Гаплогруппа L-M20
| Гаплогруппа L-M20 | |
|---|---|
 | |
| Возможное время происхождения | 30,000 [ 1 ] - 43 000 лет назад. [ 2 ] |
| Возможное место происхождения | Ближний Восток , Западная Азия , Южная Азия или горы Памира |
| Предок | LT |
| Определение мутаций | М11, М20, М61, М185, Л656, Л863, Л878, Л879 [ сеть 1 ] |
| Самые высокие частоты | Сирия Ракка , Белуджистан , Северный Афганистан , Карнатака , Тархан , Джаты , Калаш , Нуристанцы , Бурушо , Пуштуны , Лазы , деревня Афшар , Фасция , Венето , Южный Тироль |
Гаплогруппа L-M20 представляет собой гаплогруппу Y-ДНК человека , которая определяется SNP M11, M20, M61 и M185. Будучи вторичным потомком гаплогруппы K и первичной ветвью гаплогруппы LT , гаплогруппа L в настоящее время имеет альтернативное филогенетическое название K1a и является родственным братом гаплогруппы T (также известной как K1b).
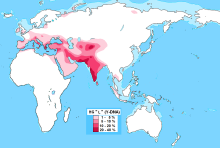
Присутствие L-M20 наблюдалось на разных уровнях по всей Южной Азии , достигая пика в популяциях, обитающих в Белуджистане (28%), [ 3 ] Северный Афганистан (25%), [ 4 ] и Южная Индия (19%). [ 5 ] Клада также встречается в Таджикистане и Анатолии , а также, реже, в Иране . Он также присутствовал на протяжении тысячелетий в очень небольших количествах на Кавказе , в Европе и Центральной Азии . Субклад L2 (L-L595) обнаружен в Европе и Западной Азии, но встречается крайне редко.
Филогенетическое дерево
[ редактировать ]Существует несколько подтвержденных и предложенных филогенетических деревьев гаплогруппы L-M20. Научно признанным является вариант Консорциума Y-хромосомы (YCC), опубликованный в Karafet 2008 и впоследствии обновленный. Черновик дерева, показывающего развивающуюся науку, предоставлен Томасом Краном из Центра геномных исследований в Хьюстоне , штат Техас. [ сеть 1 ] Международное общество генетической генеалогии (ISOGG) также предоставляет любительское дерево.
Этот раздел нуждается в расширении . Вы можете помочь, добавив к нему . ( январь 2013 г. ) |
Это Томас Кран из проекта предлагаемого дерева гаплогруппы L-M20 Центра геномных исследований: [ сеть 1 ]
- L-M20 M11, M20, M61, M185, L656, L863, L878, L879
- L-M22 (L1) M22, M295, СТРАНИЦЫ00121
- Л-М317 (Л1б) М317, Л655
- Л-Л656 (Л1б1) Л656
- Л-М349 (Л1б1а) М349
- Л-М274 М274
- L1310 L1310
- Л-СК1412
- Л-Л656 (Л1б1) Л656
- Л-Л1304 Л1304
- Л-М27 (Л1а1) М27, М76, П329.1, Л1318, Л1319, Л1320, Л1321
- Л-М357 (Л1а2) М357, Л1307
- Л-ПК3 ПК3
- Л-Л1305 Л1305, Л1306, Л1307
- Л-М317 (Л1б) М317, Л655
- Л-Л595 (Л2) Л595
- Л864 Л864 , Л865, Л866, Л867, Л868, Л869, Л870, Л877
- L-M22 (L1) M22, M295, СТРАНИЦЫ00121
Происхождение
[ редактировать ]L-M20 — потомок гаплогруппы LT , [ 6 ] [ 7 ] который является потомком гаплогруппы К-М9 . [ 8 ] [ 7 ] По мнению доктора Спенсера Уэллса , L-M20 произошел от евразийского клана K-M9, который мигрировал на восток с Ближнего Востока , а затем на юг от Памирского узла в современные Пакистан и Индию . [ 9 ] [ 10 ] Эти люди прибыли в Индию примерно 30 000 лет назад. [ 9 ] [ 10 ] Следовательно, предполагается, что первый носитель маркера M20 родился либо в Индии, либо на Ближнем Востоке. [ 9 ] Другие исследования предполагают происхождение L-M20 либо из Западной Азии , либо из Южной Азии и связывают его распространение в долине Инда с фермерами эпохи неолита . [ 11 ] [ 12 ] [ 13 ] [ 14 ] [ 15 ] [ 16 ] Генетические исследования предполагают, что L-M20 может быть одной из гаплогрупп первоначальных создателей цивилизации долины Инда . [ 3 ] [ 17 ] [ 15 ] МакЭлриви и Кинтана-Мурси, пишущие о цивилизации долины Инда, утверждают, что
Одна гаплогруппа Y-хромосомы (L-M20) имеет высокую среднюю частоту - 14% в Пакистане и поэтому отличается от всех других гаплогрупп своим распределением частот. L-M20 также наблюдается, хотя и на более низких частотах, в соседних странах, таких как Индия, Таджикистан, Узбекистан и Россия. Как частотное распределение, так и предполагаемое время распространения (~ 7000 лет назад) этой линии позволяют предположить, что ее распространение в долине Инда может быть связано с расширением местных земледельческих групп в период неолита. [ 15 ]
Сенгупта и др. (2006) наблюдали три подветви гаплогруппы L: L1-M76 (L1a1), L2-M317 (L1b) и L3-M357 (L1a2), имеющие различную географическую принадлежность. [ 5 ] Почти все индийские члены гаплогруппы L происходят от L1, при этом L3-M357 встречается лишь спорадически (0,4%). [ 18 ] [ 19 ] Напротив, в Пакистане субклад L3-M357 составляет 86% хромосом L-M20 и в целом достигает промежуточной частоты 6,8%. [ 20 ] L1-M76 встречается с частотой 7,5% в Индии и 5,1% в Пакистане, демонстрируя пиковое распределение дисперсии в регионе Махараштры в прибрежной западной Индии. [ 21 ]
Географическое распространение
[ редактировать ]В Индии L-M20 чаще встречается среди дравидийских каст, но несколько реже встречается в индоарийских кастах. [ 5 ] В Пакистане наибольшая частота встречается в Белуджистане . [ 22 ]
Он также был обнаружен с низкой частотой среди населения Центральной Азии и Юго-Западной Азии (включая Аравию , Ирак , Сирию , Турцию , Ливан , Египет и Йемен ), а также в Южной Европе (особенно в районах, прилегающих к Средиземному морю ). [ нужна ссылка ]
Предварительные данные, полученные из ненаучных источников, например, от людей, чьи Y-хромосомы были проверены в коммерческих лабораториях, [ сеть 2 ] предполагает, что большинство европейских примеров гаплогруппы L-M20 могут принадлежать к субкладу L2-M317, который среди населения Южной Азии, как правило, является самым редким из субкладов гаплогруппы L. [ сеть 2 ]
Южная Азия
[ редактировать ]Индия
[ редактировать ]Он встречается чаще среди дравидийских каст (около 17-19%), но несколько реже в индоарийских кастах (около 5-6%). [ 5 ] Присутствие гаплогруппы L-M20 довольно редко встречается среди племенных групп (около 5,6-7%) ( Cordaux 2004 , Sengupta 2006 и Thamseem 2006 ). Однако племя Корова в штате Уттара-Каннада , в котором L-M11 встречается на уровне 68%, является исключением. [ 23 ]
L-M20 был обнаружен у 38% представителей касты Бхарвад и 21% у Чаран касты из района Джунагарх в Гуджарате . ( Шах, 2011 ). Сообщалось также, что он встречается у 17% в племени Каре Воккал из Уттара-Каннада в Карнатаке . ( Шах, 2011 ). ) Он также встречается с низкой частотой у других популяций из района Джунагарх и Уттара-Каннада. L-M20 является крупнейшей мужской линией (36,8%) среди народа джат в Северной Индии и встречается на уровне 16,33% среди гуджаров Джамму и Кашмира . [ 17 ] [ 24 ] Это также встречается на уровне 18,6% среди конканастха-брахманов региона Конкан . [ 25 ] % среди маратхов Махараштры . и 15 [ 26 ] L-M20 также встречается в 32,35% у воккалигасов и в 17,82% у лингаятов Карнатаки. [ 27 ]
И доступные данные [ 28 ] показывает, что среди тамилов L-M20 встречается у 48% у калларов , 20,56% у тамильских ядавов , 28,57% у ваннияров , 28,81% у надаров , 26% у народа саураштры , 20,7% у амбалакараров , 16,7% у народа саураштры, 20,7% у тамильских ядавов , 16,7% у Айенгара и 17,2% среди Айерские касты Тамилнада . [ 29 ] [ 26 ] L-M11 встречается с частотой 8–16% среди индийских евреев . [ 30 ] составляет 12% Общая частота L-M20 в Пенджабе . [ 19 ] 2% сидди Сообщается также, что у есть L-M11. ( Shah 2011 ) Гаплогруппа L-M20 в настоящее время присутствует в индийском населении с общей частотой ок. 7-15%. [ Сноска 1 ]
Пакистан
[ редактировать ]Наибольшая концентрация гаплогруппы L-M20 находится вдоль реки Инд в Пакистане , где цивилизация долины Инда процветала в 3300–1300 годах до нашей эры, а период ее зрелости - между 2600 и 1900 годами до нашей эры. Самая высокая частота и разнообразие L-M357 обнаружены в провинции Белуджистан - 28%. [ 22 ] с умеренным распространением среди населения Пакистана в целом - 11,6% ( Firasat et al. 2007 )). Он также встречается и у этнических аналогов Афганистана , например, у пуштунов и белуджей . L-M357 часто встречается среди бурушо (около 12% ( Firasat et al. 2007 )) и пуштунов (около 7% ( Firasat et al. 2007 )),
L1a и L1c-M357 встречаются у 24% среди белуджей, L1a и L1c встречаются у 8% среди дравидоязычных и брахуи , L1c встречается у 25% среди калашей , L1c встречается у 15% среди бурушо , L1a-M76 L1b-M317 встречается у 2% среди Макрани , а L1c встречается у 3,6% синдхов По данным Джули ди Кристофаро и др., . 2013. [ 31 ] L-M20 встречается у парсов у 17,78% . [ 11 ] L3a встречается у 23% нуристанцев как в Пакистане , так и в Афганистане . [ 32 ]
L-PK3 встречается примерно у 23% калашей на северо-западе Пакистана ( Firasat et al. 2007 ).
