a1 . МтДНК принадлежала исключительно гаплогруппе U (особенно субкладам U2 , U4 и U5 ). [28] Также были проанализированы сорок особей из трех стоянок мезолита Железных ворот на Балканах . По оценкам, эти люди имели 85% WHG и 15% EHG происхождения. Самцы на этих участках несли исключительно гаплотипы гаплогруппы R1b1a и I (в основном субклады I2a ). мтДНК принадлежала в основном U (особенно субкладам U5 и U4 ). [28] Было обнаружено, что люди балканского неолита на 98% имеют анатолийское происхождение и на 2% - WHG-происхождение. К эпохе энеолита было обнаружено , что люди культуры Кукутени-Триполье имеют около 20% предков охотников-собирателей, что занимало промежуточное положение между EHG и WHG. Было обнаружено, что люди культуры шаровидных амфор укрывают ок. 25% предков WHG, что значительно выше, чем у групп среднего неолита Центральной Европы. [28]
Замена неолитическими фермерами
[ редактировать ]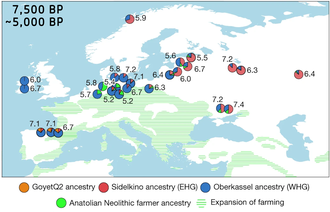

В плодотворном исследовании 2014 года впервые был выявлен вклад трех основных компонентов в современные европейские линии: западных охотников-собирателей (WHG, в пропорциях до 50% у северных европейцев), древних северных евразийцев (ANE, сибиряки верхнего палеолита, позже связанные с более поздняя индоевропейская экспансия , присутствующая в пропорциях до 20%), и, наконец, ранние европейские земледельцы (EEF, земледельцы в основном ближневосточного происхождения, которые мигрировали в Европу примерно с 8000 лет назад, сейчас присутствуют в пропорциях примерно 30% в Прибалтике). регионе до примерно 90% в Средиземноморье). Компонент « Ранние европейские фермеры» (EEF) был идентифицирован на основе генома женщины, похороненной ок. 7000 лет назад в могиле культуры линейной керамики в Штутгарте , Германия. [30]
Это исследование 2014 года обнаружило доказательства генетического смешения между WHG и EEF по всей Европе, с наибольшим вкладом EEF в Средиземноморской Европе (особенно на Сардинии, Сицилии, Мальте и среди евреев-ашкенази), а также наибольшим вкладом WHG в Северной Европе и среди басков. люди. [31]
С 2014 года дальнейшие исследования уточнили картину скрещивания EEF и WHG. В ходе анализа 180 наборов древних наборов данных ДНК периодов энеолита и неолита из Венгрии, Германии и Испании в 2017 году были обнаружены доказательства длительного периода скрещивания. Смешение происходило на региональном уровне, из местных популяций охотников-собирателей, так что популяции из трех регионов (Германия, Иберия и Венгрия) были генетически различимы на всех этапах периода неолита, с постепенно увеличивающимся соотношением предков WHG среди земледельческих популяций с течением времени. . Это говорит о том, что после первоначальной экспансии первых земледельцев не было дальнейших дальних миграций, достаточно существенных, чтобы гомогенизировать земледельческое население, и что земледельческое население и население охотников-собирателей существовали бок о бок на протяжении многих столетий, с продолжающимся постепенным смешением на протяжении всего V века. до 4-го тысячелетия до нашей эры (а не единичное событие примеси при первом контакте). [32] Уровень примеси варьировался в зависимости от географического положения; в эпоху позднего неолита доля WHG у фермеров в Венгрии составляла около 10%, в Германии - около 25%, а в Иберии - до 50%. [33]
Анализ останков из Грота Континенца в Италии показал, что из шести останков три захоронены между ок. 10 000 г. до н. э. и 7 000 г. до н. э. принадлежали I2a-P214 ; и вдвое больше материнских гаплогрупп U5b1 и одна U5b3 . [34] [35] Около 6000 г. до н. э. WHG Италии были почти полностью генетически заменены EEF (два G2a2 ) и одной гаплогруппой R1b , хотя в последующие тысячелетия родословная WHG немного увеличилась. [36]
Люди неолита на Британских островах были близки к иберийским и центральноевропейским популяциям раннего и среднего неолита, которые, согласно модели, имели около 75% предков от EEF, а остальная часть - от WHG в континентальной Европе. Впоследствии они заменили большую часть населения WHG на Британских островах, не смешиваясь с ними. [37]
По оценкам, WHG внесли 20-30% предков в неолитические группы EEF по всей Европе. Специфические приспособления против местных патогенов, возможно, были внесены посредством мезолитической примеси WHG в неолитические популяции EEF. [38]
