Гаплогруппа
| Часть серии о |
| Генетическая генеалогия |
|---|
| Концепции |
| Связанные темы |
Гаплотип – это группа аллелей в организме, унаследованных вместе от одного родителя. [1] [2] и гаплогруппа ( гаплоид от греческого : ἁπλοῦς , haploûs , «однократный, простой» и английского: группа ) — это группа схожих гаплотипов, которые имеют общего предка с однонуклеотидного полиморфизма мутацией . [3] Более конкретно, гаплотип — это комбинация аллелей в разных хромосомных регионах, которые тесно связаны и имеют тенденцию наследоваться вместе. Поскольку гаплогруппа состоит из схожих гаплотипов, гаплогруппу обычно можно предсказать по гаплотипам. Гаплогруппы относятся к одной линии происхождения . Таким образом, членство любого человека в гаплогруппе зависит от относительно небольшой части генетического материала, которым обладает этот человек.

и остается ее частью Каждая гаплогруппа происходит от предыдущей отдельной гаплогруппы (или парагруппы ) . Таким образом, любая родственная группа гаплогрупп может быть точно смоделирована как вложенная иерархия , в которой каждый набор (гаплогруппа) также является подмножеством одного более широкого набора (в отличие от моделей с двумя родителями, таких как генеалогические древа человека). . Гаплогруппы можно разделить на субклады.
Гаплогруппы обычно идентифицируются по начальной букве алфавита, а уточнения состоят из дополнительных комбинаций цифр и букв, таких как (например) A → A1 → A1a . Алфавитная номенклатура была опубликована в 2002 году Консорциумом Y-хромосомы . [4]
В генетике человека наиболее часто изучаемыми гаплогруппами являются гаплогруппы Y-хромосомы (Y-ДНК) и гаплогруппы митохондриальной ДНК (мтДНК) , каждая из которых может использоваться для определения генетических популяций . Y-ДНК передается исключительно по отцовской линии, от отца к сыну, тогда как мтДНК передается по материнской линии, от матери к потомкам обоих полов. Ни один из них не рекомбинирует , и, таким образом, Y-ДНК и мтДНК изменяются только в результате случайной мутации в каждом поколении без смешения генетического материала родителей.
Формирование гаплогруппы
[ редактировать ]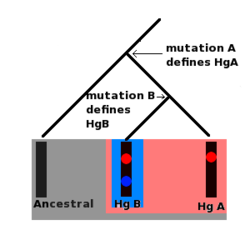
Митохондрии — это небольшие органеллы , которые лежат в цитоплазме эукариотических клеток , например клеток человека. Их основная функция – обеспечение клетки энергией. Считается, что митохондрии являются уменьшенными потомками симбиотических бактерий , которые когда-то жили свободно. Одним из признаков того, что митохондрии когда-то были свободноживущими, является то, что каждая из них содержит кольцевую ДНК , называемую митохондриальной ДНК (мтДНК), структура которой больше похожа на бактерии, чем на эукариотические организмы (см. Эндосимбиотическую теорию ). Подавляющее большинство ДНК человека содержится в хромосомах ядра . клетки, но мтДНК является исключением Индивидуум наследует свою цитоплазму и органеллы, содержащиеся в этой цитоплазме, исключительно от материнской яйцеклетки (яйцеклетки); Сперматозоиды передают только хромосомную ДНК, все отцовские митохондрии перевариваются в ооците . Таким образом, когда мутация возникает в молекуле мтДНК, она передается по прямой женской линии происхождения. Мутации – это изменения азотистых оснований Последовательность ДНК . Одиночные изменения исходной последовательности называются однонуклеотидными полиморфизмами (SNP). [ сомнительно – обсудить ]
человека Y-хромосомы представляют собой мужские половые хромосомы ; будут почти все люди, обладающие Y-хромосомой, морфологически мужчинами. Хотя Y-хромосомы расположены в ядре клетки и спарены с Х-хромосомами , они рекомбинируют с Х-хромосомой только на концах Y-хромосомы ; оставшиеся 95% Y-хромосомы не рекомбинируются. Таким образом, Y-хромосома и любые возникающие в ней мутации передаются по прямой мужской линии.
Другие хромосомы, аутосомы и Х-хромосомы (когда доступна другая Х-хромосома для спаривания с ней), делятся своим генетическим материалом во время мейоза — процесса деления клеток , в результате которого образуются гаметы . Фактически это означает, что генетический материал этих хромосом смешивается в каждом поколении, и поэтому любые новые мутации передаются случайным образом от родителей к потомству.
