Гаплогруппа R1b
| Гаплогруппа R1b | |
|---|---|
 | |
| Возможное время происхождения | Вероятно, вскоре после R1, возможно, между 18 000–14 000 г. до н.э. [1] |
| Возможное место происхождения | Западная Азия , Северная Евразия или Восточная Европа [2] |
| Предок | Р1 |
| Потомки |
|
| Определение мутаций | М343 |
Гаплогруппа R1b ( R-M343 ), ранее известная как Hg1 и Eu18 , представляет собой гаплогруппу Y-хромосомы человека .
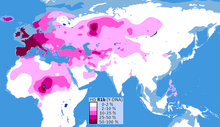
Это наиболее часто встречающаяся отцовская линия в Западной Европе , а также в некоторых частях России (например, у Башкир ) и по всему Сахелю в Центральной Африке , а именно: Камерун , Чад , Гвинея , Мавритания , Мали , Нигер , Нигерия и Сенегал ( сконцентрированы в некоторых частях Чада с концентрацией в племени хауса и среди чадоязычных этнических групп Камеруна).
Клада также присутствует на более низких частотах в Восточной Европе , Западной Азии , Центральной Азии, а также в некоторых частях Северной Африки , Южной Азии и Центральной Азии .
R1b имеет две основные ветви: R1b1-L754 и R1b2-PH155. R1b1-L754 имеет два основных субклада: R1b1a1b-M269, который преобладает в Западной Европе, и R1b1b-V88, который сегодня распространен в некоторых частях Центральной Африки. Другая ветвь, R1b2-PH155, настолько редка и широко распространена, что трудно сделать какие-либо выводы о ее происхождении. Он был обнаружен в Бахрейне , Индии , Непале , Бутане , Ладакхе , Таджикистане , Турции и Западном Китае .
Согласно древним исследованиям ДНК, большинство линий R1a и R1b произошли из Понтийской степи вместе с индоевропейскими языками . [2] [3] [4] [5] [6]
Происхождение и распространение
[ редактировать ]
Возраст R1 оценили Татьяна Карафет и др. (2008) между 12 500 и 25 700 лет назад и, скорее всего, произошло около 18 500 лет назад. [7] Поскольку самый ранний известный пример датируется примерно 14 000 лет назад и принадлежит R1b1 (R-L754), [1] R1b должен был возникнуть относительно вскоре после появления R1.
Ранние человеческие останки, содержащие R1b, включают:
- Виллабруна 1 (особь I9030), западный охотник-собиратель (WHG), найденный в среде эпиграветтской культуры в долине Сисмон (современный Венето , Италия), который жил около 14000 лет назад и принадлежал к R1b1a. [1] [8]
- Несколько мужчин из мезолита Железных Ворот на Балканах, похороненных между 11200 и 8200 годами назад, несли R1b1a1a. Было установлено, что эти люди в основном имеют происхождение от WHG с небольшой примесью восточных охотников-собирателей (EHG). [9]
- Несколько мужчин мезолитической культуры Кунда и неолитической нарвской культуры похоронены в могильнике Звейниеки на территории современной Латвии ок. 9500–6000 л.н. несли R1b1b. [10] [11] Было установлено, что эти люди в основном имели происхождение от WHG с небольшой примесью EHG. [10]
- Несколько мужчин эпохи мезолита и неолита похоронены в Дереевке и Васильевке на территории современной Украины ок. 9500-7000 л.н. несли R1b1a. [12] Эти люди в основном имели происхождение от EHG со значительной примесью WHG. [13]
- Мужчина WHG похоронен в Островуле Корбули, Румыния ок. 8700 BP несли R1b1c. [14] [15] [16]
- Мужчина похоронен в Лепенски Вир , Сербия ок. 8200-7900 лет назад несли R1b1a. [17]
- ЭХГ, похороненный недалеко от Самары , Россия, 7500 лет назад, содержал R1b1a1a. [18]
- Мужчина эпохи энеолита, похороненный в Хвалынске , Россия ок. 7200-6000 лет назад несли R1b1a. [19]
- Мужчина эпохи неолита , похороненный в Эльс-Трокс , Испания ок. 7178-7066 BP, которые, возможно, принадлежали к эпикардиальной культуре, [20] оказался носителем R1b1. [21] [22] [23]
- Мужчина позднего энеолита, похороненный в Смядово , Болгария ок. 6500 BP несли R1b1a. [24]
- Мужчина раннего медного века, похороненный в Каннас-ди-Сотто, Карбония, Сардиния ок. 6450 BP несли R1b1b2. [25]
- Мужчина из группы Баальберге в Центральной Европе похоронен ок. 5600 лет назад несли R1b1a. [26]
- Мужчина ботайской культуры в Средней Азии похоронен ок. 5500 лет назад несли R1b1a1 (R1b-M478). [27]
- Все семь протестированных мужчин ямной культуры оказались принадлежащими к субкладу M269 гаплогруппы R1b. [28]
R1b — субклад внутри «макрогаплогруппы » K ( M9), наиболее распространенной группы мужских линий человека за пределами Африки. Считается, что K возник в Азии (как и в случае с еще более ранней предковой гаплогруппой F (F-M89). Карафет Т. и др. (2014) предположили, что «быстрый процесс диверсификации K-M526, вероятно, произошел в Юго-Восточная Азия с последующей экспансией на запад предков гаплогрупп R и Q ». [29] Однако самый старый образец R* был обнаружен в образце Древней Северной Евразии из Сибири ( Мальтийский мальчик , 24 000 лет назад), а его предшественник P1 был обнаружен в другом образце Древней Северной Евразии из Северной Сибири ( Яна RHS ), датируемом из ц. 31600 лет назад. [30]
Три генетических исследования в 2015 году подтвердили гипотезу курганскую Марии Гимбутас относительно протоиндоевропейской родины . Согласно этим исследованиям, гаплогруппы R1b-M269 и R1a, наиболее распространенные сейчас в Европе (R1a также распространена в Южной Азии), должны были распространиться из Западной Евразийской Степи вместе с индоевропейскими языками ; они также обнаружили аутосомный компонент, присутствующий у современных европейцев, которого не было у европейцев эпохи неолита , который должен был быть введен с отцовскими линиями R1b и R1a, а также с индоевропейскими языками. [2] [3] [4]
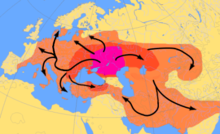
Анализ древней Y-ДНК из останков из поселений культуры линейной керамики раннего неолита Центральной и Северной Европы пока не обнаружил мужчин, принадлежащих к гаплогруппе R1b-M269. [31] [32] Олальде и др. (2017) связывают распространение гаплогруппы R1b-M269 в Западной Европе, особенно в Великобритании, с распространением культуры кубков с внезапным появлением многих гаплогрупп R1b-M269 в Западной Европе ок. 5000–4500 лет назад, в эпоху ранней бронзы. [33]
Структура
[ редактировать ]Внешняя филогения R1b
[ редактировать ]Более широкая гаплогруппа R (M207) является первичным субкладом гаплогруппы P1 (M45), которая сама по себе является первичной ветвью P (P295), которая также известна как гаплогруппа K2b2. Таким образом, R-M207 является вторичной ветвью K2b (P331) и прямым потомком K2 (M526).
произошла «первоначальная быстрая диверсификация» K-M526 По словам Карафета и др., . (2014), что «вероятно произошло в Юго-Восточной Азии с последующей экспансией на запад предков гаплогрупп R и Q ».
- Филогения внутри K2b
- P P295/PF5866/S8 (также известный как K2b2 ).
Внутренняя структура R1b
[ редактировать ]Такие имена, как R1b, R1b1 и т. д., являются филогенетическими (т.е. «генеалогическим древом») именами, которые проясняют их место в разветвлении гаплогрупп или филогенетическом дереве. Альтернативный способ обозначения тех же гаплогрупп и субкладов относится к их определяющим SNP- мутациям: например, R-M343 эквивалентен R1b. [34] Филогенетические названия меняются с новыми открытиями , и имена на основе SNP, следовательно, переклассифицируются в филогенетическом дереве. В некоторых случаях SNP оказывается ненадежным в качестве определяющей мутации, и имя на основе SNP полностью удаляется. Например, до 2005 года R1b был синонимом R-P25, который позже был реклассифицирован как R1b1; в 2016 году R-P25 был полностью удален как определяющий SNP из-за значительной частоты обратных мутаций. [35] (Ниже приведена базовая схема R1b в соответствии с деревом ISOGG по состоянию на 30 января 2017 года. [36] )
|
Географическое распространение
[ редактировать ]R1b* (Р-М343*)
[ редактировать ]не сообщалось о подтвержденных случаях R1b* (R-M343*), то есть R1b (xR1b1, R1b2), также известного как R-M343 (xL754, PH155) В рецензируемой литературе .
- Р-М343 (хМ73, М269, В88)
В ранних исследованиях, поскольку R-M269, R-M73 и R-V88 на сегодняшний день являются наиболее распространенными формами R1b, иногда предполагалось, что примеры R1b (xM73, xM269) обозначают базальные примеры «R1b *». [35] Однако, хотя парагруппа R-M343 (xM73, M269, V88) встречается редко, это не исключает членства в редких и/или обнаруженных впоследствии относительно базальных субкладах R1b, таких как R-L278* (R1b*), R -L389* (R1b1a*), R-P297* (R1b1a1*), R-V1636 (R1b1a2) или R-PH155 (R1b2).
