Гаплогруппа J-M267
| Гаплогруппа J-M267 | |
|---|---|
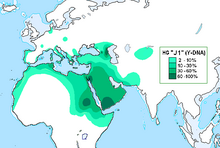 Интерполированное географическое частотное распределение. [ 1 ] | |
| Возможное время происхождения | 17,000 [ 2 ] –24000 лет назад ( Ди Джакомо, 2004 г. ) |
| Возможное место происхождения | Западная Азия [ 3 ] [ 4 ] |
| Предок | J-P209 |
| Потомки | ДЖ-М62, ДЖ-М365.1, ДЖ-Л136, ДЖ-З1828 |
| Определение мутаций | М267, Л255, Л321, Л765, Л814, Л827, Л1030 |
Гаплогруппа J-M267 , также широко известная как гаплогруппа J1 , представляет собой субклад (ветвь) гаплогруппы Y-ДНК J-P209 (широко известной как гаплогруппа J ) вместе с ее родственной гаплогруппой J-M172 (широко известной как гаплогруппа J2 ). (Все эти гаплогруппы имели другие исторические названия, перечисленные ниже. [ Филогенетика 1 ] [ Филогенетика 2 ] )
Мужчины этой линии имеют общего предка по отцовской линии , что подтверждается и определяется наличием мутации однонуклеотидного полиморфизма (SNP), называемой M267, о которой было объявлено в ( Cinnioğlu 2004 ). Эта гаплогруппа сегодня встречается со значительной частотой во многих районах Аравийского полуострова и Западной Азии или вблизи них . За пределами своего родного азиатского континента он очень часто встречается в Судане . Он также встречается в очень высокой, но меньшей степени, в некоторых частях Кавказа , Эфиопии и некоторых частях Северной Африки , а также среди большинства народов Леванта, включая еврейские группы , особенно тех, кто носит Коэн фамилию . Его также можно встретить гораздо реже, но все же иногда в значительных количествах, в некоторых частях южной Европы и на востоке, вплоть до Центральной Азии . [ нужна ссылка ]
Происхождение
[ редактировать ]С момента открытия гаплогруппы J-P209 было общепризнано, что она имеет признаки того, что она возникла примерно 20 000 лет назад где-то на северо-западе Ирана, на Кавказе, в Армянском нагорье и в северной Месопотамии. [ 5 ] [ 6 ] [ 3 ] Частота и разнообразие обеих его основных ветвей, J-M267 и J-M172, в этом регионе делают их кандидатами в качестве генетических маркеров распространения сельскохозяйственных технологий во время неолита , которые, как предполагается, оказали серьезное влияние на человеческое население.
J-M267 имеет несколько признанных субкладов, некоторые из которых были признаны до того, как был признан сам J-M267, например J-M62 Y Chromosome Consortium "YCC" 2002 . За одним примечательным исключением, J-P58, большинство из них встречаются нечасто ( Tofanelli 2009 ). Из-за доминирования J-P58 в популяциях J-M267 во многих областях обсуждение происхождения J-M267 требует одновременного обсуждения J-P58.
Распределение
[ редактировать ]Африка
[ редактировать ]Северная Африка и Африканский Рог
[ редактировать ]Северная Африка получила миграцию семитов; согласно некоторым исследованиям, она могла быть распространена в последнее время арабами, которые, в основном с 7-го века нашей эры, распространились на северную Африку ( Arredi 2004 и Semino 2004 ). Однако Канарских островах неизвестно, чтобы на существовал какой-либо семитский язык. В Северной Африке J-M267 доминирует J-P58 и, согласно исследованиям, распределен очень неравномерно, часто, но не всегда, ниже среди берберского и/или негородского населения . В Эфиопии есть признаки более старых перемещений J-M267 в Африку через Красное море, причем не только в форме J-P58. Похоже, это также связано с семитскими языками. Согласно исследованию 2011 года, в Тунисе J-M267 значительно более распространен среди городского населения (31,3%), чем среди сельского населения (2,5%) ( Ennafaa 2011 ).
| Население | Размер выборки | Дж*(xJ-M172) | всего J-M267 | ДЖ-М267(xP58) | J-P58 | публикация | предыдущие исследования на тех же образцах |
|---|---|---|---|---|---|---|---|
| Алжир ( арабы из Орана ) | 102 | ЧТО | 22.5% | ЧТО | ЧТО | Робин, 2007 год. | |
| Алжир | 20 | ЧТО | 35% | ЧТО | ЧТО | Семино 2004 г. | |
| Египет | 147 | ЧТО | 21.1% | 1.4% | 19.7% | Кьярони 2009 | Луис 2004 г. |
| Египет | 124 | ЧТО | 19.8% | ЧТО | ЧТО | Эль-Сибай 2009 г. | |
| Египет ( Сива , Западная пустыня ) | 35 | ЧТО | 31.4% | ЧТО | ЧТО | Куянова 2009 г. | |
| Libya ( Tuareg ) | 47 | ЧТО | 0% | ЧТО | ЧТО | Латунь 2011 г. | |
| Ливия ( Бенгази ) | 238 | ЧТО | 39.5% | ЧТО | ЧТО | Альварес 2014 [ 7 ] | Эльмргни 2012 |
| Марокко ( арабы ) | 87 | ЧТО | 26.4% | ЧТО | ЧТО | Фадхлауи-Зид 2013 [ 8 ] | |
| Марокко ( арабы ) | 49 | ЧТО | 20.4% | ЧТО | ЧТО | Семино 2004 г. | |
| Марокко ( арабы ) | 28 | ЧТО | 60.7% | ЧТО | ЧТО | Андерхилл 2000 [ 9 ] | |
| Марокко ( арабы ) | 19 | ЧТО | 31.5% | ЧТО | ЧТО | Франкалаччи 2008 г. [ 10 ] | |
| Марокко ( берберы ) | 64 | ЧТО | 6.3% | ЧТО | ЧТО | Семино 2004 г. | |
| Марокко ( берберы ) | 103 | ЧТО | 10.7% | ЧТО | ЧТО | Семино 2004 г. | |
| Марокко ( Рабат ) | 267 | ЧТО | 21.3% | ЧТО | ЧТО | Альварес 2014 | Абухалид 2010 г. |
| Марокко ( Касабланка ) | 166 | ЧТО | 15.7% | ЧТО | ЧТО | Альварес 2014 | Лауина 2011 г. |
| Марокко ( Фигуиг ) | 96 | ЧТО | 29.2% | ЧТО | ЧТО | Альварес 2014 | Палитра 2010 |
| Марокко ( Эль-Джадида ) | 49 | ЧТО | 8.2% | ЧТО | ЧТО | Альварес 2014 | |
| Марокко ( Фес ) | 108 | ЧТО | 16.7% | 0.0% | 16.7% | Стрим 2015 | |
| Тунис | 73 | ЧТО | 34.2% | ЧТО | ЧТО | Семино 2004 г. | |
| Тунис | 601 | Уже | 16.64% | ЧТО | ЧТО | Пестано Дж. и др. (2013) [ 11 ] | |
| Тунис ( Сус ) | 220 | ЧТО | 25.9% | 0.0% | 25.9% | Фадхлауи-Зид 2015 [ 12 ] | |
| Тунис ( Тунис ) | 148 | ЧТО | 32.4% | 1.3% | 31.1% | Грюни 2012 | Мебель 2004 г. |
| Тунис | 52 | ЧТО | 34.6% | ЧТО | ЧТО | Онофри 2008 г. | |
| Тунис ( Бу Омран Берберс ) | 40 | ЧТО | 0% | ЧТО | ЧТО | Эннафаа 2011 | |
| Тунис (Бу Саад Берберс ) | 40 | ЧТО | 5% | 0% | 5% | Эннафаа 2011 | |
| Тунис ( жербские арабы ) | 46 | ЧТО | 8.7% | ЧТО | ЧТО | Эннафаа 2011 | |
| Тунис ( джербские берберы ) | 47 | ЧТО | 0% | ЧТО | ЧТО | Эннафаа 2011 | |
| Тунис ( сенед- берберы ) | 35 | ЧТО | 31.4% | 0% | 31.4% | Фадлауи-Зид 2011 г. | |
| Тунис ( Андалуси Загуан ) | 32 | ЧТО | 43.8% | 0% | 43.8% | Фадлауи-Зид 2011 г. | |
| Тунис (Cosmopolitan Tunis ) | 33 | ЧТО | 24.2 | 0% | 24.2% | Фадлауи-Зид 2011 г. | |
| Тунис ( Сеженан ) | 47 | ЧТО | 34.0% | ЧТО | ЧТО | Альварес 2014 | Выпуск 2011 г. |
| Тунис ( Сфакс ) | 56 | ЧТО | 25% | 0.0% | 25% | Стрим 2015 | |
| Тунис ( Бежа ) | 72 | ЧТО | 15.3% | 0.0% | 15.3% | Стрим 2015 | |
| Канарские острова (доиспаноязычные) | 30 | ЧТО | 16.7% | ЧТО | ЧТО | Фрегель 2009 | |
| Канарские острова (17-18 вв.) | 42 | ЧТО | 11.9% | ЧТО | ЧТО | Фрегель 2009 | |
| Канарские острова | 652 | ЧТО | 3.5% | ЧТО | ЧТО | Фрегель 2009 | |
| сахарцы | 89 | ЧТО | 20.2% | ЧТО | ЧТО | Фрегель 2009 | Бош 2001 г. и Флорес 2001 г. |
| Судан ( Хартум ) | 35 | ЧТО | 74.3% | 0.0% | 74.3% | Кьярони 2009 | Тофанелли 2009 г. и Хасан 2008 г. |
| Судан ( суданские арабы ) | 35 | ЧТО | 17.1% | 0.0% | 17.1% | Кьярони 2009 | Хасан 2008 г. |
| Судан ( нило-сахарцы ) | 61 | ЧТО | 4.9% | 3.3% | 1.6% | Кьярони 2009 | Хасан 2008 г. |
| Эфиопия ( английский ) | 78 | ЧТО | 2.6% | 2.6% | 0.0% | Кьярони 2009 | Семино 2004 г. |
| Эфиопия ( Амхара ) | 48 | ЧТО | 29.2% | 8.3% | 20.8% | Кьярони 2009 | Семино 2004 г. |
| Эфиопия ( Арси ) | 85 | 22% | ЧТО | ЧТО | ЧТО | Моран 2004 г. | |
| Эфиопия | 95 | 21% | ЧТО | ЧТО | ЧТО | Моран 2004 г. | |
| сомалийцы [1] | 201 | 0.5% | 2.5% | ЧТО | 2.5% | Санчес 2005 г. | J-P58 может составить 5% в предстоящем исследовании |
| Коморские острова | 293 | ЧТО | 5.0% | ЧТО | ЧТО | Помогите ему 2011 | |
| Южная Африка ( текст ) | 76 | ЧТО | 39.5% | 26.3% | 13.2% | Содиалл 2011 | |
| Зимбабве ( текст ) | 54 | ЧТО | 9.3% | 9.3% | ЧТО | Содиалл 2011 |
Азия
[ редактировать ]Южная Азия
[ редактировать ]J*(xJ-M172) был найден в Индии среди индийских мусульман. [ 13 ]
| Население | Размер выборки | Дж*(xJ-M172) | всего J-M267 | ДЖ-М267(xP58) | J-P58 | Публикация |
|---|---|---|---|---|---|---|
| Индия ( шииты ) | 161 | 10.6% | ЧТО | ЧТО | ЧТО | Иасвархант 2009 г. |
| Индия ( сунниты ) | 129 | 2.3% | ЧТО | ЧТО | ЧТО | Иасвархант 2009 г. |
| Индия (Маппла) | 40 | 10% | ЧТО | ЧТО | ЧТО | Иасвархант 2009 г. |
Западная Азия
[ редактировать ]Территория, включающая восточную Турцию, а также горы Загрос и Тавр, была определена как вероятная территория древнего разнообразия J-M267. Присутствуют как J-P58, так и другие типы J-M267, иногда с похожими частотами.