Ближний Восток и Анатолия
[ редактировать ]L-M20 был обнаружен у 51% сирийцев из Ракки , северного сирийского города, прежние жители которого были уничтожены монгольским геноцидом и в последнее время вновь заселены местным бедуинским населением и чеченскими беженцами из России ( El-Sibai 2009 ). В небольшой выборке израильских друзов гаплогруппа L-M20 была обнаружена у 7 из 20 (35%). Однако исследования, проведенные на более крупных выборках, показали, что L-M20 составляет в среднем 5% среди израильских друзов . [ Сноска 2 ] 8% у ливанских друзов , [ Сноска 3 ] и он не был обнаружен в выборке из 59 сирийских друзов . Гаплогруппа L-M20 была обнаружена у 2,0% (1/50) ( Wells 2001 ) до 5,25% (48/914) ливанцев ( Zalloua 2008 ).
| Население | Распределение | Источник |
|---|---|---|
| Турция | 57% в селе Афшар , 12% (10/83) в Причерноморье , 6,6% (7/106) турок в Турции, 4,2% (1/523 L-M349 и 21/523 L-M11(xM27, M349) )) | Чинниоглу 2004 г. , Гоккумен 2008 г. |
| Иран | 54,9% (42/71) L в священнике зороастрийском парсе 22,2% L1b и L1c в Южном Иране (2/9) От 8% до 16% L2-L595, L1a, L1b и L1c курдов в Курдистане (2-4/25) 9,1% L-M20 (7/77) персов в Восточном Иране 3,4% L-M76 (4/117) и 2,6% L-M317 (3/117) всего 6,0% (7/117) гаплогруппы L-M20 в Южном Иране 3,0% (1/33) L-M357 в Северном Иране 4,2% L1c-M357 азербайджанцев Восточного Азербайджана (1/21) 4,8% L1a и L1b персов в Исфахане (2/42) |
Регейро 2006 г. , Ди Кристофаро 2013 г. , Малярчук 2013 г. |
| Сирия | 51,0% (33/65) сирийцев в Ракке , 31,0% восточных сирийцев | Эль-Сибай 2009 г. |
| Лаз | 41,7% (15/36) Л1б-М317 | [ нужна ссылка ] |
| Саудовские арабы | 15,6% (4/32 от L-M76 и 1/32 от L-317) 1,91% (2/157=1,27% L-M76 и 1/157=0,64% L-M357) | Абу-Амеро 2009 г. |
| Курды | 3,2% курдов на юго-востоке Турции | Флорес 2005 г. |
| Ирак | 3,1% (2/64) М22 | Санчес 2005 г. |
| Армяне | От 1,63% (12/734) до 4,3% (2/47) | Уил 2001 г. и Уэллс 2001 г. |
| Оманцы | 1% Л-М11 | Луис 2004 г. |
| Катарцы | 2,8% (2/72 L-M76) | Цепи 2008 |
| Арабские Эмираты | 3,0% (4/164 L-M76 и 1/164 L-M357) | Цепи 2008 |
Центральная Азия
[ редактировать ]Афганистан
[ редактировать ]Исследование пуштунских мужских линий в Афганистане показало, что гаплогруппа L-M20 с общей частотой 9,5% является второй по численности мужской линией среди них. [ 33 ] Он демонстрирует существенные различия в своем распределении по обе стороны хребта Гиндукуш: к этой линии принадлежат 25% северных афганских пуштунов по сравнению с только 4,8% мужчин с юга. [ 33 ] В частности, на парагруппу L3*-M357 приходится большая часть хромосом L-M20 у афганских пуштунов как на севере (20,5%), так и на юге (4,1%). [ 33 ] Более раннее исследование с меньшим количеством образцов показало, что L1c составляет 12,24% мужских линий афганских пуштунов . [ 34 ] [ 35 ] L1c также встречается у 7,69% белуджей Афганистана . [ 34 ] Однако L1a-M76 встречается гораздо чаще среди белуджей (20 [ 35 ] до 61,54%), [ 35 ] и встречается на более низких уровнях у кыргызского, таджикского, узбекского и туркменского населения. [ 35 ]
Восточная Азия
[ редактировать ]Исследователи, изучающие образцы Y-ДНК популяций Восточной Азии, редко проверяли свои образцы на наличие каких-либо мутаций, определяющих гаплогруппу L. Однако мутации гаплогруппы L были протестированы и обнаружены в образцах балийцев (13/641 = 2,0% L). -М20), ханьцы (1/57 = 1,8%), [ 37 ] Долганы из Саха и Таймыра (1/67 = 1,5% L-M20) и корейцы (3/506 = 0,6% L-M20). [ 38 ] [ 39 ] [ 40 ]
Европа
[ редактировать ]Статья О. Семино и др. опубликовано в журнале Science (том 290, 10 ноября 2000 г.) сообщается об обнаружении мутации M11-G, которая является одной из мутаций, определяющих гаплогруппу L, примерно в 1–3% образцов из Грузии , Греции , Венгрии , Калабрия (Италия) и Андалусия (Испания). Размеры выборок, проанализированных в этом исследовании, как правило, были довольно небольшими, поэтому вполне возможно, что фактическая частота гаплогруппы L-M20 среди популяций средиземноморской Европы может быть немного ниже или выше, чем сообщалось Semino et al. , но, похоже, на сегодняшний день не существует исследования, которое бы более точно описало распространение гаплогруппы L-M20 в Юго-Западной Азии и Европе.
| Население | Распределение | Источник |
|---|---|---|
| Фасция , Италия | 19,2% фашистов L-M20 | [ нужна ссылка ] |
| Нонсталь , Италия | 10% Нонеси L-M20 | Ди Джакомо 2003 |
| Самниум , Италия | 10% Аквиланис Л-М20 | Боаттини 2013 |
| Виченца , Италия | 10% венецианцев L-M20 | Боаттини 2013 |
| Южный Тироль , Италия | 8,9% говорящих на ладинском языке из Валь Бадии , 8,3% из Валь Бадии , 2,9% из долины Пустер , 2,2% по-немецки говорящих из Валь Бадии , 2% говорящих по-немецки из Верхнего Виншгау , 1,9% говорящих по-немецки из Нижнего Виншгау и 1,7% говорящих на немецком языке итальянских Больцано из спикеров | Пихлер 2006 г. и Томас 2007 г. |
| грузины | 20% (2/10) грузин в Гали , 14,3% (2/14) грузин в Чохатаури , 12,5% (2/16) грузин в Мартвили , 11,8% (2/17) грузин в Абаше , 11,1% (2/18) грузин в Багдати , 10% (1/10) грузин в Гардабани , 9,1% (1/11) грузин в Адигени , 6,9% (2/29) грузин в Омало , 5,9% (1/17) грузин в Гурджаани , 5,9% (1/17) грузин в Лентехи и от 1,5% (1/66) L-M357(xPK3) до 1,6%. (1/63) Л-М11 | Батталья 2008 г. , Семино 2000 г. и Тархнишвили 2014 г. |
| Дагестан , Россия | 10% В Дагестане чеченцев , 9,5% (4/42) аварцев , 8,3% (2/24) татов , 3,7% (1/27) чамалинцев. | Юнусбаев 2006 г. , Качиагли 2009 г. |
| Arkhangelsk Oblast , Russia | 5,9% россиян L1c-M357 | [ нужна ссылка ] |
| Эстония | L2-L595 и L1-M22 встречаются у 5,3%, 3,5%, 1,4% и 0,8% эстонцев. | Скоццари, 2001 г. и Лаппалайнен, 2008 г. |
| Балкарцы , Россия | 5,3% (2/38) Л-М317 | Битва 2008 |
| Португалия | 5,0% Коимбры | Красота 2006 |
| Болгария | 3,9% болгар | [ нужна ссылка ] |
| Фландрия | L1a*: 3,17% Мехелена, 2,4% Тюрнхаута и 1,3% Кемпена . L1b*: 0,74% Западной Фландрии и Восточной Фландрии. | Лармюсо 2010 г. и Лармюсо 2011 г. |
| Antsiferovo , Novgorod | 2,3% россиян | [ нужна ссылка ] |
| Восточный Тироль , Австрия | L-M20 встречается у 1,9% тирольцев в регионе B ( Изель , Нижняя Драу , Деферегген , Вирген и долина Кальс ). | [ нужна ссылка ] |
| Гипускоа , Страна Басков | L1b встречается у 1,7% гипускоанцев. | Молодой 2011 |
| Северный Тироль , Австрия | L-M20 встречается у 0,8% тирольцев в Ройтте. | [ нужна ссылка ] |
Южная Африка и побережье Суахили
[ редактировать ]В 2013 году исследователи, изучающие происхождение народа лемба , имеющего южноаравийское происхождение по отцовской линии, обнаружили, что 13,8% мужчин лемба носят Y-ДНК L-M20, в частности субклад L-M349, что делает его четвертой по распространенности линией среди них. . [ 41 ] Было обнаружено, что образец лембы из Южной Африки, представленный Familytreedna в 2023 году, несет еще неназванный субклад L-M349 L-FT408126, который был наиболее близок к двум образцам из Ирака и Ирана . [ 42 ]
Исследователи также обнаружили следы L-M20 на побережье Суахили в Кении, составляющие 4,2% от общей численности населения.