Особенностью Y-хромосомы и мтДНК является то, что мутации могут накапливаться в определенном сегменте обеих молекул, и эти мутации остаются фиксированными на ДНК. Кроме того, можно также предположить историческую последовательность этих мутаций. Например, если набор из десяти Y-хромосом (полученных от десяти разных людей) содержит мутацию А, но только пять из этих хромосом содержат вторую мутацию В, то должно быть так, что мутация В произошла после мутации А.
Более того, все десять человек, несущих хромосому с мутацией А, являются прямыми потомками по мужской линии того же человека, который был первым человеком, носителем этой мутации. Первый мужчина, несущий мутацию B, также был прямым потомком этого человека по мужской линии, но также является прямым предком по мужской линии всех мужчин, несущих мутацию B. Серии таких мутаций образуют молекулярные линии. Более того, каждая мутация определяет набор определенных Y-хромосом, называемых гаплогруппой.
Все люди, несущие мутацию A, образуют одну гаплогруппу, и все люди, несущие мутацию B, являются частью этой гаплогруппы, но мутация B также определяет более позднюю гаплогруппу (которая является подгруппой или субкладом ), к которой относятся люди, несущие только мутацию A. не принадлежать. И мтДНК, и Y-хромосомы сгруппированы в линии и гаплогруппы; они часто представлены в виде древовидных диаграмм.
Гаплогруппы ДНК Y-хромосомы человека
[ редактировать ]человека Гаплогруппы ДНК Y-хромосомы (Y-ДНК) называются от А до Т и далее подразделяются с использованием цифр и строчных букв. Обозначения гаплогрупп Y-хромосомы устанавливаются Консорциумом Y-хромосомы. [5]
Y-хромосомный Адам — это имя, данное исследователями мужчине, который является по отцовской (мужской) линии самым недавним общим предком всех ныне живущих людей .
Основные гаплогруппы Y-хромосомы и их географические регионы распространения (донедавняя европейская колонизация), включают:
Группы без мутации М168
[ редактировать ]- Гаплогруппа A (M91) (Африка, особенно койсаны и нилоты )
- Гаплогруппа B (M60) (Африка, особенно пигмеи и хадзабе )
Группы с мутацией М168
[ редактировать ](произошла мутация M168 ~50 000 п.н.)
- Гаплогруппа C (M130) (Океания, Северная/Центральная/Восточная Азия, Северная Америка и незначительное присутствие в Южной Америке, Юго-Восточной Азии, Южной Азии, Западной Азии и Европе)
- YAP+ гаплогруппы
- Гаплогруппа DE (M1, M145, M203)
- Гаплогруппа D (CTS3946) (Тибет, Непал, Япония, Андаманские острова, Центральная Азия и спорадическое присутствие в Нигерии, Сирии и Саудовской Аравии)
- Гаплогруппа Е (M96)
- Гаплогруппа E1b1a (V38) Западная Африка и прилегающие регионы; ранее известный как E3a
- Гаплогруппа E1b1b (M215) Связана с распространением афроазиатских языков; в настоящее время сосредоточены в Северной Африке и на Африканском Роге, а также в некоторых частях Ближнего Востока, Средиземноморья и Балкан; ранее известный как E3b
- Гаплогруппа DE (M1, M145, M203)
Группы с мутацией М89
[ редактировать ](произошла мутация M89 ~45 000 п.н.)
- Гаплогруппа F (M89) Океания, Европа, Азия, Северная и Южная Америка.
- Гаплогруппа FT (P14, M213) (Китай, Вьетнам, [6] Сингапур [7] )
- Гаплогруппа G (M201) (присутствует среди многих этнических групп Евразии, обычно с низкой частотой; наиболее распространена на Кавказе, Иранском нагорье и Анатолии; в Европе преимущественно в Греции, Италии, Иберии , Тироле, Богемии; редко в Северной Европа)
- Гаплогруппа H (L901/M2939)
- H1'3 (Z4221/M2826, Z13960)
- H2 (P96) Ранее известная как гаплогруппа F3. Встречается с низкой частотой в Европе и Западной Азии.