| Население | Размер выборки | Тотал J-M267 | ДЖ-М267(xP58) | J-P58 | Публикация | Предыдущие исследования на тех же образцах |
|---|---|---|---|---|---|---|
| Турция | 523 | 9.0% | 3.1% | 5.9% | Кьярони 2009 | Чинниоглу 2004 г. |
| Иран | 150 | 11.3% | 2.7% | 8.7% | Кьярони 2009 | Река 2006 |
| Иран ( Хузестан ) | ЧТО | 33.4% | ЧТО | ЧТО | Кивисилд 2012 [ 14 ] | |
| Ирак ( курды ) | 93 | 11.8% | 4.3% | 7.5% | Кьярони 2009 | |
| Ирак ( ассирийцы ) | 28 | 28.6% | 17.9% | 10.7% | Кьярони 2009 | |
| Ирак ( арабы ) | 56 | 64.1% | 1.8% | 62.3% | Кьярони 2009 | Тофанелли 2009 |
| Иран ( ассирийцы ) | 31 | 16.1% | 9.7% | 6.5% | Кьярони 2009 | |
| Иран | 92 | 3.2% | ЧТО | ЧТО | Эль-Сибай 2009 г. | |
| Турция ( ассирийцы ) | 25 | 20.0% | 16.0% | 4.0% | Кьярони 2009 |
Левантийское и семитское население
[ редактировать ]J-M267 очень распространен во всем этом регионе, где доминирует J-P58, но некоторые определенные подгруппы популяций имеют особенно низкие частоты.
| Население | Размер выборки | Тотал J-M267 | ДЖ-М267(xP58) | J-P58 | Публикация | Предыдущие исследования на тех же образцах |
|---|---|---|---|---|---|---|
| Сирия | 554 | 33.6% | ЧТО | ЧТО | Эль-Сибай 2009 г. | Заллуа 2008a |
| Сирия ( Джабель Друзы ) | 34 | 14.7% | 2.9% | 11.8% | Кьярони 2009 | |
| Сирия ( Хама -сунниты ) | 36 | 47.2% | 2.8% | 44.4% | Кьярони 2009 | |
| Сирия ( малула арамейцы ) | 44 | 6.8% | 4.5% | 2.3% | Кьярони 2009 | |
| Сирия ( Седная сирийско-католическая ) | 14 | 14.3% | 0.0% | 14.3% | Кьярони 2009 | |
| Сирия ( Дамаск сирийско-католический ) | 42 | 9.5% | 0.0% | 9.5% | Кьярони 2009 | |
| Сирия ( алавиты ) | 45 | 26.7% | 0.0% | 26.7% | Кьярони 2009 | |
| Сирия (северо-восточные ассирийцы ) | 30 | 3.3% | 0.0% | 3.3% | Кьярони 2009 | |
| Сирия ( Дамасские исмаилиты ) | 51 | 58.8% | 0.0% | 58.8% | Кьярони 2009 | |
| Ливан | 951 | 25% | ЧТО | ЧТО | Заллуа 2008a | |
| Галилейские друзы | 172 | 13.4% | 1.2% | 12.2% | Кьярони 2009 | Слэш 2008 |
| Палестинцы ( Акка ) | 101 | 39.2% | ЧТО | ЧТО | Заллуа 2008b | |
| палестинцы | 49 | 32.7% | 0.0% | 32.7% | Кьярони 2009 | |
| Иордания | 76 | 48.7% | 0.0% | 48.7% | Кьярони 2009 | |
| Иордания | 273 | 35.5% | ЧТО | ЧТО | Эль-Сибай 2009 г. | |
| Иордания ( Амман ) | 101 | 40.6% | ЧТО | ЧТО | Флорес 2005 г. | |
| Иордания ( Мёртвое море ) | 45 | 8.9% | ЧТО | ЧТО | Флорес 2005 г. | |
| Евреи ( Трас-уш-Монтес , Португалия ) | 57 | 12.3% | ЧТО | ЧТО | Ногейро 2009 г. | |
| Евреи ( коэны ) | 215 | 46.0% | 0.0% | 46.0% | Молот 2009 | |
| Евреи ( некоэны- ашкенази ) | 1,360 | 14.9% | 0.9% | 14.0% | Молот 2009 | |
| Бедуины ( Негев ) | 28 | 67.9% | 3.6% | 64.3% | Кьярони 2009 | Канн 2002 г. |
Аравийский полуостров
[ редактировать ]Эту статью необходимо обновить . ( январь 2018 г. ) |
J-P58 является наиболее распространенной гаплогруппой Y-хромосомы среди мужчин всего этого региона.
| Население | Размер выборки | Тотал J-M267 | ДЖ-М267(xP58) | J-P58 | Публикация | Предыдущие исследования на тех же образцах |
|---|---|---|---|---|---|---|
| Саудовская Аравия | 157 | 40.1% | ЧТО | ЧТО | Абу-Амеро 2009 г. | |
| Катар | 72 | 58.3% | 1.4% | 56.9% | Кьярони 2009 | Цепи 2008 |
| Объединенные Арабские Эмираты | 164 | 34.8% | 0.0% | 34.8% | Кьярони 2009 | Цепи 2008 |
| Йемен | 62 | 72.6% | 4.8% | 67.7% | Кьярони 2009 | Цепи 2008 |
| Кувейт | 117 | 45.2% | ЧТО | ЧТО | [ 15 ] | |
| Мой собственный | 121 | 38.0% | 0.8% | 37.2% | Кьярони 2009 | Луис 2004 г. |
Европа
[ редактировать ]J-M267 редко встречается в большей части Северной и Центральной Европы. Однако он встречается в значительных очагах на уровне 5–10% среди многих групп населения в южной Европе. Недавнее исследование существующей вариации пришло к выводу, что Кавказ, вероятно, является источником хромосом греческой и итальянской гаплогруппы J1-M267. [ 16 ]
Кавказ
[ редактировать ]На Кавказе есть области как с высокой, так и с низкой частотой J-M267. J-M267 на Кавказе также примечателен тем, что большая его часть не входит в субклад J-P58.
| Население | Размер выборки | Тотал J-M267 | ДЖ-М267(xP58) | J-P58 | Публикация |
|---|---|---|---|---|---|
| аварцы | 115 | 59% | 58% | 1% | Балановский 2011 г. |
| Даргинцы | 101 | 70% | 69% | 1% | Балановский 2011 г. |
| Кубачи | 65 | 99% | 99% | 0% | Балановский 2011 г. |
| Кайтак | 33 | 85% | 85% | 0% | Балановский 2011 г. |
| Лезгины | 81 | 44.4% | 44.4% | 0% | Балановский 2011 г. |
| Шапсуг | 100 | 0% | 0% | 0% | Балановский 2011 г. |
| Абхазский | 58 | 0% | 0% | 0% | Балановский 2011 г. |
| черкесы | 142 | 11.9% | 4.9% | 7% | Балановский 2011 г. |
| Ингушский | 143 | 2.8% | 2.8% | 0% | Балановский 2011 г. |
| осетины | 357 | 1.3% | 1.3% | 0.0% | Балановский 2011 г. |
| Чеченцы ( Ингушетия ) | 112 | 21% | 21% | 0% | Балановский 2011 г. |
| Чеченцы ( Чечня ) | 118 | 25% | 25% | 0% | Балановский 2011 г. |
| Чеченцы ( Дагестан ) | 100 | 16% | 16% | 0% | Балановский 2011 г. |
| Азербайджан | 46 | 15.2% | ЧТО | ЧТО | Ди Джакомо 2004 г. |
Распределение субкладов
[ редактировать ]J-P58
[ редактировать ]Маркер P58, который определяет подгруппу J1c3, был анонсирован в ( Karafet 2008 ), но ранее был анонсирован под названием Page08 в ( Repping 2006) и назван снова в Chiaroni 2009 . Он очень распространен во многих регионах, где распространен J-M267, особенно в некоторых частях Северной Африки и на всем Аравийском полуострове. Он также составляет примерно 70% J-M267 среди амхара в Эфиопии. Примечательно, что он не распространен среди J-M267 Кавказа.
