Гаплогруппа M-P256
| Гаплогруппа M-P256 | |
|---|---|
 | |
| Возможное время происхождения | 32 000–47 000 лет назад ( Шейнфельдт, 2006 г. ) |
| Возможное место происхождения | Уоллеса (восточная Индонезия) или Новая Гвинея [ 1 ] |
| Предок | К2б1 |
| Определение мутаций | Р256 |
Гаплогруппа M , также известная как M-P256 и гаплогруппа K2b1b (ранее K2b1d), представляет собой гаплогруппу ДНК Y-хромосомы . M-P256 является потомком гаплогруппы K2b1 и, как полагают, впервые появилась между 32 000 и 47 000 лет назад ( Scheinfeldt 2006 ).
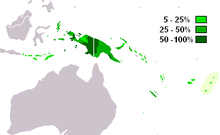
M-P256 является наиболее часто встречающейся гаплогруппой Y-хромосомы в Западном Папуа и западной части Папуа-Новой Гвинеи ( Kayser 2003 ). Он также встречается в соседних частях Меланезии , Индонезии и среди коренных австралийцев .
И M-P256, и гаплогруппа S (B254) являются единственными первичными субкладами K2b1 , также известными как гаплогруппа MS.
Филогенетическая структура
[ редактировать ]Это филогенетическое древо субкладов гаплогрупп основано в первую очередь на деревьях, опубликованных ISOGG в 2016 году ( ISOGG 2016 ) и YCC в 2008 году ( Карафет 2008 ).
- М* (P256)
- М1 (М4, М5/П73, М106, М186, М189, М296, П35)
- М1а (П34_1, П34_2, П34_3, П34_4, П34_5)
- М1а1 (П51)
- М1а2 (Р94)
- М1б (Р87)
- М1б1 (М104_1/П22_1, М104_2/П22_2)
- М1б1а (М16)
- М1b1b (М83)
- М1б1 (М104_1/П22_1, М104_2/П22_2)
- М1а (П34_1, П34_2, П34_3, П34_4, П34_5)
- М2 (М353, М387)
- М2а (M177/SRY9138)
- М3 (П117, П118)
- М1 (М4, М5/П73, М106, М186, М189, М296, П35)
Распределение
[ редактировать ]М* (М-П256*)
[ редактировать ]Парагруппа M-P256* встречается редко. [ написание? ] в Новой Гвинее (6,3%) и Флоресе (2,5%). [ 1 ]
М1 (М-М4)
[ редактировать ]| Гаплогруппа М-М4 | |
|---|---|
| Возможное время происхождения | 8200 [3800–20600] лет назад ( Кайзер 2003 ) |
| Возможное место происхождения | Юго-Восточная Азия – Меланезия [ нужна ссылка ] |
| Предок | М-П256 |
| Определение мутаций | М4, М5/П73, М106, М186, М189, М296, П35 [ нужна ссылка ] |
Часто встречается в Новой Гвинее и Меланезии , умеренно распространен в соседних частях Индонезии , Микронезии и Полинезии .
- 100 % ( Кайзер, 2003 г. )
- Кетенгбан 100% ( Кайзер, 2003 )
- Воздух 100% ( Кайзер, 2003 г. )
- Распечатать 86% ( Кайзер, 2003 г. )
- Асмат 75% ( Кайзер 2003 )
- Западное Папуа
- Низины/побережье 77,5% ( Кайзер, 2003 г. )
- Хайленд 74,5% ( Кайзер, 2003 г. )
- Комбайн / Крокодил 46% ( Кайзер 2003)
- Папуа-Новая Гвинея
- Побережье 29% ( Кайзер, 2003 г. )
- Хайленд 35,5% ( Кайзер, 2003 г. )
- Толай (Новая Британия) 31% ( Кайзер, 2003 г.)
- Острова Тробриан 30% ( Кайзер, 2003 г. )
- Малуку (Молуккские острова) 21% ( Кайзер, 2003 г. )
- Жители островов Торресова пролива (Австралия): до 2,0% – т.е. 0,9% выборок, при этом 45% от общего числа считались «некоренными» ( Nagle 2015 ) .