Считается, что наибольшая доля R-M343 (xM73, M269, V88) принадлежит курдам юго-восточного Казахстана - 13%. [41] Однако совсем недавно крупное исследование вариаций Y-хромосомы в Иране показало, что R-M343 (xV88, M73, M269) достигает 4,3% среди иранских субпопуляций. [42]
Остается вероятность того, что некоторые или даже большинство из этих случаев могут быть R-L278* (R1b*), R-L389* (R1b1a*), R-P297* (R1b1a1*), R-V1636 (R1b1a2), R-PH155 (R1b2), R1b* (R-M343*), R1a* (R-M420*), иначе недокументированная ветвь R1 (R-M173) и/или обратные мутации маркера, от положительного результата. к отрицательному наследственному состоянию, [43] и, следовательно, составляют недокументированные субклады R1b.
Подборку предыдущих исследований распределения R1b можно найти у Cruciani et al. (2010). [44] Это суммировано в следующей таблице. (Кручиани не включил некоторые исследования, предполагающие еще более высокую частоту R1b1a1b [R-M269] в некоторых частях Западной Европы.)
| Континент | Область | Образец размер | Всего 1 млрд руб. | Р-П25 (ненадежный маркер для Р1б1*) | Р-В88 (Р1б1б) | Р-М269 (R1b1a1a2) | Р-М73 (R1b1a1a1) |
|---|---|---|---|---|---|---|---|
| Африка | Северная Африка | 691 | 5.9% | 0.0% | 5.2% | 0.7% | 0.0% |
| Африка | Центральный Сахельский регион | 461 | 23.0% | 0.0% | 23.0% | 0.0% | 0.0% |
| Африка | Западная Африка | 123 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| Африка | Восточная Африка | 442 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| Африка | Южная Африка | 105 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| Европа | Западные европейцы | 465 | 57.8% | 0.0% | 0.0% | 57.8% | 0.0% |
| Европа | Северо-западные европейцы | 43 | 55.8% | 0.0% | 0.0% | 55.8% | 0.0% |
| Европа | Центральноевропейцы | 77 | 42.9% | 0.0% | 0.0% | 42.9% | 0.0% |
| Европа | Северо-восточные европейцы | 74 | 1.4% | 0.0% | 0.0% | 1.4% | 0.0% |
| Европа | русские | 60 | 6.7% | 0.0% | 0.0% | 6.7% | 0.0% |
| Европа | Восточные европейцы | 149 | 20.8% | 0.0% | 0.0% | 20.8% | 0.0% |
| Европа | Юго-восточные европейцы | 510 | 13.1% | 0.0% | 0.2% | 12.9% | 0.0% |
| Азия | выходцы из Западной Азии | 328 | 5.8% | 0.0% | 0.3% | 5.5% | 0.0% |
| Азия | жители Южной Азии | 288 | 4.8% | 0.0% | 0.0% | 1.7% | 3.1% |
| Азия | Юго-восточные азиаты | 10 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| Азия | Северо-восточные азиаты | 30 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| Азия | Восточные азиаты | 156 | 0.6% | 0.0% | 0.0% | 0.6% | 0.0% |
| Общий | 5326 | ||||||
R1b (R-L278)
[ редактировать ]R-L278 среди современных мужчин относится к субкладам R-L754 и R-PH155, хотя возможно, что некоторые очень редкие R-L278* могут существовать, поскольку не все примеры были протестированы для обеих ветвей. Примеры могут также существовать в древней ДНК, хотя из-за низкого качества часто невозможно определить, несли ли древние мутации, определяющие субклады.
Некоторые примеры описаны в старых статьях, например два найдены в Турции. [34] Сейчас считается, что они в основном относятся к недавно открытому субкладу R1b1b (R-V88). Таким образом, большинство примеров R1b попадают в субклады R1b1b (R-V88) или R1b1a (R-P297). Кручиани и др. В крупном исследовании 2010 года было обнаружено 3 случая среди 1173 итальянцев, 1 из 328 жителей Западной Азии и 1 из 156 жителей Восточной Азии. [44] Варзари обнаружил 3 случая в Украине при исследовании 322 человек из региона Днестровско - Карпатских гор , у которых был положительный результат P25, но отрицательный результат M269. [45] Случаи из более ранних исследований происходят в основном из Африки, Ближнего Востока или Средиземноморья и обсуждаются ниже как вероятные случаи R1b1b (R-V88).
R1b1 (R-L754)
[ редактировать ]R-L754 содержит подавляющее большинство R1b. Единственный известный пример R-L754* (xL389, V88) также является самым ранним известным человеком, носителем R1b: « Вильябруна 1 », который жил около 14 000 лет назад (северо-восток Италии). Виллабруна 1 принадлежала к эпиграветтской культуре.
R1b1a (R-L389)
[ редактировать ]R-L389, также известный как R1b1a (L388/PF6468, L389/PF6531), содержит очень распространенный субклад R-P297 и редкий субклад R-V1636. Неизвестно, принадлежат ли все ранее зарегистрированные R-L389* (xP297) к R-V1636 или нет.
R1b1a1 (Р-П297)
[ редактировать ]Маркер SNP P297 был признан в 2008 году предком значимых субкладов M73 и M269, объединив их в один кластер. [7] Ему было присвоено филогенетическое название R1b1a1a (а ранее - R1b1a).
Большая часть евразийских R1b попадает в этот субклад, представляя очень большую современную популяцию. Хотя сам P297 еще не подвергался серьезным исследованиям, та же популяция относительно хорошо изучена с точки зрения других маркеров. Следовательно, ветвление внутри этой клады можно относительно подробно объяснить ниже.
Р1б1а1а (Р-М73)
[ редактировать ]Малярчук и др. (2011) обнаружили R-M73 у 13,2% (5/38) шорцев, 11,4% (5/44) телеутов, 3,3% (2/60) калмыков, 3,1% (2/64) хакасов, 1,9% (2/108) тувинцев и 1,1% (1/89) алтайцев. [46] Калмыки, тувинцы и алтайцы принадлежат к кластеру Y-STR, отмеченному DYS390=19, DYS389=14–16 (или 14–15 в случае алтайца) и DYS385=13–13.
Дулик и др. (2012) обнаружили R-M73 у 35,3% (6/17) выборки кумандинцев Республики Алтай в России. [47] Трое из этих шести кумандинцев имеют одинаковый гаплотип Y-STR из 15 локусов, а еще двое отличаются только локусом DYS458, имея DYS458=18 вместо DYS458=17. Эта пара кумандинских гаплотипов R-M73 напоминает гаплотипы двух калмыков, двух тувинцев и одного алтайца, Y-ДНК которого была проанализирована Малярчуком и др. (2011). Оставшийся кумандинец R-M73 имеет гаплотип Y-STR, который резко отличается от гаплотипов других кумандинцев R-M73, напоминая вместо этого гаплотипы пяти шорцев, пяти телеутов и двух хакасов. [46]
Хотя ранние исследования R-M73 утверждали, что он был широко представлен среди хазарейцев Афганистана и башкир Урала, это, по-видимому, было опровергнуто. Например, вспомогательный материал исследования Behar et al., проведенного в 2010 году. предположили, что Сенгупта и др. (2006) могли ошибочно идентифицировать хазарейцев, которые вместо этого принадлежали к «PQR2», а не к «R(xR1a)». [48] [41] [49] Однако отнесение Y-ДНК этих хазарейцев к категории «PQR2» Бехаром и др. (2010), вероятно, объясняется популярной в течение некоторого времени привычкой обозначать R-M269 как «R1b» или «R(xR1a)», при этом любые члены R-M343 (xM269) помещаются в полифилетическую, привлекательную группу. все категории «R*» или «P». Майрес и др. (2011), Ди Кристофаро и др. (2013) и Липпольд и др. (2014) все согласны с тем, что Y-ДНК 32% (8/25) образца HGDP пакистанских хазарейцев должна принадлежать к гаплогруппе R-M478/M73. [41] [50] [51] Аналогично, было обнаружено, что большинство башкирских мужчин принадлежат к U-152 (R1b1a1a2a1a2b), а некоторые, в основном из юго-восточного Башкортостана, принадлежат к гаплогруппе Q-M25 (Q1a1b), а не к R1b; против этого Myres et al. (2011) обнаружили высокую частоту R-M73 среди своей выборки башкир юго-восточного Башкортостана (77/329 = 23,4% R1b-M73), что согласуется с более ранним исследованием башкир. [41] Помимо высокой частоты R-M73 у юго-восточных башкир, Myres et al. также сообщили об обнаружении R-M73 в следующих образцах: 10,3% (14/136) балкарцев с северо-западного Кавказа, 9,4% (8/85) образцов HGDP из северного Пакистана (это вышеупомянутые пакистанские хазарейцы), 5,8% (4/69) карачаевцев с северо-западного Кавказа, 2,6% (1/39) татар из Башкортостана, 1,9% (1/54) башкир с юго-запада Башкортостана, 1,5% (1/67) мегрелов с Южного Кавказа , 1,4% (1/70) башкир северного Башкортостана, 1,3% (1/80) татар из Казани, 1,1% (1/89) выборки из Каппадокии, Турция, 0,7% (1/141) кабардинцев с северо-западного Кавказа, 0,6% (3/522) пула выборок из Турции и 0,38% (1/263) русских из Центральной России. [41]
Помимо вышеупомянутых пакистанских хазарейцев, Ди Кристофаро и др. (2013) обнаружили R-M478/M73 у 11,1% (2/18) монголов из центральной Монголии, 5,0% (1/20) кыргызов с юго-запада Кыргызстана, 4,3% (1/23) монголов из юго-восточной Монголии, 4,3 % (4/94) узбеков из Джаузжана, Афганистан, 3,7% (1/27) иранцев из Гилана , 2,5% (1/40) кыргызов из центрального Кыргызстана, 2,1% (2/97) монголов из северо-западной Монголии и 1,4% (1/74) туркмен из Джаузджана, Афганистан. [50] Монголы, а также человек из юго-западного Кыргызстана, человек из Гилана и один из узбеков из Джауздана принадлежат к тому же кластеру гаплотипов Y-STR, что и пять из шести кумандинцев R-M73, изученных Дуликом и др. (2012). Наиболее характерное значение Y-STR этого кластера — DYS390=19. [41]
Карафет и др. (2018) обнаружили R-M73 у 37,5% (15/40) выборки Телеутов из Беково Кемеровской области, 4,5% (3/66) выборки уйгуров Синьцзян-Уйгурского автономного района, 3,4% (1/29 ) выборки казахов из Казахстана, 2,3% (3/129) выборки селькупов, 2,3% (1/44) выборки туркмен из Туркменистана и 0,7% (1/136) выборки иранцев из Ирана. [52] Четверо из этих людей (один из телеутов, один из уйгуров, казах и иранец), по-видимому, принадлежат к вышеупомянутому кластеру, отмеченному DYS390=19 (кумандинско-монгольский кластер R-M73); Телеуты и уйгуры также имеют общие модальные значения в локусах DYS385 и DYS389. Иранский отличается от модального для этого кластера наличием 13–16 (или 13–29) в DYS389 вместо 14–16 (или 14–30). Казахский отличается от модального наличием 13–14 в DYS385 вместо 13–13. Остальные четырнадцать Телеутов и трое селькупов, судя по набору данных Малярчука и др., принадлежат к Телеут-Шор-Хакасскому кластеру R-M73. (2011); этот кластер имеет модальные значения DYS390=22 (но 21 в случае двух телеутов и одного хакаса), DYS385=13-16 и DYS389=13-17 (или 13–30, но 14–31 в случае один селькуп).
