Гаплогруппа A-L1085
| Гаплогруппа A-L1085 | |
|---|---|
| Возможное время происхождения | 140 000 юаней, [ 1 ] 125 000–382 000 юаньских фунтов [ 2 ] |
| Возможное место происхождения | Центральная - Северо-Западная Африка [ 1 ] |
| Предок | Гомо Y-MRCA |
| Потомки | А-В148 (А0), А-П305 (А1) |
| Самые высокие частоты | Намибия (Цумкве Сан, Нама ) 60-70% Южный Судан ( динка , шиллук , нуэр ) 33%-61,5% Эфиопия ( Бета-Израиль ) 41%-46% |
Гаплогруппа A-L1085 , также известная как гаплогруппа A0-T, человека представляет собой гаплогруппу Y-ДНК . Это часть отцовской линии почти всех ныне живущих людей. SNP отошли L1085 сыграл две роли в популяционной генетике : во-первых, большинство гаплогрупп Y-ДНК от него и; во-вторых, он определяет недивергентную базальную кладу A-L1085*.
A0-T имеет две основные ветви: A-V148 (также известную как гаплогруппа A0) и гаплогруппа A-P305 (гаплогруппа A1).
Источник
[ редактировать ]
Многие предположения о происхождении гаплогруппы A-L1085 предполагают, что она была связана с предковым населением охотников-собирателей Южной Африки. Это связано с тем, что линии гаплогруппы A-L1085 часто встречаются среди народа сан .
Однако линии A-L1085 Южной Африки являются субкладами линий A, обнаруженными в других частях Африки, в основном среди нилотских народов , но также и среди других африканцев. Это говорит о том, что линии A-L1085 прибыли в Южную Африку из других мест. [ 3 ] Две наиболее базальные линии гаплогруппы A-L1085, A-V148 и A-P305, были обнаружены в Западной Африке, Северо-Западной Африке и Центральной Африке. Кручиани и др. 2011 год предполагает, что эти линии могли возникнуть где-то между Центральной и Северо-Западной Африкой, хотя такая интерпретация все еще является предварительной из-за неполного географического охвата африканских Y-хромосом. [ 1 ]
Первоначальные исследования показали, что линии гаплогруппы A-L1085 возникли около 60 000 лет назад, что было значительно позже, чем TMRCA для линий митохондриальной ДНК, которые сливаются между 150-200 тысячами лет назад. Кручиани и др. 2011 год, в результате серьезной реструктуризации ветвей, отодвинул корень дерева Y-хромосомы на 142 000 лет назад. [ 1 ]
В ноябре 2012 г. новое исследование Scozzari et al. подтвердили «гипотезу о происхождении гаплогруппы A1b в северо-западном квадранте африканского континента и вместе с недавними находками древних линий Y-хромосомы в центрально-западной Африке предоставили новые доказательства относительно географического происхождения человеческого MSY. разнообразие". [ 4 ]
Географическое распространение
[ редактировать ]Центральная Африка
[ редактировать ]Гаплогруппа A-M13 наблюдалась в популяциях северного Камеруна (2/9 = 22% Тупури , [ 5 ] 4/28 = 14% Мандары , [ 5 ] 2/17 = 12% фульбе [ 6 ] ) и восточной ДРК (2/9 = 22% Алура , [ 5 ] 1/18 = 6% Юг , [ 5 ] 1/47 = 2% Коза [ 5 ] ).
Гаплогруппа A-M91 (xA-M31, A-M6, A-M32) наблюдалась у народа бакола на юге Камеруна (3/33 = 9%). [ 5 ]
Без тестирования на какой-либо субклад гаплогруппа A-L1085 наблюдалась в выборках нескольких популяций Габона , в том числе 9% (3/33) выборки Бака , 3% (1/36) выборки Ндуму , 2% (1/46) выборки Думы , 2% (1/57) выборки Нзеби и 2% (1/60) выборки Цого . [ 7 ]
Восточная Африка
[ редактировать ]Гаплогруппа А-М13 распространена среди жителей Южного Судана (53%), [ 8 ] особенно динка (61,5%). [ 9 ] Гаплогруппа A-M13 также наблюдалась в другой выборке населения Южного Судана с частотой 45% (18/40), включая 1/40 A-M171. [ 10 ] Гаплогруппа А также была обнаружена в 14,6% (7/48) выборки из Амхары . [ 11 ] 10,3% (8/78) образца оромо , [ 11 ] 13,6% (12/88) другой выборки из Эфиопии, [ 10 ] и 41% выборки Бета-Израиля (Cruciani et al., 2002), а важные проценты также разделяют банту в Кении (14%, Луис и др., 2004) и иракцы в Танзании (3/43 = 7,0% ( Луис и др., 2004) до 1/6 = 17% (Найт и др., 2003)).
Северная Африка
[ редактировать ]Субклад A1 наблюдался у ливийских берберов , а субклад A-M13 наблюдался примерно у 3% египетских мужчин.
Южная Африка
[ редактировать ]Одно исследование обнаружило гаплогруппу А в выборках различных койсаноязычных групп с частотой от 10% до 70%. [ 5 ] Эта конкретная гаплогруппа не была обнаружена в выборке хадзабе из Танзании, популяции, которая иногда группировалась с другими койсанскими группами из-за присутствия щелкающих согласных в их языке, но чей язык был тщательно продемонстрирован как изолят, столь же не связанный с ними, как и другие группы. языки работы лингвиста Бонни Сэндс.
