Гаплогруппа J-M172
Эта статья , возможно, содержит оригинальные исследования . ( Апрель 2014 г. ) |
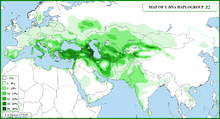
В генетике человека гаплогруппа J-M172 или J2. [ Филогенетика 1 ] представляет собой Y-хромосомы гаплогруппу , которая является субкладом (ветвью) гаплогруппы J-M304 . [ Филогенетика 2 ] Гаплогруппа J-M172 распространена у современных популяций Западной Азии , Центральной Азии , Южной Азии , Южной Европы , Северо-Западного Ирана и Северной Африки . Считается, что J-M172 мог возникнуть на Кавказе , в Анатолии и/или Западном Иране . [ 9 ] [ 10 ]
Далее он делится на две взаимодополняющие клады: J-M410 и J-M12 (M12, M102, M221, M314).
Происхождение
[ редактировать ]Дата происхождения гаплогруппы J-M172 была оценена Батини и др. в 2015 году как от 19 000 до 24 000 лет назад (BP). [ 11 ] Самино и др. в 2004 году датировали происхождение родительской гаплогруппы J-P209 периодом от 18 900 до 44 500 лет назад. [ 12 ] Древний J-M410 , а именно субклад J-Y12379*, был найден в мезолитическом контексте в зубе из пещеры Котиас Клде в западной Грузии, датируемом 9.529-9.895 кал. БП. [ 13 ] Этот образец был отнесен к аутосомному компоненту Кавказских охотников-собирателей (CHG). [ 14 ] J-M410, а точнее его субклад J-PF5008 , также был обнаружен в мезолитическом образце из пещер Хоту и Камарбанд , расположенных в провинции Мазандаран в Иране , датируемом 9100–8600 годами до нашей эры (приблизительно 11 000 лет назад). [ 15 ] Оба образца принадлежат триалетской культуре. Вполне вероятно, что люди J2 расселились по большей части Анатолии, Южного Кавказа и гор Загроса к концу Последнего оледенения 12 000 лет назад. [ 16 ]
Заллуа и Уэллс 2004 и аль-Захери 2003 утверждали, что обнаружили самую раннюю известную миграцию J2, распространившуюся, возможно, из Анатолии и Кавказа . [ 9 ] [ 10 ] [ 17 ] В 2001 году Небель и др. обнаружили, что «согласно Андерхиллу и др. (2000), Eu 9 (H58) эволюционировал из Eu 10 (H71) посредством трансверсии T→G на M172 (выделено автором)», и что в сегодняшних популяциях Eu 9 (H71) постмутационная форма M172) наиболее сильна на Кавказе , в Малой Азии и Леванте , в то время как Eu 10 становится сильнее и заменяет частоту Eu 9 по мере продвижения на юг, на Аравийский полуостров . [ 18 ] так что люди с Кавказа встретились с арабами вблизи и между Месопотамией (Шумер/Ассирия) и пустыней Негев , поскольку « арабизация » распространилась из Аравии на Плодородный полумесяц и Турцию.
Согласно исследованию Ди Джакомо, проведенному в 2004 году , гаплогруппа J-M172 распространилась в Южную Европу либо из Леванта , либо из Анатолии , вероятно, параллельно с развитием сельского хозяйства. [ 19 ] Что касается времени его распространения в Европе, Ди Джакомо указывает на события, произошедшие после неолита, в частности на демографический расцвет, связанный с возникновением древнегреческого мира. Семино и др. получил более старые оценки возраста для всего J2 (использовав метод Животовского, см. Ди Джакомо), постулируя его первоначальное распространение среди неолитических фермеров с Ближнего Востока. Однако его распространение по субкладам, демонстрирующее локализованные пики на Южных Балканах, в южной Италии, на севере/центральной Италии и на Кавказе, не соответствует единому сценарию «волны наступления», выдавая ряд все еще плохо изученных постнеолитических процессы, которые создали его нынешний образец. предположил, что, как и Ди Джакомо, южные Балканы бронзового века Семино в 2004 году были важным вектором распространения. [ 12 ]
Распределение
[ редактировать ]Гаплогруппа J-M172 встречается в основном на территории Плодородного полумесяца , Кавказа ( Насидзе 2003 ), Анатолии , Италии , средиземноморского побережья и Иранского нагорья ( Семино 2004 ). Y-ДНК: J2 (J-M172): Сирид/Нахрейнид Арабид(ы).
Самая высокая зарегистрированная частота J-M172 за всю историю составила 87,4% среди ингушей в Малгобеке ( Балановский, 2011 ).
В частности, он встречается в Ираке ( Аль-Захери 2003 ), Кувейте , [ 20 ] Сирия ( Луис 2004 ), Ливан ( Заллуа 2008l ), Турция ( Чинниоглу 2004 ), Грузия ( Насидзе 2003 ), Азербайджан ( Ди Джакомо 2004 ), Северный Кавказ ( Насидзе 2004 ), Армения ( Уэллс 2001 ), Иран ( Насидзе 2004 ), Израиль ( Семино 2004 ), Палестина ( Семино 2004 ), Кипр ( Капелли 2005 ), Греция ( Мартинес 2007 ), Албания ( Семино 2000 ), Италия ( Капелли 2007 ), Испания ( Ди Джакомо 2003 ), и чаще у иракцев 24% ( Аль-Захери 2011 ), чеченцы 51,0%-58,0% ( Балановский 2011 ), грузины 21% ( Уэллс 2001 )-72% ( Уэллс 2001 ), ливанцы 30% ( Семино 2004 ), осетины 24% ( Насидзе 2004 ), балкарцы 24% ( Батталья 2008 ), сирийцы 23% ( Луис 2004 ), турки 13% ( Чинниоглу 2004 )-40% ( Семино 2000 ), киприоты 12,9% ( Эль-Сибай 2009 )-37% ( Капелли 2005 ), армяне 21% ( Уэллс 2001 ) - 24% ( Насидзе 2004 ), черкесы 21,8% ( Балановский 2011 ), иранцы 10% ( Насидзе 2004 ) -25% ( Уэллс 2001 ), албанцы 16% ( Батталья 2008 ) и ( Семино 2000 ), итальянцы 9 %-36% ( Капелли 2007 ), евреи-сефарды 15% ( Небель 2001 )-29% ( Семино 2004 ), мальтийцы 21% ( Капелли 2005 ), палестинцы 17% ( Семино 2004 ), саудовцы 14% ( Абу-Амеро 2009 ), иорданцы 14%, оманцы 10-15% ( Ди Джакомо 2004 ) и ( Луис 2004 Северной Индии ) и мусульмане-шииты 18% ( Иасвархант 2009 ).
Северная Африка
[ редактировать ]Гаплогруппа J2 с низкой частотой встречается в Северной Африке. с точкой доступа в регионе Сус Фадлауи-Зид 2014 г. большинство образцов Суса имеют те же гаплотипы , что и гаплогруппа J-L271, обнаруженная в Мсакене .
| Страна/регион | Выборка | Н | J-M172 | Изучать |
| Тунис | Тунис | 62 | 8 | Эль-Сибай 2009 г. |
| Тунис | Сус | 220 | 8.2 | Фадхлауи-Зид 2014 г. |
| Алжир | Оран | 102 | 4.9 | Робин, 2008 год. |
| Египет | 124 | 7.6 | Эль-Сибай 2009 г. | |
| Египет | 147 | 12.0 | Абу-Амеро 2009 г. | |
| Марокко | 221 | 4.1 | Фрегель 2009 | |
| Северная Африка | Алжир, Тунис | 202 | 3.5 | Фрегель 2009 |
Центральная Азия
[ редактировать ]| Страна/регион | Выборка | Н | J-M172 | Изучать |
| Синьцзян | Лоп-уйгуры | 64 | 57.8 | Лю 2018 |
| Синьцзян | уйгуры | 50 | 34 | Шоу 2010 г. |
| Таджикистан | Ягнобис | 31 | 32 | Уэллс 2001 г. |
| Dushanbe | таджики | 16 | 31 | Уэллс 2001 г. |
| Синьцзян | Узбеки | 23 | 30.4 | Шоу 2010 г. |
| Афганистан | Хазарейский | 60 | 26.6 | Хабер 2012 г. |
| Синьцзян | Уйгуры | 39 | 25.6 | Лю 2018 |
| Казахстан | уйгуры | 41 | 20 | Уэллс 2001 г. |
| Самарканд | таджики | 40 | 20 | Уэллс 2001 г. |
| Таджикистан | таджики | 38 | 18.4 | Уэллс 2001 г. |
| Туркменистан | туркмены | 30 | 17 | Уэллс 2001 г. |
| Синьцзян | Памирские таджики | 31 | 16.1 | Шоу 2010 г. |
| Афганистан | Узбеки | 126 | 16 | Кристофаро, 2013 г. |
| Бухара | Узбеки | 58 | 16 | Уэллс 2001 г. |
| Самарканд | Узбеки | 45 | 16 | Уэллс 2001 г. |
| Сурхандарьинская | Узбеки | 68 | 16 | Уэллс 2001 г. |
| Узбекистан | Узбеки | 366 | 13.4 | Уэллс 2001 г. |
| Казахстан | казахи | 30 | 13.3 | Карафет 2001 |
| Турфанский район | уйгуры | 143 | 9.8 | [ нужна ссылка ] |
| Горячая зона | уйгуры | 478 | 9.2 | [ нужна ссылка ] |
| Чанцзи | Хуэй | 175 | 9.1 | [ нужна ссылка ] |
| Синьцзян | Долан Уйгуры | 76 | 7.9 | Лю 2018 |
| Нинся | Хуэй | 65 | 7.7 | [ нужна ссылка ] |
| Кызылсу | киргизский | 241 | 6.64% | Го 2020 |
| Казахстан | казахи | 1294 | 4.33% | Аширбеков 2017 |
| Кыргызстан | киргизский | 132 | 3.79% | Кристофаро, 2013 г. |
J-M172 встречается с умеренной частотой среди народов Центральной Азии, таких как уйгуры , узбеки , туркмены , таджики , казахи и ягнобцы . Согласно генетическому исследованию Shou et al., проведенному в Северо-Западном Китае. (2010), заметная высокая частота J-M172 наблюдается, особенно у уйгуров 34% и узбеков 30,4% в Синьцзяне , Китай . Лю Шуху и др. (2018) обнаружили J2a1 (L26/Page55/PF5110/S57, L27/PF5111/S396) у 43,75% (28/64) и J2a2 (L581/S398) у 14,06% (9/64) выборки лоп-уйгуров из Деревня Карчуга уезда Ютянь (Лопнур), Синьцзян, J2a1b1 (M92, M260/Page14) в 25,64% (10/39) выборки керианских уйгуров /Page55/PF5110/S57, L27/PF5111/S396) в 3,95%; (3/76) и J2a2 (L581/S398) в 3,95% (3/76) выборки доланских уйгуров из поселка Хорикол уезда Ават Синьцзяна. [ 21 ] Только этнические меньшинства крайнего северо-запада имели гаплогруппу J в Синьцзяне, Китай. У узбеков в выборке было 30,4% J2-M172, он также был у таджиков Синьцзяна и уйгуров. [ 22 ]
Гаплогруппа имеет древнее присутствие в Центральной Азии и, по-видимому, предшествовала распространению ислама ( Шоу 2010 ). Кроме того, непосредственный предок J-M172, а именно J* (J-M304*, он же J-P209*, J-12f2.1*), встречается также у сибо , казахов , дунсянов и узбеков Северо-Западного Китая.