В казахстанской статье, опубликованной в 2017 году, гаплогруппа R1b-M478 Y-ДНК обнаружена у 3,17% (41/1294) выборки казахов из Казахстана, при этом эта гаплогруппа наблюдается с частотой выше средней среди представителей кыпшаков (12/29 = 41,4%), Ысты (6/57 = 10,5%), Конгырат (8/95 = 8,4%), Ошакты (2/29 = 6,9%), Керей (1/28 = 3,6%) и Джетыру (3/86). = 3,5%) племен . [53] Китайская статья, опубликованная в 2018 году, обнаружила гаплогруппу R1b-M478 Y-ДНК у 9,2% (7/76) выборки уйгуров Долан из поселка Хорикол уезда Ават в Синьцзяне. [54]
R1b1a1b (R-M269)
[ редактировать ]R-M269 или R1b1a1b (по состоянию на 2018 год) среди других имен, [55] в настоящее время является наиболее распространенной линией Y-ДНК у европейских мужчин. В Европе его носителями являются около 110 миллионов мужчин. [56]

R-M269 вызвал значительный научный и популярный интерес из-за его возможной связи с индоевропейской экспансией в Европе. В частности, было обнаружено, что субклад R-Z2103 преобладает в древней ДНК, связанной с культурой Ямна . [2] Все семь особей в одной были определены как принадлежащие к субкладу R1b-M269. [2]
Более ранние исследования, опубликованные до того, как исследователи смогли изучить ДНК древних останков, предположили, что R-M269, вероятно, возник в Западной Азии и присутствовал в Европе в период неолита. [36] [41] [57] [58] Но результаты, основанные на реальной древней ДНК, показали, что до бронзового века в Европе был дефицит R-M269. [2] а распространение субкладов внутри Европы в значительной степени обусловлено различными миграциями бронзового и железного веков . Аналогичным образом, самые старые образцы, отнесенные к R-M269, были найдены в Восточной Европе и Понтийско-Каспийской степи, а не в Западной Азии. Популяции Западной Европы разделены на субклады R-P312/S116 и R-U106/S21 R-M412 (R-L51).
Распространение R-M269 в Европе увеличивается с востока на запад. Пик на национальном уровне он достигает в Уэльсе в размере 92%, в Ирландии 82% , в Шотландии 70% , в Испании 68% , во Франции 60% (76% в Нормандии ), около 60% в Португалии , [41] 50% в Германии , 50% в Нидерландах , 47% в Италии , [59] 45% в Восточной Англии и 42% в Исландии .Уровень R-M269 достигает 95% в некоторых частях Ирландии. Он также был обнаружен с более низкой частотой по всей Центральной Евразии . [60] но с относительно высокой частотой среди башкир Пермского края (84,0%). [61] Этот маркер присутствует в Китае и Индии с частотой менее одного процента. В Северной Африке и на прилегающих островах, хотя R-V88 (R1b1b) представлен более широко, R-M269, по-видимому, присутствовал с древних времен. R-M269 был обнаружен, например, в количестве около 44% среди останков, датируемых 11-13 веками в Пунта-Азуле , на Канарских островах . Эти останки были связаны с бимбаче (или бимапе), подгруппой гуанчей. [62] У ныне живущих мужчин он достигает максимума в некоторых частях Северной Африки, особенно в Алжире , и составляет 10%. [63] В странах Африки к югу от Сахары пик R-M269, по-видимому, наблюдается в Намибии , где его уровень составляет 8% среди мужчин гереро . [64] В Западной Азии R-M269 зарегистрирован у 40% армянских мужчин и более чем у 35% туркменских мужчин. [65] [66] (В таблице ниже более подробно перечислены частоты M269 в регионах Азии, Европы и Африки.)
Помимо базальной R-M269*, которая не разошлась, существуют (по состоянию на 2017 год) две основные ветви R-M269:
- R-L23 (R1b1a1b1; L23/PF6534/S141) и
- Р-ПФ7558 (Р1б1а1б2; ПФ7558/ПФ7562.)
R-L23 (Z2105/Z2103; он же R1b1a1b1) отмечен среди народов Идель -Урала (по Трофимова и др., 2015): у 21 из 58 (36,2%) башкир Бурзянского района , у 11 из 52 (21,2%). %) удмуртов , 4 из 50 (8%) коми , 4 из 59 (6,8%) мордвы , 2 из 53 (3,8%) бесермянцев и 1 из 43 (2,3%) чувашей были R1b -Л23. [67]
Субклады внутри парагруппы R-M269(xL23) – то есть R-M269* и/или R-PF7558 – по-видимому, наиболее часто встречаются на центральных Балканах , особенно в Косово (7,9%), Северной Македонии ( 5,1%) и Сербии. 4,4%. [41] В отличие от большинства других территорий со значительным процентом R-L23, Косово , Польша и башкиры юго-восточного Башкортостана отличаются высоким процентом R-L23 (xM412) – при показателях 11,4% (Косово), 2,4% ( Польша) и 2,4% юго-восток Башкортостана. [41] (Эта башкирская популяция также отличается высоким уровнем R-M73 (R1b1a1a1) - 23,4%. [41] ) Пять особей из 110, протестированных в Араратской долине Армении , принадлежали к R-M269(xL23) и 36 к R-L23*, при этом ни один не принадлежал к известным субкладам L23. [68]
В 2009 году ДНК, извлеченная из бедренных костей шести скелетов в раннесредневековом захоронении в Эргольдинге (Бавария, Германия), датированная примерно 670 годом нашей эры, дала следующие результаты: было обнаружено, что 4 относятся к гаплогруппе R1b с наиболее близкими совпадениями в современных популяциях. Германии, Ирландии и США, а двое принадлежали к гаплогруппе G2a . [69]
Ниже приводится краткое изложение большинства исследований, в которых конкретно проверялось наличие M269, показывающее его распространение (в процентах от общей численности населения) в Европе, Северной Африке , на Ближнем Востоке и в Центральной Азии, вплоть до Китая и Непала .
Филогения R-M269 по данным ISOGG 2017:
| M269/PF6517 |
| |||||||||||||||||||||||||||||||||||||||||||||
R1b1b (Р-В88)
[ редактировать ]R1b1b (PF6279/V88; ранее R1b1a2) определяется наличием маркера SNP V88, об открытии которого было объявлено в 2010 году Cruciani et al. [44] Помимо особей в Южной Европе и Западной Азии, большая часть R-V88 была обнаружена в Сахеле, особенно среди популяций, говорящих на языках чадской афроазиатских ветви.