Кьярони 2009 предположил, что J-P58 (который они называют J1e) мог впервые расселиться в период докерамического неолита B «из географической зоны, включая северо-восток Сирии, северный Ирак и восточную Турцию, в сторону Средиземноморской Анатолии, исмаилиты из южной Сирия, Иордания, Палестина и северный Египет». Они также предполагают, что Зарзиана материальная культура могла быть древней. Они также предполагают, что это перемещение людей также может быть связано с распространением семитских языков охотниками -скотоводами , которые переселялись в засушливые районы в периоды, когда, как известно, было мало осадков. Таким образом, в то время как другие гаплогруппы, включая J-M267, покинули этот район вместе с земледельцами, которые последовали за дождями, популяции, несущие J-M267, остались со своими стадами ( King 2002 и Chiaroni 2008 ).
Согласно этому сценарию, после первоначальной неолитической экспансии с участием семитских языков , которая, возможно, дошла до Йемена, более позднее распространение произошло во время энеолита или раннего бронзового века (приблизительно 3000–5000 гг. до н.э.), и это затронуло ветвь семитского языка, которая ведет к арабскому языку . Авторы предполагают, что это связано с распространением некоторого количества J-P58 со стороны Сирии в сторону арабского населения Аравийского полуострова и Негева .
С другой стороны, авторы согласны с тем, что более поздние волны рассеяния в этой области и вокруг нее также оказали сложное влияние на распространение некоторых типов J-P58 в некоторых регионах. Они перечисляют три региона, которые особенно важны для их предложения:
- Левант (Сирия, Иордания, Израиль и Палестина). В этой области Кьярони 2009 отмечает «неравномерное распределение частот J1c3 или J-P58», которое трудно интерпретировать и которое «может отражать сложную демографическую динамику религии и этнической принадлежности в регионе».
- Армянское нагорье, северный Ирак и западный Иран. В этой области Кьярони 2009 признает признаки того, что J-M267 мог иметь более древнее присутствие, и в целом они принимают доказательства, но отмечают, что они могут быть ошибочными.
- Южный район Омана, Йемена и Эфиопии. В этой области Кьярони 2009 признает аналогичные признаки, но отвергает их как возможный результат «либо изменчивости выборки, либо демографической сложности, связанной с несколькими основателями и множественными миграциями».
Кластер «YCAII=22-22 и DYS388≥15»
[ редактировать ]Исследования показывают, что группа J-P58 не только сама по себе доминирует во многих областях, где распространены J-M267 или J1, но также содержит большой кластер, который был обнаружен до открытия P58. Хотя это все еще предмет исследования.
Этот относительно молодой кластер, по сравнению с J-M267 в целом, был идентифицирован по гаплотипам маркеров STR - в частности, YCAII как 22-22 и DYS388, имеющих необычные значения повторов 15 или выше вместо более типичных 13 ( Chiaroni 2009 ). Этот кластер был обнаружен быть актуальным в некоторых широко разрекламированных исследованиях еврейского и палестинского населения ( Небель 2000 и Хаммер 2009 ). В более общем плане с тех пор было обнаружено, что этот кластер часто встречается среди мужчин на Ближнем Востоке и в Северной Африке, но реже в районах Эфиопии и Европы, где J-M267, тем не менее, распространен. Таким образом, генетический паттерн аналогичен паттерну J-P58 в целом, описанному выше, и может быть вызван теми же перемещениями/миграцией людей ( Chiaroni 2009 ).
Тофанелли 2009 называет этот общий кластер с YCAII = 22-22 и высокими значениями DYS388 «арабским» в отличие от « евразийского » типа J-M267. Этот арабский тип включает носителей арабского языка из Магриба , Судана , Ирака и Катара , и это относительно однородная группа, подразумевающая, что она могла рассеяться относительно недавно по сравнению с J-M267 в целом. Более разнообразная «евразийская» группа включает европейцев , курдов , иранцев и эфиопов (несмотря на то, что Эфиопия находится за пределами Евразии) и гораздо более разнообразна. Авторы также говорят, что «оманцы, как и ожидалось, демонстрируют смесь евразийских и типичных арабских гаплотипов, учитывая роль коридора, который в разное время играл Оманский залив в распространении азиатских и восточноафриканских генов ». Кьярони 2009 также отметил аномально высокий видимый возраст оманского J-M267, если рассматривать J-P58 и J-M267 в целом.
Этот кластер, в свою очередь, содержит три хорошо известных родственных подкластера. Во-первых, он содержит большую часть еврейского « модального гаплотипа Коэна », встречающегося среди еврейского населения, но особенно у мужчин с фамилиями, связанными с Коэном. Он также содержит « модальный гаплотип Галилеи» (GMH) и «модальный гаплотип палестинских и израильских арабов », оба из которых связаны с палестинскими / израильскими арабами Небелем 2000 и Хаммером 2009 . Затем Небель (2002) отметил, что GMH также является наиболее частым типом гаплотипа J-P209, обнаруженным у жителей северо-западной Африки и йеменцев, поэтому он не ограничивается Израилем и Палестиной. Однако этот конкретный вариант «отсутствует» у двух конкретных «неарабских популяций Ближнего Востока», а именно «евреев и курдов-мусульман» (хотя обе эти популяции действительно имеют высокий уровень J-P209). Небель 2002 отметил не только присутствие GMH в Магрибе, но и то, что J-M267 в этом регионе имел очень мало разнообразия. Они пришли к выводу, что J-M267 в этом регионе является результатом двух различных миграционных событий: «рассеяния раннего неолита» и «рассеивания в эпоху раннего неолита». экспансия с Аравийского полуострова «в течение 7-го века. Семино 2004 позже согласился, что это, по-видимому, согласуется с доказательствами, и сделал из этого обобщение, что распространение всего кластера YCAII = 22-22 J-M267 в арабоязычных районах Средней Азии Восточная и Северная Африка на самом деле могли иметь свое начало в исторические времена.
Более поздние исследования подчеркнули сомнение в том, что исламские расширения достаточно стары, чтобы полностью объяснить основные закономерности частот J-M267. Кьярони 2009 отверг это для J-P58 в целом, но признал, что «некоторые популяции с низким разнообразием, такие как бедуины из Израиля, Катара, Судана и ОАЭ, плотно сгруппированы вблизи высокочастотных гаплотипов, что предполагает эффекты основателя со звездным взрывом». экспансия в Аравийскую пустыню». Они не комментировали ситуацию в Магрибе.
Тофанелли 2009 занимает более сильную позицию, отвергая любую сильную корреляцию между арабской экспансией и либо субкластером, определенным STR YCAII=22-22, как обсуждалось Семино 2004 , либо меньшим «модальным гаплотипом Галилеи», как обсуждалось ( Небель 2002 ). Они также подсчитали, что модальному гаплотипу Коэна должно быть больше 4500 лет, а может быть и 8600 лет – задолго до предполагаемого происхождения коэнов. Только модальный вариант «палестинцы и израильские арабы» имел сильную корреляцию с этнической группой, но это также было редкостью. В заключение авторы отрицательно относятся к полезности модальных окон, определенных STR, для любых «судебно-медицинских или генеалогических целей», поскольку «они были обнаружены в этнических группах с различной культурной или географической принадлежностью».
Хаммер 2009 не согласился с этим, по крайней мере, в отношении модального гаплотипа Коэна . Они сказали, что необходимо рассмотреть более подробный гаплотип STR, чтобы определить новый «расширенный модальный гаплотип Коэна», который чрезвычайно редок за пределами еврейского населения, и даже внутри еврейского населения в основном встречается только у Коэнов . Они также заявили, что, используя больше маркеров и более строгое определение, предполагаемый возраст линии Коэним ниже, чем оценки Тофанелли 2009 , и согласуется с общим предком примерного времени основания священства, которым является источник фамилий Коэн.
Тофанелли и др. 2014 ответил: «В заключение, хотя наблюдаемое распределение субкладов гаплотипов в нерекомбинантных геномах митохондрий и Y-хромосомы может быть совместимо с событиями-основателями недавнего времени, приведшими к возникновению еврейских групп, таких как коэниты, левиты, ашкеназиты, общий существенный полифилетизм, а также их систематическое появление в нееврейских группах подчеркивают отсутствие поддержки их использования как маркеров еврейского происхождения или Библейские сказки». [ 18 ]
J-M368
[ редактировать ]Соответствие между P58 и высокими значениями DYS388 и YCAII=22-22 не является идеальным. Например, субклад J-M267 J-P58, определенный SNP M368, имеет DYS388=13 и YCAII=19-22, как и другие типы J-M267 за пределами «арабского» типа J-M267, и поэтому считается, что он быть относительно старым ответвлением J-P58, которое не принимало участия в последних волнах экспансии J-M267 на Ближнем Востоке ( Кьярони 2009 ). Эти гаплотипы DYS388=13 наиболее распространены на Кавказе и в Анатолии , но также встречаются и в Эфиопии ( Tofanelli 2009 ).