Исследование мезолитических охотников-собирателей из Дании показало, что они были связаны с современными западными охотниками-собирателями и связаны с культурами Маглемосе , Конгемосе и Эртебёлле . Они демонстрировали «генетическую однородность примерно от 10 500 до 5 900 лет назад», пока «не прибыли неолитические фермеры анатолийского происхождения». Переход к периоду неолита был «очень резким и привел к смене населения с ограниченным генетическим вкладом местных охотников-собирателей. Последующая неолитическая популяция была связана с культурой воронкообразных . [39]
Внешний вид
[ редактировать ]
По словам Дэвида Райха , анализ ДНК показал, что западные охотники-собиратели обычно были темнокожими, темноволосыми и голубоглазыми. [42] Темная кожа была обусловлена их происхождением из Африки (все популяции Homo sapiens изначально имели темную кожу), а голубые глаза были результатом изменения гена OCA2, который вызывал радужной оболочки . депигментацию [43]
Археолог Грэм Уоррен сказал, что цвет их кожи варьировался от оливкового до черного, и предположил, что у них могли быть некоторые региональные различия цвета глаз и волос. [44] Это разительно отличается от дальних родственников восточных охотников-собирателей (EHG), которые, как предполагалось, были светлокожими, кареглазыми или голубоглазыми, темноволосыми или светловолосыми. [45]
По прогнозам, два скелета WHG с неполными SNP , Ла Бранья и Человек из Чеддера , имели темную или от темной до черной кожу, тогда как два других скелета WHG с полными SNP, «Свен» и человек из Лошбора , по прогнозам, имели темную или средне- черную кожу. к темной и промежуточной коже соответственно. [46] [27] [б] Испанский биолог Карлес Лалуэса-Фокс сказал, что у особи Ла Бранья-1 была темная кожа, «хотя мы не можем знать точный оттенок». [48]
Согласно исследованию 2020 года, прибытие ранних европейских фермеров (EEF) из Западной Анатолии от 8500 до 5000 лет назад вместе с пастухами западных степей в бронзовом веке вызвало быструю эволюцию европейского населения в сторону более светлой кожи и волос. [43] Смешение охотников-собирателей и земледельцев, очевидно, было случайным, но не обширным. [49]

Некоторые авторы выражают осторожность в отношении реконструкций пигментации кожи : Quillen et al. (2019) признают исследования, которые в целом показывают, что «более светлый цвет кожи был редкостью на большей части Европы во время мезолита», включая исследования, касающиеся предсказаний «темного или темного или темно-черного» человека Чеддера, но предупреждают, что «реконструкции мезолита и неолита Фенотип пигментации с использованием локусов, распространенных в современных популяциях, следует интерпретировать с некоторой осторожностью, поскольку вполне возможно, что другие, еще не изученные локусы, также могли влиять на фенотип». [50]
Генетик Сьюзан Уолш из Университета Индианы – Университета Пердью в Индианаполисе , которая работала над проектом «Человек Чеддер», сказала, что «мы просто не знаем цвет его кожи». [51] Немецкий биохимик Йоханнес Краузе заявил, что мы не знаем, был ли цвет кожи западноевропейских охотников-собирателей больше похож на цвет кожи людей из современной Центральной Африки или людей из арабского региона . Достоверно лишь то, что они не несли какой-либо известной мутации, ответственной за светлую кожу у последующих популяций европейцев. [52]
Исследование геномного происхождения и социальной динамики последних охотников-собирателей Атлантической Франции, проведенное в 2024 году, показало, что «фенотипически мы обнаруживаем некоторое разнообразие во время позднего мезолита во Франции», при котором два из секвенированных в исследовании WHG «вероятно имели пигментация кожи от бледной до средней», но «большинство людей имеют темную кожу и голубые глаза, характерные для WHG» изученных образцов. [53]
Примечания
[ редактировать ]- ^ Восточные охотники-собиратели (EHG) на 3/4 своего происхождения происходят от ANE... Скандинавские охотники-собиратели (SHG) представляют собой смесь EHG и WHG; и WHG представляют собой смесь EHG и верхнепалеолитического бишона из Швейцарии. [21]
- ^ Эти прогнозы были получены с использованием модели полиномиальной логистической регрессии , основанной на панели из 36 тщательно отобранных SNP с низкой чувствительностью 0,26 для классификации промежуточной кожи (по сравнению с 0,99 и 0,90 для белой и черной кожи соответственно). Точность используемой модели может быть дополнительно повышена за счет «дополнительных (но пока неизвестных) предикторов SNP, которые будут идентифицированы с помощью будущего GWAS ». [47]
Ссылки
[ редактировать ]- ^ Пост, Козимо; Ю, Хэ; Галичи, Айшин (март 2023 г.). «Палеогеномика от верхнего палеолита до неолита европейских охотников-собирателей» . Природа . 615 (7950): 117–126. Бибкод : 2023Natur.615..117P . дои : 10.1038/s41586-023-05726-0 . ISSN 1476-4687 . ПМЦ 9977688 . ПМИД 36859578 . S2CID 257259969 .