На основании подробного филогенетического анализа D'Atanasio et al. (2018) предположили, что R1b-V88 возник в Европе около 12 000 лет назад и перешел в Северную Африку между 8000 и 7000 лет назад, в период « Зеленой Сахары ». R1b-V1589, основной субклад внутри R1b-V88, претерпел дальнейшее расширение около 5500 лет назад, вероятно, в районе бассейна озера Чад, откуда некоторые линии повторно пересекли Сахару в Северную Африку. [70]
Маркус и др. (2020) предоставляют убедительные доказательства этой предлагаемой модели транссахарского движения с севера на юг: самые ранние базальные гаплогруппы R1b-V88 обнаружены у нескольких восточноевропейских охотников-собирателей около 11 000 лет назад. Затем гаплогруппа, по-видимому, распространилась с экспансией неолитических фермеров , которые основали сельское хозяйство в Западном Средиземноморье примерно к 7500 году до нашей эры. Гаплогруппы R1b-V88 были идентифицированы у древних людей эпохи неолита в Германии, центральной Италии, Иберии и, особенно часто, на Сардинии. Часть ветви, ведущей к современным африканским гаплогруппам (V2197), уже произошла от неолитических европейцев из Испании и Сардинии, что обеспечило дополнительную поддержку транссахарскому движению с севера на юг. [71] [72] [73] Европейское аутосомное происхождение, гаплогруппы мтДНК и аллели персистенции лактазы также были идентифицированы в африканских популяциях, которые несут R1b-V88 с высокой частотой, таких как фулани и тубу . [74] [75] [72] [76] Присутствие европейских земледельцев эпохи неолита в Африке дополнительно подтверждается образцами из Марокко, датируемыми ок. 5400 г. до н.э. и далее. [77] [78]
Исследования 2005–2008 годов показали высокий уровень R1b* в Иордании , Египте и Судане . [79] [64] [80] [примечание 1] Последующие исследования Myres et al. (2011) указывает, что рассматриваемые образцы, скорее всего, принадлежат субкладу R-V88. По данным Майреса и др. (2011), это можно объяснить обратной миграцией из Азии в Африку людей-носителей R1b. [41] [примечание 2]
В отличие от других исследований, Шрайнер и Ротими (2018) связали появление R1b в Чаде с недавними перемещениями арабов Баггара . [81]
| Область | Население | Страна | Язык | Образец размер | Общий % | R1b1b (Р-В88) | R1b1a1b (Р-М269) | R1b1b* (Р-В88*) | R1b1b2a2a1 (Р-В69) |
|---|---|---|---|---|---|---|---|---|---|
| Северная Африка | Композитный | Марокко | АА | 338 | 0.0% | 0.3% | 0.6% | 0.3% | 0.0% |
| Северная Африка | Мозабитские берберы | Алжир | АА/берберский | 67 | 3.0% | 3.0% | 0.0% | 3.0% | 0.0% |
| Северная Африка | Северные египтяне | Египет | АА/семитский | 49 | 6.1% | 4.1% | 2.0% | 4.1% | 0.0% |
| Северная Африка | Берберы из Сивы | Египет | АА/берберский | 93 | 28.0% | 26.9% | 1.1% | 23.7% | 3.2% |
| Северная Африка | Моряк | Египет | АА/семитский | 41 | 7.3% | 4.9% | 2.4% | 0.0% | 4.9% |
| Северная Африка | Гурна Оазис | Египет | АА/семитский | 34 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| Северная Африка | Южные египтяне | Египет | АА/семитский | 69 | 5.8% | 5.8% | 0.0% | 2.9% | 2.9% |
| C Африка | Сонгай | Нигер | NS/Сонхай | 10 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| C Африка | Фульбе | Нигер | Северная Каролина/Атлантика | 7 | 14.3% | 14.3% | 0.0% | 14.3% | 0.0% |
| C Африка | Туареги | Нигер | АА/берберский | 22 | 4.5% | 4.5% | 0.0% | 4.5% | 0.0% |
| C Африка | Волна | Чад | НС/Суданский | 11 | 9.1% | 9.1% | 0.0% | 9.1% | 0.0% |
| C Африка | Хауса | Нигерия (Север) | АА/Чадик | 10 | 20.0% | 20.0% | 0.0% | 20.0% | 0.0% |
| C Африка | Фульбе | Нигерия (Север) | Северная Каролина/Атлантика | 32 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| C Африка | Йоруба | Нигерия (Юг) | NC/Дефоид | 21 | 4.8% | 4.8% | 0.0% | 4.8% | 0.0% |
| C Африка | Ульдеме | Камерун (Северный) | АА/Чадик | 22 | 95.5% | 95.5% | 0.0% | 95.5% | 0.0% |
| C Африка | Тема | Камерун (Северный) | АА/Чадик | 17 | 82.4% | 82.4% | 0.0% | 76.5% | 5.9% |
| C Африка | Мафа | Камерун (Северный) | АА/Чадик | 8 | 87.5% | 87.5% | 0.0% | 25.0% | 62.5% |
| C Африка | Гуизига | Камерун (Северный) | АА/Чадик | 9 | 77.8% | 77.8% | 0.0% | 22.2% | 55.6% |
| C Африка | Назад | Камерун (Северный) | АА/Чадик | 19 | 42.1% | 42.1% | 0.0% | 36.8% | 5.3% |
| C Африка | Путеводители | Камерун (Северный) | АА/Чадик | 9 | 66.7% | 66.7% | 0.0% | 22.2% | 44.4% |
| C Африка | Масса | Камерун (Северный) | АА/Чадик | 7 | 28.6% | 28.6% | 0.0% | 14.3% | 14.3% |
| C Африка | Другой Чадский | Камерун (Северный) | АА/Чадик | 4 | 75.0% | 75.0% | 0.0% | 25.0% | 50.0% |
| C Африка | Шуа арабы | Камерун (Северный) | АА/семитский | 5 | 40.0% | 40.0% | 0.0% | 40.0% | 0.0% |
| C Африка | Канури | Камерун (Северный) | Северная Каролина/Сахара | 7 | 14.3% | 14.3% | 0.0% | 14.3% | 0.0% |
| C Африка | Фульбе | Камерун (Северный) | Северная Каролина/Атлантика | 18 | 11.1% | 11.1% | 0.0% | 5.6% | 5.6% |
| C Африка | ОСТАНАВЛИВАТЬСЯ | Камерун (Северный) | Северная Каролина/Адамава | 21 | 66.7% | 66.7% | 0.0% | 14.3% | 52.4% |
| C Африка | Волна | Камерун (Северный) | Северная Каролина/Адамава | 48 | 20.8% | 20.8% | 0.0% | 10.4% | 10.4% |
| C Африка | Такой | Камерун (Северный) | Северная Каролина/Адамава | 22 | 9.1% | 9.1% | 0.0% | 4.5% | 4.5% |
| C Африка | Мбум | Камерун (Северный) | Северная Каролина/Адамава | 9 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| C Африка | Композитный | Камерун (Sth) | Северная Каролина/банту | 90 | 0.0% | 1.1% | 0.0% | 1.1% | 0.0% |
| C Африка | Как пигмеи | МАШИНА | Северная Каролина/банту | 33 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| Западная Африка | Композитный | — | 123 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | |
| Это Африка | Композитный | — | 442 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | |
| Южная Африка | Композитный | — | 105 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | |
| Общий | 1822 | ||||||||
| В88 |
| ||||||||||||
Две ветви R-V88, R-M18 и R-V35, встречаются почти исключительно на острове Сардиния .
Как видно из приведенной выше таблицы данных, R-V88 с очень высокой частотой встречается в северном Камеруне и на западе центральной Африки, где считается, что он вызван доисламским перемещением людей из Евразии . [64] [82]
R1b1b1 (Р-М18)
[ редактировать ]R1b1b1 представляет собой субклад R-V88, который определяется наличием маркера SNP M18. [7] Он был обнаружен только на низких частотах в образцах с Сардинии. [60] [83] и Ливан . [84]
R1b2 (Р-PH155)
[ редактировать ]R1b2 встречается крайне редко и определяется наличием PH155. [36] Живые самцы, несущие субклады R-PH155, были обнаружены в Бахрейне , Бутане , Ладакхе , Таджикистане , Турции, Синьцзяне и Юньнани . ISOGG (2022) называет две основные ветви: R-M335 (R1b2a) и R-PH200 (R1b2b).
Определяющий SNP R1b2a, M335, был впервые задокументирован в 2004 году, когда экземпляр был обнаружен в Турции, хотя в то время он классифицировался как R1b4. [34] Другие примеры R-M335 были обнаружены в образце Хуэй из Юньнани , Китай. [85] и в выборке людей из Ладакха , Индия. [86]
В популярной культуре
[ редактировать ]- Брайан Сайкс в своей книге 2006 года «Кровь островов » дает членам – и условному патриарху-основателю – R1b имя « Оисин ».
- Стивен Оппенгеймер в своей книге «Происхождение британцев » 2007 года дает патриарху R1b баскское имя «Руиско» в честь того, что Оппенгеймер считал иберийским происхождением R1b.
- Режиссер Артем Лукичев создал (около 2009 г.) 14-минутный анимационный фильм по мотивам башкирского эпоса Уральских гор , связывающий эпос с возникновением и географическим расширением R1a и R1b. [87]
- Тесты ДНК, которые помогли идентифицировать российского царя Николая II, показали, что он принадлежал к R1b. [88] Это может свидетельствовать о том, что более поздние цари Дома Романовых , произошедшие по мужской линии от Дома Гольштейн-Готторп (который возник в земле Шлезвиг-Гольштейн ), также являются членами R1b.