В 2015 году два древних образца, принадлежащих J-M172 или J-M410 (J2a), были найдены на двух разных археологических объектах на Алтае , восточная Россия : Кытманово и курган Сары-бель. Оба древних образца относятся к культурам железного века Алтая. Сары-бел J2/J2a датируется 50 г. до н.э., тогда как Кытмановский образец датируется 721-889 гг. н.э. Анализ генетических примесей этих образцов также предполагает, что эти люди были более тесно связаны с западноевразийцами , чем с другими алтайцами того же периода, хотя они также, по-видимому, связаны с современными тюркскими народами региона. [ 23 ] [ 24 ] [ 25 ]
Европа
[ редактировать ]| Страна/регион | Выборка | Н | J-M172 | Изучать |
| Албания | 55 | 19.9% 11/55 |
Битва 2009 | |
| Босния и Герцеговина | сербы | 81 | 8.7 | Битва 2009 |
| Кипр | 164 | 12.9 | Эль-Сибай 2009 г. | |
| Греция | Крит | 143 | 35 | Эль-Сибай 2009 г. |
| Иберия | 655 | 7 | Фрегель 2009 | |
| Иберия | 1140 | 7.7 | Адамс 2008 г. | |
| Италия | Сицилия | 212 | 22.6 | Эль-Сибай 2009 г. |
| Италия | Материк | 699 | 20 | Волосы 2007 |
| Италия | Центральный Марке | 59 | 35.6 | Волосы 2007 |
| Италия | Западная Калабрия | 57 | 35.1 | Волосы 2007 |
| Италия | Валь Бадия | 34 | 8.8 | Волосы 2007 |
| Мальта | 90 | 21.1 | Эль-Сибай 2009 г. | |
| Португалия | Север, Центр, Юг | 303 | 6.9 | Эль-Сибай 2009 г. |
| Португалия | Трас-ос-Монтес (евреи) | 57 | 24.5 | Ногейро 2010 г. |
| Сардиния | 81 | 9.9 | Эль-Сибай 2009 г. | |
| Испания | Майорка | 62 | 8.1 | Эль-Сибай 2009 г. |
| Испания | Севилья | 155 | 7.8 | Эль-Сибай 2009 г. |
| Испания | Леон | 60 | 5 | Эль-Сибай 2009 г. |
| Испания | Ибица | 54 | 3.7 | Эль-Сибай 2009 г. |
| Испания | Кантабрия | 70 | 2.9 | Эль-Сибай 2009 г. |
| Испания | Галисия | 292 | 13 | [ нужна ссылка ] |
| Испания | Канарские острова | 652 | 10.5 | Фрегель 2009 |
В Европе частота гаплогруппы J-M172 падает по мере удаления на север от Средиземноморья . В Италии J-M172 встречается с региональной частотой от 9% до 36% ( Капелли, 2007 ). В Греции он встречается с региональной частотой от 10% до 48%. Согласно недавнему исследованию ( Cinnioglu 2004 ), около 24% турецких мужчин имеют J-M172, а региональная частота варьируется от 13% до 40% ( Semino 2000 ). В сочетании с J-M267 до половины населения Турции принадлежит к гаплогруппе J-P209 .
Было высказано предположение, что субклад гаплогруппы J-M410 был связан с населением на древнем Крите путем изучения отношений между анатолийским , критским и греческим населением из мест раннего неолита на Крите. Гаплогруппа J-M172 была связана с неолитической Грецией (ок. 8500–4300 гг. до н.э.) и, как сообщается, была обнаружена на современном Крите (3,1%) и материковой Греции ( Македония 7,0%, Фессалия 8,8%, Арголида 1,8%) ( King 2008 ). .
Северный Кавказ
[ редактировать ]| Страна/регион | Выборка | Н | J-M172 | Изучать |
| Кавказ | Абхазский | 58 | 13.8 | Балановский 2011 г. |
| Кавказ | Просторный | 115 | 6 | Балановский 2011 г. |
| Кавказ | Чеченский | 330 | 57 | Балановский 2011 г. |
| Кавказ | Адыгейский | 142 | 21.8 | Балановский 2011 г. |
| Кавказ | Даргинцы | 101 | 1 | Балановский 2011 г. |
| Кавказ | Ингушский | 143 | 88.8 | Балановский 2011 г. |
| Кавказ | Кайтак | 33 | 3 | Балановский 2011 г. |
| Кавказ | Для резины | 73 | 21 | Юнусбаев 2012 г. |
| Кавказ | Кубачи | 65 | 0 | Балановский 2011 г. |
| Кавказ | Лезгины | 81 | 2.5 | Балановский 2011 г. |
| Кавказ | маленькие медведи | 357 | 16 | Балановский 2011 г. |
| Кавказ | Шапсуг | 100 | 6 | Балановский 2011 г. |
| Кавказ | 1525 | 28.1 | Балановский 2011 г. |
J-M172 встречается с очень высокой частотой у некоторых народов Кавказа 21% -72 : среди ингушей 87,4% ( Балановский 2011 ), чеченцев 55,2% ( Балановский 2011 ), грузин %, ( Wells 2001 ), азербайджанцев 24% ( Ди Джакомо 2004 ) - 48%, ( Уэллс 2001 ) Абхазы 25%, ( Насидзе 2004 ) Балкарцы 24% ( Батталья 2008 ), Осетины 24% ( Насидзе 2004 ), Армяне 21% ( Уэллс 2001 ) -24% ( Насидзе 2004 ) , адыгейцы 21,8% ( Балановский 2011 ) и другие группы ( Насидзе 2004 и Насидзе 2003 ).
Западная Азия
[ редактировать ]| Страна/регион | Выборка | Н | J-M172 | Изучать |
| еврейский | Ашкенази еврейские | 442 | 19 | Бехар 2004 г. |
| Иран | 92 | 25 | Эль-Сибай 2009 г. | |
| Ирак | 154 | 24 | Аль-Захери 2011 г. [ 26 ] | |
| палестинский араб | Как | 101 | 18.6 | Эль-Сибай 2009 г. |
| Иордания | 273 | 14.6 | Эль-Сибай 2009 г. | |
| Ливан | 951 | 29.4 | Эль-Сибай 2009 г. | |
| Мой собственный | 121 | 10.0 | Абу-Амеро 2009 г. | |
| Катар | 72 | 8.3 | Эль-Сибай 2009 г. | |
| Саудовская Аравия | 157 | 14 | Абу-Амеро 2009 г. [ 27 ] | |
| Сирия | Сирия | 554 | 20.8 | Эль-Сибай 2009 г. |
| Турция | 523 | 24.2 | Эль-Сибай 2009 г. | |
| ОАЭ | 164 | 10.3 | Эль-Сибай 2009 г. | |
| Йемен | 62 | 9.6 | Эль-Сибай 2009 г. |
У евреев-сефардов около 15% ( Небель 2001 )-29% ( Семино 2004 ) гаплогруппы J-M172, а у евреев-ашкенази 15% ( Шен 2004 )-23% ( Семино 2004 ). ), сообщалось В раннем исследовании, в котором тестировались только четыре маркера STR ( Maspina 2001 , что небольшая выборка итальянских Коэнов принадлежала к Сети 1.2, раннему обозначению всей клады, теперь известной как J-L26, определяемой делецией в DYS413. Однако большое количество всех евреев Коэнов в мире принадлежат к гаплогруппе J-M267 (см. модальный гаплотип Коэна ).
Было показано, что гаплогруппа J-M172 имеет более северное распространение на Ближнем Востоке, хотя она существует в значительных количествах в южных регионах Ближнего Востока, ее меньшее количество было обнаружено по сравнению с ее братской гаплогруппой J-M267. который имеет высокочастотное южное распространение. Считалось, что исходная популяция J-M172 произошла из Леванта/Сирии (Syrid-J-M172), и что ее появление среди современных популяций Европы, Центральной Азии и Южной Азии было признаком неолитических земледельцев. Однако, как уже говорилось, сейчас считается, что оно, скорее всего, распространялось волнообразно в результате постнеолитических процессов.