- ^ Jump up to: а б Энтони 2019b , с. 28.
- ^ Jump up to: а б Кашуба 2019 : «Более ранние исследования адДНК предполагают наличие трех генетических групп в ранней послеледниковой Европе: западных охотников-собирателей (WHG), восточных охотников-собирателей (EHG) и скандинавских охотников-собирателей (SHG)4. SHG были смоделированы. как смесь WHG и EHG».
- ^ Jump up to: а б с д и ж г Пост, К., Ю, Х., Галичи, А. (2023). «Палеогеномика от верхнего палеолита до неолита европейских охотников-собирателей» . Природа . 615 (2 марта 2023 г.): 117–126. Бибкод : 2023Natur.615..117P . дои : 10.1038/s41586-023-05726-0 . ПМЦ 9977688 . ПМИД 36859578 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Вильяльба-Моуко, Ванесса; ван де Лоосдрехт, Марике С.; Пост, Козимо; Мора, Рафаэль; Мартинес-Морено, Хорхе; Рохо-Гуэрра, Мануэль; Саласар-Гарсия, Доминго К.; Ройо-Гильен, Хосе И.; Кунст, Майкл; Ружье, Элен; Кревкер, Изабель; Аркуса-Магаллон, Гектор; Техедор-Родригес, Кристина; Гарсиа-Мартинес де Лагран, Иньиго; Гарридо-Пенья, Рафаэль (апрель 2019 г.). «Сохранение предков охотников-собирателей позднего плейстоцена на Пиренейском полуострове» . Современная биология . 29 (7): 1169–1177.e7. дои : 10.1016/j.cub.2019.02.006 .
- ^ Дэви, Том; Джу, Дэн; Мэтисон, Иэн; Скоглунд, Понт (апрель 2023 г.). «Примесь охотников-собирателей способствовала естественному отбору у европейских земледельцев эпохи неолита» . Современная биология . 33 (7): 1365–1371.e3. Бибкод : 2023CBio...33E1365D . дои : 10.1016/j.cub.2023.02.049 . ISSN 0960-9822 . ПМЦ 10153476 . ПМИД 36963383 .
- ^ Jump up to: а б с Лазаридис 2014 .
- ^ Мэтисон 2015 .
- ^ Вязание крючком 2015 .
- ^ Фу, Цяомэй (2016). «Генетическая история Европы ледникового периода» . Природа . 534 (7606): 200–205. Бибкод : 2016Natur.534..200F . дои : 10.1038/nature17993 . ПМЦ 4943878 . ПМИД 27135931 .
Начиная с скопления Виллабруна, по крайней мере, ~ 14 000 лет назад, все проанализированные европейские особи демонстрируют близость к Ближнему Востоку. По времени это коррелирует с интерстадиалом Бёллинг-Аллерёд, первым значительным периодом потепления после ледникового периода. Археологически это коррелирует с культурными переходами внутри эпиграветта в Южной Европе и переходом от Магдалины к Азилию в Западной Европе. Таким образом, появление скопления Виллабруна может отражать миграции или популяционные сдвиги внутри Европы в конце ледникового периода, и это наблюдение также согласуется с данными об обновлении последовательностей митохондриальной ДНК в это время. Одним из сценариев, который мог бы объяснить эти закономерности, является расширение населения из рефугиумов Юго-Восточной Европы или Западной Азии после ледникового периода, объединяющее генетическое происхождение Европы и Ближнего Востока. В-шестых, в кластере Виллабруна некоторые, но не все, люди имеют близость к выходцам из Восточной Азии. Важным направлением будущей работы является получение аналогичных данных древней ДНК из Юго-Восточной Европы и Ближнего Востока, чтобы получить более полную картину истории верхнего палеолита населения Западной Евразии.
- ^ Сикора, Мартин; Питулько Владимир Владимирович; Соуза, Витор К.; Аллентофт, Мортен Э.; Победитель, Лассе; Расмуссен, Саймон; Маргарян, Ашот; де Баррос Дамгаард, Питер; Фонтана, Констанция; Рено, Габриэль; Ян, Мелинда А.; Фу, Цяомэй; Дюпанлуп, Изабель; Джампудакис, Константинос; Ногес-Браво, Давид (июнь 2019 г.). «История населения Северо-Восточной Сибири начиная с плейстоцена» . Природа . 570 (7760): 182–188. Бибкод : 2019Natur.570..182S . дои : 10.1038/s41586-019-1279-z . hdl : 1887/3198847 . ISSN 1476-4687 . ПМИД 31168093 . S2CID 174809069 .
- ^ Валлини и др. 2022 г. (4 июля 2022 г.). «Генетика и материальная культура поддерживают неоднократные экспансии в палеолитическую Евразию из популяционного центра из Африки» . Проверено 16 апреля 2023 г.