Европа
[ редактировать ]Гаплогруппа А наблюдалась как А1 у европейских мужчин в Англии. AY-хромосома наблюдалась также с низкой частотой в Малой Азии, на Ближнем Востоке и на некоторых островах Средиземноморья, у греков Эгейского моря, сицилийцев (0,2% А1а в Капо д'Орландо и 0,5% А1b на всем острове ), палестинцев. , иорданцы и йеменцы. Без тестирования на какой-либо субклад гаплогруппа A1b была обнаружена у выборки греков из Митилини на острове Лесбос в Эгейском море. [ 12 ] и A1b наблюдался также у 0,1% иберийских евреев . Авторы одного исследования сообщили об обнаружении гаплогруппы А у 3,1% (2/65) выборки киприотов . [ 13 ] хотя они окончательно не исключили возможность того, что любой из этих людей может принадлежать к гаплогруппе B.
Распределение субкладов
[ редактировать ]
А-В148* (А0)*)
[ редактировать ]A-V148 — одна из двух основных ветвей A0-T. [ 1 ]
Этот раздел нуждается в расширении . Вы можете помочь, добавив к нему . ( апрель 2018 г. ) |
А-П305* (А1*)
[ редактировать ]Гаплогруппа A-P305* в основном распространена в некоторых частях Африки , хотя несколько случаев были зарегистрированы в Европе и Западной Азии .
A-P305 встречается чаще всего у пигмеев Бакола (Южный Камерун ) - 8,3% и у берберов из Алжира - 1,5%. [ 1 ] и в Гане . [ 4 ] Клада также достигает высоких частот среди бушменских популяций охотников-собирателей в Южной Африке , за которыми следуют многие нилотские группы в Восточной Африке . Однако самые старые субклады гаплогруппы А встречаются исключительно в Центральной и Северо- Западной Африке , где она и, следовательно, Y-хромосомный Адам, как полагают, возникли около 140 000 лет назад. [ 1 ] Клада также с заметной частотой наблюдалась у некоторых популяций Эфиопии , а также у некоторых групп пигмеев в Центральной Африке.
Гаплогруппа A-L1085 менее распространена среди носителей Нигера и Конго , которые в основном принадлежат к кладе E1b1a . гаплогруппа Е возникла в Северо-Восточной Африке. Считается, что [ 14 ] и позже был завезен в Западную Африку, откуда около 5000 лет назад распространился в Центральную, Южную и Юго-Восточную Африку с экспансией банту . [ 15 ] [ 7 ] По данным Вуда и др. (2005) и Роза и др. (2007), такие относительно недавние перемещения населения из Западной Африки изменили ранее существовавшее хромосомное разнообразие Y популяции в Центральной, Южной и Юго-Восточной Африке, заменив предыдущие гаплогруппы в этих областях ныне доминирующими линиями E1b1a. Однако сегодня в этих регионах можно наблюдать следы предковых жителей по наличию гаплогрупп Y-ДНК A-M91 и B-M60, которые распространены в некоторых реликтовых популяциях, таких как пигмеи мбути и койсаны . [ 16 ] [ 5 ] [ 17 ]
| Африка | ||
| Исследуемая популяция | Частота. (в %) | |
| [ 5 ] | Цумкве Сан (Намибия) | 66% |
| [ 5 ] | Нама (Намибия) | 64 |
| [ 8 ] | Динка (Судан) | 62 |
| [ 8 ] | Шиллук (Судан) | 53 |
| [ 8 ] | Нуба (Судан) | 46 |
| [ 10 ] | Хойсан | 44 |
| [ 6 ] [ 18 ] | Эфиопские евреи | 41 |
| [ 5 ] [ 6 ] | !Если /Секеле | ~40 |
| [ 8 ] | Боргу (Судан) | 35 |
| [ 8 ] | Нуэр (Судан) | 33 |
| [ 8 ] | Мех (Судан) | 31 |
| [ 5 ] | Масаи (Кения) | 27 |
| [ 19 ] | Нара (Эритрея) | 20 |
| [ 8 ] | Масалит (Судан) | 19 |
| [ 5 ] [ 11 ] | Амхара (Эфиопия) | ~16 |
| [ 10 ] | эфиопы | 14 |
| [ 20 ] | Банту (Кения) | 14 |
| [ 5 ] | Мандара (Камерун) | 14 |
| [ 8 ] | Хауса (Судан) | 13 |
| [ 6 ] | Хве (Южная Африка) | 12 |
| [ 6 ] | Фульбе (Камерун) | 12 |
| [ 5 ] | Дама (Намибия) | 11 |
| [ 11 ] | Оромо (Эфиопия) | 10 |
| [ 19 ] | Кунама (Эритрея) | 10 |
| [ 5 ] | Южносемитский (Эфиопия) | 10 |
| [ 20 ] | Арабы (Египет) | 3 |
В составной выборке из 3551 африканского мужчины частота гаплогруппы А составила 5,4%. [ 21 ] Самая высокая частота гаплогруппы А зарегистрирована среди койсанцев Южной Африки, бета-израильтян и нило-сахарцев .