Известные люди
[ редактировать ]Спитихнев I, герцог Богемии , анализ ДНК его останков предполагает, что его Y-гаплогруппа была R1b.
Дом Бурбонов , который правил во Франции, Испании и других европейских странах, имеет гаплогруппу R1b1b. [89]
Здоровье
[ редактировать ]Исследования показали, что гаплогруппа R1b может оказывать защитное действие на иммунную систему. [90] Однако более поздние исследования подтвердили, что Y-хромосома оказывает очень ограниченное влияние, например, на ишемическую болезнь сердца (ИБС), и что ранее предполагаемая связь между гаплогруппами Y-хромосомы и здоровьем далека от научного подтверждения. [91]
См. также
[ редактировать ]Примечания
[ редактировать ]- ^ Флорес и др. (2005) обнаружили, что 20 из всех 146 протестированных мужчин (13,7%) – включая 20 из 45 мужчин, протестированных из района Мертвого моря в Иордании – были положительными на M173 (R1) и отрицательными для обоих маркеров R1a SRY10831.2 и M17, а также P25 (который позже оказался ненадежным маркером R1b1). Майрес и др. (2011) [41] указывает на то, что это в основном R-V88 (позже известный как R1b1b). Вуд и др. (2005) также сообщили о двух египетских случаях R1* (R-M173*), которые были отрицательными для SRY10831 (R1a1) и ненадежного маркера R1b1 P25, из выборки, состоящей из 1122 мужчин из африканских стран, включая 92 из Египта. Хасан и др. (2008) обнаружили столь же удивительные 14 из 26 (54%) суданских людей фула, которые имели M173+ и P25-.
- ^ Майрес и др. (2011): «Обнаружение V88 в Иране, Палестине и особенно на Мертвом море и в Иордании (дополнительная таблица S4) дает представление об обратном пути миграции в Африку.
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с Фу К., Пост С., Хайдиняк М., Петр М., Маллик С., Фернандес Д. и др. (июнь 2016 г.). «Генетическая история Европы ледникового периода» . Природа . 534 (7606): 200–5. Бибкод : 2016Natur.534..200F . дои : 10.1038/nature17993 . hdl : 10211.3/198594 . ПМЦ 4943878 . ПМИД 27135931 .
- ^ Перейти обратно: а б с д и ж Хаак и др. 2015 .
- ^ Перейти обратно: а б Аллентофт М.Э., Сикора М., Шегрен К.Г., Расмуссен С., Расмуссен М., Стендеруп Дж. и др. (июнь 2015 г.). «Популяционная геномика Евразии бронзового века» . Природа . 522 (7555): 167–72. Бибкод : 2015Natur.522..167A . дои : 10.1038/nature14507 . ПМИД 26062507 . S2CID 4399103 .
- ^ Перейти обратно: а б Мэтисон И., Лазаридис И., Роланд Н., Маллик С., Паттерсон Н., Руденберг С.А. и др. (2015). «Восемь тысяч лет естественного отбора в Европе». bioRxiv : 016477. doi : 10.1101/016477 . S2CID 7866359 .
- ^ Кэссиди Л.М., Мартиниано Р., Мерфи Э.М., Тисдейл М.Д., Мэллори Дж., Хартвелл Б., Брэдли Д.Г. (январь 2016 г.). «Миграция в Ирландию в эпоху неолита и бронзового века и установление островного атлантического генома» . Труды Национальной академии наук Соединенных Штатов Америки . 113 (2): 368–73. Бибкод : 2016PNAS..113..368C . дои : 10.1073/pnas.1518445113 . ПМК 4720318 . ПМИД 26712024 .
- ^ Мартиниано Р., Кэссиди Л.М., О'Маолдуин Р., Маклафлин Р., Сильва Н.М., Манко Л. и др. (июль 2017 г.). «Популяционная геномика археологического перехода в западной Иберии: исследование древней субструктуры с использованием методов вменения и гаплотипов» . ПЛОС Генетика . 13 (7): e1006852. дои : 10.1371/journal.pgen.1006852 . ПМЦ 5531429 . ПМИД 28749934 .
- ^ Перейти обратно: а б с Карафет Т.М., Мендес Ф.Л., Мейлерман М.Б., Андерхилл П.А., Зегура С.Л., Хаммер М.Ф. (май 2008 г.). «Новые бинарные полиморфизмы изменяют форму и увеличивают разрешение дерева гаплогрупп Y-хромосомы человека» . Геномные исследования . 18 (5): 830–8. дои : 10.1101/гр.7172008 . ПМК 2336805 . ПМИД 18385274 .
- ^ Мэтисон 2018 , Дополнительная таблица 1, строка 467.
- ^ Мэтисон 2018 , Дополнительная таблица 1, строки 251–272.
- ^ Перейти обратно: а б Джонс 2017 .
- ^ Мэтисон 2018 , Дополнительная таблица 1, строки 205–245.
- ^ Мэтисон 2018 , Дополнительная таблица 1.
- ^ Мэтисон 2018 , стр. 2–3.
- ^ Гонсалес-Фортес и др. 2017 , стр. 8–9.
- ^ Гонсалес-Фортес и др. 2017 , с. 4, таблица 1, OC1_Meso.
- ^ Санчес-Куинто и др. 2019 , Расширенный набор данных 1.3, OC1.
- ^ Мэтисон 2018 , Дополнительная таблица 1, строка 298, I4666.
- ^ Мэтисон 2018 , Дополнительная таблица 1, строка 153, I0124.
- ^ Мэтисон 2018 , Дополнительная таблица 1, строка 375, I0122.
- ^ Хаак и др. 2015 , Дополнительная информация, стр. 107-111. 26-2
- ^ Хаак и др. 2015 , Таблица расширенных данных 2, I0410.
- ^ Хаак и др. 2015 , Дополнительная информация, стр. 101-1. 44-4
- ^ Мэтисон 2018 , Дополнительная таблица 1, I0410.
- ^ Мэтисон 2018 , Дополнительная таблица 1, строка 73, I2430.
- ^ Маркус и др. 2020 , Дополнительные данные 1, Основная таблица, строка 25, MA89.
- ^ Мэтисон 2018 , Дополнительная таблица 1, строка 128, I0559.
- ^ де Баррос Дамгаард П., Мартиниано Р., Камм Дж., Морено-Майар Дж.В., Кроонен Г., Пейро М. и др. (июнь 2018 г.). «Первые пастухи и влияние степной экспансии раннего бронзового века в Азию» . Наука . 360 (6396): eaar7711. дои : 10.1126/science.aar7711 . ПМК 6748862 . ПМИД 29743352 .
- ^ Хаак и др. 2015 , с. 5.
- ^ Карафет Т.М., Мендес Ф.Л., Судойо Х., Лансинг Дж.С., Хаммер М.Ф. (март 2015 г.). «Улучшенное филогенетическое разрешение и быстрая диверсификация гаплогруппы Y-хромосомы K-M526 в Юго-Восточной Азии» . Европейский журнал генетики человека . 23 (3): 369–73. дои : 10.1038/ejhg.2014.106 . ПМЦ 4326703 . ПМИД 24896152 .
- ^ Сикора, Мартин; Питулко Владимир Владимирович; Соуза, Витор К.; Аллентофт, Мортен Э.; Победитель, Лассе; Расмуссен, Саймон; Маргарян, Ашот; де Баррос Дамгаард, Питер; Фонтана, Констанция; Рено, Габриэль; Ян, Мелинда А.; Фу, Цяомэй; Дюпанлуп, Изабель; Джампудакис, Константинос; Ногес-Браво, Давид (июнь 2019 г.). «История населения Северо-Восточной Сибири начиная с плейстоцена» . Природа . 570 (7760): 182–188. Бибкод : 2019Natur.570..182S . дои : 10.1038/s41586-019-1279-z . hdl : 1887/3198847 . ISSN 1476-4687 . ПМИД 31168093 . S2CID 174809069 .
- ^ Лакан М., Кейзер С., Рико Ф.К., Брукато Н., Дюрантон Ф., Гилейн Дж. и др. (июнь 2011 г.). «Древняя ДНК свидетельствует о распространении мужчин по неолитическому средиземноморскому маршруту» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (24): 9788–91. Бибкод : 2011PNAS..108.9788L . дои : 10.1073/pnas.1100723108 . ПМК 3116412 . ПМИД 21628562 .
- ^ Хаак В., Балановский О., Санчес Дж.Дж., Кошель С., Запорожченко В., Адлер С.Дж. и др. (ноябрь 2010 г.). Пенни Д. (ред.). «Древняя ДНК европейских фермеров раннего неолита раскрывает их ближневосточное родство» . ПЛОС Биология . 8 (11): е1000536. дои : 10.1371/journal.pbio.1000536 . ПМЦ 2976717 . ПМИД 21085689 .
- ^ Олальде И., Брейс С., Аллентофт М.Э., Армит И., Кристиансен К., Бут Т. и др. (март 2018 г.). «Феномен стакана и геномная трансформация северо-западной Европы» . Природа . 555 (7695): 190–196. Бибкод : 2018Natur.555..190O . дои : 10.1038/nature25738 . ПМЦ 5973796 . ПМИД 29466337 .