Распределение субкладов
[ редактировать ]Л1 (М295)
[ редактировать ]L-M295 встречается от Западной Европы до Южной Азии . [ Сноска 5 ]
Субклад L1 также встречается на низких частотах на Коморских островах . [ 43 ]
L1a1 (M27)
[ редактировать ]L-M27 встречается у 14,5% индийцев и 15% жителей Шри-Ланки , с умеренным распространением в других популяциях Пакистана , южного Ирана и Европы , но несколько выше в популяциях арабов Ближнего Востока . [ нужна ссылка ] очень незначительное присутствие Среди Сидди (2%), [ 44 ] также.
L1a2(M357)
[ редактировать ]L-M357 часто встречается среди бурушо , калашей , джатов , пуштунов , с умеренным распространением среди других популяций Пакистана , Грузии , [ 45 ] Чеченцы , [ 46 ] Ингуши , [ 46 ] северный Иран , Индия , ОАЭ и Саудовская Аравия . [ нужна ссылка ]
Китайское исследование, опубликованное в 2018 году, обнаружило L-M357/L1307 у 7,8% (5/64) выборки уйгуров Лоплика из деревни Карчуга, уезда Лопнур , Синьцзян. [ 36 ]
- Л-ПК3
L-PK3, расположенный после L-M357, [ 47 ] часто встречается у калашей . [ нужна ссылка ]
Л1б (М317)
[ редактировать ]L-M317 встречается с низкой частотой в Центральной Азии , Юго-Западной Азии и Европе . [ нужна ссылка ]
В Европе L-M317 был обнаружен у северо-восточных итальянцев (3/67 = 4,5%). [ 45 ] и греки (1/92 = 1,1%). [ 45 ]
На Кавказе L-M317 обнаружен у горских евреев (2/10 = 20% [ 48 ] ), аварцы (4/42 = 9,5%, [ 48 ] 3% [ 46 ] ), балкарцы (2/38 = 5,3%), [ 45 ] Абхазы (8/162 = 4,9%, [ 48 ] 2/58 = 3.4% [ 46 ] ), Чамалалы (1/27 = 3,7% [ 48 ] ), не состоящие в браке (2/88 = 2,3% [ 48 ] ), адыгейцы (3/154 = 1,9% [ 48 ] ), чеченцы (3/165 = 1,8% [ 48 ] ), армяне (1/57 = 1,8% [ 48 ] ), лезгины (1/81 = 1,2% [ 46 ] ), и осетины (1/132 = 0,76% североосетин , [ 48 ] 2/230 = 0,9% железа [ 46 ] ).
L-M317 обнаружен у Макраниса (2/20 = 10%) в Пакистане, иранцев (3/186 = 1,6%), пуштунов в Афганистане. (1/87 = 1,1%) и узбеки в Афганистане (1/127 = 0,79%). [ 35 ]
L1b1 (М349)
[ редактировать ]L-M349 встречается у некоторых крымских караимов, являющихся левитами . [ 49 ] Некоторые из филиалов L-M349 находятся в Западной Азии, в том числе L-Y31183 в Ливане , L-Y31184 в Армении и L-Y130640 в Ираке , Иране , Йемене и Южной Африке . Другие встречаются в Европе, например L-PAGE116 в Италии , L-FT304386 в Словении и L-FGC36841 в Молдове . [ 50 ] 13,8% мужчин лемба несут L-M349 клады L-Y130640. [ 41 ] Этот процент, скорее всего, обусловлен эффектом основателя их населения, что делает их единственной группой на африканском континенте со значительной долей L-M20.
Л2 (Л595)
[ редактировать ]L2-L595 встречается крайне редко и был выявлен в результате частного тестирования у людей из Европы и Западной Азии.
О двух подтвержденных людях L2-L595 из Ирана сообщалось в дополнительном исследовании 2020 года. [ 51 ] Возможные, но неподтвержденные случаи L2 включают 4% (1/25) L-M11(xM76, M27, M317, M357) в выборке иранцев в Курдистане. [ 35 ] и 2% (2/100) L-M20(xM27, M317, M357) в выборке шапсугов , [ 46 ] среди других редких зарегистрированных случаев L, которые не попадают в общие ветви.
| Область | Население | н/размер выборки | Процент | Источник |
|---|---|---|---|---|
| Западная Азия | Азербайджан | 2/204 | 1 | [ 52 ] |
| Центральная Европа | Германия | 1/8641 | 0.0000115 | [ 53 ] |
| Южная Европа | Греция | 1/753 | 0.1 | [ 54 ] |
| Западная Азия | Иран | 2/800 | 0.25 | [ 51 ] |
| Южная Европа | Италия | 3/913 | 0.3 | [ 55 ] |
Древняя ДНК
[ редактировать ]- Три человека майкопской культуры ок. Было обнаружено, что 3200 г. до н.э. принадлежат к гаплогруппе L2-L595. [ 56 ]
- три особи, жившие в эпоху энеолита (ок. 5700–6250 гг. н. э. ), найденные в пещере Арени-1 («Птичий глаз») в горах Южного Кавказа (современная Вайоцдзорская область , Армения Также были идентифицированы ). как принадлежащий к гаплогруппе L1a. Геном одного человека показал, что у него рыжие волосы и голубые глаза. Их генетические данные приведены в таблице ниже.