- Гаплогруппа IJK (L15, L16)
Группы с мутациями L15 и L16
[ редактировать ]- Гаплогруппа IJK (L15, L16)
- Гаплогруппа IJ (S2, S22)
- Гаплогруппа I (M170, P19, M258) (широко распространена в Европе, нечасто встречается в некоторых частях Ближнего Востока и практически отсутствует в других местах). [8] )
- Гаплогруппа I1 (M253, M307, P30, P40) (Северная Европа, доминирует в Скандинавии)
- Гаплогруппа I2 (S31) (Центральная и Юго-Восточная Европа, Сардиния, Балканы)
- Гаплогруппа J (M304) (Ближний Восток, Турция, Кавказ, Италия, Греция, Балканы, Северная Африка)
- Гаплогруппа J* (в основном встречается на Сокотре , с некоторыми наблюдениями в Пакистане , Омане , Греции , Чехии и среди тюркских народов )
- Гаплогруппа J1 (M267) (в основном связана с семитскими народами на Ближнем Востоке, но также встречается в Средиземноморской Европе, Эфиопии, Северной Африке, Пакистане, Индии и с Северо-Восточного Кавказа народами в Дагестане ; J1 с DYS388 = 13 связана с восточной Анатолией)
- Гаплогруппа J2 (M172) (в основном встречается в Западной Азии, Центральной Азии, Иране, Италии, Греции, на Балканах и в Северной Африке)
- Гаплогруппа I (M170, P19, M258) (широко распространена в Европе, нечасто встречается в некоторых частях Ближнего Востока и практически отсутствует в других местах). [8] )
- Гаплогруппа К (M9, P128, P131, P132)
- Гаплогруппа IJ (S2, S22)
Группы с мутацией М9
[ редактировать ](произошла мутация M9 ~40 000 п.н. )
- Гаплогруппа К
- Гаплогруппа LT (L298/P326)
- Гаплогруппа L (M11, M20, M22, M61, M185, M295) (Южная Азия, Центральная Азия, Юго-Западная Азия, Средиземноморье)
- Гаплогруппа Т (M70, M184/USP9Y+3178, M193, M272) (Северная Африка, Африканский Рог, Юго-Западная Азия, Средиземноморье, Южная Азия); ранее известная как гаплогруппа K2
- Гаплогруппа K(xLT) (rs2033003/M526)
- Гаплогруппа LT (L298/P326)
Группы с мутацией М526
[ редактировать ]- Гаплогруппа M (P256) (Новая Гвинея, Меланезия, восточная Индонезия)
- Гаплогруппа НЕТ (M214)
- Гаплогруппа N (M231) (самый север Евразии)
- Гаплогруппа O (M175) (Восточная Азия, Юго-Восточная Азия, Южная часть Тихого океана, Южная Азия, Центральная Азия)
- Гаплогруппа O1 (F265)
- Гаплогруппа O1a (M119)
- Гаплогруппа O1b (P31, M268)
- Гаплогруппа О2 (M122)
- Гаплогруппа O1 (F265)
- Гаплогруппа P-M45 (M45) (M45 произошла ~ 35 000 п.н.)
- Гаплогруппа Q-M242 (M242) (произошла ~ 15 000–20 000 п.н. Обнаружена в Азии и Америке)
- Гаплогруппа Q-M3 (M3) (Северная Америка, Центральная Америка и Южная Америка)
- Гаплогруппа R (M207)
- Гаплогруппа R1 (M173)
- Гаплогруппа R1a (M17) (Центральная Азия, Южная Азия, Центральная, Северная и Восточная Европа)
- Гаплогруппа R1b (M343) (Европа, Кавказ, Центральная Азия, Южная Азия, Северная Африка, Центральная Африка)
- Гаплогруппа R2 (M124) (Южная Азия, Кавказ, Средняя Азия)
- Гаплогруппа R1 (M173)
- Гаплогруппа Q-M242 (M242) (произошла ~ 15 000–20 000 п.н. Обнаружена в Азии и Америке)
- Гаплогруппа S (M230, P202, P204) (Новая Гвинея, Меланезия, восточная Индонезия)
Гаплогруппы митохондриальной ДНК человека
[ редактировать ]
человека Гаплогруппы мтДНК обозначаются буквами: А , Б , С , Чехия , Д , Э , Ф , Г , Х , ХВ , я , Дж , до JT , Джей Ти , К , Л0 , Л1 , Л2 , Л3 , Л4 , Л5 , Л6 , М , Н , О , П , Вопрос , Р , Р0 , С , Т , У , В , В , Х , Й , и З. Самая последняя версия дерева мтДНК поддерживается Маннисом ван Овеном на веб-сайте PhyloTree. [9]
Филогенетическое древо гаплогрупп митохондриальной ДНК (мтДНК) человека | |||||||||||||||||||||||||||||||||||||||
| Митохондриальная Ева ( L ) | |||||||||||||||||||||||||||||||||||||||
| Л0 | Л1–6 | ||||||||||||||||||||||||||||||||||||||
| Л1 | Л2 | Л3 | Л4 | Л5 | Л6 | ||||||||||||||||||||||||||||||||||
| М | Н | ||||||||||||||||||||||||||||||||||||||
| Чехия | Д | И | Г | вопрос | ТО | А | С | Р | я | В | Х | И | |||||||||||||||||||||||||||
| С | С | Б | Ф | Р0 | до JT | П | В | ||||||||||||||||||||||||||||||||
| Х.В. | Джей Ти | К | |||||||||||||||||||||||||||||||||||||
| ЧАС | V | Дж | Т | ||||||||||||||||||||||||||||||||||||
Митохондриальная Ева — это имя, данное исследователями женщине, которая является по материнской линии (женской линии) последним общим предком всех ныне живущих людей .