{{cite web}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Марки, Нина; Винкельбах, Лаура; Шульц, Электра; Брами, Максим; Хофманова, Зузана; Блёхер, Йенс; Рейна-Бланко, Карлос С.; Дикманн, Джоан; Тьери, Александр; Капопулу, Адамандиа; Линк, Вивиан; Пьюз, Валери; Крейцер, Сюзанна; Фигарска, Сильвия М.; Ганиацу, Элиссавет (май 2022 г.). «Геномное происхождение первых фермеров мира» . Клетка . 185 (11): 1842–1859.e18. дои : 10.1016/j.cell.2022.04.008 . ПМК 9166250 . ПМИД 35561686 .
- ^ Скоццари, Розария; Массайя, Андреа; Д'Атанасио, Евгения; Майрес, Натали М.; Перего, Уго А.; Тромбетта, Бениамино; Кручиани, Фульвио (7 ноября 2012 г.). «Молекулярное рассечение базальных клад филогенетического дерева Y-хромосомы человека» . ПЛОС ОДИН . 7 (11): e49170. Бибкод : 2012PLoSO...749170S . дои : 10.1371/journal.pone.0049170 . ISSN 1932-6203 . ПМЦ 3492319 . ПМИД 23145109 .
Благодаря этому анализу мы идентифицировали хромосому из южной Европы как новую глубокую ветвь гаплогруппы C (C-V20 или C7, рисунок S1). Ранее в Южной Европе было обнаружено лишь несколько примеров С-хромосом (определяемых только маркером RPS4Y711) [32], [33]. Чтобы улучшить наши знания о распространении гаплогруппы C в Европе, мы опросили 1965 европейцев на наличие мутации RPS4Y711 и идентифицировали одну дополнительную хромосому гаплогруппы C из южной Европы, которая также была классифицирована как C7 (данные не показаны).
- ^ Бортолини, Эухенио; Пагани, Лука; Оксилия, Григорий; Пост, Козимо; Фонтана, Федерика; Бадино, Федерика; Саупе, Тина; Монтинаро, Франческо; Маргаритора, Давиде; Романдини, Маттео; Лугли, Федерико; Папини, Андреа; Богджиони, Марко; Перрини, Никола; Оксилия, Антонио (июнь 2021 г.). «Ранняя альпийская оккупация возникла в прошлом, когда люди мигрировали на запад в позднеледниковую Европу» . Современная биология . 31 (11): 2484–2493.e7. Бибкод : 2021CBio...31E2484B . дои : 10.1016/j.cub.2021.03.078 . hdl : 11585/827591 . ПМИД 33887180 .
- ^ «Ученые секвенировали геномы доисторических охотников-собирателей из разных евразийских культур» . Научные новости . 2 марта 2023 г.
- ^ Ирвинг-Пиз, Эван К.; Рефойо-Мартинес, Альба; Барри, Уильям; Ингасон, Андрес; Пирсон, Элис; Фишер, Андерс; Шегрен, Карл-Йоран; Халгрен, Альма С.; Маклеод, Руайрид; Деметра, Фабрис; Хенриксен, Расмус А.; Вимала, Тарсика; Макколл, Хью; Вон, Эндрю Х.; Шпайдель, Лео (январь 2024 г.). «Селекционный ландшафт и генетическое наследие древних евразийцев» . Природа . 625 (7994): 312–320. Бибкод : 2024Natur.625..312I . дои : 10.1038/s41586-023-06705-1 . ISSN 1476-4687 . ПМЦ 10781624 . ПМИД 38200293 .
...в то время как аллели риска диабета и болезни Альцгеймера обогащены западными предками охотников-собирателей.
- ^ Чарльтон, Софи; Брейс, Селина (ноябрь 2022 г.). «Двойное происхождение и экология позднего ледникового палеолита в Британии» . Экология и эволюция природы . 6 (11): 1658–1668. Бибкод : 2022NatEE...6.1658C . дои : 10.1038/s41559-022-01883-z . ISSN 2397-334X . ПМК 9630104 . ПМИД 36280785 .
- ^ Симоэнс, Лусиана Г.; Гюнтер, Торстен; Мартинес-Санчес, Рафаэль М.; Вера-Родригес, Хуан Карлос; Ириарте, Энеко; Родригес-Варела, Рикардо; Бокбот, Юсеф; Вальдиосера, Кристина; Якобссон, Маттиас (7 июня 2023 г.). «Неолит Северо-Западной Африки, инициированный мигрантами из Иберии и Леванта» . Природа . 618 (7965): 550–556. Бибкод : 2023Nature.618..550S . дои : 10.1038/s41586-023-06166-6 . ISSN 1476-4687 . ПМК 10266975 . ПМИД 37286608 .