Южная Азия
[ редактировать ]Гаплогруппа J2 присутствовала в Южной Азии в основном как J2a-M410 и J2b-M102 со времен неолита (9500 лет назад). [ 28 ] [ 29 ] Было обнаружено, что J2-M172 значительно выше среди дравидийских каст (19%), чем среди индоарийских каст (11%). J2-M172 и J-M410 встречаются в 21% среди дравидских средних каст, за ними следуют высшие касты (18,6%) и низшие касты (14%). [ 30 ] Среди кастовых групп самая высокая частота J2-M172 наблюдалась среди тамилов Веллаларов Южной Индии - 38,7%. [ 30 ] J2 также присутствует у индийских племен и встречается в 11% у австро-азиатских племен. Среди австро-азиатских племен преобладает J2 у племени Асур (77,5%), хотя и с размером выборки 40 человек. [ 28 ] и в Лодхе (35%) Западной Бенгалии . [ 30 ] J2 также присутствует у южноиндийского горного племени Тода (38,46%), хотя размер выборки составляет всего 26 человек. [ 31 ] в племени Анд Телангана - 35,19%, [ 32 ] в племени Нарикуравар - 57,9% [ 28 ] и в племени Кол в Уттар- Прадеше - 33,34%. [ 33 ] Гаплогруппа J-P209 оказалась более распространенной у индийских мусульман-шиитов , из которых 28,7% принадлежат к гаплогруппе J, 13,7% - к J-M410, 10,6% - к J-M267 и 4,4% - к J2b ( Eaaswarkhoth 2009 ).
В Пакистане частоты J2-M172 наблюдались среди парсов - 38,89%, дравидоязычных самые высокие брахуи - 28,18% и макранских белуджей - 24%. [ 34 ] Это также встречается на уровне 18,18% в Макрани-Сиддис и на 3% в Карнатака-Сиддис . [ 34 ] [ 35 ]
J2-M172 встречается с общей частотой 16,1% у жителей Шри-Ланки . [ 36 ] На Мальдивах 22% населения Мальдив оказались положительными по гаплогруппе J2. [ 37 ] Субклады M172, такие как M67 и M92, не были обнаружены ни в индийских, ни в пакистанских образцах, что также может указывать на частичное общее происхождение. [ 30 ]
J2-M172 наблюдался в 15,9% (20/164 J2a-M410, 6/164 J2b2-M241) Тару из Уттар-Прадеша , [ 38 ] 13,4% (19/202 J2a-M410, 8/202 J2b2-M241) Тару из Непала , [ 39 ] [ 38 ] и 8,9% (4/45 J2a-M410) Тхару из Уттаракханда . [ 38 ]
Распределение субкладов
[ редактировать ]Гаплогруппа J-M172 подразделяется на две дополняющие друг друга подгаплогруппы: J-M410 , определяемая M410 генетическим маркером , и J-M12 , определяемая генетическим маркером M12 .
J-M172
[ редактировать ]J-M172 типичен для популяций Ближнего Востока , Южной Европы , Юго-Западной Азии и Кавказа , с умеренным распространением на большей части территории Центральной Азии , Южной Азии и Северной Африки . [ 40 ]
J-M410
[ редактировать ]J-M410* встречается в Грузии , Северной Осетии. [ 41 ]
J-M47
[ редактировать ]J-M47 встречается с низкой частотой в Грузии , ( Батталья, 2008 ), южном Иране ( Регейро, 2006 ), Катаре ( Каденас, 2008 ) , Саудовской Аравии ( Абу-Амеро, 2009 ), Сирии ( Ди Джакомо, 2004 ), Тунисе ( Арреди, 2004 ), Турции. ( Ди Джакомо, 2004 г. и Чинниоглу, 2004 г. ), ОАЭ ( Каденас, 2008 г. ) и Центральная Азия / Сибирь ( Андерхилл, 2000 г. ).
J-M67
[ редактировать ]J-M67 (в старых статьях называемый J2f) имеет самые высокие частоты, связанные с нахскими народами . Встречается с очень высокой (большинством) частотой среди ингушей в Малгобеке (87,4%), чеченцев в Дагестане (58%), чеченцев в Чечне (56,8%) и чеченцев в Малгобеке, Ингушетия (50,9%) ( Балановский 2011 ). На Кавказе он встречается со значительной частотой среди грузин (13,3%) ( Семино 2004 ), железных осетин (11,3%), южнокавказских балкарцев (6,3%) ( Семино 2004 ), дигорских осетин (5,5%), абхазов (6,9). %) и черкесские (5,6%) ( Балановский 2011 ). Он также встречается с заметной частотой в Средиземноморье и на Ближнем Востоке, включая критян (10,2%), северо-центральных итальянцев (9,6%), южных итальянцев (4,2%; только 0,8% среди северных итальянцев), анатолийских турок (2,7- 5,4%), греки (4-4,3%), албанцы (3,6%), евреи-ашкенази (4,9%), сефарды (2,4%), каталонцы (3,9%), андалузцы (3,2%), калабрийцы (3,3%), албанцы Калабрийцы (8,9%) (см. Di Giacomo 2004 и Semino 2004 ).
J-M92/M260, субклад J-M67, наблюдался у 25,64% (10/39) выборки керианских уйгуров из деревни Дарья Бойи уезда Ютянь (Керия), Синьцзян. [ 21 ] Эта уйгурская деревня расположена в отдаленном оазисе в пустыне Такла-Макан .
J-M319
[ редактировать ]J-M319 встречается с низкой и умеренной частотой у критских греков ( Мартинес 2007 и Кинг 2008 ), иракских евреев ( Шен 2004 ) и марокканских евреев ( Шен 2004 ).
J-M158
[ редактировать ]J-M158 (расположение под L24 неизвестно) J-M158 встречается с низкой частотой в Турции ( Cinnioglu 2004 ), Южной Азии ( Sengupta 2006 и Underhill 2000 ), Индокитае ( Underhill 2000 ) и на Пиренейском полуострове .
Филогенетика
[ редактировать ]В филогенетике Y-хромосомы субклады являются ветвями гаплогрупп. Эти субклады также определяются однонуклеотидными полиморфизмами (SNP) или полиморфизмами уникальных событий (UEP).
Филогенетическая история
[ редактировать ]До 2002 года в научной литературе существовало как минимум семь систем наименования филогенетического дерева Y-хромосомы. Это привело к значительной путанице. В 2002 году основные исследовательские группы объединились и сформировали Консорциум Y-хромосомы (YCC). Они опубликовали совместный документ, в котором было создано единое новое дерево, которое все согласились использовать. Позже группа гражданских ученых, интересующихся популяционной генетикой и генетической генеалогией, сформировала рабочую группу для создания любительского дерева, стремясь быть прежде всего своевременным. В таблице ниже собраны все эти работы, относящиеся к знаковому дереву YCC 2002 года. Это позволяет исследователю, просматривающему старую опубликованную литературу, быстро перемещаться между номенклатурами.
| YCC 2002/2008 (сокращенно) | (а) | (б) | (с) | (г) | (е) | (г) | (или) | YCC 2002 (от руки) | YCC 2005 (от руки) | YCC 2008 (от руки) | YCC 2010r (от руки) | ИСОГГ 2006 г. | ИСОГГ 2007 | ИСОГГ 2008 | ИСОГГ 2009 | ИСОГГ 2010 | ИЗОГГ 2011 | ИЗОГГ 2012 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| J-12ф2а | 9 | МЫ | С | 23 | ЕС10 | Н4 | Б | Дж* | Дж | Дж | Дж | - | - | - | - | - | - | Дж |
| J-M62 | 9 | МЫ | С | 23 | ЕС10 | Н4 | Б | J1 | J1a | J1a | J1a | - | - | - | - | - | - | Частный |
| J-M172 | 9 | МЫ | С | 24 | I9 | Н4 | Б | J2* | J2 | J2 | J2 | - | - | - | - | - | - | J2 |
| J-M47 | 9 | МЫ | С | 24 | I9 | Н4 | Б | J2a | J2a | J2a1 | J2a4a | - | - | - | - | - | - | J2a1a |
| J-M68 | 9 | МЫ | С | 24 | I9 | Н4 | Б | J2b | J2b | J2a3 | J2a4c | - | - | - | - | - | - | J2a1c |
| J-M137 | 9 | МЫ | С | 24 | I9 | Н4 | Б | J2c | J2c | J2a4 | J2a4h2a1 | - | - | - | - | - | - | J2a1h2a1a |
| J-M158 | 9 | МЫ | С | 24 | I9 | Н4 | Б | J2d | J2d | J2a5 | J2a4h1 | - | - | - | - | - | - | J2a1h1 |
| J-M12 | 9 | МЫ | С | 24 | I9 | Н4 | Б | J2e* | J2e | J2b | J2b | - | - | - | - | - | - | J2b |
| J-M102 | 9 | МЫ | С | 24 | I9 | Н4 | Б | J2e1* | J2e1 | J2b | J2b | - | - | - | - | - | - | J2b |
| J-M99 | 9 | МЫ | С | 24 | I9 | Н4 | Б | J2e1a | J2e1a | J2b2a | J2b2a | - | - | - | - | - | - | Частный |
| J-M67 | 9 | МЫ | С | 24 | I9 | Н4 | Б | J2f* | J2f | J2a2 | J2a4b | - | - | - | - | - | - | J2a1b |
| J-M92 | 9 | МЫ | С | 24 | I9 | Н4 | Б | J2f1 | J2f1 | J2a2a | J2a4b1 | - | - | - | - | - | - | J2a1b1 |
| J-M163 | 9 | МЫ | С | 24 | I9 | Н4 | Б | J2f2 | J2f2 | J2a2b | J2a4b2 | - | - | - | - | - | - | Частный |
Научные публикации
[ редактировать ]Следующие исследовательские группы в соответствии с их публикациями были представлены в создании Дерева YCC.