Крайнее географическое отклонение, очевидно, было выявлено в исследовании 2012 года, в котором сообщалось о хазарейце из Мазари-Шарифа , Афганистан, с M1 среди выборки из 60 мужчин из Мазари-Шарифа ( Haber 2012 ). У хазарейца был SNP M186 (который, как полагают, эквивалентен M4).
| Старые имена (YCC 2002/2008) | М-М4 |
| Джоблинг и Тайлер-Смит, 2000 г. | 24 |
| Андерхилл 2000 | VIII |
| Хаммер 2001 | 1U |
| Карафет 2001 | 37 |
| Семино 2000 | ЕС16 |
| Они в 1999 году | H17 |
| Волосы 2001 | И |
| YCC 2002 (от руки) | М* |
| YCC 2005 (от руки) | М |
| YCC 2008 (от руки) | М1 |
| YCC 2010r (от руки) | М1 |
М1а (М-П34)
[ редактировать ]M1a (M-P34) — наиболее часто встречающаяся гаплогруппа ДНК Y-хромосомы в Западной Новой Гвинее . Он также встречается с умеренной частотой в соседних частях Индонезии ( Малуку , Нуса-Тенгара ) и по всей Папуа-Новой Гвинее , включая прибрежные острова ( Карафет, 2005 и Кайзер, 2008 ).
| Старые имена (YCC 2002/2008) | М-П34 |
| Джоблинг и Тайлер-Смит, 2000 г. | 24 |
| Андерхилл 2000 | VIII |
| Хаммер 2001 | 1U |
| Карафет 2001 | 37 |
| Семино 2000 | ЕС16 |
| Они в 1999 году | H17 |
| Волосы 2001 | И |
| YCC 2002 (от руки) | М1 |
| YCC 2005 (от руки) | М1 |
| YCC 2008 (от руки) | М1а |
| YCC 2010r (от руки) | М1а |
М1б (М-П87)
[ редактировать ]M1b M-P87(xM104/P22) был обнаружен примерно в 18% (20/109) пула образцов из Новой Ирландии , примерно в 12% (5/43) образца Лавонгаи из Нью-Ганновера , примерно в 5%. (19/395) пула образцов из Новой Британии (и, в частности, примерно в 24% (15/63) Бейнинга из Восточной Новой Британии ), а также одна особь сапоса с севера Бугенвиля и еще одна особь с северного побережья Папуа-Новой Гвинеи ( Scheinfeldt 2006 ).
Субклад M1b1 (M104_1/P22_1, M104_2/P22_2) часто встречается в популяциях архипелага Бисмарка и острова Бугенвиль , с умеренным распространением в Новой Гвинее , Фиджи , Тонге , Восточной Футуне и Самоа . ( Кайзер 2008 и Шейнфельдт 2006 ).
| Старые имена (YCC 2002/2008) | М-П22 |
| Джоблинг и Тайлер-Смит, 2000 г. | 24 |
| Андерхилл 2000 | VIII |
| Хаммер 2001 | 1U |
| Карафет 2001 | 38 |
| Семино 2000 | ЕС16 |
| Они в 1999 году | H17 |
| Волосы 2001 | И |
| YCC 2002 (от руки) | М2* |
| YCC 2005 (от руки) | М2а |
| YCC 2008 (от руки) | М1б1 |
| YCC 2010r (от руки) | М1б1 |
М2(М-М353)
[ редактировать ]M2 встречается с низкой частотой на Фиджи и в Восточной Футуне ( Kayser 2006 ).
Субклад M2a (M-M177, также называемый M-SRY9138) был обнаружен у одной особи насиои с восточного побережья Бугенвиля и у одной особи из провинции Малаита на Соломоновых островах ( Cox 2006 ).