Филогенетика и распространение
[ редактировать ]Существует несколько подтвержденных и предложенных филогенетических деревьев гаплогруппы J-M267. Следующая филогения или генеалогическое древо субкладов гаплогруппы J-M267 основана на дереве ISOGG (2012), которое, в свою очередь, основано на дереве YCC 2008 и последующих опубликованных исследованиях.
J1 (Л255, Л321, М267)
- Кластеры J1* J1* встречаются на Армянском нагорье и в некоторых частях Кавказа.
- J1a (M62) Встречается очень редко в Европе.
- J1b (M365.1) Встречается с низкой частотой на Армянском нагорье, в Иране, Катаре и некоторых частях Европы.
- J1c (L136)
- J1c* Встречается с низкой частотой в Европе.
- J1c1 (M390)
- J1c2 (P56) Встречается спорадически в Анатолии, Восточной Африке, на Аравийском полуострове и в Европе.
- J1c3
- J1c3* Встречается с низкой частотой в Леванте и на Аравийском полуострове.
- J1c3a (M367.1, M368.1) Ранее известный как J1e1.
- J1c3b (M369) Ранее известный как J1e2.
- J1c3c (L92, L93) Встречается с низкой частотой в Южной Аравии.
- J1c3d (L147.1) Составляет большую часть J1, преобладающей гаплогруппы на Аравийском полуострове.
- J1c3d* Составляет большинство J1 в Йемене, евреи Коэна (как раввины , так и караимы ), [ 19 ] но отсутствует среди курайшитов, включая Шарифа из Мекки из клана Бану Хашем.
- J1c3d1 (L174.1)
- J1c3d2 (L222.2) Составляет большую часть J1c3d в Саудовской Аравии. Важный элемент J1c3d в Северной Африке.
- J1c3d2*
- J1c3d2a (L65.2/S159.2)
- J1c3d2*
Древняя ДНК
[ редактировать ]Алалах аморейский город-государство
[ редактировать ]Пять из 12 особей мужского пола из Алалаха , живших между 1930-1325 гг. до н.э., принадлежали к гаплогруппе J1-P58. [ 20 ] [ 21 ]
Арслантепе археологический комплекс
[ редактировать ]Один из 18 мужчин из Арслантепе, живших ок. 3491-3122 гг. до н.э., принадлежал к гаплогруппе J1-Z1824. [ 22 ] [ 23 ]
Древний город Эбла
[ редактировать ]Трое из 6 особей Эблы , живших между 2565-1896 гг. до н.э., принадлежали к J1-P58. [ 24 ] [ 25 ] Эбла была древним восточно-семитским городом и царством в Сирии в раннем бронзовом веке , которое было разрушено аккадцами .
Карелия
[ редактировать ]Член гаплогруппы J1-M267 обнаружен среди восточных охотников-собирателей из Карелии , Северо-Восточная Европа , живших примерно 8,3 тыс. лет назад. Эта ветвь отсутствует у других древнеевропейских охотников-собирателей. К сожалению, невозможно поместить этот образец в контекст текущей вариации гаплогруппы J1-M267 из-за низкого качества последовательности ДНК. [ 3 ]
Сардиния
[ редактировать ]Оливьери и др . обнаружили гаплотип J1c3 в одном из своих древних образцов с Сардинии, датированный 6190–6000 кал. [ 26 ]
Сацурбля
[ редактировать ]Древний образец J1 был найден в пещере Сацурблиа около 11 000 г. до н. э. и принадлежит к редкому субкладу J1-FT34521. [ 27 ] Древний человек из Сацурблии был мужчиной с черными волосами, карими глазами и светлой кожей.
Скажи Волку
[ редактировать ]Один из четырех особей мужского пола из Телль-Курду, живших около 5706-5622 гг. до н.э., принадлежал к J1-L620. [ 28 ] [ 29 ]
См. также
[ редактировать ]Генетика
[ редактировать ]- Генетическая история Ближнего Востока
- Генетическая история Европы
- Таблица перевода гаплогрупп Y-хромосомы
- Генетическая генеалогия
- Гаплогруппа
- Гаплотип
- Гаплогруппа ДНК Y-хромосомы человека
- Молекулярная филогенетика
- Парагруппа
- Субклад
- Y-хромосома Аарон
- Гаплогруппы Y-хромосомы в популяциях мира
- Гаплогруппы Y-ДНК в населении Европы
- Гаплогруппы Y-ДНК в популяциях Восточной и Юго-Восточной Азии
- Гаплогруппы Y-ДНК у населения Ближнего Востока
- Гаплогруппы Y-ДНК у населения Северной Африки
- Гаплогруппы Y-ДНК у населения Кавказа
- Гаплогруппы Y-ДНК по этническим группам
Субклады Y-ДНК J
[ редактировать ]Основополагающее дерево Y-ДНК
[ редактировать ]Ссылки
[ редактировать ]- ^ Сингх С; Сингх А; Раджкумар Р; Сампат Кумар К; Кадаркарай Сами С; Низамуддин С; и др. (2016). «Анализ влияния неолитической демической диффузии на индийский пул Y-хромосом через гаплогруппу J2-M172» . Научные отчеты . 6 : 19157. Бибкод : 2016NatSR...619157S . дои : 10.1038/srep19157 . ПМК 4709632 . ПМИД 26754573 .
- ^ Дж1
- ^ Перейти обратно: а б с Саакян, Оганес; Маргарян, Ашот; Сааг, Лаури; Кармин, Моника; Флорес, Родриго; Хабер, Марк; Кушнеревич Алена; Хачатрян, Заруи; Бахманимер, Ардешир; Парик, Юрий; Карафет, Татьяна; Юнусбаев, Баязит; Райсберг, Туули; Сольник, Ану; Мецпалу, Эне (23 марта 2021 г.). «Происхождение и распространение гаплогруппы J1-M267 Y-хромосомы человека» . Научные отчеты . 11 (1): 6659. Бибкод : 2021NatSR..11.6659S . дои : 10.1038/s41598-021-85883-2 . ISSN 2045-2322 . ПМЦ 7987999 . ПМИД 33758277 .
- ^ Ребай, Ахмед. «Синтетический обзор генетического родства между Северной Африкой и Аравией, выведенный на основе распределения по отцовской линии» .
- ^ Ребай, Ахмед. «Синтетический обзор генетического родства между Северной Африкой и Аравией, выведенный на основе распределения по отцовской линии» .
- ^ Кьярони, Жак; Кинг, Рой Дж.; Майрес, Натали М.; Хенн, Бренна М.; Дюкурно, Аксель; Митчелл, Майкл Дж.; Боетч, Жиль; Шейха, Исса; Лин, Алиса А.; Ник-Ахд, Махнуш; Ахмад, Джабин; Латтанци, Франческа; Эррера, Рене Дж.; Ибрагим, Мунтасер Э.; Броуди, Аарон; Семино, Орнелла; Кивисилд, Тоомас; Андерхилл, Питер А. (2010). «Появление гаплогруппы J1e Y-хромосомы среди арабоязычного населения» . Европейский журнал генетики человека . 18 (3): 348–353. дои : 10.1038/ejhg.2009.166 . ПМЦ 2987219 . ПМИД 19826455 .
- ^ Альварес, Луис; Цирия, Эстела; Маркес, София Л.; Сантос, Кристина; Алуя, Мария Пилар (2014). «Анализ Y-хромосомы у населения Северо-Западной Иберии: выяснение влияния североафриканских линий». Американский журнал биологии человека . 26 (6): 740–746. дои : 10.1002/ajhb.22602 . ПМИД 25123837 . S2CID 205303372 .
- ^ Фадхлауи-Зид, Карима; Хабер, Марк; Мартинес-Крус, Бегонья; Заллуа, Питер; Бенаммар Эльгаайед, Амель; Комас, Дэвид (27 ноября 2013 г.). «Общегеномное и отцовское разнообразие, раскрывающее недавнее происхождение человеческих популяций в Северной Африке» . ПЛОС ОДИН 8 (11):e8 Бибкод : 2013PLoSO... 880293F дои : 10.1371/journal.pone.0080293 . ISSN 1932-6203 . ПМЦ 3842387 . ПМИД 24312208 .
- ^ Андерхилл, Питер А. (декабрь 2000 г.). «Вариации последовательности Y-хромосомы и история человеческих популяций» . Природная генетика . 26 (3): 360. дои : 10.1038/81685 . ПМИД 11062480 . S2CID 12893406 .
- ^ Франкалаччи, Паоло (2008). «История и география Y-хромосомы человека в Европе: взгляд на SNP» (PDF) . Журнал антропологических наук . 86 : 59–89. ПМИД 19934469 . Архивировано из оригинала (PDF) 28 марта 2012 года . Проверено 9 октября 2022 г.