- ^ Jump up to: а б Лазаридис, Иосиф (1 декабря 2018 г.). «Эволюционная история человеческих популяций в Европе». Текущее мнение в области генетики и развития . Генетика происхождения человека. 53 : 21–27. arXiv : 1805.01579 . дои : 10.1016/j.gde.2018.06.007 . ISSN 0959-437X . ПМИД 29960127 . S2CID 19158377 .
- ^ Лазаридис 2016 .
- ^ Хаак, Вольфганг; Лазаридис, Иосиф; Паттерсон, Ник; Роланд, Надин; Маллик, Свапан; Ламас, Бастьен; Брандт, Гвидо; Норденфельт, Сюзанна; Харни, Эдаоин; Стюардсон, Кристин; Фу, Цяомэй; Миттник, Алисса; Банфи, Эстер; Эконому, Христос; Франкен, Майкл (июнь 2015 г.). «Массовая миграция из степи явилась источником индоевропейских языков в Европе» . Природа . 522 (7555): 207–211. arXiv : 1502.02783 . Бибкод : 2015Natur.522..207H . дои : 10.1038/nature14317 . ISSN 1476-4687 . ПМК 5048219 . ПМИД 25731166 .
- ^ Сикора, Мартин; Питулько Владимир Владимирович; Соуза, Витор К.; Аллентофт, Мортен Э.; Победитель, Лассе; Расмуссен, Саймон; Маргарян, Ашот; де Баррос Дамгаард, Питер; Фонтана, Констанция; Рено, Габриэль; Ян, Мелинда А.; Фу, Цяомэй; Дюпанлуп, Изабель; Джампудакис, Константинос; Ногес-Браво, Давид (июнь 2019 г.). «История населения Северо-Восточной Сибири начиная с плейстоцена» . Природа . 570 (7760): 182–188. Бибкод : 2019Natur.570..182S . дои : 10.1038/s41586-019-1279-z . hdl : 1887/3198847 . ISSN 1476-4687 . ПМИД 31168093 . S2CID 174809069 .
- ^ Jump up to: а б Миттник 2018 .
- ^ Джонс 2017 .
- ^ Видел 2017 год .
- ^ Jump up to: а б Гюнтер 2018 .
- ^ Jump up to: а б с д и Мэтисон 2018 .
- ^ Сикора М., Карпентер М.Л., Морено-Эстрада А., Хенн Б.М., Андерхилл П.А., Санчес-Куинто Ф. и др. (май 2014 г.). «Популяционный геномный анализ древних и современных геномов дает новое представление о генетическом происхождении тирольского ледяного человека и генетической структуре Европы» . ПЛОС Генетика . 10 (5): e1004353. дои : 10.1371/journal.pgen.1004353 . ПМК 4014435 . ПМИД 24809476 .
- ^ Лазаридис, Иосиф; Паттерсон, Ник; Миттник, Алисса (сентябрь 2014 г.). «Древние человеческие геномы позволяют предположить три предковые популяции современных европейцев» . Природа . 513 (7518): 409–413. arXiv : 1312.6639 . Бибкод : 2014Natur.513..409L . дои : 10.1038/nature13673 . ISSN 1476-4687 . ПМК 4170574 . ПМИД 25230663 .
Большинство современных европейцев происходят по крайней мере из трех высокодифференцированных популяций: западноевропейских охотников-собирателей, которые внесли свой вклад в происхождение всех европейцев, но не жителей Ближнего Востока; древние северные евразийцы, родственные сибирякам верхнего палеолита, которые внесли свой вклад как в европейцев, так и в жителей Ближнего Востока; и ранние европейские фермеры, которые были в основном выходцами с Ближнего Востока, но также имели предков западноевропейских охотников-собирателей.
- ^ Лазаридис и др. (2014), Дополнительная информация , с. 113.
- ^ Липсон и др., «Параллельные палеогеномные разрезы раскрывают сложную генетическую историю ранних европейских фермеров», Nature 551, 368–372 (16 ноября 2017 г.) дои : 10.1038/nature24476 .
- ^ Липсон и др. (2017), рис. 2.
- ^ Антонио и др. 2019 г. , Таблица 2. Пример информации, строки 4–6.
- ^ Антонио и др. 2019 , с. 1.
- ^ Антонио и др. 2019 , с. 2, рис. 1.
- ^ Брейс, Селина; Дикманн, Йоан; Бут, Томас Дж.; ван Дорп, Люси; Фалтыскова, Зузана; Роланд, Надин; Маллик, Свапан; Олальде, Иньиго; Ферри, Мэтью; Мишель, Меган; Оппенгеймер, Йонас; Брумандхошбахт, Насрин; Стюардсон, Кристин; Мартиниано, Руи; Уолш, Сьюзен; Кайзер, Манфред; Чарльтон, Софи; Хеллентал, Гарретт; Армит, Ян; Шультинг, Рик; Крейг, Оливер Э.; Шеридан, Элисон; Паркер Пирсон, Майк; Стрингер, Крис; Райх, Дэвид; Томас, Марк Г.; Барнс, Ян (2019). «Древние геномы указывают на смену населения в Британии раннего неолита» . Экология и эволюция природы . 3 (5): 765–771. Бибкод : 2019NatEE...3..765B . дои : 10.1038/s41559-019-0871-9 . ISSN 2397-334X . ПМК 6520225 . ПМИД 30988490 .