Альтернативные названия субклада M2a, ранее использовавшиеся в рецензируемой литературе, перечислены ниже.
| Старые имена (YCC 2002/2008) | К-SRY9138/М-SRY9138 РУЧНОЙ М-М177 |
| Джоблинг и Тайлер-Смит, 2000 г. | 23 |
| Андерхилл 2000 | VIII |
| Хаммер 2001 | 1Е |
| Карафет 2001 | 25 |
| Семино 2000 | ЕС16 |
| Они в 1999 году | Н5 |
| Волосы 2001 | Ф |
| YCC 2002 (от руки) | 1 квартал |
| YCC 2005 (от руки) | 1 квартал |
| YCC 2008 (от руки) | М2а |
| YCC 2010r (от руки) | М2а |
М3 (М-П117)
[ редактировать ]M3 (P117, P118) часто встречается в популяциях Новой Британии , а также иногда наблюдается на севере Бугенвиля , Фиджи и Восточной Футуне ( Kayser 2008 и Scheinfeldt 2006 ).
Предыдущая филогенетическая история
[ редактировать ]До 2002 года в академической литературе использовалось по крайней мере семь различных систем наименования филогенетического дерева Y-хромосомы, что приводило к значительной путанице. Чтобы решить эту проблему, в 2002 году основные исследовательские группы объединились и сформировали Консорциум Y-хромосомы (YCC), в результате чего была опубликована совместная статья, содержащая одно новое дерево, которое все они согласились использовать. Позже группа гражданских ученых, интересующихся популяционной генетикой и генетической генеалогией, сформировала рабочую группу для создания любительского дерева, стремясь быть прежде всего своевременным.
В таблице ниже собрана номенклатура, использовавшаяся в исследованиях гаплогруппы M до знаменательного дерева YCC 2002 года, что позволяет исследователям, просматривающим более старую литературу, быстро переходить между различными использовавшимися номенклатурами.
| YCC 2002/2008 (сокращенно) | (а) | (б) | (с) | (г) | (е) | (г) | (или) | YCC 2002 (от руки) | YCC 2005 (от руки) | YCC 2008 (от руки) | YCC 2010r (от руки) | ИСОГГ 2006 г. | ИСОГГ 2007 | ИСОГГ 2008 | ИСОГГ 2009 | ИСОГГ 2010 | ИЗОГГ 2011 | ИЗОГГ 2012 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| М4 | 24 | VIII | 1U | 37 | ЕС16 | H17 | И | М* | М | М1 | М1 | - | - | - | - | - | - | - |
| М-П34 | 24 | VIII | 1U | 37 | ЕС16 | H17 | И | М1 | М1 | М1а | М1а | - | - | - | - | - | - | - |
| М-П22/М-М104 | 24 | VIII | 1U | 38 | ЕС16 | H17 | И | М2* | М2а | М1б1 | М1б1 | - | - | - | - | - | - | - |
| М-М16 | 24 | VIII | 1U | 39 | ЕС16 | H17 | И | М2а | М2а1 | M1b1a | M1b1a | - | - | - | - | - | - | - |
| М-М83 | 24 | VIII | 1U | 38 | ЕС16 | H17 | И | М2б | М2а2 | M1b1b | M1b1b | - | - | - | - | - | - | - |
| К-SRY9138/М-SRY9138 | 23 | VIII | 1Е | 25 | ЕС16 | Н5 | Ф | 1 квартал | 1 квартал | М2а | М2а | - | - | - | - | - | - | - |
- Источники
Следующие исследовательские группы в соответствии с их публикациями были представлены в создании Дерева YCC.
В статье Карафета 2008 года был внесен ряд изменений по сравнению с предыдущим деревом ISOGG 2006 года . До открытия маркера P256 нынешняя подгруппа M-M4 (определяемая маркером M4) ранее представляла всю гаплогруппу M-P256; родительской гаплогруппы K. а подгруппы M2 и M3 ранее классифицировались как подгруппы K1 и K7 [ нужна ссылка ]
Ссылки
[ редактировать ]Сноски
[ редактировать ]- ^ Jump up to: а б Татьяна М. Карафет, Брайан Холлмарк, Мюррей П. Кокс, Херавати Судойо , Шон Дауни, Дж. Стивен Лансинг и Майкл Ф. Хаммер, «Основное разделение Восток-Запад лежит в основе стратификации Y-хромосомы в Индонезии», Molecular Biological Evolution , (2010), vol. 27, нет. 8, стр. 1833-1844.