А-М31
[ редактировать ]Субклад A-M31 был обнаружен примерно в 2,8% (8/282) пула из семи выборок различных этнических групп в Гвинее-Бисау , особенно среди папель-манжако-манканья (5/64 = 7,8%). [ 16 ] Более раннее исследование Gonçalves et al. В 2003 году сообщалось об обнаружении A-M31 в 5,1% (14/276) образцов из Гвинеи-Бисау и в 0,5% (1/201) пары образцов из Кабо-Верде . [ 22 ] Авторы другого исследования сообщили об обнаружении гаплогруппы A-M31 у 5% (2/39) выборки мандинка из Сенегамбии и 2% (1/55) выборки догонов из Мали . [ 5 ] Гаплогруппа A-M31 также была обнаружена у 3% (2/64) выборки берберов из Марокко. [ 6 ] и 2,3% (1/44) выборки неустановленной этнической принадлежности из Мали . [ 10 ]
По меньшей мере семь мужчин, родом из Йоркшира , Англия , и носящих характерную фамилию Ревис, были идентифицированы как принадлежащие к субкладу A-M31. В новостях говорилось, что эти мужчины были фенотипически «европейцами» и не подозревали о каком-либо африканском происхождении. Последующие исследования показали, что у них был общий предок по отцовской линии в 18 веке. [ 21 ]
А-М6
[ редактировать ]A-M6 (ранее A2) обычно встречается у койсанских народов. Авторы одного исследования сообщили об обнаружении гаплогруппы A-M6(xA-P28) у 28% (8/29) выборки Цумкве Сан и 16% (5/32) выборки !Кунг /Секеле, а также гаплогруппы A-P28 в 17% (5/29) образца Цумкве Сан, 9% (3/32) образца !Кунг /Секеле, 9% (1/11) образца Нама и 6% (1/18) образца Дама . [ 5 ] Авторы другого исследования сообщили об обнаружении гаплогруппы A-M6 у 15,4% (6/39) выборки койсанских мужчин, включая 5/39 A-M6(xA-M114) и 1/39 A-M114. [ 10 ]
А-М32
[ редактировать ]Клада A-M32 (ранее A3) содержит наиболее многочисленные ветви гаплогруппы A-L1085 и в основном встречается в Восточной Африке и Южной Африке .
А-М28
[ редактировать ]Субклад A-M28 (ранее A3a) редко наблюдался на Африканском Роге . В 5% (1/20) смешанной выборки носителей южносемитских языков из Эфиопии [ 5 ] 1,1% (1/88) выборки эфиопов, [ 10 ] и 0,5% (1/201) у сомалийцев. [ 23 ]
А-М51
[ редактировать ]Субклад A-M51 (ранее A3b1) встречается чаще всего среди койсанских народов (6/11 = 55% нама , [ 5 ] 11/39 = 28% койсан, [ 10 ] 7/32 = 22% !Если /Секеле, [ 5 ] 6/29 = 21% Цумкве Сан, [ 5 ] 1/18 = 6% шанс [ 5 ] ). Однако это также было обнаружено с меньшей частотой среди народов банту Южной Африки , включая 2/28 = 7% сото-тсвана , [ 5 ] 3/53 = 6% некойсанских жителей Южной Африки, [ 10 ] 4/80 = 5% коса , [ 5 ] и 1/29 = 3% зулусов . [ 5 ]
А-М13
[ редактировать ]Субклад A-M13 (ранее A3b2), обычно встречающийся в Восточной Африке и северном Камеруне, отличается от субкладов, обнаруженных в койсанских образцах, и лишь отдаленно связан с ними. Это открытие предполагает древнее расхождение.
В Судане гаплогруппа A-M13 обнаружена у 28/53 = 52,8% южных суданцев , 13/28 = 46,4% нубийцев центрального Судана, 25/90 = 27,8% западных суданцев , 4/32 = 12,5% местных жителей хауса и 5/216 = 2,3% жителей Севера. Суданский. [ 24 ]
В Эфиопии одно исследование сообщило об обнаружении гаплогруппы A-M13 у 14,6% (7/48) выборки Амхары и 10,3% (8/78) выборки Оромо . [ 11 ] Другое исследование показало, что гаплогруппа A-M118 обнаружена у 6,8% (6/88) и гаплогруппа A-M13(xA-M171, A-M118) у 5,7% (5/88) смешанной выборки эфиопов, что составляет общее количество 12,5% (11/88) А-М13. [ 10 ]
Гаплогруппа A-M13 также иногда наблюдалась за пределами Центральной и Восточной Африки, например, в Эгейском регионе Турции . (2/30 = 6,7%) [ 25 ] ), йеменские евреи (1/20 = 5% [ 18 ] ), Египет (4/147 = 2,7%, [ 20 ] 3/92 = 3.3% [ 5 ] ), палестинские арабы (2/143 = 1,4% [ 26 ] ), Сардиния (1/77 = 1,3%, [ 27 ] 1/22 = 4.5% [ 10 ] ), столица Иордании Амман % (1/101=1 [ 28 ] ) и Оман (1/121 = 0,8% [ 20 ] ).