- ^ Бекада, Асмахан; Фрегель, Роза; Кабрера, Винсент М.; Ларруга, Джозеф М.; Пестано, Джозеф; Бенхамамуш, Сорая; Гонсалес, Ханна М. (19 февраля 2013 г.). «Введение алжирской митохондриальной ДНК и профилей Y-хромосомы в ландшафт Северной Африки» . ПЛОС ОДИН 8 (2): e56775. Бибкод : 2013PLoSO... 856775B дои : 10.1371/journal.pone.0056775 . ISSN 1932-6203 . ПМЦ 3576335 . ПМИД 23431392 .
- ^ Фадлауи-Зид, Карима (2014). «Сус: крайняя генетическая гетерогенность в Северной Африке» . Журнал генетики человека . 60 (1): 41–49. дои : 10.1038/jhg.2014.99 . ПМИД 25471516 . S2CID 25186140 .
- ^ Иасвархант, М; Хак, я; Равеш, З; и др. (март 2010 г.). «Следы происхождения к югу от Сахары и Ближнего Востока в мусульманском населении Индии» . Европейский журнал генетики человека . 18 (3): 354–63. дои : 10.1038/ejhg.2009.168 . ПМЦ 2859343 . ПМИД 19809480 .
- ^ Грантс, Виола; Батталья, Винченца; Хушиар Кашани, Бахарак; Пароло, Сильвия; Аль-Захери, Надя; Ахилли, Алессандро; Оливьери, Анна; Гандини, Франческа; Хаушманд, Масуд; Санати, Мохаммад Хосейн; Торрони, Антонио; Семино, Орнелла (18 июля 2012 г.). «Древние миграционные события на Ближнем Востоке: новые данные по Y-хромосомной вариации современных иранцев» . ПЛОС ОДИН . 7 (7): е41252. Бибкод : 2012PLoSO...741252G . дои : 10.1371/journal.pone.0041252 . ISSN 1932-6203 . ПМЦ 3399854 . ПМИД 22815981 .
- ^ Трик-Фендри, Сумайя; Санчес-Дис, Паула; Кинг-Гонсалес, Данель; Альфадли, Суад; Аяди, Имен; Бен Марзуг, Эр-Рияд; Карраседо, Анхель; Ребай, Ахмед (4 марта 2016 г.). «Генетическая структура населения Кувейта, выявленная по отцовской линии: генетическая структура Кувейта». Американский журнал биологии человека . 28 (2): 203–212. дои : 10.1002/ajhb.22773 .
- ^ Финоккио, Андреа; Тромбетта, Бениамино; Мессина, Франческо; Д'Атанасио, Евгения; Акар, Неджат; Лутрадис, Афродита; Михалодимитракис, Эммануэль И.; Кручиани, Фульвио; Новеллетто, Андреа (10 мая 2018 г.). «Точно разрешенная филогения Y-хромосомы Hg J освещает процессы финикийских и греческих колонизаций в Средиземноморье» . Научные отчеты . 8 (1): 7465. Бибкод : 2018NatSR...8.7465F . дои : 10.1038/s41598-018-25912-9 . ISSN 2045-2322 . ПМЦ 5945646 . ПМИД 29748665 .
- ^ Эль-Сибай и др., 2009, Процент гаплогрупп.
- ^ Тофанелли, Серджио (10 ноября 2014 г.). «Мотивы гаплотипов митохондрий и Y-хромосомы как диагностические маркеры еврейского происхождения: пересмотр» . Границы генетики . 5 : 384. дои : 10.3389/fgene.2014.00384 . ПМЦ 4229899 . ПМИД 25431579 .
- ^ Брук, Кевин А. (лето 2014 г.). «Генетика крымских караимов» (PDF) . Карадениз Араштырмалары (Журнал черноморских исследований) . 11 (42): 69–84 на стр. 83. doi : 10.12787/KARAM859 .
- ^ «J-P58 YДерево» .
- ^ Скуртаниоти, Эйрини; Эрдал, Йылмаз С.; Франжипан, Марселла; Балосси Рестелли, Франческа; Йенер, К. Аслихан; Пиннок, Фрэнсис; Мэтью, Пол; Узбал, Рана; Шуп, Вольф-Дитрих; Гулиев, Фархад; Ахундов, Туфан; Лайонс, Бертиль; Хаммер, Эмили Л.; Ньюджент, Селин Э.; Бурри, Марта; Нойманн, Гуннар У.; Пенске, Сандра; Ингман, Тара; Акар, Мурат; Шафик, Рула; Палумби, Джулио; Эйзенманн, Стефани; д'Андреа, Марта; Рорлах, Адам Б.; Вариннер, Кристина; Чон, Чунгвон; Стокхаммер, Филип В.; Хаак, Вольфганг; Краузе, Джон (2020). «Геномная история от неолита до бронзового века в Анатолии, Северном Леванте и Южном Кавказе» . Ячейка 181 (5):1158–1175.e28. дои : 10.1016/j.cell.2020.04.044 . hdl : 20.500.12154/1254 . ПМИД 32470401 . S2CID 219105572 .
- ^ "J-Z1842 YДерево" .
- ^ Скуртаниоти, Эйрини; Эрдал, Йылмаз С.; Франжипан, Марселла; Балосси Рестелли, Франческа; Йенер, К. Аслихан; Пиннок, Фрэнсис; Мэтью, Пол; Узбал, Рана; Шуп, Вольф-Дитрих; Гулиев, Фархад; Ахундов, Туфан; Лайонс, Бертиль; Хаммер, Эмили Л.; Ньюджент, Селин Э.; Бурри, Марта; Нойманн, Гуннар У.; Пенске, Сандра; Ингман, Тара; Акар, Мурат; Шафик, Рула; Палумби, Джулио; Эйзенманн, Стефани; д'Андреа, Марта; Рорлах, Адам Б.; Вариннер, Кристина; Чон, Чунгвон; Стокхаммер, Филип В.; Хаак, Вольфганг; Краузе, Джон (2020). «Геномная история от неолита до бронзового века в Анатолии, Северном Леванте и Южном Кавказе» . Ячейка 181 (5):1158–1175.e28. дои : 10.1016/j.cell.2020.04.044 . hdl : 20.500.12154/1254 . ПМИД 32470401 . S2CID 219105572 .
- ^ «J-P58 YДерево» .
- ^ Скуртаниоти, Эйрини; Эрдал, Йылмаз С.; Франжипан, Марселла; Балосси Рестелли, Франческа; Йенер, К. Аслихан; Пиннок, Фрэнсис; Мэтью, Пол; Узбал, Рана; Шуп, Вольф-Дитрих; Гулиев, Фархад; Ахундов, Туфан; Лайонс, Бертиль; Хаммер, Эмили Л.; Ньюджент, Селин Э.; Бурри, Марта; Нойманн, Гуннар У.; Пенске, Сандра; Ингман, Тара; Акар, Мурат; Шафик, Рула; Палумби, Джулио; Эйзенманн, Стефани; д'Андреа, Марта; Рорлах, Адам Б.; Вариннер, Кристина; Чон, Чунгвон; Стокхаммер, Филип В.; Хаак, Вольфганг; Краузе, Джон (2020). «Геномная история от неолита до бронзового века в Анатолии, Северном Леванте и Южном Кавказе» . Ячейка 181 (5):1158–1175.e28. дои : 10.1016/j.cell.2020.04.044 . hdl : 20.500.12154/1254 . ПМИД 32470401 . S2CID 219105572 .
- ^ Оливьери, А.; Сидоре, К.; Ахиллес, А.; Ангиус, А.; Пост, К.; Фуртвенглер, А.; Брандини, С.; Розарио Каподиферро, М.; Гандини, Ф.; Золедзевска, М.; Питцалис, М.; Мале, А.; Бусонеро, Ф.; Лай, Л.; Скитс, Р. (01 мая 2017 г.). «Митогеномное разнообразие у сардинцев: генетическое окно в прошлое острова» . Молекулярная биология и эволюция . 34 (5): 1230–1239. дои : 10.1093/molbev/msx082 . ISSN 0737-4038 . ПМК 5400395 . ПМИД 28177087 .
- ^ «J-Y6313 YДерево» .
- ^ «J-L620 YДерево» .
- ^ Скуртаниоти, Эйрини; Эрдал, Йылмаз С.; Франжипан, Марселла; Балосси Рестелли, Франческа; Йенер, К. Аслихан; Пиннок, Фрэнсис; Мэтью, Пол; Узбал, Рана; Шуп, Вольф-Дитрих; Гулиев, Фархад; Ахундов, Туфан; Лайонс, Бертиль; Хаммер, Эмили Л.; Ньюджент, Селин Э.; Бурри, Марта; Нойманн, Гуннар У.; Пенске, Сандра; Ингман, Тара; Акар, Мурат; Шафик, Рула; Палумби, Джулио; Эйзенманн, Стефани; д'Андреа, Марта; Рорлах, Адам Б.; Вариннер, Кристина; Чон, Чунгвон; Стокхаммер, Филип В.; Хаак, Вольфганг; Краузе, Джон (2020). «Геномная история от неолита до бронзового века в Анатолии, Северном Леванте и Южном Кавказе» . Ячейка 181 (5):1158–1175.e28. дои : 10.1016/j.cell.2020.04.044 . hdl : 20.500.12154/1254 . ПМИД 32470401 . S2CID 219105572 .
Сноски
[ редактировать ]Цитируемые работы
[ редактировать ]Журналы
[ редактировать ]- Моран, Китай; Скотт Р.А.; Адамс С.М.; Уоррингтон С.Дж.; Джоблинг М.А.; Уилсон Р.Х.; Гудвин WH; Георгиадес Э; Вольде Б; Питсиладис Ю.П. (ноябрь 2004 г.). «Гаплогруппы Y-хромосомы элитных эфиопских бегунов на выносливость». Генетика человека . 115 (6): 492–7. дои : 10.1007/s00439-004-1202-y . ПМИД 15503146 . S2CID 13960753 .