Определение популяций
[ редактировать ]
Гаплогруппы могут использоваться для определения генетических популяций и часто имеют географическую ориентацию. Например, следующие общие подразделения гаплогрупп мтДНК:
- Африканские: L0 , L1 , L2 , L3 , L4 , L5 , L6.
- Западноевразийские: H , T , U , V , X , K , I , J , W (все перечисленные западноевразийские гаплогруппы происходят от макрогаплогруппы N ) [10]
- Восточноевразийские: A , B , C , D , E , F , G , Y , Z (примечание: C, D, E, G и Z принадлежат к макрогаплогруппе M )
- Коренной американец: A , B , C , D , X.
- Австрало-меланезийский : P , Q , S
Митохондриальные гаплогруппы делятся на три основные группы, которые обозначаются последовательными буквами L, M, N.Человечество сначала разделилось внутри группы L на L0 и L1-6. L1-6 дали начало другим группам L, одна из которых, L3, разделилась на группу M и N.
Группа М представляет собой первую волну миграции людей, которая, как полагают, возникла за пределами Африки, следуя по восточному маршруту вдоль южных прибрежных районов. Потомки гаплогруппы M сейчас встречаются по всей Азии, Америке и Меланезии, а также в некоторых частях Африканского Рога и Северной Африки; в Европе почти ничего не обнаружено. Гаплогруппа N может представлять собой еще одну макролинию, которая развилась за пределами Африки и направилась на север, а не на восток. Вскоре после миграции большая группа R отделилась от группы N.
Гаплогруппа R состоит из двух подгрупп, определенных на основе их географического распространения: одна встречается в Юго-Восточной Азии и Океании, а другая содержит почти все современное европейское население. Гаплогруппа N(xR), т.е. мтДНК, принадлежащая к группе N, но не к ее подгруппе R, типична для аборигенных популяций Австралии, но также присутствует с низкими частотами среди многих популяций Евразии и Америки.
К типу L принадлежат почти все африканцы.
Тип М состоит из:
M1 – эфиопское, сомалийское и индийское население. Вероятно, из-за большого потока генов между Африканским Рогом и Аравийским полуостровом (Саудовская Аравия, Йемен, Оман), разделенным лишь узким проливом между Красным морем и Аденским заливом.
Чехия – Много сибиряков; ветвь C – некоторые индейцы; ветвь Z – многие саами, некоторые корейцы, некоторые северокитайцы, некоторые народы Центральной Азии.
D – Некоторые американские индейцы, многие сибиряки и выходцы из северной Восточной Азии.
E – малайцы, Борнео, Филиппины, тайваньские аборигены , Папуа-Новая Гвинея.
G - Многие выходцы из Северо-Восточной Сибири, Северной Восточной Азии и Центральной Азии.
Q – меланезийское, полинезийское, новогвинейское население.
Тип N состоит из:
А – встречается у многих американских индейцев, а также у некоторых выходцев из Восточной Азии и Сибири.
I – частота 10% в Северной и Восточной Европе.
S - Некоторые коренные австралийцы (первые народы Австралии)
W – некоторые выходцы из Восточной Европы, Южной Азии и южной части Восточной Азии.
X – некоторые американские индейцы, южные сибиряки, выходцы из Юго-Западной Азии и южные европейцы.
Y – Большинство нивхов и жителей Ниаса ; многие айны, тунгусы и австронезийцы ; также встречается с низкой частотой в некоторых других популяциях Сибири, Восточной Азии и Центральной Азии.
R – большая группа, обнаруженная в типе N. Популяции, содержащиеся там, географически можно разделить на Западную Евразию и Восточную Евразию. Сегодня почти все европейское население и большое количество населения Ближнего Востока относятся к этой ветви. Меньший процент содержится в других группах типа N (см. выше). Ниже приведены субклады R :
Б – некоторые китайцы, тибетцы, монголы, выходцы из Центральной Азии, корейцы, американские индейцы, жители Южной Сибири, японцы, австронезийцы.