- ^ Перейти обратно: а б с Чинниоглу С., Кинг Р., Кивисилд Т., Калфоглу Е., Атасой С., Каваллери Г.Л. и др. (январь 2004 г.). «Раскопки слоев гаплотипов Y-хромосомы в Анатолии». Генетика человека . 114 (2): 127–48. дои : 10.1007/s00439-003-1031-4 . ПМИД 14586639 . S2CID 10763736 .
- ^ Перейти обратно: а б «ISOGG 2016 Гаплогруппа R Y-ДНК» . isogg.org .
- ^ Перейти обратно: а б с «ISOGG 2017 Гаплогруппа R Y-ДНК» . isogg.org .
- ^ YПолный YTree v10.00.00
- ^ Филогенетическое древо гаплогруппы R-V1636 на 23 мофанге.
- ^ Гаплогруппа R на гаплодереве Y-ДНК в FTDNA
- ^ «R-V1636 YTree» . www.yfull.com .
- ^ Перейти обратно: а б с д и ж г час я дж к л м Майрес Н.М., Роотси С., Лин А.А., Ярве М., Кинг Р.Дж., Кутуев И. и др. (январь 2011 г.). «Эффект основателя основной гаплогруппы Y-хромосомы R1b эпохи голоцена в Центральной и Западной Европе» . Европейский журнал генетики человека . 19 (1): 95–101. дои : 10.1038/ejhg.2010.146 . ПМК 3039512 . ПМИД 20736979 .
- ^ Гругни В., Батталья В., Хушиар Кашани Б., Пароло С., Аль-Захери Н., Ахилли А. и др. (18 июля 2012 г.). «Древние миграционные события на Ближнем Востоке: новые подсказки по вариации Y-хромосомы современных иранцев» . ПЛОС ОДИН . 7 (7): е41252. Бибкод : 2012PLoSO...741252G . дои : 10.1371/journal.pone.0041252 . ПМК 3399854 . ПМИД 22815981 .
- ^ Адамс С.М., Кинг Т.Е., Бош Э., Джоблинг М.А. (май 2006 г.). «Случай ненадежного SNP: рецидивирующая обратная мутация Y-хромосомного маркера P25 посредством конверсии гена» . Международная судебно-медицинская экспертиза . 159 (1): 14–20. doi : 10.1016/j.forsciint.2005.06.003 . HDL : 2381/443 . ПМИД 16026953 .
- ^ Перейти обратно: а б с Кручиани Ф., Тромбетта Б., Селитто Д., Массайя А., Дестро-Бисол Г., Уотсон Е. и др. (июль 2010 г.). «Гаплогруппа R-V88 Y-хромосомы человека: отцовская генетическая запись транссахарских связей начала среднего голоцена и распространения чадских языков» . Европейский журнал генетики человека . 18 (7): 800–7. дои : 10.1038/ejhg.2009.231 . ПМЦ 2987365 . ПМИД 20051990 .
- ^ Варзари А (2006). Популяционная история Днестровско-Карпатского региона: данные инсерции Alu и полиморфизмов Y-хромосомы (Диссертация). Мюнхенский университет Людвига-Максимилиана. дои : 10.5282/edoc.5868 .
- ^ Перейти обратно: а б Малярчук Б, Деренко М, Денисова Г, Максимов А, Возняк М, Гжибовский Т и др. (август 2011 г.). «Древние связи между сибиряками и коренными американцами, выявленные путем подтипирования гаплогруппы Q1a Y-хромосомы» . Журнал генетики человека . 56 (8): 583–8. дои : 10.1038/jhg.2011.64 . ПМИД 21677663 . S2CID 12015336 .
- ^ Дулик М.С., Жаданов С.И., Осипова Л.П., Аскапули А., Гау Л., Гоккумен О. и др. (февраль 2012 г.). «Вариации митохондриальной ДНК и Y-хромосомы свидетельствуют о недавнем общем происхождении коренных американцев и коренных алтайцев» . Американский журнал генетики человека . 90 (2): 229–46. дои : 10.1016/j.ajhg.2011.12.014 . ПМК 3276666 . ПМИД 22281367 .
- ^ Бехар Д.М., Юнусбаев Б., Мецпалу М., Мецпалу Е., Россет С., Парик Дж. и др. (июль 2010 г.). «Полногеномная структура еврейского народа». Природа . 466 (7303): 238–42. Бибкод : 2010Natur.466..238B . дои : 10.1038/nature09103 . ПМИД 20531471 . S2CID 4307824 .
- ^ Сенгупта С., Животовский Л.А., Кинг Р., Мехди С.К., Эдмондс К.А., Чоу К.Э. и др. (февраль 2006 г.). «Полярность и временность распределения Y-хромосомы в Индии с высоким разрешением определяют как местное, так и экзогенное распространение и выявляют незначительное генетическое влияние скотоводов Центральной Азии» . Американский журнал генетики человека . 78 (2): 202–21. дои : 10.1086/499411 . ПМК 1380230 . ПМИД 16400607 .
- ^ Перейти обратно: а б Ди Кристофаро Дж., Пеннарун Э., Мазьер С., Майрес Н.М., Лин А.А., Темори С.А. и др. (2013). «Афганский Гиндукуш: место, где сходятся потоки генов Евразийского субконтинента» . ПЛОС ОДИН . 8 (10): е76748. Бибкод : 2013PLoSO...876748D . дои : 10.1371/journal.pone.0076748 . ПМЦ 3799995 . ПМИД 24204668 .
- ^ Липпольд С., Сюй Х., Ко А., Ли М., Рено Г., Баттоф А. и др. (2014). «Демографические истории отца и матери человека: данные по Y-хромосоме высокого разрешения и последовательностям мтДНК» . Исследовательская генетика . 5:13 . дои : 10.1186/2041-2223-5-13 . ПМК 4174254 . ПМИД 25254093 .
- ^ Карафет Т.М., Осипова Л.П., Савина О.В., Холлмарк Б, Хаммер М.Ф. (ноябрь 2018). «Сибирское генетическое разнообразие раскрывает сложное происхождение самодийскоязычного населения». Американский журнал биологии человека . 30 (6): e23194. дои : 10.1002/ajhb.23194 . ПМИД 30408262 . S2CID 53238849 .
- ^ Ashirbekov EE, Botbaev DM, Belkozhaev AM, Abayldaev AO, Neupokoeva AS, Mukhataev JE, et al. (2017). "Распределение гаплогрупп И-хромосомы казахов Южно-Казахстанской, Жамбылской и Алматинской областей" [Distribution of Y-Chromosome Haplogroups of the Kazakh from the South Kazakhstan, Zhambyl, and Almaty Regions] (PDF) . Reports of the National Academy of Sciences of the Republic of Kazakhstan (in Russian). 6 (316): 85–95. Archived from the original (PDF) on 2021-09-23 . Retrieved 2021-11-20 .
- ^ Шуху Л.И., Илихаму Н.И., Баке Р.А., Бупатима А.Б., Матюсуп Д.О. (2018). «Исследование генетического разнообразия трех изолированных популяций в Синьцзяне с использованием Y-SNP». Acta Anthropologica Sinica . 37 (1): 146–56. doi : 10.16359/j.cnki.cn11-1963/q.2017.0067 .
- ^ R1b1a1b (R-M269) ранее был R1b1a1a2. С 2003 по 2005 год то, что сейчас называется R1b1a1b, называлось R1b3. С 2005 по 2008 год это был R1b1c. С 2008 по 2011 год это был R1b1b2. С 2011 по 2018 год это был R1b1a1a2.
- ^ Перейти обратно: а б Балареск П., Боуден Г.Р., Адамс С.М., Люнг Х.И., Кинг Т.Е., Россер З.Х. и др. (январь 2010 г.). Пенни Д. (ред.). «Преимущественно неолитическое происхождение европейских отцовских линий» . ПЛОС Биология . 8 (1): e1000285. дои : 10.1371/journal.pbio.1000285 . ПМЦ 2799514 . ПМИД 20087410 .
- ^ Арреди Б., Полони Э.С., Тайлер-Смит С. (2007). «Заселение Европы». В Кроуфорде, М.Х. (ред.). Антропологическая генетика: теория, методы и приложения . Кембридж, Великобритания: Издательство Кембриджского университета. п. 394. ИСБН 978-0-521-54697-3 .
- ^ Кручиани Ф., Тромбетта Б., Антонелли С., Пасконе Р., Валезини Г., Скальци В. и др. (июнь 2011 г.). «Сильная внутри- и межконтинентальная дифференциация, выявленная SNP Y-хромосомы M269, U106 и U152». Международная судебно-медицинская экспертиза. Генетика . 5 (3): е49-52. дои : 10.1016/j.fsigen.2010.07.006 . hdl : 11573/226727 . ПМИД 20732840 .
- ^ Грантс, Виола; Равеан, Александр; Маттиоли, Франческа; Батталья, Винченца; Сала, Чинция; Тониоло, Даниэла; Ферретти, Лука; Гарделла, Рита; Ахилли, Алессандро; Оливьери, Анна; Торрони, Антонио; Пассарино, Джузеппе; Семино, Орнелла (февраль 2018 г.). «Реконструкция генетической истории итальянцев: новые идеи с точки зрения мужчины (Y-хромосомы)» . Анналы биологии человека . 45 (1): 44–56. дои : 10.1080/03014460.2017.1409801 . ISSN 1464-5033 . ПМИД 29382284 . S2CID 43501209 .