Филогенетика
[ редактировать ]Филогенетическая история
[ редактировать ]До 2002 года в научной литературе существовало как минимум семь систем наименования филогенетического дерева Y-хромосомы. Это привело к значительной путанице. В 2002 году основные исследовательские группы объединились и сформировали Консорциум Y-хромосомы (YCC). Они опубликовали совместный документ, в котором было создано единое новое дерево, которое все согласились использовать. Позже группа гражданских ученых, интересующихся популяционной генетикой и генетической генеалогией, сформировала рабочую группу для создания любительского дерева, стремясь быть прежде всего своевременным. В таблице ниже собраны все эти работы, относящиеся к знаковому дереву YCC 2002 года. Это позволяет исследователю, просматривающему старую опубликованную литературу, быстро перемещаться между номенклатурами.
| YCC 2002/2008 (сокращенно) | (а) | (б) | (с) | (г) | (е) | (г) | (или) | YCC 2002 (от руки) | YCC 2005 (от руки) | YCC 2008 (от руки) | YCC 2010r (от руки) | ИСОГГ 2006 г. | ИСОГГ 2007 | ИСОГГ 2008 | ИСОГГ 2009 | ИСОГГ 2010 | ИЗОГГ 2011 | ИЗОГГ 2012 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| А-М31 | 7 | я | 1А | 1 | – | H1 | А | А1 | А1 | А1 | А1а | А1 | А1 | А1а | А1а | А1а | А1а | А1а |
| А-М6 | 27 | я | 2 | 3 | – | H1 | А | А2* | А2 | А2 | А2 | А2 | А2 | А2 | А2 | А2 | А2 | А1б1а1а |
| А-М114 | 27 | я | 2 | 3 | – | H1 | А | А2а | А2а | А2а | А2а | А2а | А2а | А2а | А2а | А2а | А2а | А1б1а1а1а |
| А-П28 | 27 | я | 2 | 4 | – | H1 | А | А2б | А2б | А2б | А2б | А2б | А2б | А2б | А2б | А2б | А2б | А1б1а1а1б |
| А-М32 | * | * | * | * | * | * | * | * | А3 | А3 | А3 | А3 | А3 | А3 | А3 | А3 | А3 | A1b1b |
| А-М28 | 7 | я | 1А | 1 | – | H1 | А | А3а | А3а | А3а | А3а | А3а | А3а | А3а | А3а | А3а | А3а | А1b1b1 |
| А-М51 | 7 | я | 1А | 1 | – | H1 | А | А3б1 | А3б1 | А3б1 | А3б1 | А3б1 | А3б1 | А3б1 | А3б1 | А3б1 | А3б1 | A1b1b2a |
| А-М13 | 7 | я | 1А | 2 | ЕС1 | H1 | А | A3b2* | А3б2 | А3б2 | А3б2 | А3б2 | А3б2 | А3б2 | А3б2 | А3б2 | А3б2 | A1b1b2b |
| А-М171 | 7 | я | 1А | 2 | ЕС1 | H1 | А | A3b2a | A3b2a | A3b2a | A3b2a | A3b2a | A3b2a | A3b2a | A3b2a | A3b2a | A3b2a | удаленный |
| А-М118 | 7 | я | 1А | 2 | ЕС1 | H1 | А | A3b2b | A3b2b | A3b2b | A3b2b | A3b2b | A3b2b | A3b2b | A3b2b | A3b2b | A3b2b | А1b1b2b1 |
Оригинальные исследовательские публикации
[ редактировать ]Следующие исследовательские группы в соответствии с их публикациями были представлены в создании Дерева YCC.
Кручиани 2011
[ редактировать ]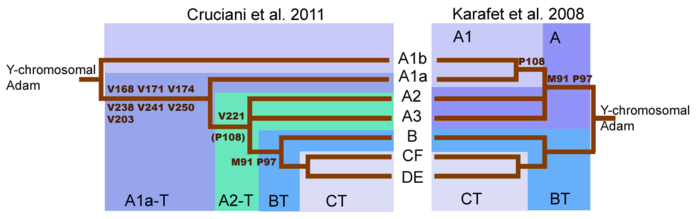
Серьезный сдвиг в понимании дерева гаплогруппы А произошел с публикацией ( Cruciani 2011 ). Первоначальное секвенирование Y-хромосомы человека показало, что первое разделение генеалогического древа Y-хромосомы произошло с мутацией M91, которая отделила гаплогруппу A от гаплогруппы BT . [ 29 ] Однако теперь известно, что самый глубокий раскол в дереве Y-хромосомы обнаружен между двумя ранее сообщенными субкладами гаплогруппы A, а не между гаплогруппой A и гаплогруппой BT. Субклады A1b и A1a-T теперь происходят непосредственно от корня дерева. Перестройка генеалогического древа Y-хромосомы подразумевает, что линии, классифицированные как гаплогруппа A, не обязательно образуют монофилетическую кладу. [ 1 ] Таким образом, гаплогруппа A относится к совокупности линий, которые не обладают маркерами, определяющими гаплогруппу BT, хотя многие линии внутри гаплогруппы A имеют лишь очень отдаленное родство.