F – В основном встречается в Юго-Восточной Азии, особенно во Вьетнаме ; 8,3% на острове Хвар в Хорватии. [11]
R0 – встречается в Аравии, среди эфиопов и сомалийцев; ветка HV (ветвь H; ветка V) – Европа, Западная Азия, Северная Африка;
Pre-JT - возник в Леванте (территория современного Ливана), встречается в 25% случаев среди бедуинов; ветка JT (ветвь J; ветка Т) – Северная, Восточная Европа, Инд, Средиземноморье
U – высокая частота в Западной Евразии, Индийском субконтиненте и Алжире, встречается от Индии до Средиземноморья и остальной Европы; U5, в частности, демонстрирует высокую частоту в странах Скандинавии и Балтии, причем самая высокая частота встречается у саамов .
Назначение географической гаплогруппы Y-хромосомы и мтДНК
[ редактировать ]Вот список географических гаплогрупп Y-хромосомы и мтДНК, предложенный Бекадой и др. 2013. [12]
Y-хромосома
[ редактировать ]Согласно гаплогруппам SNPS, возраст первого вымирания обычно составляет около 45–50 тысяч лет назад. гаплогруппы второго вымирания, по-видимому, разошлись на 32–35 тыс. лет назад По мнению Мальты, . Эпицентральным вымиранием, по-видимому, стал Тоба, во время которого гаплогруппа CDEF*, по-видимому, разделилась на C, DE и F. C и F не имеют почти ничего общего, в то время как D и E имеют много общего. Событие №1, согласно текущим оценкам, произошло после Тобы, хотя более старая древняя ДНК могла отодвинуть событие вымирания от эпицентра задолго до Тобы и отодвинуть первое событие вымирания здесь обратно к Тобе. Гаплогруппы с заметками о событиях вымирания имеют сомнительное происхождение, и это связано с тем, что события вымирания приводят к серьезным узким местам, поэтому все записи этих групп являются лишь догадками. Обратите внимание, что подсчет SNP древней ДНК может сильно различаться, а это означает, что, хотя все эти группы разошлись примерно в одно и то же время, никто не знает, когда. [13] [14]
| Источник | Гаплогруппа | Маркер |
| Европа (Второе событие вымирания?) | я | М170, М253, П259, М227, М507 |
| Европа | I1b | П215, М438, П37.2, М359, П41.2 |
| Европа | I1b2 | М26 |
| Европа | I1c | М223, М284, П78, П95 |
| Европа | J2a1 | М47 |
| Европа | J2a2 | М67, М166 |
| Европа | J2a2a | М92 |
| Европа | J2b | М12, М102, М280, М241 |
| Европа | R1b1b1a | М412, П310 |
| Европа | R1b1b1a1 | Л11 |
| Европа | R1b1b1a1a | U106 |
| Европа | R1b1b1a1b | У198, П312, С116 |
| Европа | R1b1b1a1b1 | U152 |
| Европа | R1b1b1a1b2 | М529 |
| Европа | Р1б1б1а1б3,4 | М65, М153 |
| Европа | R1b1b1a1b5 | SRY2627 |
| Южная Азия или Меланезия | C1 (ранее известный как CxC3) | Z1426 |
| Северная Азия | C2 (ранее известный как C3) | М217+ |
| Индонезия или Южная Азия | Ф | М89, М282 |
| Европа (Кавказ) | Г | М201, М285, П15, П16, М406 |
| Южная Азия | ЧАС | М69, М52, М82, М197, М370 |
| Европа или Ближний Восток | J1 | М304, М267, П58, М365, М368, М369 |
| Европа или Ближний Восток | J2 | М172, М410, М158, М319, ДИС445=6, М339, М340 |
| К западу от Бирмы в Евразии (первое вымирание?) [15] | ||
| Индонезия (первое событие вымирания?) [15] | К2 (НОПС) | М526 |
| Южная Азия | л | М11, М20, М27, М76, М317, М274, М349, М357 |
| Восточная Азия, Юго-Восточная Азия | Н | М231, М214, ЛЛИ22г, Тат, М178 |
| Восточная Азия, Юго-Восточная Азия, Южная Азия | ТО | М175, М119 |
| Индонезия, Филиппины | П (xQR) | 92Р7, М207, М173, М45 |
| Южная Азия, Сибирь | Разделение R и Q (QR) [15] | МЕХ2, М242, П36.2, М25, М346 |