- Нарасимхан и др. (2018) проанализировали скелеты из стоянок BMAC в Узбекистане и идентифицировали 2 человека как принадлежащих к гаплогруппе L1a. Один из этих экземпляров был найден в Бустане, другой – в Сапали-Тепе; оба были признаны памятниками бронзового века . [ 57 ]
- Скуртаниоти и др. (2020) проанализировали скелеты из Алалаха и идентифицировали одну особь (ALA084) c. 2006-1777 гг. до н.э. как принадлежащий к гаплогруппе L-L595 (L2). [ 58 ] Ингман и др. (2021) проанализировали дополнительные скелеты из Алалаха и идентифицировали еще одного человека, принадлежащего к гаплогруппе L-M349 (L1b). [ 59 ]
- Один человек железного века из Бэтмена в Верхней Месопотамии (современная Юго- Восточная Турция ) принадлежал к гаплогруппе L2-L595. [ 60 ]
- Было установлено, что древний викинг, живший на острове Эланд , Швеция, примерно в 847 ± 65 году нашей эры, принадлежал к L-L595. [ 61 ]
Энеолит Южный Кавказ
[ редактировать ]| Свойство | Арени-I | Арени-II | Арени-III |
|---|---|---|---|
| ИДЕНТИФИКАТОР | АР1/44 I1634 | АР1/46 I1632 | ARE12 I1407 |
| Y ДНК | Л1а | Л1а1-М27 | Л1а |
| Население | Эпоха энеолита (Горизонт III) | Эпоха энеолита (Горизонт III) | Эпоха энеолита (Горизонт II) |
| Язык | |||
| Культура | Поздний энеолит | Поздний энеолит | Поздний энеолит |
| Дата (YBP) | 6161 ± 89 | 6086 ± 72 | 6025 ± 325 |
| Похороны / Местонахождение | Погребение 2 / Пещера Арени-1 | Погребение 3 / Пещера Арени-1 | Траншея 2А, Блок 7, Площадь S33/T33, Участок 9, Коса 23 / Пещера Арени-1 |
| Члены / Размер выборки | 1/3 | 1/3 | 1/3 |
| Процент | 33.3% | 33.3% | 33.3% |
| мтДНК | H2a1 | К1а8 | ЧАС* |
| Изотоп-старший | |||
| Цвет глаз (система HIrisPlex) | Вероятно синий | ||
| Краска для волос (система HIrisPlex) | Вероятно красный | ||
| Пигментация кожи | Вероятно свет | ||
| Группа крови АВО | Скорее всего О или Б | ||
| Диета (d13C%0 / d15N%0) | |||
| Деятельность ФАДС | |||
| Стойкость лактазы | Вероятно, непереносимость лактозы | ||
| Oase-1 Общая ДНК | |||
| Ostuni1 Общая ДНК | |||
| Неандерталец Vi33.26 Общая ДНК | |||
| Неандерталец Vi33.25 Общая ДНК | |||
| Неандерталец Vi33.16 Общая ДНК | |||
| Родовой компонент (AC) | |||
| пунтDNAL K12 Древний | |||
| Додекада [dv3] | |||
| Евроген [К=36] | |||
| Додекада [Глобус13] | |||
| Генетическая дистанция | |||
| Родительское родство | |||
| Возраст смерти | 11 ± 2.5 | 15 ± 2.5 | |
| Позиция смерти | |||
| SNP | |||
| Чтение пар | |||
| Образец | |||
| Источник | [ 62 ] | ||
| Примечания | Самые ранние в мире свидетельства обувного и винодельческого производства |
Номенклатура
[ редактировать ]До 2002 года в научной литературе существовало как минимум семь систем наименования филогенетического дерева Y-хромосомы. Это привело к значительной путанице. В 2002 году основные исследовательские группы объединились и сформировали Консорциум Y-хромосомы (YCC). Они опубликовали совместный документ, в котором было создано единое новое дерево, которое все согласились использовать. Позже группа гражданских ученых, интересующихся популяционной генетикой и генетической генеалогией, сформировала рабочую группу для создания любительского дерева, стремясь быть прежде всего своевременным. В таблице ниже собраны все эти работы, относящиеся к знаковому дереву YCC 2002 года. Это позволяет исследователю, просматривающему старую опубликованную литературу, быстро перемещаться между номенклатурами.
| YCC 2002/2008 (сокращенно) | (а) | (б) | (с) | (г) | (е) | (г) | (или) | YCC 2002 (от руки) | YCC 2005 (от руки) | YCC 2008 (от руки) | YCC 2010r (от руки) | ИСОГГ 2006 г. | ИСОГГ 2007 | ИСОГГ 2008 | ИСОГГ 2009 | ИСОГГ 2010 | ИЗОГГ 2011 | ИЗОГГ 2012 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Л-М20 | 28 | VIII | 1U | 27 | ЕС17 | Н5 | Ф | Л* | л | л | л | - | - | - | - | - | - | - |
| Л-М27 | 28 | VIII | 1U | 27 | ЕС17 | Н5 | Ф | Л1 | Л1 | Л1 | Л1 | - | - | - | - | - | - | - |
- Дерево Консорциума Y-хромосомы
Это официальное научное дерево, созданное Консорциумом Y-хромосомы (YCC). Последнее крупное обновление было в 2008 году. [ нужна ссылка ] Последующие обновления были ежеквартальными и двухгодичными. Текущая версия является версией обновления 2010 года. [ 63 ]
Этот раздел нуждается в расширении . Вы можете помочь, добавив к нему . ( январь 2013 г. ) |
- Оригинальные исследовательские публикации
Следующие исследовательские группы в соответствии с их публикациями были представлены в создании Дерева YCC.
См. также
[ редактировать ]Сноски
[ редактировать ]- ^ см . Басу 2003 , Кордо 2004 , Сенгупта 2006 и Тамсим 2006 .
- ^ 12/222 Шлаш и др. 2008 год
- ^ 1/25 Шлаш и др. 2008 год
- ^ В Hammer 2005 см. дополнительный материал.
- ^ Результаты лаборатории FTDNA, май 2011 г.
Ссылки
[ редактировать ]- ^ гаплогруппы L Y-хромосомы. Узнайте об учебных пособиях по генной базе
- ^ Yfull Tree L Гаплогруппа YTree v8.09.00 (8 октября 2020 г.)
- ^ Jump up to: а б Дорогой 2018 год .
- ^ Лако Х., Гайден Т., Регейро М., Ченнакришнайя С., Бухари А., Андерхилл П.А. и др. (октябрь 2012 г.). «Афганистан с точки зрения Y-хромосомы» . Европейский журнал генетики человека . 20 (10): 1063–1070. дои : 10.1038/ejhg.2012.59 . ПМК 3449065 . ПМИД 22510847 .
- ^ Jump up to: а б с д Сенгупта 2006 .
- ^ Международное общество генетической генеалогии , 2015, Дерево гаплогрупп Y-ДНК 2015 (30 мая 2015 г.).
- ^ Jump up to: а б Кьярони Дж., Андерхилл, Пенсильвания, Кавалли-Сфорца, Л.Л. (декабрь 2009 г.). «Разнообразие Y-хромосомы, человеческая экспансия, дрейф и культурная эволюция» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (48): 20174–20179. Бибкод : 2009PNAS..10620174C . дои : 10.1073/pnas.0910803106 . JSTOR 25593348 . ПМЦ 2787129 . ПМИД 19920170 .
- ^ Международное общество генетической генеалогии , 2015 Гаплогруппа K Y-ДНК и ее субклады - 2015 (5 апреля 2015 г.).
- ^ Jump up to: а б с Уэллс С. (20 ноября 2007 г.). Глубокая родословная: эпохальный поиск ДНК для расшифровки нашего далекого прошлого . Национальные географические книги. стр. 161–162. ISBN 978-1-4262-0211-7 .
Эта часть евразийского клана М9 мигрировала на юг, как только достигла труднопроходимого и гористого региона Памирского узла. Человек, давший начало маркеру М20, возможно, родился в Индии или на Ближнем Востоке. Его предки прибыли в Индию около 30 000 лет назад и представляют собой самое раннее значительное поселение Индии.
- ^ Jump up to: а б Уэллс С. (28 марта 2017 г.). Путешествие человека: генетическая одиссея . Издательство Принстонского университета. стр. 111–113. ISBN 978-0-691-17601-7 .
- ^ Jump up to: а б Камар Р., Аюб К., Мохьюддин А., Хельгасон А., Мажар К., Мансур А. и др. (май 2002 г.). «Вариации ДНК Y-хромосомы в Пакистане» . Американский журнал генетики человека . 70 (5): 1107–1124. дои : 10.1086/339929 . ПМЦ 447589 . ПМИД 11898125 .
- ^ Чжао З., Хан Ф., Боркар М., Эррера Р., Агравал С. (2009). «Наличие трех разных отцовских линий у жителей Северной Индии: исследование 560 Y-хромосом» . Анналы биологии человека . 36 (1): 46–59. дои : 10.1080/03014460802558522 . ПМЦ 2755252 . ПМИД 19058044 .
- ^ Тансим И., Тангарадж К., Чауби Дж., Сингх В.К., Бхаскар Л.В., Редди Б.М. и др. (август 2006 г.). «Генетическое сходство среди низших каст и племенных групп Индии: вывод на основе Y-хромосомы и митохондриальной ДНК» . БМК Генетика . 7 (1): 42. дои : 10.1186/1471-2156-7-42 . ПМЦ 1569435 . ПМИД 16893451 .
- ^ Кордо Р., Онгер Р., Бентли Г., Насидзе И., Сираджуддин С.М., Стоункинг М. (февраль 2004 г.). «Независимое происхождение индийских каст и племенных отцовских линий» . Современная биология . 14 (3): 231–235. Бибкод : 2004CBio...14..231C . дои : 10.1016/j.cub.2004.01.024 . ПМИД 14761656 . S2CID 5721248 .
- ^ Jump up to: а б с МакЭлриви К., Кинтана-Мурси Л. (2005). «Перспектива популяционной генетики долины Инда с помощью маркеров, унаследованных одним родителем». Анналы биологии человека . 32 (2): 154–162. дои : 10.1080/03014460500076223 . ПМИД 16096211 . S2CID 109014 .
- ^ Тангарадж К., Найду Б.П., Кривелларо Ф., Таманг Р., Упадхьяй С., Шарма В.К. и др. (декабрь 2010 г.). «Влияние естественных барьеров на формирование генетической структуры популяций Махараштры» . ПЛОС Один . 5 (12): e15283. Бибкод : 2010PLoSO...515283T . дои : 10.1371/journal.pone.0015283 . ПМК 3004917 . ПМИД 21187967 .