- Абу-Амеро, Халед К.; Хеллани, Али; Гонсалес, Ана М; Ларруга, Хосе М; Кабрера, Висенте М; Андерхилл, Питер А. (2009). «Разнообразие Y-хромосомы Саудовской Аравии и его связь с близлежащими регионами» . БМК Генетика . 10 (1): 59. дои : 10.1186/1471-2156-10-59 . ПМК 2759955 . ПМИД 19772609 .
- Альварес, Луис; Сантос, Кристина; Монтьель, Рафаэль; Каейро, Бласкес; Баали, Абдельлатиф; Дюгужона, Жан-Мишель; Алуя, Мария Пилар (2009). «Вариации Y-хромосомы в Южной Иберии: понимание вклада Северной Африки» . Американский журнал биологии человека . 21 (3): 407–9. дои : 10.1002/ajhb.20888 . ПМИД 19213004 . S2CID 7041905 .
- Арреди, Б; Полони, Э; Параккини, С; Зержал, Т; Фаталлах, Д; Макрелуф, М; Паскали, В; Новеллетто, А; Тайлер-Смит, К. (2004). «Преимущественно неолитическое происхождение вариаций Y-хромосомной ДНК в Северной Африке» . Американский журнал генетики человека . 75 (2): 338–45. дои : 10.1086/423147 . ПМК 1216069 . ПМИД 15202071 .
- Балановский О.; Дибирова, К.; Дыбо, А.; Мудрак, О.; Фролова С.; Почешхова Е.; Хабер, М.; Платт, Д.; и др. (2011). «Параллельная эволюция генов и языков в Кавказском регионе» . Молекулярная биология и эволюция . 28 (10): 2905–20. дои : 10.1093/molbev/msr126 . ПМЦ 3355373 . ПМИД 21571925 .
- Батталья, Винченца; Форнарино, Симона; Аль-Захери, Надя; Оливьери, Анна; Лопата, Мэри; Майрес, Натали М; Кинг, Рой Дж; Корни, Сирия; и др. (2009). «Y-хромосомные свидетельства культурного распространения сельского хозяйства в Юго-Восточной Европе» . Европейский журнал генетики человека . 17 (6): 820–30. дои : 10.1038/ejhg.2008.249 . ПМК 2947100 . ПМИД 19107149 .
- Босх, Елена; Калафель, Франческ; Комас, Дэвид; Офнер, Питер Дж.; Андерхилл, Питер А.; Бертранпети, Жауме (апрель 2001 г.). «Анализ вариаций Y-хромосомы человека с высоким разрешением показывает резкий разрыв и ограниченный поток генов между Северо-Западной Африкой и Пиренейским полуостровом» . Американский журнал генетики человека . 68 (4): 1019–1029. дои : 10.1086/319521 . ПМЦ 1275654 . ПМИД 11254456 .
- Каденас, Алисия М; Животовский Лев А; Кавалли-Сфорца, Лука Л; Андерхилл, Питер А; Эррера, Рене Дж (2008). «Разнообразие Y-хромосом характеризует Оманский залив» . Европейский журнал генетики человека . 16 (3): 374–86. дои : 10.1038/sj.ejhg.5201934 . ПМИД 17928816 .
- Канн, ХМ (2002). «Панель клеточных линий разнообразия генома человека». Наука . 296 (5566): 261–262. дои : 10.1126/science.296.5566.261b . ПМИД 11954565 . S2CID 41595131 .
- Волосы, Кристиан; Онофри, Валерио; Бризигелли, Франческа; Боски, Илария; Скарниччи, Франческа; Масулло, Мара; Ферри, Джанмарко; Тофанелли, Серджио; и др. (2009). «Мавры и сарацины в Европе: оценка средневекового мужского наследия Северной Африки в южной Европе» . Европейский журнал генетики человека . 17 (6): 848–52. дои : 10.1038/ejhg.2008.258 . ПМЦ 2947089 . ПМИД 19156170 .
- Кьярони, Дж; Кинг, Рой Дж; Андерхилл, Питер А. (2008). «Корреляция годовых осадков с разнообразием Y-хромосомы человека и появлением неолитической сельскохозяйственной и скотоводческой экономики в Плодородном полумесяце». Античность . 82 (316): 281–289. дои : 10.1017/s0003598x00096800 . S2CID 163297133 .
- Кьярони, Жак; Кинг, Рой Дж; Майрес, Натали М; Хенн, Бренна М; Дюкурно, Аксель; Митчелл, Майкл Дж; Боетч, Жиль; Шейха, Исса; и др. (2010). «Появление гаплогруппы J1e Y-хромосомы среди арабоязычного населения» . Европейский журнал генетики человека . 18 (3): 348–353. дои : 10.1038/ejhg.2009.166 . ПМЦ 2987219 . ПМИД 19826455 .
- Чинниоглу, Ченгиз; Кинг, Рой; Кивисилд, Тоомас; Калфоглу, Эрси; Атасой, Севиль; Каваллери, Джанпьеро Л; Лилли, Анита С.; Роузман, Чарльз С; и др. (2004). «Раскопки слоев гаплотипов Y-хромосомы в Анатолии». Генетика человека . 114 (2): 127–148. дои : 10.1007/s00439-003-1031-4 . ПМИД 14586639 . S2CID 10763736 .
- Ди Гаэтано, Корнелия; Черутти, Николетта; Кробу, Франческа; Робино, Карло; Интурри, Серена; Джино, Сара; Гуаррера, Симонетта; Андерхилл, Питер А; и др. (2009). «Дифференциальная миграция греков и северной Африки на Сицилию подтверждается генетическими данными Y-хромосомы» . Европейский журнал генетики человека . 17 (1): 91–99. дои : 10.1038/ejhg.2008.120 . ПМЦ 2985948 . ПМИД 18685561 .
- Эль-Сибай, Мирват; Платт, Дэниел Э.; Хабер, Марк; Сюэ, Яли; Юханна, Соня С.; Уэллс, Р. Спенсер; Изаабель, Хасан; Санюра, Мэй Ф.; и др. (2009). «Географическая структура Y-хромосомного генетического ландшафта Леванта: контраст между побережьем и сушей» . Анналы генетики человека . 73 (Часть 6): 568–581. дои : 10.1111/j.1469-1809.2009.00538.x . ПМЦ 3312577 . ПМИД 19686289 .
- Эннафаа, Хаджер; Фрегель, Роза; Ходжет-эль-Хиль, Хусейн; Гонсалес, Ана М; Махмуди, Хеджер Абдалла Эль; Кабрера, Висенте М; Ларруга, Джозеф М; Бенаммар-Эльгаид, Амель (2011). «Митохондриальная ДНК и микроструктура Y-хромосомы в Тунисе» . Журнал генетики человека . 56 (10): 734–741. дои : 10.1038/jhg.2011.92 . ПМИД 21833004 .
- Фадхлауи-Зид, Карима; Мартинес-Крус, Бегона; Ходжет-эль-Хиль, Хусейн; Мендисабаль, Изабель; Бенаммар-Эльгаайед, Амель; Комас, Дэвид (2011). «Генетическая структура тунисских этнических групп, выявленная по отцовской линии». Американский журнал физической антропологии . 146 (2): 271–280. дои : 10.1002/ajpa.21581 . ПМИД 21915847 .
- Флорес, Карлос; Мака-Мейер, Николь; Пешез, Хосе А.; Кабрера, Висенте М. (сентябрь 2001 г.). «Население Канарских островов: взгляд на микросателлитный гаплотип CD4/Alu». Иммунология человека . 62 (9): 949–953. дои : 10.1016/S0198-8859(01)00311-1 . ПМИД 11543897 .
- Флорес, Карлос; Мака-Мейер, Николь; Ларруга, Хосе М.; Кабрера, Висенте М.; Карадшех, Наиф; Гонсалес, Ана М. (2005). «Изоляты в коридоре миграций: анализ вариаций Y-хромосомы в Иордании с высоким разрешением» . Журнал генетики человека . 50 (9): 435–441. дои : 10.1007/s10038-005-0274-4 . ПМИД 16142507 .
- Фрегель, Роза; ГОМЕС, Вероника; ГУСМИО, Леонор; Гонсалес, Ана М; Кабрера, Висенте М; АМОРИМ, Антонио; Ларруга, Хосе М. (2009). «Демографическая история мужского генофонда Канарских островов: замена местных линий Европой» . Эволюционная биология BMC . 9 (1): 181. дои : 10.1186/1471-2148-9-181 . ПМЦ 2728732 . ПМИД 19650893 .
- Ди Джакомо, Ф.; Лука, Ф.; Попа, Ло; Акар, Н.; Ананью, Н.; Банико Дж.; Брдичка, Р.; Барбуджани, Г.; и др. (2004). «Y-хромосомная гаплогруппа J как признак постнеолитической колонизации Европы». Генетика человека . 115 (5): 357–371. дои : 10.1007/s00439-004-1168-9 . ПМИД 15322918 . S2CID 18482536 .