| Ближний Восток, Европа, Сибирь, Южная Азия | R1a1 | М420, М17, М198, М204, М458 |
| Анатолия, Юго-Восточная Европа? | Р1б | М173, М343, П25, М73 |
| Европа | R1b1b | М269 |
| Европа | R1b1b1 | Л23 |
| Пакистан, Индия | Р2 | М479, М124 |
| Средний Восток | Т | М70 |
| Северная Африка | E1b1b1 | М35 |
| Северная Африка | E1b1b1a | М78 |
| Западная Азия | E1b1b1a2 | В13 |
| Северная Африка | E1b1b1a1 | V12 |
| Северная Африка | E1b1b1a1b | В32 |
| Северная Африка | E1b1b1a3 | В22 |
| Северная Африка | E1b1b1a4 | В65 |
| Северная Африка | E1b1b1b | М81 |
| Северная Африка | E1b1b1c | М123, М34 |
| Западная Африка, Северная Африка | А | М91, М13 |
| Восточная Африка | Б | М60, М181, SRY10831.1, М150, М109, М112 |
| Азия, Африка | ИЗ | М1, ЯП, М174, М40, М96, М75, М98 |
| Восточная Азия, Непал | Д | М174 |
| Западная Африка (первое событие вымирания?) | Е1а | М33 |
| Восточная Африка (первое событие вымирания — это раскол между E1b1 и E1a, второе событие вымирания — это раскол между E1b1b и E1b1a) | E1b1 | П2, М2, У175, М191 |
| Средний Восток | J1 | P58 |
мтДНК
[ редактировать ]| Источник | Гаплогруппа |
| Европа | H1 |
| Европа | H11a |
| Европа | H1a |
| Европа | H1b |
| Европа | H2a |
| Европа | Н3 |
| Европа | H5a |
| Европа | H6a |
| Европа | H7 |
| Европа | HV0/HV0a/V |
| Европа | я4 |
| Европа | J1c7 |
| Европа | J2b1 |
| Европа | Т2б* |
| Европа | Т2б4 |
| Европа | Т2е |
| Европа | U4c1 |
| Европа | U5* |
| Европа | U5a |
| Европа | U5a1b1 |
| Европа | U5b* |
| Европа | U5b1b* |
| Европа | U5b1c |
| Европа | U5b3 |
| Европа | X2c'e |
| Средний Восток | я |
| Средний Восток | А |
| Средний Восток | Б |
| Средний Восток | К/З |
| Средний Восток | Д/Г/М9/Е |
| Индия | Ф |
| Средний Восток | ЧАС* |
| Средний Восток | H13a1 |
| Средний Восток | H14a |
| Средний Восток | H20 |
| Средний Восток | H2a1 |
| Средний Восток | Н4 |
| Средний Восток | H6b |
| Средний Восток | Н8 |
| Средний Восток | HV1 |
| Средний Восток | я1 |
| Средний Восток | J / J1c / J2 |
| Средний Восток | J1a'b'e |
| Средний Восток | J1b1a1 |
| Средний Восток | J1b2a |
| Средний Восток | J1d / J2b |
| Средний Восток | J1d1 |
| Средний Восток | J2a |
| Средний Восток | J2a2a1 |
| Средний Восток | К* |
| Средний Восток | К1а* |
| Средний Восток | К1б1* |
| Средний Восток | Н1а* |
| Средний Восток | N1b |
| Средний Восток | N1c |
| Средний Восток | Н2 |
| Средний Восток | N9 |
| Средний Восток | Р* |
| Средний Восток | R0a |
| Средний Восток | Т |
| Средний Восток | Т1* |
| Западная Азия | Т1а |
| Средний Восток | Т2 |
| Средний Восток | Т2с |
| Средний Восток | Т2и |
| Средний Восток | U1* |
| Средний Восток | U2* |
| Средний Восток | U2e |
| Евразия | U3* |
| Средний Восток | U4 |
| Средний Восток | U4a* |
| Средний Восток | U7 |
| Средний Восток | U8* |
| Средний Восток | U9a |
| Средний Восток | Х |
| Средний Восток | Х1а |
| Средний Восток | X2b1 |
| Северная Африка | Л3е5 |
| Северная Африка | М1 |
| Северная Африка | М1а1 |
| Северная Африка | U6a |
| Северная Африка | U6a1'2'3 |
| Северная Африка | U6b'c'd |
| Восточная Африка | Л0* |
| Восточная Африка | L0a1 |
| Восточная Африка | L0a1b |
| Восточная Африка | L0a2* |
| Восточная Африка | L3c/L4/М |
| Восточная Африка | L3d1a1 |
| Восточная Африка | L3d1d |
| Восточная Африка | L3e1* |
| Восточная Африка | L3f* |
| Восточная Африка | L3h1b* |
| Восточная Африка | L3i* |
| Восточная Африка | L3x* |
| Восточная Африка | L4a'b* |
| Восточная Африка | Л5* |
| Восточная Африка | Л6 |
| Восточная Африка | Н* / М* / L3* |