- ^ Jump up to: а б Махал Д.Г., Мацукас И.Г. (20 сентября 2017 г.). «Разнообразие гаплогрупп Y-STR в популяции джатов раскрывает несколько различных древних источников» . Границы генетики . 8 : 121. дои : 10.3389/fgene.2017.00121 . ПМК 5611447 . ПМИД 28979290 .
- ^ Сенгупта 2006 , с. 218.
- ^ Jump up to: а б Каменный мост 2003г .
- ^ Сенгупта 2006 , с. 219.
- ^ Сенгупта 2006 , с. 220.
- ^ Jump up to: а б Камар 2002 .
- ^ Шах 2011 .
- ^ Шарма С., Рай Э., Шарма П., Джена М., Сингх С., Дарвиши К. и др. (январь 2009 г.). «Индийское происхождение отцовской гаплогруппы R1a1* обосновывает автохтонное происхождение брахманов и кастовой системы» . Журнал генетики человека . 54 (1): 47–55. дои : 10.1038/jhg.2008.2 . ПМИД 19158816 .
- ^ Кивисилд Т., Роотси С., Метспалу М., Мастана С., Калдма К., Парик Дж. и др. (февраль 2003 г.). «Генетическое наследие первых поселенцев сохраняется как в индийских племенах, так и в кастах» . Американский журнал генетики человека . 72 (2): 313–332. дои : 10.1086/346068 . ПМК 379225 . ПМИД 12536373 .
- ^ Jump up to: а б Сенгупта С., Животовский Л.А., Кинг Р., Мехди С.К., Эдмондс К.А., Чоу К.Э. и др. (февраль 2006 г.). «Полярность и временность распределения Y-хромосомы в Индии с высоким разрешением определяют как местное, так и экзогенное распространение и выявляют незначительное генетическое влияние скотоводов Центральной Азии» . Американский журнал генетики человека . 78 (2): 202–221. дои : 10.1086/499411 . ПМЦ 1380230 . ПМИД 16400607 .
- ^ «Анализ разнообразия Y-хромосомы в популяциях лингаятов и воккалиг Южной Индии». 2011. CiteSeerX 10.1.1.425.9132 .
- ^ Арункумар Г., Сориа-Эрнанц Д.Ф., Кавита В.Дж., Арун В.С., Шьяма А., Ашокан К.С. и др. (28 ноября 2012 г.). «Дифференциация населения мужских родов Южной Индии коррелирует с сельскохозяйственной экспансией, предшествовавшей кастовой системе» . ПЛОС Один . 7 (11): е50269. дои : 10.1371/journal.pone.0050269 . ПМК 3508930 . ПМИД 23209694 .
- ^ Уэллс 2001 .
- ^ Чаубей Г., Сингх М., Рай Н., Кариаппа М., Сингх К., Сингх А. и др. (январь 2016 г.). «Генетическое родство еврейского населения Индии» . Научные отчеты . 6 : 19166. Бибкод : 2016NatSR...619166C . дои : 10.1038/srep19166 . ПМЦ 4725824 . ПМИД 26759184 .
- ^ Ди Кристофаро Дж., Пеннарун Э., Мазьер С., Майрес Н.М., Лин А.А., Темори С.А. и др. (2013). «Афганский Гиндукуш: место, где сходятся потоки генов Евразийского субконтинента» . ПЛОС Один . 8 (10): е76748. Бибкод : 2013PLoSO...876748D . дои : 10.1371/journal.pone.0076748 . ПМЦ 3799995 . ПМИД 24204668 .
- ^ Фирасат и др. 2007 .
- ^ Jump up to: а б с Лако Х., Гайден Т., Регейро М., Ченнакришнайя С., Бухари А., Андерхилл П.А. и др. (октябрь 2012 г.). «Афганистан с точки зрения Y-хромосомы» . Европейский журнал генетики человека . 20 (10): 1063–1070. дои : 10.1038/ejhg.2012.59 . ПМК 3449065 . ПМИД 22510847 .
- ^ Jump up to: а б Хабер М., Платт Д.Э., Ашрафиан Бонаб М., Юханна С.К., Сориа-Эрнанц Д.Ф., Мартинес-Круз Б. и др. (2012). «Этнические группы Афганистана имеют общее Y-хромосомное наследие, структурированное историческими событиями» . ПЛОС Один . 7 (3): e34288. Бибкод : 2012PLoSO...734288H . дои : 10.1371/journal.pone.0034288 . ПМЦ 3314501 . ПМИД 22470552 .
- ^ Jump up to: а б с д и ж Ди Кристофаро Дж., Пеннарун Э., Мазьер С., Майрес Н.М., Лин А.А., Темори С.А. и др. (2013). «Афганский Гиндукуш: место, где сходятся потоки генов Евразийского субконтинента» . ПЛОС Один . 8 (10): е76748. Бибкод : 2013PLoSO...876748D . дои : 10.1371/journal.pone.0076748 . ПМЦ 3799995 . ПМИД 24204668 .
- ^ Jump up to: а б Лю Ш., Н., Илихаму, Р. Баке (2018). «Исследование генетического разнообразия трех изолированных популяций в Синьцзяне с использованием Y-SNP». Acta Anthropologica Sinica . 37 (1): 146–156.
- ^ Чжун 2010 .
- ^ Fedorova 2013 .
- ^ Карафет и др. 2010 .
- ^ Ким 2011 .
- ^ Jump up to: а б Судьял Х (октябрь 2013 г.). «Возврат к происхождению лембы: отслеживание происхождения Y-хромосом у южноафриканской и зимбабвийской лембы » журнал Южноафриканский медицинский журнал = Шведско -африканский 103 (12 Приложение 1): 1009–1013. дои : 10.7196/SAMJ.7297 . ПМИД 24300649 .
- ^ «Y-ДНК Haplgroup L-FT408126» .
- ^ Мсаиди С., Дюкурно А., Боетч Г., Лонгепье Г., Папа К., Аллиберт С. и др. (январь 2011 г.). «Генетическое разнообразие Коморских островов показывает, что раннее мореплавание стало основным фактором, определяющим биокультурную эволюцию человека в западной части Индийского океана» (PDF) . Европейский журнал генетики человека . 19 (1): 89–94. дои : 10.1038/ejhg.2010.128 . ПМК 3039498 . ПМИД 20700146 . Архивировано из оригинала (PDF) 7 октября 2016 г. Проверено 5 октября 2016 г.
- ^ Шах А.М., Таманг Р., Мурджани П., Рани Д.С., Говиндарадж П., Кулкарни Г. и др. (июль 2011 г.). «Индийские Сидди: африканские потомки с индийской примесью» . Американский журнал генетики человека . 89 (1): 154–161. дои : 10.1016/j.ajhg.2011.05.030 . ПМК 3135801 . ПМИД 21741027 .
- ^ Jump up to: а б с д Винченца Батталья, Симона Форнарино, Надя Аль-Захери и др . (2009), «Y-хромосомные свидетельства культурного распространения сельского хозяйства в Юго-Восточной Европе». Европейский журнал генетики человека (2009) 17, 820–830; дои:10.1038/ejhg.2008.249; опубликовано онлайн 24 декабря 2008 г.
- ^ Jump up to: а б с д и ж г Балановский О, Дибирова К, Дыбо А, Мудрак О, Фролова С, Почешхова Е и др. (октябрь 2011 г.). «Параллельная эволюция генов и языков в Кавказском регионе » Молекулярная биология и эволюция . 28 (10): 2905–2 дои : 10.1093/molbev/msr126 . ПМЦ 3355373 . ПМИД 21571925 .
- ^ ИЗОГГ 2016.
- ^ Jump up to: а б с д и ж г час я Юнусбаев Б, Мецпалу М, Ярве М, Кутуев И, Роотси С, Мецпалу Е и др. (январь 2012 г.). «Кавказ как асимметричный полупроницаемый барьер на пути древних миграций человека» . Молекулярная биология и эволюция . 29 (1): 359–365. дои : 10.1093/molbev/msr221 . ПМИД 21917723 .
- ^ Брук К.А. (лето 2014 г.). «Генетика крымских караимов» (PDF) . Карадениз Араштырмалары (Журнал черноморских исследований) . 11 (42): 69–84 на стр. 76. doi : 10.12787/KARAM859 .
- ^ «L-M349 YДерево» .
- ^ Jump up to: а б Платт Д.Э., Артиниан Х., Музая Ф., Халил В., Камар Ф.Г., Матису-Смит Э. и др. (апрель 2021 г.). «Аутосомная генетика и гаплогруппа Y-хромосомы L1b-M317 показывают, что марониты Горного Ливана являются постоянно неэмигрирующей популяцией» . Европейский журнал генетики человека . 29 (4): 581–592. дои : 10.1038/s41431-020-00765-x . ПМЦ 8182888 . ПМИД 33273712 .