Мутации M91 и P97 отличают гаплогруппу A от гаплогруппы BT . В хромосомах гаплогруппы А маркер M91 состоит из участка из 8 Т нуклеиновых оснований. В гаплогруппе ВТ и хромосомах шимпанзе этот маркер состоит из 9 Т нуклеиновых оснований. Эта закономерность предполагает, что участок 9T гаплогруппы BT был наследственной версией и что гаплогруппа A образовалась путем делеции одного нуклеинового основания . [ 1 ] [ 29 ]
Но по данным Cruciani et al. В 2011 году область, окружающая маркер M91, представляет собой горячую точку мутаций, склонную к повторяющимся мутациям. Поэтому возможно, что участок 8T гаплогруппы A может быть предковым состоянием M91, а участок 9T гаплогруппы BT может быть производным состоянием, возникшим в результате вставки 1T. Это могло бы объяснить, почему субклады A1b и A1a-T, самые глубокие ветви гаплогруппы A, обладают удлинением 8T. Более того, Cruciani et al. В 2011 году было установлено, что маркер P97, который также используется для идентификации гаплогруппы A, обладал наследственным состоянием в гаплогруппе A, но производным состоянием в гаплогруппе BT . [ 1 ]
Филогенетические деревья
[ редактировать ]Это филогенетическое древо субкладов гаплогрупп основано на дереве Консорциума Y-хромосомы (YCC), [ 30 ] дерево гаплогрупп ISOGG Y-ДНК, [ 15 ] и последующие опубликованные исследования.
Y-хромосомный Адам
- A0 (ранее A1b) (P305, V148, V149, V154, V164, V166, V172, V173, V177, V190, V196, V223, V225, V229, V233, V239)
- A1 (A1a-T по Cruciani 2011) (L985, L989, L990, L1002, L1003, L1004, L1009, L1013, L1053, V161, V168, V171, V174, V203, V238, V241, V250, V238, В241, В250)
- А1а (М31, Р82, В4, В14, В15, В25, В26, В28, В30, В40, В48, В53, В57, В58, В63, В76, В191, В201, В204, В214, В215, В236)
- A1b (A2-T по Cruciani 2011) (P108, V221)
- А1б1 (Л419)
- А1б1а (В50, В82, В198, В224)
- A1b1a1 ранее A2 (M14, M23, L968/M29/P3/PN3, M71, M135, M141, M206, M276/P247, M277/P248, MEH1, P4, P5, P36.1, Страница71, Страница87, Страница95)
- А1б1а1а (М6, М196)
- А1б1а1а1 (М212)
- A1b1a1a1a ранее A2a (M114)
- A1b1a1a1b ранее A2b (P28)
- A1b1a1a1c ранее A2c (P262)
- А1б1а1а1 (М212)
- А1б1а1а (М6, М196)
- A1b1a1 ранее A2 (M14, M23, L968/M29/P3/PN3, M71, M135, M141, M206, M276/P247, M277/P248, MEH1, P4, P5, P36.1, Страница71, Страница87, Страница95)
- A1b1b ранее A3 (M32)
- A1b1b1 ранее A3a (M28, M59)
- A1b1b2 ранее A3b (M144, M190, M220, P289)
- A1b1b2a ранее A3b1 (M51, P100, P291)
- A1b1b2a1 ранее A3b1a (P71, P102)
- A1b1b2b ранее A3b2 (M13, M127, M202, M219, M305):
- А1б1б2б1 (М118)
- A1b1b2a ранее A3b1 (M51, P100, P291)
- А1б1а (В50, В82, В198, В224)
- БТ (M42, M94, M139, M299, M60, M181/страница32, P85, P90, P97, страница65.1/SRY1532.1/SRY10831.1, V21, V29, V31, V59, V64, V102, V187, V202, В216, В235)
- А1б1 (Л419)
См. также
[ редактировать ]Генетика
[ редактировать ]Субклады Y-ДНК А
[ редактировать ]Основополагающее дерево Y-ДНК
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я дж Кручиани Ф, Тромбетта Б, Массайя А, Дестро-Бисол Дж, Селитто Д, Скоццари Р (июнь 2011 г.). «Пересмотренный корень филогенетического дерева Y-хромосомы человека: происхождение отцовского разнообразия в Африке» . Американский журнал генетики человека . 88 (6): 814–8. дои : 10.1016/j.ajhg.2011.05.002 . ПМК 3113241 . ПМИД 21601174 .
- ^ Мендес Ф.Л., Кран Т., Шрак Б., Кран А.М., Вирама К.Р., Вернер А.Е., Фомин Ф.Л., Брэдман Н., Томас М.Г., Карафет Т.М., Хаммер М.Ф. (март 2013 г.). «Афроамериканская линия по отцовской линии добавляет чрезвычайно древний корень к филогенетическому дереву Y-хромосомы человека» . Американский журнал генетики человека . 92 (3): 454–9. дои : 10.1016/j.ajhg.2013.02.002 . ПМК 3591855 . ПМИД 23453668 .
- ^ Батини К , Ферри Дж , Дестро-Бисол Дж , Бризигелли Ф , Луизелли Д , Санчес-Диз П , Роша Дж , Симонсон Т , Брем А , Монтано В , Эльвали Н.Э. , Спедини Дж , Д'Амато М.Э. , Майрес Н , Эббесен П. , Комас Д., Капелли С. (сентябрь 2011 г.). «Признаки доземледельческих процессов заселения в странах Африки к югу от Сахары, выявленные с помощью филогеографии ранних линий Y-хромосомы» (PDF) . Молекулярная биология и эволюция . 28 (9): 2603–13. дои : 10.1093/molbev/msr089 . ПМИД 21478374 .