Филогенетические деревья
[ редактировать ]Существует несколько подтвержденных и предложенных филогенетических деревьев гаплогруппы J-M172. Научно признанным является вариант Консорциума Y-хромосомы (YCC), опубликованный в Karafet 2008 и впоследствии обновленный. Черновик дерева, показывающего новые научные достижения, предоставлен Томасом Краном из Центра геномных исследований в Хьюстоне, штат Техас . Международное общество генетической генеалогии (ISOGG) также предоставляет любительское дерево. [ Филогенетика 3 ] [ 42 ]
Черновик дерева Центра геномных исследований
[ редактировать ]для гаплогруппы J-M172 Центра геномных исследований Это Томас Кран из проекта предлагаемого дерева ( Krahn & FTDNA 2013 ). Для краткости показаны только первые три уровня субкладов.
- М172, Л228
- М410, Л152, Л212, Л505, Л532, Л559
- ПФ5008
- Y182822
- Л581
- Z37823
- Л581
- Y182822
- ПФ4610
- Z6046
- Л26
- ПФ5008
- М12, М102, М221, М314, Л282
- М205
- М241
- М99
- М280
- М321
- Р84
- Л283
- М410, Л152, Л212, Л505, Л532, Л559
Дерево Консорциума Y-хромосомы
[ редактировать ]Это официальное научное дерево, созданное Консорциумом Y-хромосомы (YCC). Последнее крупное обновление было в 2008 году ( Карафет 2008 ). Последующие обновления были ежеквартальными и двухгодичными. Текущая версия является версией обновления 2010 года. [ 43 ]
Этот раздел нуждается в расширении . Вы можете помочь, добавив к нему . ( январь 2013 г. ) |
Дерево ISOGG
[ редактировать ]Ниже представлены субклады гаплогруппы J-M172 с определяющей мутацией согласно дереву ISOGG (по состоянию на январь 2020 г.). Обратите внимание, что идентификаторы на основе спуска могут быть изменены по мере обнаружения новых SNP, которые дополняют и уточняют дерево. Для краткости показаны только первые три уровня субкладов.
- J2 M172/Page28/PF4908, L228/PF4895/S321
- J2a M410, L152, L212/PF4988, L559/PF4986
- J2a1 DYS413≤18, L26/Page55/PF5110/S57, F4326/L27/PF5111/S396
- J2a1a М47, М322
- J2a1b M67/PF5137/S51
- J2a1c M68
- J2a1d M319
- J2a1e M339
- J2a1f M419
- J2a1g P81/PF4275
- J2a1h L24/S286, L207.1
- J2a1i L88.2, L198
- J2a2 L581/PF5026/S398
- J2a2a P279/PF5065
- J2a1 DYS413≤18, L26/Page55/PF5110/S57, F4326/L27/PF5111/S396
- J2b М12
- J2b1 М205
- J2b1a~ A11525, PH4306, Y22059, Y22060, Y22061, Y22062, Y22063
- J2b1b~ CTS1969
- J2b2~ CTS2622/Z1827, CTS11335/Z2440, Z575
- J2b2a М241
- J2b1 М205
- J2a M410, L152, L212/PF4988, L559/PF4986
См. также
[ редактировать ]Генетика
[ редактировать ]- Генетическая история Ближнего Востока
- Генетическая история Европы
- Таблица перевода гаплогрупп Y-хромосомы
- Генетическая генеалогия
- Гаплогруппа
- Гаплотип
- Гаплогруппа ДНК Y-хромосомы человека
- Молекулярная филогения
- Парагруппа
- Субклад
- Y-хромосома Аарон
- Гаплогруппы Y-хромосомы в популяциях мира
- Гаплогруппы Y-ДНК в населении Европы
- Гаплогруппы Y-ДНК у населения Южной Азии
- Гаплогруппы Y-ДНК в популяциях Восточной и Юго-Восточной Азии
- Гаплогруппы Y-ДНК у населения Ближнего Востока
- Гаплогруппы Y-ДНК у населения Северной Африки
- Гаплогруппы Y-ДНК у населения Кавказа
- Гаплогруппы Y-ДНК по этническим группам
Другие субклады J-ДНК
[ редактировать ]Основополагающее дерево Y-ДНК
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б «YFull YTree v7.05.00» . yfull.com . Архивировано из оригинала 18 июня 2019 г. Проверено 27 сентября 2019 г.
- ^ Степень дифференциации Hg J, наблюдаемая как с биаллельными, так и с микросателлитными маркерами, указывает на Ближний Восток как на его вероятную родину. В этой области J-M172 и J-M267 представлены в равной степени и демонстрируют наибольшую степень внутренней изменчивости, что указывает на то, что, скорее всего, эти субклады возникли также на Ближнем Востоке. ( Ди Джакомо 2004 )
- ^ Шоу, Вэй-Хуа; Цяо, Эн-Фа; Вэй, Чуан-Юй; Донг, Юн-Ли; Тан, Си-Цзе; Ши, Хун; Тан, Вэнь-Ру; Сяо, Чун-Цзе (2010). «Распределение Y-хромосомы среди популяций Северо-Западного Китая указывает на значительный вклад скотоводов Центральной Азии и меньшее влияние западных евразийцев» . Журнал генетики человека . 55 (5): 314–322. дои : 10.1038/jhg.2010.30 . ПМИД 20414255 .
- ^ Хабер М., Платт Д.Э., Ашрафиан Бонаб М., Юханна С.К., Сориа-Эрнанц Д.Ф., Мартинес-Круз Б. и др. (2012). «Этнические группы Афганистана имеют общее Y-хромосомное наследие, структурированное историческими событиями» . ПЛОС ОДИН . 7 (3): e34288. Бибкод : 2012PLoSO...734288H . дои : 10.1371/journal.pone.0034288 . ПМЦ 3314501 . ПМИД 22470552 .
- ^ Капелли С., Редхед Н., Романо В., Кали Ф., Лефранк Г., Делаг В. и др. (2006). «Структура населения в Средиземноморском бассейне: перспектива хромосомы AY» . Анналы генетики человека . 70 (2): 207–225. дои : 10.1111/j.1529-8817.2005.00224.x . hdl : 2108/37090 . ПМИД 16626331 . S2CID 25536759 .
- ^ «Гаплогруппа Y J в Иране, автор Альфред А. Абурто-младший» . Архивировано из оригинала 13 октября 2012 г. Проверено 6 апреля 2014 г.
- ^ Jump up to: а б с "Трофимова Наталья Вадимовна, ИЗМЕНЧИВОСТЬ МИТОХОНДРИАЛЬНОЙ ДНК И Y-ХРОМОСОМЫВ ПОПУЛЯЦИЯХ ВОЛГО-УРАЛЬСКОГО РЕГИОНА, 03.02.07" (PDF) . Archived from the original (PDF) on 2015-01-14 . Retrieved 2015-01-07 .
- ^ Опубликованное генетическое исследование особей калаша под руководством Фирасата (2007) выявило высокие и разнообразные частоты.
- ^ Jump up to: а б Zalloua & Wells: Журнал National Geographic, октябрь 2004 г. [1] Архивировано 7 апреля 2014 г. в Wayback Machine и [2] Архивировано 10 февраля 2012 г. в Wayback Machine .
- ^ Jump up to: а б «Н. Аль-Захери и др. «Полиморфизмы Y-хромосомы и мтДНК в Ираке, перекресток раннего расселения человека и постнеолитических миграций» (2003)» (PDF) . ДНК генеалогического древа. Архивировано из оригинала (PDF) 27 декабря 2010 года . Проверено 1 сентября 2013 г.
- ^ Батини С., Халласт П., Задик Д., Дельсер П.М., Бенаццо А., Гиротто С. и др. (май 2015 г.). «Крупномасштабное недавнее расширение европейских отцов, продемонстрированное изменением последовательности населения» . Природные коммуникации . 6 : 7152. Бибкод : 2015NatCo...6.7152B . дои : 10.1038/ncomms8152 . ПМЦ 4441248 . ПМИД 25988751 .
- ^ Jump up to: а б Семино О., Магри С., Бенуцци Г., Лин А.А., Аль-Захери Н., Батталья В. и др. (май 2004 г.). «Происхождение, распространение и дифференциация гаплогрупп E и J Y-хромосомы: выводы о неолитизации Европы и более поздних миграционных событиях в Средиземноморье» . Американский журнал генетики человека . 74 (5): 1023–34. дои : 10.1086/386295 . ПМК 1181965 . ПМИД 15069642 .
- ^ «[Домашняя страница]» . YПолный . Архивировано из оригинала 23 апреля 2020 г. Проверено 11 мая 2020 г.
- ^ Джонс Э.Р., Гонсалес-Фортес Г., Коннелл С., Сиска В., Эрикссон А., Мартиниано Р. и др. (ноябрь 2015 г.). «Геномы верхнего палеолита раскрывают глубокие корни современных евразийцев» . Природные коммуникации . 6 : 8912. Бибкод : 2015NatCo...6.8912J . дои : 10.1038/ncomms9912 . ПМК 4660371 . ПМИД 26567969 .