- ^ «ДНК Азербайджана» . СемейноеДревоДНК . Джин бай Джин, ООО
- ^ «Германия-ЯДНА» . СемейноеДревоДНК . Джин бай Джин, ООО
- ^ «Греческий проект ДНК» . СемейноеДревоДНК . Джин бай Джин, ООО
- ^ «L-Y Гаплогруппа L» . СемейноеДревоДНК . Джин бай Джин, ООО
- ^ Чуан-Чао Ван, Сабина Рейнхольд, Алексей Калмыков, Антье Виссготт, Гвидо Брандт, Чунгвон Чжон, Оливия Черонет, Мэтью Ферри, Иадаоин Харни, Дениз Китинг, Свапан Маллик, Надин Роланд, Кристин Стюардсон, Анатолий Р. Кандимирович, Владимир Э. Маслов, Петренко Владимир Григорьевич, Эрлих Владимир Романович, Биаслан Ч. Атабиев, Рабадан Г. Магомедов, Филипп Л. Коль, Курт В. Альт, Сандра Л. Пихлер, Клаудия Герлинг, Харальд Меллер, Беник Варданян, Лариса Еганян, Алексей Д. Резепкин, Дирк Мариашк, Наталья Березина, Юлия Грески, Катарина Фукс , Корина Книппер, Стефан Шиффельс, Елена Балановская, Олег Балановский, Иэн Мэтисон, Томас Хайэм, Яков Б. Березин, Александра Бужилова, Виктор Трифонов, Рон Пинхаси, Андрей Б. Белинский, Дэвид Райх, Свенд Хансен, Йоханнес Краузе, Вольфганг Хаакан биоRxiv 322347; дои: https://doi.org/10.1101/322347 Опубликовано в журнале Nature Communications doi: 10.1038/s41467-018-08220-8.
- ^ Нарасимхан В.М., Паттерсон Н., Мурджани П., Роланд Н., Бернардос Р., Маллик С. и др. (06.09.2019). «Формирование человеческих популяций в Южной и Центральной Азии» . Наука . 365 (6457). bioRxiv 10.1101/292581 . дои : 10.1126/science.aat7487 . ISSN 0036-8075 . ПМЦ 6822619 . ПМИД 31488661 .
- ^ Скуртаниоти Э., Эрдал Ю.С., Франжипан М., Балосси Рестелли Ф., Йенер К.А., Пиннок Ф. и др. (май 2020 г.). «Геномная история от неолита до бронзового века в Анатолии, Северном Леванте и Южном Кавказе» . Клетка . 181 (5): 1158–1175.e28. дои : 10.1016/j.cell.2020.04.044 . hdl : 20.500.12154/1254 . ПМИД 32470401 . S2CID 219105572 .
- ^ Ингман Т., Эйзенманн С., Скуртаниоти Е., Акар М., Илгнер Дж., Гнекки Русконе Г.А. и др. (2021). «Человеческая мобильность в Телль-Атчане (Алалах), Хатай, Турция, во 2-м тысячелетии до нашей эры: интеграция изотопных и геномных данных» . ПЛОС Один . 16 (6): e0241883. Бибкод : 2021PLoSO..1641883I . дои : 10.1371/journal.pone.0241883 . ПМЦ 8244877 . ПМИД 34191795 .
- ^ Лазаридис И., Алпаслан-Руденберг С., Ачар А., Ачиккол А., Агеларакис А., Агикян Л. и др. (август 2022 г.). «Генетическая история Южной дуги: мост между Западной Азией и Европой» . Наука . 377 (6609): eabm4247. дои : 10.1126/science.abm4247 . ПМЦ 10064553 . ПМИД 36007055 .
- ^ Маргарян А., Лоусон Д.Д., Сикора М., Расимо Ф., Расмуссен С., Мольтке И. и др. (сентябрь 2020 г.). «Популяционная геномика мира викингов» . Природа . 585 (7825): 390–396. Бибкод : 2020Natur.585..390M . дои : 10.1038/s41586-020-2688-8 . hdl : 10852/83989 . ПМИД 32939067 . S2CID 221769227 .
- ^ Лазаридис И., Надель Д., Роллефсон Г., Мерретт Д.С., Роланд Н., Маллик С. и др. (август 2016 г.). «Геномный взгляд на происхождение земледелия на древнем Ближнем Востоке» . Природа . 536 (7617): 419–424. Бибкод : 2016Natur.536..419L . bioRxiv 10.1101/059311 . дои : 10.1038/nature19310 . ПМЦ 5003663 . ПМИД 27459054 .
- ^ «Гаплодерево Y-ДНК» . ДНК генеалогического древа использует дерево Консорциума Y-хромосомы и публикует его на своем веб-сайте.
Источники
[ редактировать ]- Абу-Амеро К.К., Хеллани А., Гонсалес А.М., Ларруга Х.М., Кабрера В.М., Андерхилл, Пенсильвания (сентябрь 2009 г.). «Разнообразие Y-хромосомы Саудовской Аравии и его связь с близлежащими регионами» . БМК Генетика . 10:59 . дои : 10.1186/1471-2156-10-59 . ПМК 2759955 . ПМИД 19772609 .
- Басу А., Мукерджи Н., Рой С., Сенгупта С., Банерджи С., Чакраборти М. и др. (октябрь 2003 г.). «Этническая Индия: геномный взгляд с особым упором на заселение и структуру» . Геномные исследования . 13 (10): 2277–2290. дои : 10.1101/гр.1413403 . ПМК 403703 . ПМИД 14525929 .
- Батталья В., Форнарино С., Аль-Захери Н., Оливьери А., Пала М., Майрес Н.М. и др. (июнь 2009 г.). «Y-хромосомные свидетельства культурного распространения сельского хозяйства в Юго-Восточной Европе» . Европейский журнал генетики человека . 17 (6): 820–830. дои : 10.1038/ejhg.2008.249 . ПМК 2947100 . ПМИД 19107149 .
- Махал Д.Г., Мацукас И.Г. (23 января 2018 г.). «Географическое происхождение этнических групп на Индийском субконтиненте: исследование древних следов с помощью гаплогрупп Y-ДНК» . Границы генетики . 9 :4. дои : 10.3389/fgene.2018.00004 . ПМЦ 5787057 . ПМИД 29410676 .
- Белеза С., Гужман Л., Лопес А., Алвес С., Гомес И., Гиузели М. и др. (март 2006 г.). «Микрофилогеографическая и демографическая история португальских мужских линий». Анналы генетики человека . 70 (Часть 2): 181–194. дои : 10.1111/j.1529-8817.2005.00221.x . ПМИД 16626329 . S2CID 4652154 .
- Боаттини А., Кастри Л., Сарно С., Усели А., Чиоффи М., Саццини М. и др. (март 2013 г.). «Вариации мтДНК в Восточной Африке раскрывают историю афро-азиатских групп». Американский журнал физической антропологии . 150 (3): 375–385. дои : 10.1002/ajpa.22212 . ПМИД 23283748 .
- Качиагли Л., Булаева К., Булаев О., Бертончини С., Тальоли Л., Пагани Л. и др. (декабрь 2009 г.). «Ключевая роль отцовской наследственности в формировании генетической изменчивости горцев Дагестана» . Журнал генетики человека . 54 (12): 689–694. дои : 10.1038/jhg.2009.94 . ПМИД 19911015 .
- Каденас А.М., Животовский Л.А., Кавалли-Сфорца Л.Л., Андерхилл П.А., Эррера Р.Дж. (март 2008 г.). «Разнообразие Y-хромосом характеризует Оманский залив» . Европейский журнал генетики человека . 16 (3): 374–386. дои : 10.1038/sj.ejhg.5201934 . ПМИД 17928816 .
- Чинниоглу С., Кинг Р., Кивисилд Т., Калфоглу Е., Атасой С., Каваллери Г.Л. и др. (январь 2004 г.). «Раскопки слоев гаплотипов Y-хромосомы в Анатолии». Генетика человека . 114 (2): 127–148. дои : 10.1007/s00439-003-1031-4 . ПМИД 14586639 . S2CID 10763736 .
- Кордо Р., Онгер Р., Бентли Г., Насидзе И., Сираджуддин С.М., Стоункинг М. (февраль 2004 г.). «Независимое происхождение индийских каст и племенных отцовских линий» . Современная биология . 14 (3): 231–235. Бибкод : 2004CBio...14..231C . дои : 10.1016/j.cub.2004.01.024 . ПМИД 14761656 . S2CID 5721248 .
- Дулик М.С., Жаданов С.И., Осипова Л.П., Аскапули А., Гау Л., Гоккумен О. и др. (февраль 2012 г.). «Вариации митохондриальной ДНК и Y-хромосомы свидетельствуют о недавнем общем происхождении коренных американцев и коренных алтайцев» . Американский журнал генетики человека . 90 (2): 229–246. дои : 10.1016/j.ajhg.2011.12.014 . ПМК 3276666 . ПМИД 22281367 .
- Эль-Сибай М., Платт Д.Э., Хабер М., Сюэ Ю., Юханна С.С., Уэллс Р.С. и др. (ноябрь 2009 г.). «Географическая структура Y-хромосомного генетического ландшафта Леванта: контраст между побережьем и сушей» . Анналы генетики человека . 73 (Часть 6): 568–581. дои : 10.1111/j.1469-1809.2009.00538.x . ПМЦ 3312577 . ПМИД 19686289 .