Цитируемые работы
[ редактировать ]- Кокс, Мюррей П.; Мирасон Лар, Марта (2006). «Разнообразие Y-хромосом обратно пропорционально связано с языковой принадлежностью в парных австронезийских и папуасоязычных общинах Соломоновых Островов». Американский журнал биологии человека . 18 (1): 35–50. дои : 10.1002/ajhb.20459 . ПМИД 16378340 . S2CID 4824401 .
- Хабер, Марк; Платт, Дэниел Э.; Ашрафиан Бонаб, Мазиар; Юханна, Соня С.; Сория-Эрнанц, Дэвид Ф.; Мартинес-Крус, Бегонья; Дуэи, Бушра; Гассибе-Саббах, Мишелла; Рафатпана, Хошанг (2012). Кайзер, Манфред (ред.). «Этнические группы Афганистана разделяют Y-хромосомное наследие, структурированное историческими событиями» . ПЛОС ОДИН . 7 (3): e34288. Бибкод : 2012PLoSO...734288H . дои : 10.1371/journal.pone.0034288 . ПМЦ 3314501 . ПМИД 22470552 .
- Карафет Татьяна М.; Лансинг, Дж. С. ; Редд, Алан Дж.; Уоткинс, Джозеф К.; Сурата, СПК; Артхавигуна, Вашингтон; Майер, Лаура; Бамшад, Майкл; Джорд, Линн Б. (2005). «Балийский взгляд на Y-хромосому на заселение Индонезии: генетический вклад донеолитических охотников-собирателей, австронезийских фермеров и индийских торговцев» (PDF) . Биология человека . 77 (1): 93–114. дои : 10.1353/hub.2005.0030 . hdl : 1808/13586 . ПМИД 16114819 . S2CID 7953854 .
- Карафет, ТМ; Мендес, Флорида; Мейлерман, МБ; Андерхилл, Пенсильвания; Зегура, СЛ; Хаммер, МФ (2008). «Новые бинарные полиморфизмы изменяют форму и увеличивают разрешение дерева гаплогрупп Y-хромосомы человека» . Геномные исследования . 18 (5): 830–8. дои : 10.1101/гр.7172008 . ПМК 2336805 . ПМИД 18385274 .
- Кайзер, Манфред; Брауэр, Силке; Вайс, Гюнтер; Шифенхёвель, Вульф; Андерхилл, Питер; Шен, Пейдун; Офнер, Питер; Томмазео-Понцетта, Мила; Стоункинг, Марк (2003). «Уменьшенная Y-хромосома, но не митохондриальная ДНК, разнообразие человеческих популяций из Западной Новой Гвинеи» . Американский журнал генетики человека . 72 (2): 281–302. дои : 10.1086/346065 . ПМК 379223 . ПМИД 12532283 .
- Кайзер, М.; Брауэр, С; Кордо, Р; Касто, А; Лао, О; Животовский, Л.А.; Мойс-Фори, К; Ратледж, РБ; Шифенхувел, W (2006). «Меланезийское и азиатское происхождение полинезийцев: градиенты мтДНК и Y-хромосомы в Тихом океане» . Молекулярная биология и эволюция . 23 (11): 2234–44. дои : 10.1093/molbev/msl093 . hdl : 11858/00-001M-0000-0010-0145-0 . ПМИД 16923821 .
- Кайзер, М.; Чой, Ю.; Ван Овен, М.; Мона, С.; Брауэр, С.; Трент, Р.Дж.; Суаркиа, Д.; Шифенховел, В.; Стоункинг, М. (2008). «Влияние австронезийской экспансии: данные о разнообразии мтДНК и Y-хромосомы на Адмиралтейских островах Меланезии» . Молекулярная биология и эволюция . 25 (7): 1362–74. дои : 10.1093/molbev/msn078 . ПМИД 18390477 .