- ^ Лазаридис, Джозеф; Надель, Дэни; Роллефсон, Гэри; Мерретт, Дебора Дж.; Роланд, Надин; Маллик, Свапан; Фернандес, Даниэль; Новак, Марио; Гамарра, Беатрис; Сирак, Кендра; Коннелл, Сара; Стюардсон, Кристин; Харни, Эдаоин; Фу, Цяомэй; Гонсалес-Фортес, Глория; Джонс, Эппи Р.; Руденберг, Сонгул Алпаслан; Лендьель, Дьёрдь; Бокантен, Фанни; Гаспарян, Борис; Монж, Джанет М.; Грегг, Майкл; Эшед, Веред; Мизрахи, Ахува-Сиван; Мейкледжон, Кристофер; Герритсен, Фокке; Беженару, Луминица; Блюхер, Матиас; Кэмпбелл, Арчи; Каваллери, Джанпьеро; Комас, Дэвид; Фрогель, Филипп; Гилберт, Эдмунд; Керр, Шона М.; Ковач, Питер; Краузе, Джон; МакГеттиган, Даррен; Мерриган, Майкл; Мерриуэзер, Д. Эндрю; О'Рейли, Симус; Ричардс, Мартин Б.; Семино, Орнелла; Шамун-Пур, Мишель; Стефанеску, Георге; Стамволл, Майкл; Тёньес, Анке; Торрони, Антонио; Уилсон, Джеймс Ф.; Йенго, Лоик; Оганесян, Нелли А.; Паттерсон, Ник; Пинхаси, Рон; Райх, Дэвид (август 2016 г.). «Геномный взгляд на происхождение земледелия на древнем Ближнем Востоке» . Природа . 536 (7617): 419–424. Бибкод : 2016Natur.536..419L . дои : 10.1038/nature19310 . ПМЦ 5003663 . ПМИД 27459054 .
- ^ «Распространение быка» . Колыбель цивилизации . 15 июля 2018 г. Проверено 21 октября 2023 г.
- ^ Аль-Захери, Н. (2003). «Полиморфизмы Y-хромосомы и мтДНК в Ираке, перекресток раннего расселения человека и постнеолитических миграций». Молекулярная филогенетика и эволюция . 28 (3): 458–72. дои : 10.1016/s1055-7903(03)00039-3 . ПМИД 12927131 .
- ^ Небель А., Филон Д., Бринкманн Б., Маджумдер П.П., Фаерман М., Оппенгейм А. (ноябрь 2001 г.). «Пул Y-хромосомы евреев как часть генетического ландшафта Ближнего Востока» . Американский журнал генетики человека . 69 (5): 1095–1112. дои : 10.1086/324070 . ПМЦ 1274378 . ПМИД 11573163 . См. особенно рисунок шесть. Semino 2000 — это источник, который также утверждает, что Eu 9 происходит от Eu 10 (Eu 10 — это другой субклад гаплогруппы J (мтДНК) ).
- ^ Ди Джакомо Ф., Лука Ф., Попа Л.О., Акар Н., Ананью Н., Банико Дж. и др. (2004). «Y-хромосомная гаплогруппа J как признак постнеолитической колонизации Европы». Генетика человека . 115 (5): 357–71. дои : 10.1007/s00439-004-1168-9 . ПМИД 15322918 . S2CID 18482536 .
- ^ Аль-Захери, Н.; Семино, О.; Бенуцци, Г.; Магри, К.; Пассарино, Г.; Торрони, А.; Сантакьяра-Бенерекетти, AS (1 сентября 2003 г.). «Полиморфизм Y-хромосомы и мтДНК в Ираке, перекресток раннего расселения человека и постнеолитических миграций» . Молекулярная филогенетика и эволюция . Специальный выпуск: доклады, представленные на симпозиуме по филогении млекопитающих во время ежегодного собрания Общества молекулярной биологии и эволюции 2002 г., Сорренто, Италия, 13–16 июня 2002 г. 28 (3): 458–472. дои : 10.1016/S1055-7903(03)00039-3 . ISSN 1055-7903 .
- ^ Jump up to: а б Лю Шуху; Низам Ильхам; Рабиаб Баке; Абдулла Бупатима; Долкун Матюсуп (2018). «Инъюн Ю. Ринсэту SNP duì Синьцзян Сан Гели Ренцюнь Ичуань Дуоян из Синьцзяна» Исследование генетического разнообразия трех изолированных популяций в Синьцзяне с использованием SNP Y-хромосомы [Исследование генетического разнообразия трех изолированных популяций в Синьцзяне с использованием Y-SNP]. Anthropologica Sinica / Acta Anthropologica Sinica (на китайском языке). 37 (1): 146–156. doi : 10.16359/j.cnki.cn11-1963/q.2017.0067 .
- ^ Шоу, Вэй-Хуа; Цяо, Внь-Фа; Вэй, Чуан-Юй; Донг, Юн-Ли; Тан, Си-Цзе; Ши, Хун; Тан, Вэнь-Ру; Сяо, Чун-Цзе (2010). «Распределение Y-хромосомы среди популяций Северо-Западного Китая указывает на значительный вклад скотоводов Центральной Азии и меньшее влияние западных евразийцев» . Джей Хум Жене . 55 (5): 314–322. дои : 10.1038/jhg.2010.30 . ПМИД 20414255 . S2CID 23002493 .
- ^ Аллентофт; и др. (2015). «Популяционная геномика Евразии бронзового века» . Природа . 522 (7555): 167–172. Бибкод : 2015Natur.522..167A . дои : 10.1038/nature14507 . ПМИД 26062507 . S2CID 4399103 . Архивировано из оригинала 04 февраля 2020 г. Проверено 21 января 2020 г.
- ^ C. Rоttensteiner, J2a2-PH3085, SK1403: Древний Алтай, современный уйгурский и турецкий язык. Архивировано 26 июня 2015 г. в Wayback Machine , Исследование гаплогрупп J2-M172.
- ^ Ф. Иммануэль, Коды для результатов Gedmatch, страница древней ДНК. Архивировано 5 сентября 2015 г. в Wayback Machine , F999962 для RISE504, образец Кытманово и F999965 для RISE602, образец Сары-Бел.
- ^ Аль-Захери и др., 2011, Дополнительный файл 3. Абсолютные частоты гаплогрупп и субгаплогрупп Y-хромосомы в 48 популяциях, включенных в PCA. Архивировано 6 ноября 2015 г. в Wayback Machine. Примечание. Только 37 из 154 образцов (24%) относятся к J2 в Ираке согласно списку Аль-Захери за 2011 г. 43,6% - это частота J2 среди всех иракцев гаплогруппы J, а не у всех. гаплогруппы.
- ^ Абу-Амеро (2009), Разнообразие Y-хромосомы Саудовской Аравии и его связь с близлежащими регионами. Архивировано 3 апреля 2017 г. в Wayback Machine , Цитата: Самые распространенные гаплогруппы в Саудовской Аравии, J1-M267 (42%), J2. -М172 (14%), Е1-М2 (8%), R1-M17 (5%) и К2-М184 (5%) также хорошо представлены в других арабских популяциях (Таблица (Таблица1).1).
- ^ Jump up to: а б с Сингх С., Сингх А., Раджкумар Р., Сампат Кумар К., Кадаркарай Сами С., Низамуддин С. и др. (январь 2016 г.). «Анализ влияния неолитической демической диффузии на индийский пул Y-хромосом через гаплогруппу J2-M172» . Научные отчеты . 6 : 19157. Бибкод : 2016NatSR...619157S . дои : 10.1038/srep19157 . ПМК 4709632 . ПМИД 26754573 .
- ^ Эррера, Рене Дж.; Гарсиа-Бертран, Ральф (2018). ДНК предков, происхождение человека и миграции . Академическая пресса. п. 250. ИСБН 978-0-12-804128-4 .
- ^ Jump up to: а б с д Сенгупта С., Животовский Л.А., Кинг Р., Мехди С.К., Эдмондс К.А., Чоу С.Э. и др. (февраль 2006 г.). «Полярность и временность распределения Y-хромосомы в Индии с высоким разрешением определяют как местное, так и экзогенное распространение и выявляют незначительное генетическое влияние скотоводов Центральной Азии» . Американский журнал генетики человека . 78 (2): 202–21. дои : 10.1086/499411 . ПМЦ 1380230 . ПМИД 16400607 .
- ^ Арункумар Г., Сориа-Эрнанц Д.Ф., Кавита В.Дж., Арун В.С., Шьяма А., Ашокан К.С. и др. (2012). «Дифференциация населения мужских родов Южной Индии коррелирует с сельскохозяйственной экспансией, предшествовавшей кастовой системе» . ПЛОС ОДИН . 7 (11): е50269. Бибкод : 2012PLoSO...750269A . дои : 10.1371/journal.pone.0050269 . ПМК 3508930 . ПМИД 23209694 .
- ^ Тансим И., Тангарадж К., Чауби Дж., Сингх В.К., Бхаскар Л.В., Редди Б.М. и др. (август 2006 г.). «Генетическое сходство среди низших каст и племенных групп Индии: вывод на основе Y-хромосомы и митохондриальной ДНК» . БМК Генетика . 7:42 . дои : 10.1186/1471-2156-7-42 . ПМЦ 1569435 . ПМИД 16893451 .
- ^ Шарма С., Рай Э., Шарма П., Джена М., Сингх С., Дарвиши К. и др. (январь 2009 г.). «Индийское происхождение отцовской гаплогруппы R1a1* обосновывает автохтонное происхождение брахманов и кастовой системы» . Журнал генетики человека . 54 (1): 47–55. дои : 10.1038/jhg.2008.2 . ПМИД 19158816 .
- ^ Jump up to: а б Камар Р., Аюб К., Мохьюддин А. и др. (май 2002 г.). «Вариант ДНК Y-хромосомы в Пакистане» . Являюсь. Дж. Хум. Жене . 70 (5): 1107–24. дои : 10.1086/339929 . ПМЦ 447589 . ПМИД 11898125 .
- ^ Шах А.М., Таманг Р., Мурджани П., Рани Д.С., Говиндарадж П., Кулкарни Дж., Бхаттачарья Т., Мустак М.С., Бхаскар Л.В., Редди А.Г., Гадхви Д., Гай П.Б., Чаубей Дж., Паттерсон Н., Райх Д., Тайлер-Смит С., Сингх Л., Тангарадж К. (2011). «Индийские сидди: африканские потомки с индийской примесью» . Являюсь. Дж. Хум. Жене . 89 (1): 154–61. дои : 10.1016/j.ajhg.2011.05.030 . ПМК 3135801 . ПМИД 21741027 .