- Гонсалвес, Рита; Фрейтас, Ана; Бранко, Марта; РОЗА, Александра; ФЕРНАНДЕС, Ана Т.; Животовский Лев А.; Андерхилл, Питер А.; Кивисилд, Тоомас; Брем, Антонио (2005). «Линии Y-хромосомы из Португалии, Мадейры и Азорских островов фиксируют элементы сефардского и берберского происхождения». Анналы генетики человека . 69 (Часть 4): 443–454. дои : 10.1111/j.1529-8817.2005.00161.x . hdl : 10400.13/3018 . ПМИД 15996172 . S2CID 3229760 .
- Хаммер, Майкл Ф.; Бехар, Дорон М.; Карафет Татьяна М.; Мендес, Фернандо Л.; Холлмарк, Брайан; Эрез, Тамар; Животовский Лев А.; Россе, Сахарон; Скорецкий, Карл (2009). «Расширенные гаплотипы Y-хромосомы определяют многочисленные и уникальные линии еврейского духовенства» . Генетика человека . 126 (5): 707–717. дои : 10.1007/s00439-009-0727-5 . ПМЦ 2771134 . ПМИД 19669163 .
- Хасан, Хишам Ю.; Андерхилл, Питер А.; Кавалли-Сфорца, Лука Л.; Ибрагим, Мунтасер Э. (2008). «Вариации Y-хромосомы у суданцев: ограниченный поток генов, соответствие языку, географии и истории». Американский журнал физической антропологии . 137 (3): 316–323. дои : 10.1002/ajpa.20876 . ПМИД 18618658 .
- Карачанак, Сена; Грантс, Виола; Форнарино, Симона; Нешева, Десислава; Аль-Захери, Надя; Батталья, Винченца; Каросса, Валерия; Йорданов, Йордан; Торрони, Антонио; Галабов, Ангел С.; Тончева, Драга; Семино, Орнелла (6 марта 2013 г.). «Разнообразие Y-хромосомы у современных болгар: новые сведения об их происхождении» . ПЛОС ОДИН . 8 (3): e56779. Бибкод : 2013PLoSO...856779K . дои : 10.1371/journal.pone.0056779 . ПМК 3590186 . ПМИД 23483890 .
- Карафет, ТМ; Мендес, Флорида; Мейлерман, МБ; Андерхилл, Питер А.; Зегура, СЛ; Хаммер, МФ (2008). «Новые бинарные полиморфизмы изменяют форму и увеличивают разрешение дерева гаплогруппы Y-хромосомы человека» . Геномные исследования . 18 (5): 830–838. дои : 10.1101/гр.7172008 . ПМК 2336805 . ПМИД 18385274 . См. также «Дополнительный материал».
- Кинг, Р; Андерхилл, Питер А. (2002). «Согласованное распределение расписной керамики и керамических фигурок эпохи неолита с линиями Y-хромосомы» . Античность . 76 (293): 707–714. дои : 10.1017/s0003598x00091158 . S2CID 160359661 .
- Кинг, Р.Дж.; Озджан, СС; Картер, Т.; Калфоглу, Э.; Атасой, С.; Триантафиллидис, К.; Коваци, А.; Лин, А.А.; и др. (2008). «Дифференциальное анатолийское влияние Y-хромосомы на греческий и критский неолит». Анналы генетики человека . 72 (Часть 2): 205–214. дои : 10.1111/j.1469-1809.2007.00414.x . ПМИД 18269686 . S2CID 22406638 .
- Кинг, Рой Дж; Ди Кристофаро, Джули; Куваци, Анастасия; Триантафиллидис, Костас; Шайдель, Уолтер; Майрес, Натали М; Лин, Алиса А; Эйсотье, Александр; Митчелл, Майкл; Биндер, Дидье; Семино, Орнелла; Новеллетто, Андреа; Андерхилл, Питер А; Кьярони, Жак (декабрь 2011 г.). «Приход греков в Прованс и Корсику: модели Y-хромосомы архаической греческой колонизации западного Средиземноморья» . Эволюционная биология BMC . 11 (1): 69. дои : 10.1186/1471-2148-11-69 . ПМК 3068964 . ПМИД 21401952 .
- Куянова, Мартина; Перейра, Луиза; ФЕРНАНДЕС, Вероника; Перейра, Джоана Б.; Черный, Виктор (2009). «Генетический вклад ближневосточного неолита в небольшой оазис Западной египетской пустыни». Американский журнал физической антропологии . 140 (2): 336–46. дои : 10.1002/ajpa.21078 . ПМИД 19425100 .
- Луис, Дж; Роуолд, Д; Регейро, М; Каейро, Б; Чинниоглу, К; Роузман, К; Андерхилл, П; Кавалли-Сфорца, Л; Эррера, Р. (2004). «Левант против Африканского Рога: свидетельства двунаправленных коридоров миграции людей» . Американский журнал генетики человека . 74 (3): 532–544. дои : 10.1086/382286 . ПМЦ 1182266 . ПМИД 14973781 . . (Также см. Ошибки )
- Мсаиди, Саид; Дюкурно, Аксель; Боетч, Жиль; Лонгепье, Гай; Папа, Кассим; Аллибер, Клод; Яхая, Али Ахмед; Кьярони, Жак; Митчелл, Майкл Дж (2011). «Генетическое разнообразие Коморских островов показывает, что раннее мореплавание стало основным фактором, определяющим биокультурную эволюцию человека в западной части Индийского океана» . Европейский журнал генетики человека . 19 (1): 89–94. дои : 10.1038/ejhg.2010.128 . ПМК 3039498 . ПМИД 20700146 .
- Небель, А; Филон, Д; Вайс, Д.А.; Уил, М; Фаерман, М; Оппенгейм, А; Томас, МГ (2000). «Гаплотипы Y-хромосомы высокого разрешения израильских и палестинских арабов обнаруживают географическую субструктуру и существенное совпадение с гаплотипами евреев». Хм. Жене . 107 (6): 630–641. дои : 10.1007/s004390000426 . ПМИД 11153918 . S2CID 8136092 .
- Небель, А; Филон, Д; Бринкманн, Б; Маджамдер, П; Фаерман, М; Оппенгейм, А (2001). «Пул Y-хромосомы евреев как часть генетического ландшафта Ближнего Востока» . Американский журнал генетики человека . 69 (5): 1095–1112. дои : 10.1086/324070 . ПМЦ 1274378 . ПМИД 11573163 .
- Небель, А; Ландау-Тассерон, Э; Филон, Д; Оппенгейм, А; Фаерман, М (2002). «Генетические доказательства экспансии арабских племен в Южный Левант и Северную Африку» . Американский журнал генетики человека . 70 (6): 1594–1596. дои : 10.1086/340669 . ПМК 379148 . ПМИД 11992266 .
- Ногейро, И.; Манко, Л.; Гомес, В.; Аморим, А.; Гужман, Л. (2010). «Филогеографический анализ отцовских линий в еврейских общинах северо-восточной Португалии» . Американский журнал физической антропологии . 141 (3): 373–381. дои : 10.1002/ajpa.21154 . ПМИД 19918998 .
- Онофри, Валерио; Алессандрини, Федерика; Турки, Кьяра; Песарези, Мауро; Тальябраччи, Адриано (2008). «Распределение маркеров Y-хромосомы в Северной Африке: анализ SNP и STR высокого разрешения в популяциях Туниса и Марокко». Forensic Science International: Серия дополнений по генетике . 1 : 235–236. дои : 10.1016/j.fsigss.2007.10.173 .
- Оттони, Клаудио; Лармюсо, Мартен HD; Вандерхейден, Нэнси; Мартинес-Лабарга, Кристина; Приматив, Жозефина; Бионди, Джанфранко; Декорте, Ронни; Рикардс, Ольга (2011). «Глубоко в корни ливийских туарегов: генетический обзор их отцовского наследия». Американский журнал физической антропологии . 145 (1): 118–124. дои : 10.1002/ajpa.21473 . ПМИД 21312181 .
- Регейру, М.; Каденас, AM; Гайден, Т.; Андерхилл, Пенсильвания; Эррера, Р.Дж. (2006). «Иран: трехконтинентальная связь для миграции, вызванной Y-хромосомой». Наследственность человека . 61 (3): 132–143. дои : 10.1159/000093774 . ПМИД 16770078 . S2CID 7017701 .
- Реппинг, С; ван Даален, Словакия; Браун, LG; Корвер, Синди М; Ланге, Джулиан; Маршалек, Джанет Д; Пынтикова Татьяна; ван дер Вин, Фулько; и др. (2006). «Высокая частота мутаций привела к обширному структурному полиморфизму среди Y-хромосом человека». Нат Жене . 38 (4): 463–467. CiteSeerX 10.1.1.537.1822 . дои : 10.1038/ng1754 . ПМИД 16501575 . S2CID 17083896 .
- Робино, К.; Кробу, Ф.; Ди Гаэтано, К.; Бекада, А.; Бенхамамуш, С.; Черутти, Н.; Пьяцца, А.; Интурри, С.; Торре, К. (2008). «Анализ гаплогрупп SNP Y-хромосомы и гаплотипов STR в выборке населения Алжира». Международный журнал юридической медицины . 122 (3): 251–255. дои : 10.1007/s00414-007-0203-5 . ПМИД 17909833 . S2CID 11556974 .
- Семино, Орнелла; Магри, Кьяра; Бенуцци, Джорджия; Лин, Алиса А.; Аль-Захери, Надя; Батталья, Винченца; Маччиони, Лилиана; Триантафиллидис, Костас; и др. (2004). «Происхождение, распространение и дифференциация гаплогрупп E и J Y-хромосомы: выводы о неолитизации Европы и более поздних миграционных событиях в Средиземноморье» . Американский журнал генетики человека . 74 (5): 1023–1034. дои : 10.1086/386295 . ПМК 1181965 . ПМИД 15069642 .