- ^ Дэви, Том; Джу, Дэн; Мэтисон, Иэн; Скоглунд, Понт (апрель 2023 г.). «Примесь охотников-собирателей способствовала естественному отбору у европейских земледельцев эпохи неолита» . Современная биология . 33 (7): 1365–1371.e3. Бибкод : 2023CBio...33E1365D . дои : 10.1016/j.cub.2023.02.049 . ISSN 0960-9822 . ПМЦ 10153476 . ПМИД 36963383 .
- ^ Аллентофт, Мортен Э.; Сикора, Мартин; Фишер, Андерс; Шегрен, Карл-Горан; Ингасон, Андрес; Маклеод, Руайрид; Розенгрен, Андерс; Шульц Паулссон, Беттина; Йорков, Мария Луиза Шеллеруп; Новосолова Мария; Стендерап, Йеспер; Прайс, Т. Дуглас; Фишер Мортенсен, Мортен; Нильсен, Энн Биргитте; Ульфельдт Хеде, Миккель (10 января 2024 г.). «100 древних геномов показывают повторяющуюся смену населения в неолитической Дании» . Природа . 625 (7994): 329–337. Бибкод : 2024Nature.625..329A . дои : 10.1038/s41586-023-06862-3 . ISSN 1476-4687 . ПМЦ 10781617 . ПМИД 38200294 .
- ^ Коннеллер, Шанталь (29 ноября 2021 г.). Мезолит в Британии: ландшафт и общество во времена перемен . Рутледж. п. 126. ИСБН 978-1-000-47515-9 .
- ^ Подробности о реконструкции из Музея естественной истории: «Человек Чеддер: голубоглазый мальчик мезолитической Британии» . www.nhm.ac.uk. Музей естественной истории.
- ^ Райх, Дэвид (2018). Кто мы и как нам здесь жарко: древняя ДНК и новая наука о человеческом прошлом (первое изд.). Нью-Йорк: Издательская группа Knopf Doubleday. ISBN 978-1101870334 . «Анализ данных древней ДНК показывает, что охотники-собиратели Западной Европы около восьми тысяч лет назад имели голубые глаза, но темную кожу и темные волосы — сочетание, которое сегодня редко встречается».
- ^ Jump up to: а б с Ханель, Андреа; Карлберг, Карстен (сентябрь 2020 г.). «Цвет кожи и витамин D: обновленная информация» . Экспериментальная дерматология . 29 (9): 867. doi : 10.1111/exd.14142 . ISSN 0906-6705 . ПМИД 32621306 . S2CID 220335539 .
Человек разумный прибыл в Европу с Ближнего Востока около 42 000 лет назад. Как и в случае с африканским происхождением, эти люди имели темную кожу, но из-за вариаций гена OCA2 (вызывающего депигментацию радужной оболочки) у многих из них были голубые глаза» (...) «в южной и центральной Европе, где они [аллели светлой кожи] были завезен фермерами из Западной Анатолии, расширявшимися от 8500 до 5000 лет назад. Это было началом неолитической революции в этих регионах, характеризующейся более оседлым образом жизни и одомашниванием некоторых видов животных и растений. (...) «Быстрый рост численности населения в результате неолитической революции, такой как использование молочных продуктов в качестве источника пищи для взрослых и развитие сельского хозяйства, а также массовое распространение ямных скотоводов, вероятно, вызвали быструю селекцию европейское население стремится к светлой коже и волосам
- ^ Уоррен, Грэм (2021). Охотник-собиратель Ирландия: налаживание связей в островном мире . Книги Оксбоу. ISBN 978-1789256840 . «Например, WHG имели пигментацию кожи от оливкового до коричневого и черного цвета, голубые или сине-зеленые глаза. В некоторых частях Европы это также могло быть связано со светлыми волосами».