- ^ Тоомас Кивисилд; Сиири Роотси; Мейт Метспалу; Эне Мецпалу; Юри Парик; Катрин Калдма; Эсиен Усанга; Сарабджит Мастана; Суриндер С. Папиха; Ричард Виллемс (2002). «Генетика языка и распространения сельского хозяйства в Индии» (PDF) . В Питере Беллвуде; Колин Ренфрю (ред.). Исследование гипотезы расселения сельского хозяйства/языка . Монографии Института Макдональда. Институт археологических исследований Макдональда. ISBN 9781902937205 .
- ^ «Происхождение мальдивских островитян в свете популяционной генетики: мальдивское происхождение в свете генетики» . 24 мая 2013. Архивировано из оригинала 29 октября 2013 года . Проверено 6 августа 2016 г.
- ^ Jump up to: а б с Гьянешвер Чаубей, Манвендра Сингх, Федерика Кривелларо и др. , «Распутывание различных линий происхождения Тару». Европейский журнал генетики человека (2014), 1–9. doi:10.1038/ejhg.2014.36
- ^ Симона Форнарино, Мария Пала, Винченца Батталья и др. , «Митохондриальное и Y-хромосомное разнообразие народа Тарус (Непал): резервуар генетических вариаций». Эволюционная биология BMC 2009, 9:154. дои: 10.1186/1471-2148-9-154
- ^ Сингх, Сакши; Сингх, Ашиш; Раджкумар, Раджа; Сампат Кумар, Катакам; Кадаркарай Сами, Суббурадж; Низамуддин, шейх; Сингх, Амита; Ахмед Шейх, Шахнаваз; Педдада, Видья; Ханна, Вини; Вирайя, Пандичелвам; Пандит, Аридаман; Чаубей, Гьянешвер; Сингх, Лалджи; Тангарадж, Кумарасами (12 января 2016 г.). «Анализ влияния неолитической демической диффузии на индийский пул Y-хромосом через гаплогруппу J2-M172» . Научные отчеты . 6 (1): 19157. doi : 10.1038/srep19157 . ISSN 2045-2322 . ПМК 4709632 . ПМИД 26754573 .
- ^ «Осетинский проект ДНК — классическая диаграмма Y-ДНК» . семьятридна.
- ^ «Гаплогруппа J2 (Y-ДНК)» . Генетический атлас . Архивировано из оригинала 24 сентября 2010 г. Проверено 27 декабря 2010 г.
- ^ «Гаплодерево Y-ДНК» . Архивировано из оригинала 27 января 2013 г. Проверено 5 января 2013 г. ДНК генеалогического древа использует дерево Консорциума Y-хромосомы и публикует его на своем веб-сайте.
Источники таблиц преобразования
[ редактировать ]- Капелли, Кристиан; Уилсон, Джеймс Ф.; Ричардс, Мартин; Штумпф, Майкл П.Х.; и др. (февраль 2001 г.). «Отцовское наследие преимущественно коренных народов австронезийскоязычных народов островной Юго-Восточной Азии и Океании» . Американский журнал генетики человека . 68 (2): 432–443. дои : 10.1086/318205 . ПМЦ 1235276 . ПМИД 11170891 .
- Хаммер, Майкл Ф.; Карафет Татьяна М.; Редд, Алан Дж.; Джарджанази, Хамди; и др. (1 июля 2001 г.). «Иерархические закономерности глобального разнообразия Y-хромосом человека» . Молекулярная биология и эволюция . 18 (7): 1189–1203. doi : 10.1093/oxfordjournals.molbev.a003906 . ПМИД 11420360 .
- Джоблинг, Марк А.; Тайлер-Смит, Крис (2000), «Новое использование новых гаплотипов», Trends in Genetics , 16 (8): 356–62, doi : 10.1016/S0168-9525(00)02057-6 , PMID 10904265
- Каладжиева, Люба; Калафель, Франческ; Джоблинг, Марк А; Ангеличева, Дора; и др. (февраль 2001 г.). «Закономерности меж- и внутригруппового генетического разнообразия у влакс-рома, выявленные с помощью Y-хромосомы и линий митохондриальной ДНК» . Европейский журнал генетики человека . 9 (2): 97–104. дои : 10.1038/sj.ejhg.5200597 . ПМИД 11313742 . S2CID 21432405 .
- Карафет, Татьяна; Сюй, Липин; Ду, Руофу; Ван, Уильям; и др. (сентябрь 2001 г.). «История отцовского населения Восточной Азии: источники, закономерности и микроэволюционные процессы» . Американский журнал генетики человека . 69 (3): 615–628. дои : 10.1086/323299 . ПМЦ 1235490 . ПМИД 11481588 .
- Карафет, ТМ; Мендес, Флорида; Мейлерман, МБ; Андерхилл, Пенсильвания; Зегура, СЛ; Хаммер, М.Ф. (2008), «Новые бинарные полиморфизмы изменяют форму и повышают разрешение дерева гаплогрупп Y-хромосомы человека», Genome Research , 18 (5): 830–8, doi : 10.1101/gr.7172008 , PMC 2336805 , PMID 18385274
- Семино, О.; Пассарино, Дж; Оефнер, П.Дж.; Лин, А.А.; и др. (2000), «Генетическое наследие палеолитического Homo sapiens sapiens у современных европейцев: хромосомная перспектива AY», Science , 290 (5494): 1155–9, Бибкод : 2000Sci...290.1155S , doi : 10.1126/science.290.5494. 1155 , ПМИД 11073453
- Су, Бинг; Сяо, Цзюньхуа; Андерхилл, Питер; Дека, Ранджан; и др. (декабрь 1999 г.). «Свидетельства Y-хромосомы о миграции современных людей на север в Восточную Азию во время последнего ледникового периода» . Американский журнал генетики человека . 65 (6): 1718–1724. дои : 10.1086/302680 . ПМЦ 1288383 . ПМИД 10577926 .
- Андерхилл, Питер А.; Шен, Пейдун; Лин, Алиса А.; Джин, Ли; и др. (ноябрь 2000 г.). «Вариации последовательности Y-хромосомы и история человеческих популяций». Природная генетика . 26 (3): 358–361. дои : 10.1038/81685 . ПМИД 11062480 . S2CID 12893406 .
Примечания
[ редактировать ]- ^ Ренфрю AC (1998). Археология и язык: загадка индоевропейского происхождения (изд. Пимлико). Лондон: Пимлико. ISBN 978-0-7126-6612-1 .
- ^ Небель А., Филон Д., Бринкманн Б., Маджумдер П.П., Фаерман М., Оппенгейм А. (2001). «Пул Y-хромосомы евреев как часть генетического ландшафта Ближнего Востока» . Являюсь. Дж. Хум. Жене . 69 (5): 1095–112. дои : 10.1086/324070 . ПМЦ 1274378 . ПМИД 11573163 .
- ^ Маласпина П., Цопаномихалу М., Думан Т., Стефан М., Сильвестри А., Ринальди Б. и др. (2001). «Многоэтапный процесс распространения Y-хромосомной линии в Средиземноморье» (PDF) . Энн. Хм. Жене . 65 (Часть 4): 339–49. дои : 10.1046/j.1469-1809.2001.6540339.x . hdl : 2108/44448 . ПМИД 11592923 . S2CID 221448190 .
Библиография
[ редактировать ]- Абу-Амеро К.К., Хеллани А., Гонсалес А.М., Ларруга Х.М., Кабрера В.М., Андерхилл, Пенсильвания (2009). «Разнообразие Y-хромосомы Саудовской Аравии и его связь с близлежащими регионами» . БМК Генетика . 10:59 . дои : 10.1186/1471-2156-10-59 . ПМК 2759955 . ПМИД 19772609 .
- Адамс, С.М.; Бош, Э; Балареск, Польша; Баллеро, SJ; Ли, AC; Арройо, Э; Лопес-Парра, AM; Алер, М; и др. (2008). «Генетическое наследие религиозного разнообразия и нетерпимости: отцовские линии христиан, евреев и мусульман на Пиренейском полуострове» . Американский журнал генетики человека . 83 (6): 725–36. дои : 10.1016/j.ajhg.2008.11.007 . ПМК 2668061 . ПМИД 19061982 .
- Аль-Захери Н., Семино О., Бенуцци Г., Магри С., Пассарино Дж., Торрони А., Сантакьяра-Бенерекетти А.С. (2003). «Полиморфизмы Y-хромосомы и мтДНК в Ираке, перекресток раннего расселения человека и постнеолитических миграций». Молекулярная филогенетика и эволюция . 28 (3): 458–72. дои : 10.1016/S1055-7903(03)00039-3 . ПМИД 12927131 .
- Аль-Захери Н., Пала М., Батталья В., Гругни В., Хамод М.А., Хушиар Кашани Б. и др. (2011). «В поисках генетических следов шумеров: исследование вариаций Y-хромосомы и мтДНК у болотных арабов Ирака» . Эволюционная биология BMC . 11 : 288. дои : 10.1186/1471-2148-11-288 . ПМЦ 3215667 . ПМИД 21970613 .
- Арреди Б., Полони Э.С., Параккини С., Зержал Т., Фаталлах Д.М., Макрелуф М., Паскали В.Л., Новеллетто А., Тайлер-Смит С. (2004). «Преимущественно неолитическое происхождение вариаций ДНК Y-хромосомы в Северной Африке» . Американский журнал генетики человека . 75 (2): 338–45. дои : 10.1086/423147 . ПМК 1216069 . ПМИД 15202071 .
- Аширбеков Э.Э.; и др. (2017). «Распространение гаплогрупп Y-хромосомы казахов Южно-Казахстанской, Жамбылской и Алматинской областей» (PDF) . Доклады Национальной академии наук Республики Казахстан . 6 (316): 85–95. Архивировано из оригинала (PDF) 23 сентября 2021 г. Проверено 26 мая 2023 г.