- Сенгупта, Сангамитра; Животовский Лев А.; Кинг, Рой; Мехди, SQ; Эдмондс, Кристофер А.; Чоу, Шерил-Эмилиан Т.; Лин, Алиса А.; Митра, Миташри; и др. (2006). «Полярность и временность распределения Y-хромосомы в Индии с высоким разрешением определяют как местное, так и экзогенное распространение и выявляют незначительное генетическое влияние скотоводов Центральной Азии» . Американский журнал генетики человека . 78 (2): 202–221. дои : 10.1086/499411 . ПМЦ 1380230 . ПМИД 16400607 .
- Шен, Пейдун; Лави, Таль; Кивисилд, Тоомас; Чоу, Вивиан; Сенгун, Дениз; Гефель, Дов; Шпирер, Иссак; Вульф, Эйлон; и др. (2004). «Реконструкция отцовских и матрилиний самаритян и других израильских популяций на основе вариаций последовательностей Y-хромосомы и митохондриальной ДНК». Человеческая мутация . 24 (3): 248–260. дои : 10.1002/humu.20077 . ПМИД 15300852 . S2CID 1571356 .
- Шлаш, Лиран И.; Бехар, Дорон М.; Юдковский, Геннадий; Темплтон, Алан; Хадид, Ярин; Базис, Фуад; Хаммер, Майкл; Ицковиц, Шалев; Скорецкий, Карл (2008). «Друзы: популяционно-генетический рефугиум Ближнего Востока» . ПЛОС ОДИН . 3 (5): е2105. Бибкод : 2008PLoSO...3.2105S . дои : 10.1371/journal.pone.0002105 . ПМК 2324201 . ПМИД 18461126 .
- Тофанелли, Серджио; Ферри, Джанмарко; Булаева, Казима; Качиагли, Лаура; Онофри, Валерио; Тальоли, Лука; Булаев Олег; Боски, Илария; и др. (2009). «Происхождение J1-M267 Y отмечает доисторическое перемещение людей, вызванное климатом» . Европейский журнал генетики человека . 17 (11): 1520–1524. дои : 10.1038/ejhg.2009.58 . ПМЦ 2986692 . ПМИД 19367321 .
- Консорциум Y-хромосомы «YCC» (2002). «Система номенклатуры дерева бинарных гаплогрупп Y-хромосомы человека» . Геномные исследования . 12 (2): 339–48. дои : 10.1101/гр.217602 . ПМК 155271 . ПМИД 11827954 .
- Заллуа, Пьер А.; Сюэ, Яли; Халифе, Джейд; Махул, Надин; Дебиан, Лабиб; Платт, Дэниел Э.; Ройюру, Аджай К.; Эррера, Рене Дж.; и др. (2008). «Разнообразие Y-хромосом в Ливане структурировано недавними историческими событиями» . Американский журнал генетики человека . 82 (4): 873–882. дои : 10.1016/j.ajhg.2008.01.020 . ПМЦ 2427286 . ПМИД 18374297 .
- Заллуа, Питер А.; Платт, Дэниел Э.; Сибай, Мирват; Халифе, Джейд; Махул, Надин; Хабер, Марк; Сюэ, Он был; Изабель, Хасан; и др. (2008). «Выявление генетических следов исторической экспансии: финикийские следы в Средиземноморье» . Американский журнал генетики человека . 83 (5): 633–642. дои : 10.1016/j.ajhg.2008.10.012 . ПМК 2668035 . ПМИД 18976729 .
Веб-сайты
[ редактировать ]Гаплогруппы/Филогения
- ИЗОГГ ; Шрак, Янцен (2013). «Гаплогруппа J Y-ДНК и ее субклады» . Международное общество специалистов по генетической генеалогии «ISOGG». Текущие исправления/дополнения гражданских ученых.
Исследовательские проекты гаплотипов/SNP. См. также проекты гаплогрупп Y-ДНК (ISOGG Wiki).
- Шрак; Янзен; Роттенштайнер; Риччи; Мас (2013). «Проект гаплогруппы Y-ДНК J» . ДНК генеалогического древа. Это продолжающийся исследовательский проект гражданских ученых. Более 2300 участников.
- Гиваргидзе; Гречдакян (2013). «Проект J1* Y-ДНК» . ДНК генеалогического древа. Это продолжающийся исследовательский проект гражданских ученых. Более 150 участников.
- Аль Хаддад (2013). «J1c3 (J-L147)» . ДНК генеалогического древа. Это продолжающийся исследовательский проект гражданских ученых. Более 550 участников.
- Конус; Аль Газза; Сандерс (2013). «Проект J-M172 Y-ДНК (J2)» . ДНК генеалогического древа. Это продолжающийся исследовательский проект гражданских ученых. Более 1050 участников.
- Абурто; Кац; Аль Газза; Янзен (2013). «Проект гаплогруппы J-L24-Y-ДНК (J2a1h)» . ДНК генеалогического древа. Это продолжающийся исследовательский проект гражданских ученых. Более 450 участников.
Проекты этнических/географических групп, специфичных для гаплогрупп
- Эддали (2013). «Арабские племена» . ДНК генеалогического древа. Это продолжающийся исследовательский проект гражданских ученых. Более 950 членов J.
- Аль Газза (2013). «Проект J2-Ближний Восток: проект династии J2 в арабском мире и на Ближнем Востоке » . ДНК генеалогического древа. Это продолжающийся исследовательский проект гражданских ученых. Более 400 участников.
Дальнейшее чтение
[ редактировать ]- Бехар, Дорон М.; Томас, Марк Г.; Скорецкий, Карл; Хаммер, Майкл Ф.; Булыгина Екатерина; Розенгартен, Дрор; Джонс, Эбигейл Л.; Держись, Карен; и др. (2003). «Множественное происхождение ашкеназских левитов: свидетельства Y-хромосомы как ближневосточного, так и европейского происхождения» . Американский журнал генетики человека . 73 (4): 768–79. дои : 10.1086/378506 . ПМК 1180600 . ПМИД 13680527 .
- Бехар, Дорон М.; Гарриган, Дэниел; Каплан, Мэтью Э.; Мобашер, Захра; Розенгартен, Дрор; Карафет Татьяна М.; Кинтана-Мурси, Луис; Острер, Гарри; и др. (2004). «Контрастные закономерности вариаций Y-хромосомы у евреев-ашкенази и нееврейского европейского населения». Генетика человека . 114 (4): 354–65. дои : 10.1007/s00439-003-1073-7 . ПМИД 14740294 . S2CID 10310338 .
- Фирасат, Садаф; Халик, Шагуфта; Мохьюддин, Аиша; Папайоанну, Мирто; Тайлер-Смит, Крис; Андерхилл, Питер А; Аюб, Касим (2006). «Y-хромосомные доказательства ограниченного вклада греков в патанское население Пакистана» . Европейский журнал генетики человека . 15 (1): 121–6. дои : 10.1038/sj.ejhg.5201726 . ПМЦ 2588664 . ПМИД 17047675 .
- Флорес, Карлос; Мака-Мейер, Николь; Гонсалес, Ана М; Офнер, Питер Дж; Шен, Пейдун; Перес, Хосе А; Рохас, Антонио; Ларруга, Хосе М; Андерхилл, Питер А. (2004). «Сокращенная генетическая структура Пиренейского полуострова, выявленная с помощью анализа Y-хромосомы: последствия для демографии населения» . Европейский журнал генетики человека . 12 (10): 855–63. дои : 10.1038/sj.ejhg.5201225 . ПМИД 15280900 .
- Семино, О.; Пассарино, Дж; Оефнер, П.Дж.; Лин, А.А.; Арбузова С; Бекман, Ле; Де Бенедиктис, Дж; Франкалаччи, П; и др. (2000). «Генетическое наследие палеолитического Homo sapiens sapiens у современных европейцев: хромосомная перспектива AY». Наука . 290 (5494): 1155–9. Бибкод : 2000Sci...290.1155S . дои : 10.1126/science.290.5494.1155 . ПМИД 11073453 .
Филогенетические заметки
[ редактировать ]- ^ В этой таблице показаны исторические названия J-M267 и его ранее открытого и названного субклада J-M62 в опубликованной рецензируемой литературе.
YCC 2002/2008 (сокращенно) J-M267 J-M62 Джоблинг и Тайлер-Смит, 2000 г. - 9 Андерхилл 2000 - МЫ Хаммер 2001 - С Карафет 2001 - 23 Семино 2000 - ЕС10 Они в 1999 году - Н4 Волосы 2001 - Б YCC 2002 (от руки) - J1 YCC 2005 (от руки) J1 J1a YCC 2008 (от руки) J1 J1a YCC 2010r (от руки) J1 J1a - ^ В этой таблице показаны исторические названия J-P209 (AKA J-12f2.1 или J-M304) в опубликованной рецензируемой литературе. Обратите внимание, что в Semino 2000 Eu09 является субкладом Eu10, а в Karafet 2001 24 является субкладом 23 .
YCC 2002/2008 (сокращенно) J-P209
(АКА J-12f2.1 или J-M304)Джоблинг и Тайлер-Смит, 2000 г. 9 Андерхилл 2000 МЫ Хаммер 2001 С Карафет 2001 23 Семино 2000 ЕС10 Они в 1999 году Н4 Волосы 2001 Б YCC 2002 (от руки) Дж* YCC 2005 (от руки) Дж YCC 2008 (от руки) Дж YCC 2010r (от руки) Дж