- ^ Популяционная геномика мезолитической Скандинавии: исследование маршрутов ранней послеледниковой миграции и адаптации к высоким широтам S8 Text. Функциональная вариация древних образцов. , doi : 10.1371/journal.pbio.2003703.s013
- ^ Брейс, Селина; Дикманн, Йоан; Бут, Томас Дж.; Фалтыскова, Зузана; Роланд, Надин; Маллик, Свапан; Ферри, Мэтью; Мишель, Меган; Оппенгеймер, Йонас; Брумандхошбахт, Насрин; Стюардсон, Кристин; Уолш, Сьюзен; Кайзер, Манфред; Шультинг, Рик; Крейг, Оливер Э.; Шеридан, Элисон; Пирсон, Майк Паркер; Стрингер, Крис; Райх, Дэвид; Томас, Марк Г.; Барнс, Ян (2019), «Замещение населения в Британии раннего неолита», Nature Ecology & Evolution , 3 (5): 765–771, Бибкод : 2019NatEE...3..765B , doi : 10.1038/s41559-019-0871 -9 , PMC 6520225 , PMID 30988490 Дополнительный материал. Страница 22: «По прогнозам, у двух WHG (Чеддер-Мэн и Ла-Бранья из северной Испании) была темная кожа или кожа от темного до черного, тогда как у одного (Лошбур44 из Люксембурга), по прогнозам, кожа была промежуточной, но мы находим потенциальную временную и / или географические различия в характеристиках пигментации, что позволяет предположить, что различные уровни пигментации кожи сосуществовали у WHG, по крайней мере, примерно на 8 kBP. Было предсказано, что у Свена была кожа от темного до среднего или темного цвета, что соответствует текущей гипотезе о том, что аллели обычно связаны с более светлой кожей. в европейцах были завезены в Северо-Западную Европу благодаря ANF».
- ^ Уолш, Сьюзен (2017). «Глобальное предсказание цвета кожи по ДНК» . Генетика человека . 136 (7): 847–863. дои : 10.1007/s00439-017-1808-5 . ПМЦ 5487854 . ПМИД 28500464 .
- ^ «Темная кожа, голубые глаза: гены рисуют картину европейца 7000-летней давности» . Новости Эн-Би-Си . 26 января 2014 г.
- ^ Каллауэй, Юэн (12 мая 2022 г.). «Древние карты ДНК «на заре земледелия» » . Природа . дои : 10.1038/d41586-022-01322-w . ПМИД 35552521 . S2CID 248765487 .
Команда Экскофье обнаружила, что, обосновавшись в Анатолии, ранние земледельческие популяции двинулись на запад, в Европу, словно ступенька, начиная примерно 8000 лет назад. Время от времени они смешивались, но не слишком часто, с местными охотниками-собирателями.
- ^ Куиллен, Эллен (2019). «Оттенки сложности: новые взгляды на эволюцию и генетическую архитектуру кожи человека» . Американский журнал биологической антропологии . 168 (С67): 4–26. дои : 10.1002/ajpa.23737 . ПМИД 30408154 . S2CID 53237190 .
Их анализ показывает, что цвет кожи обоих людей, вероятно, был темным, а цвет кожи мезолитического человека Чеддер, по прогнозам, был «темным или от темного до черного». Эти данные позволяют предположить, что более светлый цвет кожи был редкостью на большей части территории Европы в эпоху мезолита. Однако это не противоречит оценкам дат <20 тыс. лет назад, которые касаются начала отбора, а не времени фиксации предпочтительных аллелей (Beleza et al., 2013; Beleza, Johnson и др., 2013). . Хотя исследования древнего генома предсказывают, что среди мезолитических европейцев в целом более темный цвет кожи, производные аллели rs1426654 и rs16891982 разделялись в европейских популяциях во время мезолита (González-Fortes et al., 2017; Günther et al., 2018; Mittnik et al., 2018). ), что позволяет предположить, что к этому времени, вероятно, уже присутствовали фенотипические вариации, обусловленные этими локусами. Однако реконструкции фенотипа пигментации мезолита и неолита с использованием локусов, распространенных в современных популяциях, следует интерпретировать с некоторой осторожностью, поскольку возможно, что другие, еще не изученные локусы также могли повлиять на фенотип.
- ^ «Находка древнего темнокожего человека из Чеддера может быть неправдой» . Новый учёный . 21 февраля 2018 г.
- ^ Краузе, Йоханнес (2021). Краткая история человечества. Новая история старой Европы . l: Издательская группа Random House. ISBN 9780593229446 .
- ^ Симоэнс, Лусиана Г.; Пейротео-Стьерна, Рита; Маршан, Грегор; Бернхардссон, Каролина; Виалет, Амели; Четти, Даршан; Алачамлы, Эркин; Эдлунд, Ханна; Букен, Дени; Дина, Кристиан; Гармонд, Николас; Гюнтер, Торстен; Якобссон, Маттиас (5 марта 2024 г.). «Геномное происхождение и социальная динамика последних охотников-собирателей Атлантической Франции» . Труды Национальной академии наук . 121 (10): e2310545121. Бибкод : 2024PNAS..12110545S . дои : 10.1073/pnas.2310545121 . ISSN 0027-8424 . ПМЦ 10927518 . ПМИД 38408241 .