- ^ Jump up to: а б Скоццари Р., Массайя А., Д'Атанасио Э., Майрес Н.М., Перего У.А., Тромбетта Б., Круциани Ф. (2012). «Молекулярное рассечение базальных клад филогенетического дерева Y-хромосомы человека» . ПЛОС ОДИН 7 (11):e4 Бибкод : 2012PLoSO...749170S . дои : 10.1371/journal.pone.0049170 . ПМЦ 3492319 . ПМИД 23145109 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа Вуд Э.Т., Стовер Д.А., Эрет С., Дестро-Бисол Дж., Спедини Дж., Маклеод Х., Луи Л., Бамшад М., Страссманн Б.И., Судьялл Х., Хаммер М.Ф. (июль 2005 г.). «Контрастные закономерности вариаций Y-хромосомы и мтДНК в Африке: свидетельства демографических процессов с учетом пола» . Европейский журнал генетики человека . 13 (7): 867–76. дои : 10.1038/sj.ejhg.5201408 . ПМИД 15856073 .
ср. Приложение A: Частоты гаплотипов Y-хромосомы
- ^ Jump up to: а б с д и ж Кручиани Ф, Сантоламазца П, Шен П, Маколей В, Морал П, Олкерс А, Модиано Д, Холмс С, Дестро-Бисоль Г, Койя В, Уоллес Д.С., Офнер П.Дж., Торрони А, Кавалли-Сфорца ЛЛ, Скоццари Р., Андерхилл ПА (май 2002 г.). «Обратная миграция из Азии в страны Африки к югу от Сахары подтверждается анализом гаплотипов Y-хромосомы человека с высоким разрешением» . Американский журнал генетики человека . 70 (5): 1197–214. дои : 10.1086/340257 . ПМЦ 447595 . ПМИД 11910562 .
- ^ Jump up to: а б Берниелл-Ли Г., Калафелл Ф., Бош Э., Хейер Э., Сика Л., Мугиама-Дауда П., ван дер Вин Л., Хомберт Дж.М., Кинтана-Мурси Л., Комас Д. (июль 2009 г.). «Генетические и демографические последствия экспансии банту: данные об отцовских линиях человека» . Молекулярная биология и эволюция . 26 (7): 1581–9. дои : 10.1093/molbev/msp069 . ПМИД 19369595 .
- ^ Jump up to: а б с д и ж г час я 28/53 (Динка, Нуэр и Шиллук), Хасан Х.И., Андерхилл П.А., Кавалли-Сфорца Л.Л., Ибрагим М.Е. (ноябрь 2008 г.). «Вариации Y-хромосомы у суданцев: ограниченный поток генов, соответствие языку, географии и истории» (PDF) . Американский журнал физической антропологии . 137 (3): 316–23. дои : 10.1002/ajpa.20876 . ПМИД 18618658 . Архивировано из оригинала (PDF) 4 марта 2009 г.
- ^ 16/26, Хасан и др. 2008 год
- ^ Jump up to: а б с д и ж г час я дж к Андерхилл П.А., Шен П., Лин А.А., Джин Л., Пассарино Г., Ян В.Х., Кауфман Э., Бонне-Тамир Б., Бертранпетит Дж., Франкалаччи П., Ибрагим М., Дженкинс Т., Кидд Дж.Р., Мехди С.К., Сейелстад М.Т., Уэллс Р.С., Пьяцца А, Дэвис Р.В., Фельдман М.В., Кавалли-Сфорца Л.Л., Офнер П.Дж. (ноябрь 2000 г.). «Вариации последовательности Y-хромосомы и история человеческих популяций». Природная генетика . 26 (3): 358–61. дои : 10.1038/81685 . ПМИД 11062480 . S2CID 12893406 .
- ^ Jump up to: а б с д и Семино О, Сантакьяра-Бенерекетти А.С., Фаласки Ф, Кавалли-Сфорца ЛЛ, Андерхилл, Пенсильвания (январь 2002 г.). «Эфиопы и койсанцы разделяют самые глубокие клады филогении Y-хромосомы человека » Американский журнал генетики человека . 70 (1): 265–8. дои : 10.1086/338306 . ПМЦ 384897 . ПМИД 11719903 .
- ^ Ди Джакомо Ф, Лука Ф, Ананью Н, Чаварелла Дж, Корбо Р.М., Креста М, Куччи Ф, Ди Стази Л, Агостиано В, Гипараки М, Лутрадис А, Мамми К, Михалодимитракис Э.Н., Папола Ф, Педичини Г, Плата Е , Терренато Л., Тофанелли С., Маласпина П., Новеллетто А. (сентябрь 2003 г.). «Клинические закономерности разнообразия Y-хромосомы человека в континентальной Италии и Греции определяются эффектами дрейфа и основателя». Молекулярная филогенетика и эволюция . 28 (3): 387–95. дои : 10.1016/S1055-7903(03)00016-2 . ПМИД 12927125 .
- ^ Капелли С, Рыжая Н, Романо В, Кали Ф, Лефранк Г, Делаг В, Мегарбан А, Феличе А.Э., Паскали В.Л., Неофиту П.И., Пулли З., Новеллетто А., Маласпина П., Терренато Л., Беребби А., Феллоус М., Томас М.Г. , Гольдштейн Д.Б. (март 2006 г.). «Структура населения в бассейне Средиземноморья: перспектива Y-хромосомы». Анналы генетики человека 70 (Часть 2): 207–25. дои : 10.1111/j.1529–8817.2005.00224.x . hdl : 2108/37090 . ПМИД 16626331 . S2CID 25536759 .