- Федорова С.А., Рейдла М, Метспалу Е, Метспалу М, Роотси С, Тамбетс К и др. (июнь 2013 г.). «Аутосомные и однородительские портреты коренного населения Саха (Якутии): значение для заселения Северо-Восточной Евразии» . Эволюционная биология BMC . 13 (127): 127. Бибкод : 2013BMCEE..13..127F . дои : 10.1186/1471-2148-13-127 . ПМЦ 3695835 . ПМИД 23782551 .
- Фирасат С., Халик С., Мохьюддин А., Папайоанну М., Тайлер-Смит С., Андерхилл П.А. и др. (январь 2007 г.). «Y-хромосомные доказательства ограниченного вклада греков в патанское население Пакистана» . Европейский журнал генетики человека . 15 (1): 121–126. дои : 10.1038/sj.ejhg.5201726 . ПМЦ 2588664 . ПМИД 17047675 .
- Флорес С., Мака-Мейер Н., Ларруга Дж.М., Кабрера В.М., Карадше Н., Гонсалес А.М. (2005). «Изоляты в коридоре миграций: анализ вариаций Y-хромосомы с высоким разрешением в Иордании» . Журнал генетики человека . 50 (9): 435–441. дои : 10.1007/s10038-005-0274-4 . ПМИД 16142507 .
- Ди Джакомо Ф., Лука Ф., Ананью Н., Чаварелла Дж., Корбо Р.М., Креста М. и др. (сентябрь 2003 г.). «Клинические закономерности разнообразия Y-хромосомы человека в континентальной Италии и Греции определяются эффектами дрейфа и основателя». Молекулярная филогенетика и эволюция . 28 (3): 387–395. дои : 10.1016/S1055-7903(03)00016-2 . ПМИД 12927125 .
- Гоккумен О (2008). Этноисторическое и генетическое обследование четырех поселений Центральной Анатолии . Пенсильванский университет. ISBN 978-0-549-80966-1 . Проверено 13 мая 2014 г.
- Хаммер М.Ф., Карафет Т.М., Парк Х., Омото К., Харихара С., Стоункинг М. и др. (январь 2006 г.). «Двойное происхождение японцев: общая основа Y-хромосом охотников-собирателей и фермеров» . Журнал генетики человека . 51 (1): 47–58. дои : 10.1007/s10038-005-0322-0 . ПМИД 16328082 . S2CID 6559289 .
- Карафет Т., Сюй Л., Ду Р., Ван В., Фэн С., Уэллс Р.С. и др. (сентябрь 2001 г.). «История отцовского населения Восточной Азии: источники, закономерности и микроэволюционные процессы» . Американский журнал генетики человека . 69 (3): 615–628. дои : 10.1086/323299 . ПМЦ 1235490 . ПМИД 11481588 .
- Карафет Т.М., Холлмарк Б., Кокс М.П., Судойо Х., Дауни С., Лансинг Дж.С. и др. (август 2010 г.). «Основное деление с востока на запад лежит в основе стратификации Y-хромосомы по всей Индонезии» . Молекулярная биология и эволюция . 27 (8): 1833–1844. дои : 10.1093/molbev/msq063 . ПМИД 20207712 .
- Ким Ш., Ким К.С., Шин DJ, Джин Х.Дж., Квак К.Д., Хан М.С. и др. (апрель 2011 г.). «Высокие частоты линий гаплогруппы Y-хромосомы O2b-SRY465 в Корее: генетический взгляд на заселение Кореи» . Исследовательская генетика . 2 (1): 10. дои : 10.1186/2041-2223-2-10 . ПМК 3087676 . ПМИД 21463511 .
- Лаппалайнен Т., Лайтинен В., Салмела Е., Андерсен П., Хуопонен К., Савонтаус М.Л. и др. (май 2008 г.). «Миграционные волны в регион Балтийского моря» . Анналы генетики человека . 72 (Часть 3): 337–348. дои : 10.1111/j.1469-1809.2007.00429.x . ПМИД 18294359 . S2CID 32079904 .
- Лармюсо М.Х., Вандерхейден Н., Джейкобс М., Куманс М., Ларно Л., Декорте Р. (март 2011 г.). «Микрогеографическое распространение вариаций Y-хромосомы в центрально-западном европейском регионе Брабант». Международная судебно-медицинская экспертиза. Генетика . 5 (2): 95–99. дои : 10.1016/j.fsigen.2010.08.020 . ПМИД 21036685 .
- Кивисилд Т., Роотси С., Метспалу М., Мастана С., Калдма К., Парик Дж. и др. (февраль 2003 г.). «Генетическое наследие первых поселенцев сохраняется как в индийских племенах, так и в кастах» . Американский журнал генетики человека . 72 (2): 313–332. дои : 10.1086/346068 . ПМК 379225 . ПМИД 12536373 .
- Лармюсо М.Х., Оттони С., Раймакерс Дж.А., Вандерхейден Н., Лармюсо Х.Ф., Декорте Р. (апрель 2012 г.). «Временная дифференциация западноевропейской линии Y-хромосомы: генеалогия как инструмент популяционной генетики человека» . Европейский журнал генетики человека . 20 (4): 434–440. дои : 10.1038/ejhg.2011.218 . ПМК 3306861 . ПМИД 22126748 .
- Лобов А.С. и др. (2009). «Структура генофонда башкирских субпопуляций» (PDF) (на русском языке). Архивировано из оригинала (PDF) 16 августа 2011 г.
- Луис Дж.Р., Роуолд Д.Д., Регейро М., Каейро Б., Чинниоглу С., Роузман С. и др. (март 2004 г.). «Левант против Африканского Рога: свидетельства двунаправленных коридоров миграции людей» . Американский журнал генетики человека . 74 (3): 532–544. дои : 10.1086/382286 . ПМЦ 1182266 . ПМИД 14973781 .
- Малярчук Б, Деренко М, Денисова Г, Хойт С, Возняк М, Гжибовский Т и др. (декабрь 2013 г.). «Y-хромосомное разнообразие у калмыков на этническом и племенном уровнях» . Журнал генетики человека . 58 (12): 804–811. дои : 10.1038/jhg.2013.108 . ПМИД 24132124 .
- Пихлер И., Мюллер Дж.К., Стефанов С.А., Де Гранди А., Вольпато С.Б., Пинггера Г.К. и др. (декабрь 2009 г.). «Генетическая структура современных изолированных популяций Южного Тироля, выявленная путем анализа полиморфизмов Y-хромосомы, мтДНК и Alu. 2006». Биология человека . 81 (5–6): 875–898. дои : 10.3378/027.081.0629 . ПМИД 20504204 . S2CID 46073270 .
- Камар Р., Аюб К., Мохьюддин А., Хельгасон А., Мажар К., Мансур А. и др. (май 2002 г.). «Вариации ДНК Y-хромосомы в Пакистане» . Американский журнал генетики человека . 70 (5): 1107–1124. дои : 10.1086/339929 . ПМЦ 447589 . ПМИД 11898125 .
- Регейру М., Каденас А.М., Гайден Т., Андерхилл П.А., Эррера Р.Дж. (2006). «Иран: трехконтинентальная связь для миграции, вызванной Y-хромосомой». Наследственность человека . 61 (3): 132–143. дои : 10.1159/000093774 . ПМИД 16770078 . S2CID 7017701 .
- Саху С., Сингх А., Химабинду Г., Банерджи Дж., Ситалакшими Т., Гайквад С. и др. (январь 2006 г.). «Предыстория индийских Y-хромосом: оценка сценариев демического распространения» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (4): 843–848. Бибкод : 2006PNAS..103..843S . дои : 10.1073/pnas.0507714103 . ПМК 1347984 . ПМИД 16415161 .
- Санчес Дж. Дж., Халленберг С., Бёрстинг С., Эрнандес А., Морлинг Н. (июль 2005 г.). «Высокие частоты линий Y-хромосомы, характеризующиеся E3b1, DYS19-11, DYS392-12 у сомалийских мужчин» . Европейский журнал генетики человека . 13 (7): 856–866. дои : 10.1038/sj.ejhg.5201390 . ПМИД 15756297 .
- Скоццари Р., Кручиани Ф., Панграцио А., Сантоламазца П., Вона Г., Морал П. и др. (сентябрь 2001 г.). «Изменения Y-хромосомы человека в западном Средиземноморье: последствия для заселения региона» Иммунология человека . 62 (9): 871–884. CiteSeerX 10.1.1.408.4857 . дои : 10.1016/S0198-8859(01) 00286-5 ПМИД 11543889 .
- Семино О., Пассарино Г., Офнер П.Дж., Лин А.А., Арбузова С., Бекман Л.Е. и др. (ноябрь 2000 г.). «Генетическое наследие палеолитического Homo sapiens sapiens у современных европейцев: взгляд на Y-хромосому». Наука . 290 (5494): 1155–1159. Бибкод : 2000Sci...290.1155S . дои : 10.1126/science.290.5494.1155 . ПМИД 11073453 .