Библиография
[ редактировать ]- Энтони, Дэвид В. (2019b). «Древняя ДНК, сети спаривания и анатолийский раскол». В Серанджели Матильда; Оландер, Томас (ред.). Рассредоточение и диверсификация: лингвистические и археологические перспективы ранних этапов индоевропейского происхождения . БРИЛЛ . стр. 21–54. ISBN 978-9004416192 .
- Антонио, Маргарет Л.; и др. (8 ноября 2019 г.). «Древний Рим: генетический перекресток Европы и Средиземноморья» . Наука . 366 (6466). Американская ассоциация содействия развитию науки : 708–714. Бибкод : 2019Sci...366..708A . дои : 10.1126/science.aay6826 . ПМК 7093155 . ПМИД 31699931 .
- Гюнтер, Торстен (1 января 2018 г.). «Популяционная геномика мезолитической Скандинавии: исследование маршрутов ранней послеледниковой миграции и адаптации к высоким широтам» . ПЛОС Биология . 16 (1). ПЛОС : e2003703. дои : 10.1371/journal.pbio.2003703 . ПМК 5760011 . ПМИД 29315301 .
- Хаак, Вольфганг (11 июня 2015 г.). «Массовая миграция из степи явилась источником индоевропейских языков в Европе» . Природа . 522 (7555). Исследования природы : 207–211. arXiv : 1502.02783 . Бибкод : 2015Natur.522..207H . дои : 10.1038/nature14317 . ПМК 5048219 . ПМИД 25731166 .
- Джонс, Эппи Р. (20 февраля 2017 г.). «Переход к неолиту в Прибалтике не был вызван смешением с ранними европейскими земледельцами» . Современная биология . 27 (4). Cell Press : 576–582. Бибкод : 2017CBio...27..576J . дои : 10.1016/j.cub.2016.12.060 . ПМК 5321670 . ПМИД 28162894 .
- Кашуба, Наталья (15 мая 2019 г.). «Древняя ДНК из мастики укрепляет связь между материальной культурой и генетикой мезолитических охотников-собирателей в Скандинавии» . Коммуникационная биология . 2 (105). Исследования природы : 185. doi : 10.1038/s42003-019-0399-1 . ПМК 6520363 . ПМИД 31123709 .
- Лазаридис, Иосиф (17 сентября 2014 г.). «Древние человеческие геномы позволяют предположить три предковые популяции современных европейцев» . Природа . 513 (7518). Исследования природы : 409–413. arXiv : 1312.6639 . Бибкод : 2014Natur.513..409L . дои : 10.1038/nature13673 . hdl : 11336/30563 . ПМК 4170574 . ПМИД 25230663 .
- Лазаридис, Иосиф (25 июля 2016 г.). «Геномный взгляд на происхождение земледелия на древнем Ближнем Востоке» . Природа . 536 (7617). Исследования природы : 419–424. Бибкод : 2016Natur.536..419L . дои : 10.1038/nature19310 . ПМЦ 5003663 . ПМИД 27459054 .
- Мэтисон, Иэн (23 ноября 2015 г.). «Общегеномные закономерности отбора у 230 древних евразийцев» . Природа . 528 (7583). Исследования природы : 499–503. Бибкод : 2015Natur.528..499M . дои : 10.1038/nature16152 . ПМЦ 4918750 . ПМИД 26595274 .
- Мэтисон, Иэн (21 февраля 2018 г.). «Геномная история Юго-Восточной Европы» . Природа . 555 (7695). Исследования природы : 197–203. Бибкод : 2018Natur.555..197M . дои : 10.1038/nature25778 . ПМК 6091220 . ПМИД 29466330 .
- Миттник, Алиса (30 января 2018 г.). «Генетическая предыстория региона Балтийского моря» . Природные коммуникации . 16 (1). Исследования природы : 442. Бибкод : 2018NatCo...9..442M . дои : 10.1038/s41467-018-02825-9 . ПМЦ 5789860 . ПМИД 29382937 .
- Сааг, Лехти (24 июля 2017 г.). «Экстенсивное земледелие в Эстонии началось благодаря миграции из степи по признаку пола» . Современная биология . 27 (14). Cell Press : 2185–2193. Бибкод : 2017CBio...27E2185S . дои : 10.1016/j.cub.2017.06.022 . ПМИД 28712569 .
Дальнейшее чтение
[ редактировать ]- Энтони, Дэвид (весна – лето 2019 г.). «Археология, генетика и язык в степях: комментарий к Бомхарду» . Журнал индоевропейских исследований . 47 (1–2) . Проверено 9 января 2020 г.
- Лазаридис, Иосиф (декабрь 2018 г.). «Эволюционная история человеческих популяций в Европе». Текущее мнение в области генетики и развития . 53 . Эльзевир : 21–27. arXiv : 1805.01579 . дои : 10.1016/j.gde.2018.06.007 . ПМИД 29960127 . S2CID 19158377 .