| Западная Африка | Л1б* |
| Западная Африка | Л1б3 |
| Западная Африка | L1c* |
| Западная Африка | L1c2 |
| Западная Африка | Л2* |
| Западная Африка | Л2а |
| Западная Африка | L2a1* |
| Западная Африка | Л2а1а2'3'4 |
| Западная Африка | L2a1b |
| Западная Африка | L2a1b'f |
| Западная Африка | L2a1c1'2 |
| Западная Африка | L2a1(16189) |
| Западная Африка | Л2а2 |
| Западная Африка | Л2б* |
| Западная Африка | L2c1'2 |
| Западная Африка | Л2д |
| Западная Африка | L2e |
| Западная Африка | L3b |
| Западная Африка | L3b1a3 |
| Западная Африка | L3b(16124!) |
| Западная Африка | L3b2a |
| Западная Африка | L3d* |
| Западная Африка | L3e2'3'4 |
| Западная Африка | L3f1b* |
См. также
[ редактировать ]- Международный проект HapMap
- Молекулярная эволюция
- Молекулярная филогенетика
- Эволюционная генетика человека
- Раса (биология) / Раса (категоризация человека)
- Генетическая генеалогия
- Генеалогический ДНК-тест
- Список тем генетической генеалогии
- Список гаплогрупп известных людей
Ссылки
[ редактировать ]- ^ К. Барри Кокс, Питер Д. Мур, Ричард Лэдл. Уайли-Блэквелл, 2016. ISBN 978-1-118-96858-1 стр. 106. Биогеография: экологический и эволюционный подход.
- ^ Редакционная коллегия V&S Publishers, 2012, ISBN 9381588643 с. 137. Краткий научный словарь .
- ^ Международного общества генетической генеалогии, 2015 г. Генетический словарь
- ^ Консорциум, Y-хромосома (1 февраля 2002 г.). «Система номенклатуры дерева бинарных гаплогрупп Y-хромосомы человека» . Геномные исследования . 12 (2). Лаборатория Колд-Спринг-Харбор: 339–348. дои : 10.1101/гр.217602 . ISSN 1088-9051 . ПМК 155271 . ПМИД 11827954 .
- ^ «Консорциум Y-хромосомы» . Архивировано из оригинала 16 января 2017 г. Проверено 27 июля 2005 г.
- ^ Позник, Г. Дэвид; Сюэ, Яли; Мендес, Фернандо Л.; и др. (2016). «Периодические всплески демографии мужского пола, выявленные на основе 1244 последовательностей Y-хромосомы по всему миру» . Природная генетика . 48 (6): 593–599. дои : 10.1038/ng.3559 . ПМЦ 4884158 . ПМИД 27111036 .
- ^ Перейти обратно: а б Кармин, Моника; Сааг, Лаури; Висенте, Марио; и др. (2015). «Недавнее ограничение разнообразия Y-хромосом совпало с глобальными изменениями в культуре» . Геномные исследования . 25 (4): 459–466. дои : 10.1101/гр.186684.114 . ПМЦ 4381518 . ПМИД 25770088 .
- ^ Роотси С, Магри С, Кивисилд Т, Бенуцци Г, Хелп Х, Бермишева М, Кутуев И, Барак Л, Перичич М, Балановский О, Пшеничнов А, Дион Д, Гробей М, Животовский Л.А., Батталья В, Ахилли А, Ал- Захери Н, Парик Дж, Кинг Р, Чинниоглу Дж, Хуснутдинова Е, Рудан П, Балановска Е, Шеффран В, Симонеску М, Брем А, Гонсалвес Р, Роза А, Мойсан Дж. П., Чавентр А, Ферак В, Фуреди С, Оефнер П. Дж. , Шен П., Бекман Л., Микереси И., Терзич Р., Приморак Д., Камбон-Томсен А., Крумина А., Торрони А., Андерхилл П.А., Сантакьяра-Бенересетти А.С., Виллемс Р., Семино О (июль 2004 г.). «Филогеография гаплогруппы I Y-хромосомы выявляет отдельные области доисторического потока генов в Европе» (PDF) . Американский журнал генетики человека . 75 (1): 128–37. дои : 10.1086/422196 . ПМК 1181996 . ПМИД 15162323 . Архивировано из оригинала (PDF) 24 июня 2009 г. Проверено 8 марта 2007 г.
- ^ «PhyloTree.org» .
- ^ Лоогвяли Е.Л., Роосталу У., Малярчук Б.А., Деренко М.В., Кивисилд Т., Мецпалу Е. и др. (2004). «Разъединяющее единообразие: пестрая кладистическая картина гаплогруппы H мтДНК в Евразии» . Мол. Биол. Эвол . 21 (11): 2012–21. дои : 10.1093/molbev/msh209 . ПМИД 15254257 .