- Сенгупта С., Животовский Л.А., Кинг Р., Мехди С.К., Эдмондс К.А., Чоу К.Э. и др. (февраль 2006 г.). «Полярность и временность распределения Y-хромосомы в Индии с высоким разрешением определяют как местное, так и экзогенное распространение и выявляют незначительное генетическое влияние скотоводов Центральной Азии» . Американский журнал генетики человека . 78 (2): 202–221. дои : 10.1086/499411 . ПМЦ 1380230 . ПМИД 16400607 .
- Шах А.М., Таманг Р., Мурджани П., Рани Д.С., Говиндарадж П., Кулкарни Г. и др. (июль 2011 г.). «Индийские Сидди: африканские потомки с индийской примесью» . Американский журнал генетики человека . 89 (1): 154–161. дои : 10.1016/j.ajhg.2011.05.030 . ПМК 3135801 . ПМИД 21741027 .
- Тархнишвили Д, Гавашелишвили А, Мурцхваладзе М, Габелая М, Тевзадзе Г (2014). «Человеческие отцовские линии, языки и окружающая среда на Кавказе». Биология человека . 86 (2): 113–130. дои : 10.3378/027.086.0205 . ПМИД 25397702 . S2CID 7733899 .
- Тансим И., Тангарадж К., Чауби Дж., Сингх В.К., Бхаскар Л.В., Редди Б.М. и др. (август 2006 г.). «Генетическое сходство среди низших каст и племенных групп Индии: вывод на основе Y-хромосомы и митохондриальной ДНК» . БМК Генетика . 7:42 . дои : 10.1186/1471-2156-7-42 . ПМЦ 1569435 . ПМИД 16893451 .
- Томас М.Г., Барнс И., Уил М.Э., Джонс А.Л., Форстер П., Брэдман Н. и др. (январь 2008 г.). «Новые генетические данные подтверждают изоляцию и дрейф общин ладинов в Южно-Тирольских Альпах, но не подтверждают их древнее происхождение на Ближнем Востоке» . Европейский журнал генетики человека . 16 (1): 124–134. дои : 10.1038/sj.ejhg.5201906 . ПМИД 17712356 .
- Уил М.Э., Епископосян Л., Джагер Р.Ф., Оганесян Н., Худоян А., Бербедж-Холл О. и др. (декабрь 2001 г.). «Армянские гаплотипы Y-хромосомы демонстрируют сильную региональную структуру внутри одной этнонациональной группы». Генетика человека . 109 (6): 659–674. дои : 10.1007/s00439-001-0627-9 . ПМИД 11810279 . S2CID 23113666 .
- Уэллс Р.С., Юлдашева Н., Рузибакиев Р., Андерхилл П.А., Евсеева И., Блю-Смит Дж. и др. (август 2001 г.). «Центр Евразии: континентальный взгляд на разнообразие Y-хромосомы» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (18): 10244–10249. Бибкод : 2001PNAS...9810244W . дои : 10.1073/pnas.171305098 . ПМК 56946 . ПМИД 11526236 .
- Янг К.Л., Сан Дж., Дека Р., Кроуфорд М.Х. (август 2011 г.). «Отцовская генетическая история баскского населения Испании». Биология человека . 83 (4): 455–475. дои : 10.3378/027.083.0402 . hdl : 1808/16387 . ПМИД 21846204 . S2CID 3191418 .
- Заллуа П.А., Сюэ Ю., Халифе Дж., Махул Н., Дебиан Л., Платт Д.Э. и др. (апрель 2008 г.). «Разнообразие Y-хромосомы в Ливане структурировано недавними историческими событиями» . Американский журнал генетики человека . 82 (4): 873–882. дои : 10.1016/j.ajhg.2008.01.020 . ПМЦ 2427286 . ПМИД 18374297 .
Веб-источники
[ редактировать ]- ^ Jump up to: а б с Кран Т. «Проект дерева Y-ДНК FTDNA (AKA YTree)» . ДНК генеалогического древа . Архивировано из оригинала 15 августа 2015 г. Проверено 1 января 2013 г.
- ^ Jump up to: а б Хенсон Г., Гречдакян П.О. (2013). «L - Проект Y-гаплогруппы L» . ДНК генеалогического древа . Проверено 1 января 2013 г.
Источники таблиц преобразования
[ редактировать ]- Капелли С., Уилсон Дж. Ф., Ричардс М., Стампф М. П., Гратрикс Ф., Оппенгеймер С. и др. (февраль 2001 г.). «Преимущественно коренное отцовское наследие австронезийскоязычных народов островной Юго-Восточной Азии и Океании» . Американский журнал генетики человека . 68 (2): 432–443. дои : 10.1086/318205 . ПМЦ 1235276 . ПМИД 11170891 .
- Хаммер М.Ф., Карафет Т.М., Редд А.Дж., Джарджанази Х., Сантакьяра-Бенерекетти С., Судьялл Х. и др. (июль 2001 г.). «Иерархические закономерности глобального разнообразия Y-хромосом человека» . Молекулярная биология и эволюция . 18 (7): 1189–1203. doi : 10.1093/oxfordjournals.molbev.a003906 . ПМИД 11420360 .
- Джоблинг М.А., Тайлер-Смит С. (август 2000 г.). «Новое использование новых гаплотипов Y-хромосомы человека, болезни и отбор». Тенденции в генетике . 16 (8): 356–362. дои : 10.1016/S0168-9525(00)02057-6 . ПМИД 10904265 .
- Калайджиева Л., Калафель Ф., Джоблинг М.А., Ангеличева Д., де Книфф П., Россер З.Х. и др. (февраль 2001 г.). «Закономерности меж- и внутригруппового генетического разнообразия у Vlax Roma, выявленные с помощью Y-хромосомы и линий митохондриальной ДНК» . Европейский журнал генетики человека . 9 (2): 97–104. дои : 10.1038/sj.ejhg.5200597 . ПМИД 11313742 . S2CID 21432405 .
- Карафет Т., Сюй Л., Ду Р., Ван В., Фэн С., Уэллс Р.С. и др. (сентябрь 2001 г.). «История отцовского населения Восточной Азии: источники, закономерности и микроэволюционные процессы» . Американский журнал генетики человека . 69 (3): 615–628. дои : 10.1086/323299 . ПМЦ 1235490 . ПМИД 11481588 .
- Карафет Т.М., Мендес Ф.Л., Мейлерман М.Б., Андерхилл П.А., Зегура С.Л., Хаммер М.Ф. (май 2008 г.). «Новые бинарные полиморфизмы изменяют форму и увеличивают разрешение дерева гаплогрупп Y-хромосомы человека» . Геномные исследования . 18 (5): 830–838. дои : 10.1101/гр.7172008 . ПМК 2336805 . ПМИД 18385274 .
- Су Б., Сяо Дж., Андерхилл П., Дека Р., Чжан В., Акей Дж. и др. (декабрь 1999 г.). «Y-хромосома свидетельствует о миграции современных людей на север в Восточную Азию во время последнего ледникового периода» . Американский журнал генетики человека . 65 (6): 1718–1724. дои : 10.1086/302680 . ПМЦ 1288383 . ПМИД 10577926 .
- Андерхилл П.А., Шен П., Лин А.А., Джин Л., Пассарино Г., Ян В.Х. и др. (ноябрь 2000 г.). «Вариации последовательности Y-хромосомы и история человеческих популяций». Природная генетика . 26 (3): 358–361. дои : 10.1038/81685 . ПМИД 11062480 . S2CID 12893406 .
- Yunusbaev BB (2006). ПОПУЛЯЦИОННО-ГЕНЕТИЧЕСКОЕ ИССЛЕДОВАНИЕ НАРОДОВ ДАГЕСТАНА ПО ДАННЫМ О ПОЛИМОРФИЗМЕ У-ХРОМОСОМЫ И АТД-ИНСЕРЦИЙ [ Population-genetic study of the peoples of Dagestan on the data on Y-chromosome and ATD-insertion polymorphism ] (PDF) (PhD). Moscow: Russian Academy of Sciences. Archived from the original (PDF) on 5 February 2007.
- Чжун Х., Ши Х., Ци С.Б., Дуань З.Ю., Тан П.П., Цзинь Л. и др. (январь 2011 г.). «Расширенное исследование Y-хромосомы позволяет предположить, что современные люди мигрировали в Восточную Азию северным путем после ледникового периода» . Молекулярная биология и эволюция . 28 (1): 717–727. дои : 10.1093/molbev/msq247 . ПМИД 20837606 .
Внешние ссылки
[ редактировать ]- ИЗОГГ, [1]
- Генная база (2006). «Учебные пособия по генной базе: узнайте о гаплогруппе L Y-хромосомы» . Архивировано из оригинала 23 октября 2012 г.
- Распространение гаплогруппы L по данным National Geographic.
- Индийский генеалогический проект
- Y Гаплогруппа L