- ^ Абу-Амеро К.К., Хеллани А., Гонсалес А.М., Ларруга Х.М., Кабрера В.М., Андерхилл, Пенсильвания (2009). «Разнообразие Y-хромосомы Саудовской Аравии и его связь с близлежащими регионами» . БМК Генетика . 10 (59): 59. дои : 10.1186/1471-2156-10-59 . ПМК 2759955 . ПМИД 19772609 .
- ^ Jump up to: а б Международное общество генетической генеалогии. «Дерево гаплогрупп Y-ДНК» .
- ^ Jump up to: а б Роза А., Орнелас С., Джоблинг М.А., Брем А., Виллемс Р. (2007). «Y-хромосомное разнообразие населения Гвинеи-Бисау: многоэтническая перспектива» . Эволюционная биология BMC . 7 : 124. дои : 10.1186/1471-2148-7-124 . ЧВК 1976131 . ПМИД 17662131 .
- ^ Андерхилл П.А., Пассарино Г., Лин А.А., Шен П., Мирасон Лар М., Фоли Р.А., Офнер П.Дж., Кавалли-Сфорца Л.Л. (январь 2001 г.). «Филогеография бинарных гаплотипов Y-хромосомы и происхождение современных человеческих популяций» . Анналы генетики человека . 65 (Часть 1): 43–62. дои : 10.1046/j.1469-1809.2001.6510043.x . ПМИД 11415522 . S2CID 9441236 .
- ^ Jump up to: а б Шен П., Лави Т., Кивисилд Т., Чоу В., Сенгун Д., Гефель Д., Шпирер И., Вульф Э., Хиллель Дж., Фельдман М.В., Офнер П.Дж. (сентябрь 2004 г.). «Реконструкция отцовских и матрилиний самаритян и других израильских популяций на основе вариаций последовательностей Y-хромосомы и митохондриальной ДНК». Человеческая мутация . 24 (3): 248–60. дои : 10.1002/humu.20077 . ПМИД 15300852 . S2CID 1571356 .
- ^ Jump up to: а б Кручиани Ф, Тромбетта Б, Селитто Д, Массаи А, Дестро-Бисол Г, Уотсон Е, Беро Коломб Е, Дюгужон Дж. М., Мораль П., Скозцари Р. (июль 2010 г.). «Гаплогруппа R-V88 Y-хромосомы человека: отцовская генетическая запись транссахарских связей начала среднего голоцена и распространения чадских языков» . Европейский журнал генетики человека . 18 (7): 800–7. дои : 10.1038/ejhg.2009.231 . ПМЦ 2987365 . ПМИД 20051990 .
- ^ Jump up to: а б с д Луис-младший, Роуолд Д.Д., Регейру М., Каейро Б., Чинниоглу С., Роузман С., Андерхилл П.А., Кавалли-Сфорца Л.Л., Эррера Р.Дж. (март 2004 г.). «Левант против Африканского Рога: свидетельства двунаправленных коридоров миграции людей» . Американский журнал генетики человека . 74 (3): 532–44. дои : 10.1086/382286 . ПМЦ 1182266 . ПМИД 14973781 .
- ^ Jump up to: а б Кинг Т.Э., Паркин Э.Дж., Суинфилд Г., Кручиани Ф., Скоццари Р., Роза А., Лим С.К., Сюэ Ю., Тайлер-Смит С., Джоблинг М.А. (март 2007 г.). «Африканцы в Йоркшире? Самая глубоко укоренившаяся клада филогении Y в английской генеалогии» . Европейский журнал генетики человека . 15 (3): 288–93. дои : 10.1038/sj.ejhg.5201771 . ПМК 2590664 . ПМИД 17245408 .
Новостная статья: «Йоркширский клан связан с Африкой» . Новости Би-би-си . 24 января 2007 г. Проверено 27 января 2007 г. - ^ Гонсалвес Р., Роза А., Фрейтас А., Фернандес А., Кивисилд Т., Виллемс Р., Брем А. (ноябрь 2003 г.). «Линии Y-хромосомы на островах Кабо-Верде свидетельствуют о разнообразном географическом происхождении первых поселенцев-мужчин». Генетика человека . 113 (6): 467–72. дои : 10.1007/s00439-003-1007-4 . hdl : 10400.13/3047 . ПМИД 12942365 . S2CID 63381583 .
- ^ Абу-Амеро К.К., Хеллани А., Гонсалес А.М., Ларруга Х.М., Кабрера В.М., Андерхилл, Пенсильвания (22 сентября 2009 г.). «Разнообразие Y-хромосомы Саудовской Аравии и его связь с близлежащими регионами» . БМК Генетика . 10:59 . дои : 10.1186/1471-2156-10-59 . ПМК 2759955 . ПМИД 19772609 .
- ^ Хишам Ю. Хасан и др. (2008). «Южный Судан» включает 26 динка, 15 шиллуков и 12 нуэров. «Западный Судан» включает 26 боргу, 32 масалита и 32 фура. «Северный Судан» включает 39 нубийцев, 42 беджа, 33 копта, 50 гаалиен, 28 месерий и 24 аракиена.
- ^ Чинниоглу С., Кинг Р., Кивисилд Т. и др. (2004). «Раскопки слоев гаплотипов Y-хромосомы в Анатолии». Генетика человека . 114 (2): 127–148. дои : 10.1007/s00439-003-1031-4 . ПМИД 14586639 . S2CID 10763736 .