- Шейнфельдт, Л.; Фридлендер, Ф; Фридлендер, Дж; Лэтэм, К; Коки, Г; Карафет, Т; Хаммер, М; Лоренц, Дж (2006). «Неожиданная хромосомная вариация NRY на Северном острове Меланезии» . Молекулярная биология и эволюция . 23 (8): 1628–41. дои : 10.1093/molbev/msl028 . ПМИД 16754639 .
- Нэгл, Н.; и др. (2015). «Древность и разнообразие Y-хромосом аборигенов Австралии». Американский журнал физической антропологии . 159 (3): 367–81. дои : 10.1002/ajpa.22886 . ПМИД 26515539 .
- «ISOGG 2016 Гаплогруппа M Y-ДНК» . www.isogg.org .
Источники таблиц преобразования
[ редактировать ]- Капелли, Кристиан; Уилсон, Джеймс Ф.; Ричардс, Мартин; Штумпф, Майкл П.Х.; и др. (февраль 2001 г.). «Отцовское наследие преимущественно коренных народов австронезийскоязычных народов островной Юго-Восточной Азии и Океании» . Американский журнал генетики человека . 68 (2): 432–443. дои : 10.1086/318205 . ПМЦ 1235276 . ПМИД 11170891 .
- Хаммер, Майкл Ф.; Карафет Татьяна М.; Редд, Алан Дж.; Джарджанази, Хамди; и др. (1 июля 2001 г.). «Иерархические закономерности глобального разнообразия Y-хромосом человека» . Молекулярная биология и эволюция . 18 (7): 1189–1203. doi : 10.1093/oxfordjournals.molbev.a003906 .
- Джоблинг, Марк А.; Тайлер-Смит, Крис (2000), «Новое использование новых гаплотипов», Trends in Genetics , 16 (8): 356–62, doi : 10.1016/S0168-9525(00)02057-6 , PMID 10904265
- Каладжиева, преп.; Калафель, Франческ; Джоблинг, Марк А; Ангеличева, Дора; и др. (февраль 2001 г.). «Закономерности меж- и внутригруппового генетического разнообразия у влакс-рома, выявленные с помощью Y-хромосомы и линий митохондриальной ДНК» . Европейский журнал генетики человека . 9 (2): 97–104. дои : 10.1038/sj.ejhg.5200597 .
- Карафет, Татьяна; Сюй, Липин; Ду, Руофу; Ван, Уильям; и др. (сентябрь 2001 г.). «История отцовского населения Восточной Азии: источники, закономерности и микроэволюционные процессы» . Американский журнал генетики человека . 69 (3): 615–628. дои : 10.1086/323299 . ПМЦ 1235490 . ПМИД 11481588 .
- Семино, О.; Пассарино, Дж; Оефнер, П.Дж.; Лин, А.А.; и др. (2000), «Генетическое наследие палеолитического Homo sapiens sapiens у современных европейцев: хромосомная перспектива AY», Science , 290 (5494): 1155–9, Бибкод : 2000Sci...290.1155S , doi : 10.1126/science.290.5494. 1155 , ПМИД 11073453
- Су, Бинг; Сяо, Цзюньхуа; Андерхилл, Питер; Дека, Ранджан; и др. (декабрь 1999 г.). «Y-хромосома свидетельствует о миграции современных людей на север в Восточную Азию во время последнего ледникового периода» . Американский журнал генетики человека . 65 (6): 1718–1724. дои : 10.1086/302680 . ПМЦ 1288383 . ПМИД 10577926 .
- Андерхилл, Питер А.; Шен, Пейдун; Лин, Алиса А.; Джин, Ли; и др. (ноябрь 2000 г.). «Вариации последовательности Y-хромосомы и история человеческих популяций». Природная генетика . 26 (3): 358–361. дои : 10.1038/81685 .
Внешние ссылки
[ редактировать ]См. также
[ редактировать ]- Генетическая генеалогия
- Гаплогруппа
- Гаплотип
- Гаплогруппа ДНК Y-хромосомы человека
- Молекулярная филогения
- Парагруппа
- Субклад
- Гаплогруппы Y-хромосомы в популяциях мира
- Гаплогруппы Y-ДНК по этническим группам
- Гаплогруппы Y-ДНК в популяциях Восточной и Юго-Восточной Азии