- Балановский О, Дибирова К, Дыбо А, Мудрак О, Фролова С, Почешхова Е и др. (2011). «Параллельная эволюция генов и языков в Кавказском регионе » Молекулярная биология и эволюция . 28 (10): 2905–20. дои : 10.1093/molbev/msr126 . ПМЦ 3355373 . ПМИД 21571925 .
- Батталья В, Форнарино С, Аль-Захери Н, Оливьери А, Пала М, Майрес Н.М., Кинг Р.Дж., Роотси С, Марьянович Д, Приморак Д, Хаджиселимович Р, Видович С, Дробнич К, Дурмиши Н, Торрони А, Сантакьяра-Бенерекетти А.С., Андерхилл П.А., Семино О. (2008). «Y-хромосомные свидетельства культурного распространения сельского хозяйства в Юго-Восточной Европе» . Европейский журнал генетики человека . 17 (6): 820–30. дои : 10.1038/ejhg.2008.249 . ПМК 2947100 . ПМИД 19107149 .
- Бехар, Дорон М.; Гарриган, Дэниел; Каплан, Мэтью Э.; Мобашер, Захра; Розенгартен, Дрор; Карафет Татьяна М.; Кинтана-Мурси, Луис; Острер, Гарри; Скорецкий, Карл; Хаммер, Майкл Ф. (1 марта 2004 г.). «Контрастные закономерности вариаций Y-хромосомы у евреев-ашкенази и нееврейского европейского населения». Генетика человека . 114 (4): 354–365. дои : 10.1007/s00439-003-1073-7 . ПМИД 14740294 . S2CID 10310338 .
- Каденас А.М., Животовский Л.А., Кавалли-Сфорца Л.Л., Андерхилл П.А., Эррера Р.Дж. (2007). «Разнообразие Y-хромосом характеризует Оманский залив» . Европейский журнал генетики человека . 16 (3): 374–86. дои : 10.1038/sj.ejhg.5201934 . ПМИД 17928816 .
- Капелли С., Редхед Н., Романо В., Кали Ф., Лефранк Г., Делаг В. и др. (2006). «Структура населения в Средиземноморском бассейне: хромосомная перспектива AY». Анналы генетики человека . 70 (2): 207–25. дои : 10.1111/j.1529-8817.2005.00224.x . hdl : 2108/37090 . ПМИД 16626331 . S2CID 25536759 .
- Капелли С., Бризигелли Ф., Скарниччи Ф., Арреди Б., Калья А., Ветруньо Г. и др. (2007). «Генетическая вариация Y-хромосомы на итальянском полуострове является клинальной и подтверждает модель примеси для встречи мезолита и неолита». Молекулярная филогенетика и эволюция . 44 (1): 228–39. дои : 10.1016/j.ympev.2006.11.030 . ПМИД 17275346 .
- Чинниоглу С., Кинг Р., Кивисилд Т., Калфоглу Е., Атасой С., Каваллери Г.Л. и др. (2004). «Раскопки слоев гаплотипов Y-хромосомы в Анатолии». Генетика человека . 114 (2): 127–48. дои : 10.1007/s00439-003-1031-4 . ПМИД 14586639 . S2CID 10763736 .
- Ди Джакомо Ф, Лука Ф, Ананью Н, Чаварелла Дж, Корбо Р.М., Креста М, Куччи Ф, Ди Стази Л, Агостиано В, Гипараки М, Лутрадис А, Мамми К, Михалодимитракис Э.Н., Папола Ф, Педичини Г, Плата Е , Терренато Л., Тофанелли С., Маласпина П., Новеллетто А. (2003). «Клинические закономерности разнообразия Y-хромосомы человека в континентальной Италии и Греции определяются эффектами дрейфа и основателя». Молекулярная филогенетика и эволюция . 28 (3): 387–95. дои : 10.1016/S1055-7903(03)00016-2 . ПМИД 12927125 .
- Иасвархант М., Хак И., Равеш З., Ромеро И.Г., Меганатан П.Р., Дубей Б. и др. (2009). «Следы происхождения к югу от Сахары и Ближнего Востока в мусульманском населении Индии» . Европейский журнал генетики человека . 18 (3): 354–63. дои : 10.1038/ejhg.2009.168 . ПМЦ 2859343 . ПМИД 19809480 .
- Эль-Сибай М., Платт Д.Э., Хабер М., Сюэ Ю., Юханна С.С., Уэллс Р.С. и др. (2009). «Географическая структура Y-хромосомного генетического ландшафта Леванта: контраст между побережьем и сушей» . Анналы генетики человека . 73 (6): 568–81. дои : 10.1111/j.1469-1809.2009.00538.x . ПМЦ 3312577 . ПМИД 19686289 . В этом документе представлены результаты нескольких исследований: Ди Джакомо, 2003 г., Аль-Захери, 2003 г., Флорес, 2004 г., Чинниоглу, 2004 г., Капелли, 2005 г., Гонсалвес, 2005 г., Заллуа, 2008 г., Каденас, 2008 г.
- Фадлауи-Зид, Карима (2014). «Сус: крайняя генетическая гетерогенность в Северной Африке» . Журнал генетики человека . 60 (1): 41–49. дои : 10.1038/jhg.2014.99 . ПМИД 25471516 . S2CID 25186140 .
- Фрегель, Роза; ГОМЕС, Вероника; ГУСМИО, Леонор; Гонсалес, Ана М; Кабрера, Висенте М; АМОРИМ, Антониу; Ларруга, Хосе М. (2009). «Демографическая история мужского генофонда Канарских островов: замена местных линий Европой» . Эволюционная биология BMC . 9 (1): 181. дои : 10.1186/1471-2148-9-181 . ПМЦ 2728732 . ПМИД 19650893 .
- Фирасат, Садаф; Халик, Шагуфта; Мохьюддин, Аиша; Папайоанну, Мирто; Тайлер-Смит, Крис; Андерхилл, Питер А.; Аюб, Касим (2007). «Y-хромосомные доказательства ограниченного вклада греков в патанское население Пакистана» . Европейский журнал генетики человека . 15 (1): 121–126. дои : 10.1038/sj.ejhg.5201726 . ПМЦ 2588664 . ПМИД 17047675 .
- Ди Джакомо Ф., Лука Ф., Попа Л.О., Акар Н., Ананью Н., Банико Дж. и др. (2004). «Y-хромосомная гаплогруппа J как признак постнеолитической колонизации Европы». Генетика человека . 115 (5): 357–71. дои : 10.1007/s00439-004-1168-9 . ПМИД 15322918 . S2CID 18482536 .
- Гругни В., Батталья В., Хушиар Кашани Б., Пароло С., Аль-Захери Н., Ахилли А. и др. (2012). Кивисилд Т (ред.). «Древние миграционные события на Ближнем Востоке: новые данные по Y-хромосомной вариации современных иранцев» . ПЛОС ОДИН . 7 (7): е41252. Бибкод : 2012PLoSO...741252G . дои : 10.1371/journal.pone.0041252 . ПМЦ 3399854 . ПМИД 22815981 .
- Го, Юйсинь; Ся, Чжию; Цуй, Вэй; Чен, Чонг; Джин, Сяое; Чжу, Бофэн (18 мая 2020 г.). «Совместный генетический анализ молекулярных маркеров митохондрий и Y-хромосомы для населения Северо-Западного Китая» . Гены . 11 (5): 564. doi : 10.3390/genes11050564 . ПМК 7290686 . ПМИД 32443545 .
- Хабер М., Платт Д.Э., Ашрафиан Бонаб М., Юханна С.К., Сориа-Эрнанц Д.Ф., Мартинес-Круз Б. и др. (2011). Кайзер М. (ред.). «Этнические группы Афганистана разделяют Y-хромосомное наследие, структурированное историческими событиями» . ПЛОС ОДИН . 7 (3): e34288. Бибкод : 2012PLoSO...734288H . дои : 10.1371/journal.pone.0034288 . ПМЦ 3314501 . ПМИД 22470552 .
- Кинг Р., Андерхилл, Пенсильвания (2002). «Согласованное распределение расписной керамики и керамических фигурок эпохи неолита с линиями Y-хромосомы». Античность . 76 (293): 707–714. дои : 10.1017/s0003598x00091158 . S2CID 160359661 .
- Кинг Р.Дж., Озджан С.С., Картер Т., Калфоглу Э., Атасой С., Триантафиллидис С. и др. (2008). «Дифференциальное анатолийское влияние Y-хромосомы на греческий и критский неолит». Анналы генетики человека . 72 (2): 205–14. дои : 10.1111/j.1469-1809.2007.00414.x . ПМИД 18269686 . S2CID 22406638 .
- Кран; ФТДНА (2003). «Черновик дерева Центра геномных исследований (AKA Y-TRee)» . Архивировано из оригинала 15 августа 2015 г.
- Луис Дж.Р., Роуолд Д.Д., Регейру М., Каейро Б., Чинниоглу С., Роузман С. и др. (2004). «Левант против Африканского Рога: свидетельства двунаправленных коридоров миграции людей» . Американский журнал генетики человека . 74 (3): 532–44. дои : 10.1086/382286 . ПМЦ 1182266 . ПМИД 14973781 .
- Маласпина П., Цопаномихалу М., Думан Т., Стефан М., Сильвестри А., Ринальди Б. и др. (2001). «Многоэтапный процесс распространения Y-хромосомной линии в Средиземноморье». Анналы генетики человека . 65 (4): 339–49. дои : 10.1046/j.1469-1809.2001.6540339.x . hdl : 2108/44448 . ПМИД 11592923 . S2CID 221448190 .
- Мартинес Л., Андерхилл П.А., Животовский Л.А., Гайден Т., Мошонас Н.К., Чоу CE и др. (2007). «На Критском высокогорном плато преобладает палеолитическое наследие Y-гаплогруппы» . Европейский журнал генетики человека . 15 (4): 485–93. дои : 10.1038/sj.ejhg.5201769 . PMID 17264870 .