- ^ Перейти обратно: а б Андерхилл П.А., Шен П., Линь А.А., Джин Л., Пассарино Г., Ян В.Х. и др. (ноябрь 2000 г.). «Вариации последовательности Y-хромосомы и история человеческих популяций». Природная генетика . 26 (3): 358–361. дои : 10.1038/81685 . ПМИД 11062480 . S2CID 12893406 .
- ^ Лобов А.С. (2009). Структура генофонда башкирских субпопуляций (PDF) (кандидатская диссертация) (на русском языке). Институт биохимии и генетики Уфимского научного центра РАН. Архивировано из оригинала (PDF) 16 августа 2011 г.
- ^ Ордоньес А.С., Фрегель Р., Трухильо-Медерос А., Эрвелла М., де-ла-Руа К., Арне-де-ла-Роса М. (2017). «Генетические исследования доиспанского населения, захороненного в пещере Пунта-Асуль (Эль-Йерро, Канарские острова)». Журнал археологической науки . 78 : 20–28. Бибкод : 2017JArSc..78...20O . дои : 10.1016/j.jas.2016.11.004 .
- ^ Робино С., Кробу Ф., Ди Гаэтано С., Бекада А., Бенхамамуш С., Черутти Н. и др. (май 2008 г.). «Анализ гаплогрупп SNP Y-хромосомы и гаплотипов STR в выборке населения Алжира». Международный журнал юридической медицины . 122 (3): 251–5. дои : 10.1007/s00414-007-0203-5 . ПМИД 17909833 . S2CID 11556974 .
- ^ Перейти обратно: а б с Вуд Э.Т., Стовер Д.А., Эрет С., Дестро-Бисол Г., Спедини Г., МакЛеод Х. и др. (июль 2005 г.). «Контрастные закономерности вариаций Y-хромосомы и мтДНК в Африке: свидетельства демографических процессов с учетом пола» . Европейский журнал генетики человека . 13 (7): 867–76. дои : 10.1038/sj.ejhg.5201408 . ПМИД 15856073 . S2CID 20279122 .
- ^ Епископосян Л, Худоян А, Арутюнян А (2006). «Генетическое тестирование гипотезы замены языка в Юго-Западной Азии». Иран и Кавказ . 10 (2): 191–208. дои : 10.1163/157338406780345899 . JSTOR 4030922 . S2CID 162345193 .
- ^ Масьямо Хэй. «Гаплогруппа R1b (Y-ДНК)» . Евпедия .
- ^ Vadimovna TN (February 2015). Изменчивость Митохондриальной ДНК и Y-Хромосомы в Популяциях Волго-Уральского Региона [ Mitochondrial DNA variation and the Y-chromosome in the population of the Volga-Ural Region ] (PDF) (Biological Sciences thesis) (in Russian). Archived from the original (PDF) on 2017-04-02.
- ^ Эррера К.Дж., Лоури Р.К., Хэдден Л., Кальдерон С., Чиу С., Епископосян Л. и др. (март 2012 г.). «Неолитические отцовские сигналы указывают на то, что Армянское нагорье было заселено земледельцами» . Европейский журнал генетики человека . 20 (3): 313–20. дои : 10.1038/ejhg.2011.192 . ПМК 3286660 . ПМИД 22085901 .
- ^ Ванек Д., Саскова Л., Кох Х. (июнь 2009 г.). «Анализ родства и Y-хромосомы человеческих останков VII века: новая процедура извлечения ДНК и типирования древнего материала» . Хорватский медицинский журнал . 3. 50 (3): 286–95. дои : 10.3325/cmj.2009.50.286 . ПМК 2702742 . ПМИД 19480023 .
- ^ Д'Атанасио Э., Тромбетта Б., Бонито М., Финоккио А., Ди Вито Г., Сегиззи М. и др. (февраль 2018 г.). «Заселение последней Зеленой Сахары, выявленное путем широкомасштабного повторения последовательности транссахарских отцов» . Геномная биология . 19 (1): 20. дои : 10.1186/s13059-018-1393-5 . ПМК 5809971 . ПМИД 29433568 .
- ^ Маркус Дж. Х., Пост С., Рингбауэр Х., Лай Л., Скитс Р., Сидоре С. и др. (февраль 2020 г.). «Генетическая история от среднего неолита до представления на средиземноморском острове Сардиния» . Природные коммуникации . 11 (1): 939. Бибкод : 2020NatCo..11..939M . дои : 10.1038/s41467-020-14523-6 . ПМК 7039977 . ПМИД 32094358 .
- ^ Перейти обратно: а б Грюни, Виола; и др. (2019). «Анализ Y-хромосомы и фамилий для реконструкции прошлых популяционных структур: население Сардинии как тестовый пример» . Международный журнал молекулярных наук . 20 (5763): 5763. doi : 10.3390/ijms20225763 . ПМК 6888588 . ПМИД 31744094 .
- ^ Аллентофт, Мэн (2024). «Популяционная геномика послеледниковой Западной Евразии» . Природа . 625 (7994): Дополнительная информация, стр. 48. Бибкод : 2024Natur.625..301A . дои : 10.1038/s41586-023-06865-0 . ПМЦ 10781627 . ПМИД 38200295 .
- ^ Хабер, Марк; Меззавилла, Массимо; Бергстрем, Андерс; Прадо-Мартинес, Хавьер; Халласт, Пилле; Саиф-Али, Эр-Рияд; Аль-Хабори, Молхам; Дедусси, Джордж; Зеггини, Элефтерия; Блю-Смит, Джейсон; Уэллс, Р. Спенсер; Сюэ, Яли; Заллуа, Пьер А.; Тайлер-Смит, Крис (декабрь 2016 г.). «Генетическое разнообразие Чада раскрывает африканскую историю, отмеченную многочисленными голоценовыми евразийскими миграциями» (PDF) . Американский журнал генетики человека . 99 (6): 1316–1324. дои : 10.1016/j.ajhg.2016.10.012 . ПМК 5142112 . ПМИД 27889059 . S2CID 38169172 .
- ^ Куличова, Ива; и др. (2017). «Внутренняя диверсификация гаплогрупп, не принадлежащих к югу от Сахары, в сахельском населении и распространение скотоводства за пределы Сахары» . Американский журнал биологической антропологии . 164 (2): 424–434. дои : 10.1002/ajpa.23285 . ПМИД 28736914 .
- ^ Висенте, Марио; и др. (2019). «Популяционная история и генетическая адаптация кочевников фулани: выводы на основе полногеномных данных и признака персистенции лактазы» . БМК Геномика . 20 (1): 915. doi : 10.1186/s12864-019-6296-7 . ПМК 6888939 . ПМИД 31791255 .
- ^ Симойнс, Л.Г.; и др. (2023). «Неолит Северо-Западной Африки, инициированный мигрантами из Иберии и Леванта» . Природа . 618 (7965): 550–556. Бибкод : 2023Natur.618..550S . дои : 10.1038/s41586-023-06166-6 . ПМК 10266975 . ПМИД 37286608 .
- ^ Фрегель, Роза; Мендес, Фернандо Л.; Бокбот, Юсеф; Мартин-Сокас, Димас; Камалич-Массие, Мария Д.; Сантана, Джонатан; Моралес, Джейкоб; Авила-Аркос, Мария К.; Андерхилл, Питер А.; Шапиро, Бет; Войчик, Женевьева; Расмуссен, Мортен; Соарес, Андре ЭР; Капп, Джошуа; Сокелл, Александра (26 июня 2018 г.). «Древние геномы из Северной Африки свидетельствуют о доисторических миграциях в Магриб как из Леванта, так и из Европы» . Труды Национальной академии наук . 115 (26): 6774–6779. Бибкод : 2018PNAS..115.6774F . дои : 10.1073/pnas.1800851115 . ISSN 0027-8424 . ПМК 6042094 . ПМИД 29895688 .
- ^ Флорес С., Мака-Мейер Н., Ларруга Дж.М., Кабрера В.М., Карадше Н., Гонсалес А.М. (2005). «Изоляты в коридоре миграций: анализ вариаций Y-хромосомы с высоким разрешением в Иордании» . Журнал генетики человека . 50 (9): 435–441. дои : 10.1007/s10038-005-0274-4 . ПМИД 16142507 . S2CID 6490283 .
- ^ Хасан Х.И., Андерхилл П.А., Кавалли-Сфорца Л.Л., Ибрагим М.Е. (ноябрь 2008 г.). «Вариации Y-хромосомы у суданцев: ограниченный поток генов, соответствие языку, географии и истории». Американский журнал физической антропологии . 137 (3): 316–323. дои : 10.1002/ajpa.20876 . ПМИД 18618658 .
13/32
- ^ Шрайнер, Дэниел; Ротими, Чарльз Н. (декабрь 2018 г.). «Генетическая история Чада» . Американский журнал физической антропологии . 167 (4): 804–812. дои : 10.1002/ajpa.23711 . ISSN 0002-9483 . ПМК 6240361 . ПМИД 30259956 .
- ^ Кручиани Ф., Сантоламазца П., Шен П., Маколей В., Морал П., Олкерс А. и др. (май 2002 г.). «Обратная миграция из Азии в страны Африки к югу от Сахары подтверждается анализом гаплотипов Y-хромосомы человека с высоким разрешением» . Американский журнал генетики человека . 70 (5): 1197–214. дои : 10.1086/340257 . ПМЦ 447595 . ПМИД 11910562 . , стр. 13–14.