- ^ Толк Х.В., Барак Л., Перичич М., Кларик И.М., Яничичевич Б., Кэмпбелл Х., Рудан И., Кивисилд Т., Виллемс Р., Рудан П. (сентябрь 2001 г.). «Свидетельства наличия гаплогруппы F мтДНК у европейского населения и их этноисторические последствия» . Европейский журнал генетики человека . 9 (9): 717–23. дои : 10.1038/sj.ejhg.5200709 . ПМИД 11571562 .
- ^ Бекада А., Фрегель Р., Кабрера В.М., Ларруга Х.М., Пестано Х., Бенхамамуш С., Гонсалес А.М. (2013). «Введение алжирской митохондриальной ДНК и профилей Y-хромосомы в ландшафт Северной Африки» . ПЛОС ОДИН . 8 (2): e56775. Бибкод : 2013PLoSO...856775B . дои : 10.1371/journal.pone.0056775 . ПМЦ 3576335 . ПМИД 23431392 .
- ^ «Общие генетические предки жили примерно в один и тот же период времени» . 1 августа 2013 г. Проверено 23 января 2015 г.
- ^ Рагхаван М, Скоглунд П, Граф КЕ, Метспалу М, Альбрехцен А, Мольтке И, Расмуссен С, Стаффорд Т.В., Орландо Л, Метспалу Е, Кармин М, Тамбетс К, Роотси С, Мяги Р, Кампос П.Ф., Балановска Е, Балановский О , Хуснутдинова Е, Литвинов С, Осипова ЛП, Федорова С.А., Воевода М.И., ДеДжорджио М, Зихеритц-Понтен Т, Брунак С, Демещенко С, Кивисилд Т, Виллемс Р, Нильсен Р, Якобссон М, Виллерслев Е (январь 2014 г.). «Геном Сибири верхнего палеолита свидетельствует о двойном происхождении коренных американцев» . Природа . 505 (7481): 87–91. Бибкод : 2014Natur.505...87R . дои : 10.1038/nature12736 . ПМК 4105016 . ПМИД 24256729 .
- ^ Перейти обратно: а б с Карафет Т.М., Мендес Ф.Л., Судойо Х., Лансинг Дж.С., Хаммер М.Ф. (март 2015 г.). «Улучшенное филогенетическое разрешение и быстрая диверсификация гаплогруппы Y-хромосомы K-M526 в Юго-Восточной Азии» . Европейский журнал генетики человека . 23 (3): 369–73. дои : 10.1038/ejhg.2014.106 . ПМЦ 4326703 . ПМИД 24896152 .
Внешние ссылки
[ редактировать ]Общий
[ редактировать ]все гаплогруппы ДНК
[ редактировать ]Y-хромосома
- Карты гаплогрупп мира (PDF)
Гаплогруппы ДНК Y-хромосомы
[ редактировать ]- Консорциум Y-хромосомы
- Дерево гаплогрупп ISOGG Y-ДНК
- Y-дерево PhyloTree. Минимальная эталонная филогения Y-хромосомы человека.
- Предсказатель гаплогруппы
- Консорциум Y-хромосомы (2002), Номенклатурная система для дерева бинарных гаплогрупп Y-хромосомы человека , Genome Research, Vol. 12 (2), 339–48, февраль 2002 г. (Подробная иерархическая диаграмма содержит преобразования из предыдущих схем именования)
- Семино и др. (2000), Генетическое наследие палеолита Homo sapiens sapiens in Extant Europeans , Science, Vol 290 (статья, в которой представлены гаплогруппы «Eu»).
- Y-ДНК Этнографический и генографический атлас и сборник данных из открытых источников
Гаплогруппы митохондриальной ДНК
[ редактировать ]- PhyloTree - Филогенетическое древо глобальных вариаций митохондриальной ДНК человека.
- PhyloD3 - филогенетическое дерево на основе D3.js на основе PhyloTree.
- MitoTool - веб-сервер для анализа и поиска вариаций последовательностей митохондриальной ДНК человека. Архивировано 19 июня 2016 г. на Wayback Machine.
- HaploGrep - автоматическая классификация гаплогрупп митохондриальной ДНК на основе PhyloTree. Архивировано 12 июня 2016 г. на Wayback Machine.
- HaploFind - быстрый автоматический конвейер назначения гаплогрупп для митохондриальной ДНК человека. Архивировано 11 июня 2016 г. в Wayback Machine.
- Графический скелет гаплогруппы мтДНК
- Создание африканского ландшафта мтДНК
- Эволюционируют ли четыре клады гаплогруппы L2 мтДНК с разной скоростью?