- Насидзе И., Саркисян Т., Керимов А., Стоункинг М. (2003). «Проверка гипотезы замены языка на Кавказе: данные Y-хромосомы». Генетика человека . 112 (3): 255–61. дои : 10.1007/s00439-002-0874-4 . ПМИД 12596050 . S2CID 13232436 .
- Насидзе И., Линг Е.Ю., Куинке Д., Дюпанлуп И., Кордо Р., Рычков С. и др. (2004). «Митохондриальная ДНК и вариации Y-хромосомы на Кавказе» . Анналы генетики человека . 68 (3): 205–21. дои : 10.1046/j.1529-8817.2004.00092.x . ПМИД 15180701 . S2CID 27204150 .
- Небель А., Филон Д., Бринкманн Б., Маджумдер П.П., Фаерман М., Оппенгейм А. (2001). «Пул Y-хромосомы евреев как часть генетического ландшафта Ближнего Востока» . Американский журнал генетики человека . 69 (5): 1095–112. дои : 10.1086/324070 . ПМЦ 1274378 . ПМИД 11573163 .
- Ногейро, И.; Манко, Л.; Гомес, В.; Аморим, А.; Гужман, Л. (2010). «Филогеографический анализ отцовских линий в еврейских общинах северо-восточной Португалии» . Американский журнал физической антропологии . 141 (3): 373–381. дои : 10.1002/ajpa.21154 . ПМИД 19918998 .
- Регейро М., Каденас А.М., Гайден Т., Андерхилл П.А., Эррера Р.Дж. (2006). «Иран: трехконтинентальная связь для миграции, вызванной Y-хромосомой». Наследственность человека . 61 (3): 132–43. дои : 10.1159/000093774 . ПМИД 16770078 . S2CID 7017701 .
- Робино, К.; Кробу, Ф.; Ди Гаэтано, К.; Бекада, А.; Бенхамамуш, С.; Черутти, Н.; Пьяцца, А.; Интурри, С.; Торре, К. (2008). «Анализ гаплогрупп SNP Y-хромосомы и гаплотипов STR в выборке населения Алжира». Международный журнал юридической медицины . 122 (3): 251–255. дои : 10.1007/s00414-007-0203-5 . ПМИД 17909833 . S2CID 11556974 .
- Санчес Дж.Дж., Халленберг С., Бёрстинг С., Эрнандес А., Морлинг Н. (2005). «Высокие частоты линий Y-хромосомы, характеризующиеся E3b1, DYS19-11, DYS392-12 у сомалийских мужчин» . Европейский журнал генетики человека . 13 (7): 856–66. дои : 10.1038/sj.ejhg.5201390 . ПМИД 15756297 .
- Семино О., Пассарино Г., Офнер П.Дж., Лин А.А., Арбузова С., Бекман Л.Е. и др. (2000). «Генетическое наследие палеолитического Homo sapiens sapiens у современных европейцев: хромосомная перспектива AY». Наука . 290 (5494): 1155–9. Бибкод : 2000Sci...290.1155S . дои : 10.1126/science.290.5494.1155 . ПМИД 11073453 .
- Семино О., Магри С., Бенуцци Г., Лин А.А., Аль-Захери Н., Батталья В. и др. (2004). «Происхождение, распространение и дифференциация гаплогрупп E и J Y-хромосомы: выводы о неолитизации Европы и более поздних миграционных событиях в Средиземноморье» . Американский журнал генетики человека . 74 (5): 1023–34. дои : 10.1086/386295 . ПМК 1181965 . ПМИД 15069642 .
- Сенгупта С., Животовский Л.А., Кинг Р., Мехди С.К., Эдмондс К.А., Чоу С.Э. и др. (2006). «Полярность и временность распределения Y-хромосомы с высоким разрешением в Индии определяют как местное, так и экзогенное распространение и выявляют незначительное генетическое влияние скотоводов Центральной Азии» . Американский журнал генетики человека . 78 (2): 202–21. дои : 10.1086/499411 . ПМЦ 1380230 . ПМИД 16400607 .
- Шен П., Лави Т., Кивисилд Т., Чоу В., Сенгун Д., Гефель Д. и др. (2004). «Реконструкция отцовских и матрилиний самаритян и других израильских популяций на основе вариаций Y-хромосомы и последовательности митохондриальной ДНК». Человеческая мутация . 24 (3): 248–60. дои : 10.1002/humu.20077 . ПМИД 15300852 . S2CID 1571356 .
- Судьял, Х (2013). «Возврат к происхождению лембы: отслеживание происхождения Y-хромосом у южноафриканской и зимбабвийской лембы» . Южноафриканский медицинский журнал . 103 (12): 1009–1013. дои : 10.7196/SAMJ.7297 . ПМИД 24300649 . Проверено 9 мая 2014 г.
- Тофанелли С., Ферри Г., Булаева К., Качиагли Л., Онофри В., Тальоли Л. и др. (2009). «Происхождение J1-M267 Y знаменует собой вызванное климатом доисторическое перемещение людей» . Европейский журнал генетики человека . 17 (11): 1520–4. дои : 10.1038/ejhg.2009.58 . ПМЦ 2986692 . ПМИД 19367321 .
- Андерхилл П.А., Шен П., Лин А.А., Джин Л., Пассарино Г., Ян В.Х., Кауфман Э., Бонне-Тамир Б., Бертранпетит Дж., Франкалаччи П., Ибрагим М., Дженкинс Т., Кидд Дж.Р., Мехди С.К., Сейелстад М.Т., Уэллс Р.С., Пьяцца А, Дэвис Р.В., Фельдман М.В., Кавалли-Сфорца Л.Л., Офнер П.Дж. (2000). «Вариации последовательности Y-хромосомы и история человеческих популяций». Природная генетика . 26 (3): 358–61. дои : 10.1038/81685 . ПМИД 11062480 . S2CID 12893406 .
- Уэллс Р.С., Юлдашева Н., Рузибакиев Р., Андерхилл П.А., Евсеева И., Блю-Смит Дж. и др. (2001). «Евразийский Хартленд: континентальный взгляд на разнообразие Y-хромосомы» . Труды Национальной академии наук . 98 (18): 10244–9. Бибкод : 2001PNAS...9810244W . дои : 10.1073/pnas.171305098 . ПМК 56946 . ПМИД 11526236 .
- Юнусбаев Б, Мецпалу М, Ярве М, Кутуев И, Роотси С, Мецпалу Э, Бехар ДМ, Варенди К, Саакян Х, Хусаинова Р, Епископосян Л, Хуснутдинова Е.К., Андерхилл П.А., Кивисилд Т, Виллемс Р (2012). «Кавказ как асимметричный полупроницаемый барьер на пути древних миграций человека» . Молекулярная биология и эволюция . 29 (1): 359–65. дои : 10.1093/molbev/msr221 . ПМИД 21917723 .
- Заллуа П.А., Сюэ Ю., Халифе Дж., Махул Н., Дебиан Л., Платт Д.Э. и др. (2008л). «Разнообразие Y-хромосомы в Ливане структурировано недавними историческими событиями» . Американский журнал генетики человека . 82 (4): 873–82. дои : 10.1016/j.ajhg.2008.01.020 . ПМЦ 2427286 . ПМИД 18374297 .
- Ди Кристофаро Дж., Пеннарун Э., Мазьер С., Майрес Н.М., Лин А.А., Темори С.А. и др. (2013). «Афганский Гиндукуш: место, где сходятся потоки генов Евразийского субконтинента» . ПЛОС ОДИН . 8 (10): е76748. Бибкод : 2013PLoSO...876748D . дои : 10.1371/journal.pone.0076748 . ПМЦ 3799995 . ПМИД 24204668 .
- Шоу В.Х., Цяо Э.Ф., Вэй С.И., Донг Ю.Л., Тан С.Дж., Ши Х. и др. (2010). «Распределение Y-хромосомы среди популяций Северо-Западного Китая указывает на значительный вклад скотоводов Центральной Азии и меньшее влияние западных евразийцев» . Журнал генетики человека . 55 (5): 314–22. дои : 10.1038/jhg.2010.30 . ПМИД 20414255 .
Внешние ссылки
[ редактировать ]- Миграция индейцев по континентам на протяжении поколений: история семьи Салуджа.
- В Ливане ДНК еще может залечить трещины
Филогенетические заметки
[ редактировать ]- ^ В этой таблице показаны исторические названия J-M172 в опубликованной рецензируемой литературе. Обратите внимание, что в Semino 2000 Eu09 является субкладом Eu10, а в Karafet 2001 24 является субкладом 23 .
YCC 2002/2008 (сокращенно) J-M172 Джоблинг и Тайлер-Смит, 2000 г. 9 Андерхилл 2000 МЫ Хаммер 2001 С Карафет 2001 24 Семино 2000 I9 Они в 1999 году Н4 Волосы 2001 Б YCC 2002 (от руки) J2* YCC 2005 (от руки) J2 YCC 2008 (от руки) J2 YCC 2010r (от руки) J2 - ^ В этой таблице показаны исторические названия J-P209 (AKA J-12f2.1 или J-M304) в опубликованной рецензируемой литературе. Обратите внимание, что в Semino 2000 Eu09 является субкладом Eu10, а в Karafet 2001 24 является субкладом 23 .
YCC 2002/2008 (сокращенно) J-P209
(АКА J-12f2.1 или J-M304)Джоблинг и Тайлер-Смит, 2000 г. 9 Андерхилл 2000 МЫ Хаммер 2001 С Карафет 2001 23 Семино 2000 ЕС10 Они в 1999 году Н4 Волосы 2001 Б YCC 2002 (от руки) Дж* YCC 2005 (от руки) Дж YCC 2008 (от руки) Дж YCC 2010r (от руки) Дж - ^ «ISOGG 2018 Гаплогруппа J Y-ДНК» . www.isogg.org . Архивировано из оригинала 18 августа 2017 г. Проверено 11 апреля 2010 г.