- ^ Небель А., Филон Д., Бринкманн Б., Маджумдер П.П., Фаерман М., Оппенгейм А. (ноябрь 2001 г.). «Пул Y-хромосомы евреев как часть генетического ландшафта Ближнего Востока» . Американский журнал генетики человека . 69 (5): 1095–112. дои : 10.1086/324070 . ПМЦ 1274378 . ПМИД 11573163 .
- ^ Семино О, Пассарино Г, Офнер П.Дж., Лин А.А., Арбузова С., Бекман Л.Е., Де Бенедиктис Г., Франкалаччи П., Коуваци А., Лимборска С., Марсикиаэ М., Мика А., Мика Б., Приморак Д., Сантакьяра-Бенеректи А.С., Кавалли- Сфорца Л.Л., Андерхилл, Пенсильвания (ноябрь 2000 г.). «Генетическое наследие палеолитического Homo sapiens sapiens у современных европейцев: взгляд на Y-хромосому». Наука 290 (5494): 1155–9. Бибкод : 2000Sci... 290.1155S дои : 10.1126/science.290.5494.1155 . ПМИД 11073453 .
- ^ Флорес С., Мака-Мейер Н., Ларруга Дж.М., Кабрера В.М., Карадше Н., Гонсалес А.М. (2005). «Изоляты в коридоре миграций: анализ вариаций Y-хромосомы с высоким разрешением в Иордании» . Журнал генетики человека . 50 (9): 435–41. дои : 10.1007/s10038-005-0274-4 . ПМИД 16142507 .
- ^ Jump up to: а б Карафет Т.М., Мендес Ф.Л., Мейлерман М.Б., Андерхилл П.А., Зегура С.Л., Хаммер М.Ф. (май 2008 г.). «Новые бинарные полиморфизмы изменяют форму и увеличивают разрешение дерева гаплогрупп Y-хромосомы человека» . Геномные исследования . 18 (5): 830–8. дои : 10.1101/гр.7172008 . ПМК 2336805 . ПМИД 18385274 .
- ^ Кран, Томас. «Дерево YCC» . Хьюстон, Техас: FTDNA. Архивировано из оригинала 26 июля 2011 года . Проверено 16 мая 2011 г.
Источники таблиц преобразования
[ редактировать ]- Капелли, Кристиан; Уилсон, Джеймс Ф.; Ричардс, Мартин; Штумпф, Майкл П.Х.; и др. (февраль 2001 г.). «Отцовское наследие преимущественно коренных народов австронезийскоязычных народов островной Юго-Восточной Азии и Океании» . Американский журнал генетики человека . 68 (2): 432–443. дои : 10.1086/318205 . ПМЦ 1235276 . ПМИД 11170891 .
- Хаммер, Майкл Ф.; Карафет Татьяна М.; Редд, Алан Дж.; Джарджанази, Хамди; и др. (1 июля 2001 г.). «Иерархические закономерности глобального разнообразия Y-хромосом человека» . Молекулярная биология и эволюция . 18 (7): 1189–1203. doi : 10.1093/oxfordjournals.molbev.a003906 . ПМИД 11420360 .
- Джоблинг, Марк А.; Тайлер-Смит, Крис (2000), «Новое использование новых гаплотипов», Trends in Genetics , 16 (8): 356–62, doi : 10.1016/S0168-9525(00)02057-6 , PMID 10904265
- Каладжиева, преп.; Калафель, Франческ; Джоблинг, Марк А; Ангеличева, Дора; и др. (февраль 2001 г.). «Закономерности меж- и внутригруппового генетического разнообразия у влакс-рома, выявленные с помощью Y-хромосомы и линий митохондриальной ДНК» . Европейский журнал генетики человека . 9 (2): 97–104. дои : 10.1038/sj.ejhg.5200597 . ПМИД 11313742 .
- Карафет, Татьяна; Сюй, Липин; Ду, Руофу; Ван, Уильям; и др. (сентябрь 2001 г.). «История отцовского населения Восточной Азии: источники, закономерности и микроэволюционные процессы» . Американский журнал генетики человека . 69 (3): 615–628. дои : 10.1086/323299 . ПМЦ 1235490 . ПМИД 11481588 .
- Семино, О.; Пассарино, Дж; Оефнер, П.Дж.; Лин, А.А.; и др. (2000), «Генетическое наследие палеолитического Homo sapiens sapiens у современных европейцев: хромосомная перспектива AY», Science , 290 (5494): 1155–9, Бибкод : 2000Sci...290.1155S , doi : 10.1126/science.290.5494. 1155 , PMID 11073453
- Су, Бинг; Сяо, Цзюньхуа; Андерхилл, Питер; Дека, Ранджан; и др. (декабрь 1999 г.). «Y-хромосома свидетельствует о миграции современных людей на север в Восточную Азию во время последнего ледникового периода» . Американский журнал генетики человека . 65 (6): 1718–1724. дои : 10.1086/302680 . ПМЦ 1288383 . ПМИД 10577926 .
- Андерхилл, Питер А.; Шен, Пейдун; Лин, Алиса А.; Джин, Ли; и др. (ноябрь 2000 г.). «Вариации последовательности Y-хромосомы и история человеческих популяций». Природная генетика . 26 (3): 358–361. дои : 10.1038/81685 . ПМИД 11062480 . S2CID 12893406 .