- ^ Конту Д., Морелли Л., Сантони Ф., Фостер Дж.В., Франкалаччи П., Кукка Ф. (январь 2008 г.). «Доказательства донеолитического происхождения генетически однородного, но разнообразного населения Сардинии, основанные на Y-хромосоме: выводы для сканирования ассоциаций» . ПЛОС ОДИН . 3 (1): e1430. Бибкод : 2008PLoSO...3.1430C . дои : 10.1371/journal.pone.0001430 . ПМК 2174525 . ПМИД 18183308 .
- ^ Заллуа П.А., Сюэ Ю., Халифе Дж., Махул Н., Дебиан Л., Платт Д.Э. и др. (апрель 2008 г.). «Разнообразие Y-хромосом в Ливане структурировано недавними историческими событиями» . Американский журнал генетики человека . 82 (4): 873–882. дои : 10.1016/j.ajhg.2008.01.020 . ПМЦ 2427286 . ПМИД 18374297 .
- ^ Чжун Х., Ши Х., Ци С.Б., Дуань З.Ю., Тан П.П., Цзинь Л. и др. (январь 2011 г.). «Расширенное исследование Y-хромосомы позволяет предположить, что современные люди мигрировали в Восточную Азию северным путем после ледникового периода» . Молекулярная биология и эволюция . 28 (1): 717–27. дои : 10.1093/molbev/msq247 . ПМИД 20837606 .
- ^ Роуолд Д.Д., Перес Бенедико Д., Гарсия-Бертран Р., Ченнакришнайя С., Альфонсо-Санчес М.А., Гайден Т., Эррера Р.Дж. (март 2016 г.). «Ладакх, Индия: земля высоких перевалов и генетической гетерогенности демонстрирует слияние миграций» . Европейский журнал генетики человека . 24 (3): 442–9. дои : 10.1038/ejhg.2015.80 . ПМЦ 4755386 . ПМИД 25966630 .
- ^ "О Р1а и Р1б из уральской былины. Артем Лукичев (с)" . 5 августа 2014 г. Архивировано из оригинала 15 декабря 2021 г. – на сайте www.youtube.com.
- ^ Coble MD, Loreille OM, Wadhams MJ, Edson SM, Maynard K, Meyer CE и др. (2009). «Тайна раскрыта: идентификация двух пропавших детей Романовых с помощью анализа ДНК» . ПЛОС ОДИН . 4 (3): e4838. Бибкод : 2009PLoSO...4.4838C . дои : 10.1371/journal.pone.0004838 . ПМЦ 2652717 . ПМИД 19277206 .
- ^ Лармюсо, Маартен HD; Делорм, Филипп; Жермен, Патрик; Вандерхейден, Нэнси (9 октября 2013 г.). «Генетическая генеалогия обнаруживает истинную гаплогруппу Y Дома Бурбонов, что противоречит недавней идентификации предполагаемых останков двух французских королей» . Eur J Hum Genet . 22 (5): 681–687. дои : 10.1038/ejhg.2013.211 . ПМЦ 3992573 . ПМИД 24105374 .
- ^ Маан А.А., Илс Дж., Акбаров А., Роуленд Дж., Сюй X, Джоблинг М.А. и др. (ноябрь 2017 г.). «Y-хромосома: основа мужского здоровья?» . Европейский журнал генетики человека . 25 (11): 1181–1188. дои : 10.1038/ejhg.2017.128 . ПМЦ 5643963 . ПМИД 28853720 .
- ^ Тиммерс П., Уилсон Дж. Ф. (июль 2022 г.). «Ограниченное влияние вариаций Y-хромосомы на ишемическую болезнь сердца и смертность в биобанке Великобритании — краткий отчет» . Атеросклероз, тромбоз и сосудистая биология . 42 (9): 1198–1206. дои : 10.1161/ATVBAHA.122.317664 . ПМЦ 9394501 . ПМИД 35861954 .
Библиография
[ редактировать ]- Джонс Э.Р., Зарина Г., Моисеев В., Лайтфут Э., Нигст П.Р., Маника А. и др. (февраль 2017 г.). «Переход к неолиту в Прибалтике не был вызван смешением с ранними европейскими земледельцами» . Современная биология . 27 (4): 576–582. Бибкод : 2017CBio...27..576J . дои : 10.1016/j.cub.2016.12.060 . ПМК 5321670 . ПМИД 28162894 .
- Гонсалес-Фортес Дж., Джонс Э.Р., Лайтфут Е., Бонсалл С., Лазар С., Грандаль-д'Англад А. и др. (июнь 2017 г.). «Палеогеномные доказательства смешения нескольких поколений неолитических земледельцев и мезолитических охотников-собирателей в бассейне Нижнего Дуная» . Современная биология . 27 (12): 1801–1810.e10. Бибкод : 2017CBio...27E1801G . дои : 10.1016/j.cub.2017.05.023 . ПМЦ 5483232 . ПМИД 28552360 .
- Хаак В., Лазаридис И., Паттерсон Н., Роланд Н., Маллик С., Ламас Б. и др. (июнь 2015 г.). «Массовая миграция из степи явилась источником индоевропейских языков в Европе» . Природа . 522 (7555): 207–211. arXiv : 1502.02783 . Бибкод : 2015Natur.522..207H . дои : 10.1038/nature14317 . ПМК 5048219 . ПМИД 25731166 .
- Холлард С., Звенигорский В., Ковалев А., Кирюшин Ю., Тишкин А., Лазаретов И. и др. (сентябрь 2018 г.). «Новые генетические свидетельства сходства и различий между сибирскими популяциями бронзового века». Американский журнал физической антропологии . 167 (1): 97–107. дои : 10.1002/ajpa.23607 . ПМИД 29900529 . S2CID 205337212 .
- Липсон М., Сеченьи-Надь А., Маллик С., Поса А., Стегмар Б., Кирл В. и др. (ноябрь 2017 г.). «Параллельные палеогеномные разрезы раскрывают сложную генетическую историю ранних европейских земледельцев» . Природа . 551 (7680): 368–372. Бибкод : 2017Natur.551..368L . дои : 10.1038/nature24476 . ПМК 5973800 . ПМИД 29144465 .
- Маркус Дж. Х., Пост С., Рингбауэр Х., Лай Л., Скитс Р., Сидоре С. и др. (февраль 2020 г.). «Генетическая история от среднего неолита до представления на средиземноморском острове Сардиния» . Природные коммуникации . 11 (1): 939. Бибкод : 2020NatCo..11..939M . дои : 10.1038/s41467-020-14523-6 . ПМК 7039977 . ПМИД 32094358 .
- Мэтисон И., Алпаслан-Руденберг С., Пост С., Сечени-Надь А., Роланд Н., Маллик С. и др. (март 2018 г.). «Геномная история Юго-Восточной Европы» . Природа . 555 (7695): 197–203. Бибкод : 2018Natur.555..197M . дои : 10.1038/nature25778 . ПМК 6091220 . ПМИД 29466330 .
- Нарасимхан В.М., Паттерсон Н., Мурджани П., Роланд Н., Бернардос Р., Маллик С. и др. (сентябрь 2019 г.). «Формирование человеческих популяций в Южной и Центральной Азии » Наука 365 (6457): eaat7487. дои : 10.1126/science.aat7487 . ПМЦ 6822619 . ПМИД 31488661 .
- Олальде И., Брейс С., Аллентофт М.Э., Армит И., Кристиансен К., Бут Т. и др. (март 2018 г.). «Феномен стакана и геномная трансформация северо-западной Европы» . Природа . 555 (7695): 190–196. Бибкод : 2018Natur.555..190O . дои : 10.1038/nature25738 . ПМЦ 5973796 . ПМИД 29466337 .
- Хаак, Вольфганг; Лазаридис, Иосиф; Паттерсон, Ник; Роланд, Надин; Маллик, Свапан; Ламас, Бастьен; Брандт, Гвидо; Норденфельт, Сюзанна; и др. (2015). «Массовая миграция из степи является источником индоевропейских языков в Европе» . Природа . 522 (7555): 207–211. arXiv : 1502.02783 . Бибкод : 2015Natur.522..207H . bioRxiv 10.1101/013433 . дои : 10.1038/NATURE14317 . ПМК 5048219 . ПМИД 25731166 .
- Санчес-Куинто Ф., Мальмстрем Х., Фрейзер М., Гирдланд-Флинк Л., Свенссон Э.М., Симойнс Л.Г. и др. (май 2019 г.). «Мегалитические гробницы в западной и северной неолитической Европе были связаны с родственным обществом» . Труды Национальной академии наук Соединенных Штатов Америки . 116 (19): 9469–9474. Бибкод : 2019PNAS..116.9469S . дои : 10.1073/pnas.1818037116 . ПМК 6511028 . ПМИД 30988179 .
- Фу К., Пост С., Хайдиньяк М., Петр М., Маллик С., Фернандес Д. и др. (июнь 2016 г.). «Генетическая история Европы ледникового периода» . Природа . 534 (7606): 200–5. Бибкод : 2016Natur.534..200F . дои : 10.1038/nature17993 . ПМЦ 4943878 . ПМИД 27135931 .
{{|url= https://www.dotyk.cz/magazin/premyslovci-dna-30000216.html}}