Гаплогруппа А (Y-ДНК)
| Гаплогруппа А | |
|---|---|
 | |
| Возможное время происхождения | 270 000 лет назад, [1] [2] 275 000 BP (303 000–241 000 BP), [3] [4] 291 000 лет назад [5] |
| Возраст слияния | 275 000 BP (разделено с другими линиями) [6] |
| Возможное место происхождения | Северо-Западная Африка , Центральная Африка [7] |
| Предок | Человеческий Y-MRCA (A00-T) |
| Потомки | первичный: A00 (AF6/L1284), A0-T (Их субклады включают гаплогруппы A00a, A00b, A00c, A0 , A1 , A1a , A1b , A1b1 и BT .) |
Гаплогруппа А — гаплогруппа ДНК Y-хромосомы человека , которая включает в себя все живые Y-хромосомы человека. Носители дошедших до нас субкладов гаплогруппы А почти исключительно встречаются в Африке (или среди африканской диаспоры ), в отличие от гаплогруппы ВТ , носители которой участвовали в миграции из Африки ранних современных людей . Известные ветви гаплогруппы A — A00 , A0 , A1a и A1b1 ; эти ветви связаны лишь очень отдаленно и не более тесно связаны друг с другом, чем с гаплогруппой BT.
Источник
[ редактировать ]
Хотя существуют терминологические проблемы с определением ее как гаплогруппы, гаплогруппа А стала означать «основную гаплогруппу» (то есть современных человеческих популяций ); оно не определяется какой-либо мутацией, а относится к любой гаплогруппе, не происходящей от гаплогруппы BT ; другими словами, это определяется отсутствием определяющей мутации этой группы (M91). Согласно этому определению, гаплогруппа A включает все мутации, которые произошли между самым последним общим предком Y-хромосомы (по оценкам, около 270 тысяч лет назад ) и мутацией, определяющей гаплогруппу BT (по оценкам, около 140–150 тысяч лет назад). [8] включая любые существующие субклады, которые, возможно, еще предстоит открыть.
Носители гаплогруппы А (т.е. отсутствие определяющей мутации гаплогруппы BT) были обнаружены в районах проживания охотников-собирателей Южной Африки, особенно среди народа сан . Кроме того, большинство базальных линий L0 митохондриальной ДНК также в значительной степени ограничены San. Однако линии A Южной Африки являются субкладами линий A, обнаруженными в других частях Африки, что позволяет предположить, что субгаплогруппы A прибыли в Южную Африку из других мест. [9]
Две наиболее базальные линии гаплогруппы A, A0 и A1 (до объявления об открытии гаплогруппы A00 в 2013 году) были обнаружены в Западной Африке, Северо-Западной Африке и Центральной Африке. Кручиани и др. (2011) предполагают, что эти линии могли возникнуть где-то между Центральной и Северо-Западной Африкой. [10] Скоццари и др. ] в северо-западном квадранте Африканского континента (2012) также поддержали «гипотезу о происхождении гаплогруппы A1b [ т.е. A0 ». [11]
Гаплогруппа A1b1b2 была обнаружена среди древних окаменелостей, раскопанных в заливе Балито в Квазулу-Натале , Южная Африка , которые были датированы примерно 2149-1831 годами назад (2/2; 100%). [12]
Распределение
[ редактировать ]Согласно определению гаплогруппы А как «не- BT », она почти полностью ограничена Африкой зарегистрировано очень небольшое количество ее носителей , хотя в Европе и Западной Азии .
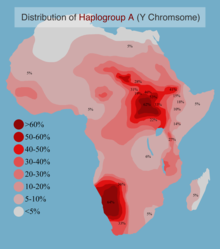
Клада достигает своей самой высокой современной частоты среди бушменских популяций охотников-собирателей в Южной Африке , за которыми следуют многие нилотские группы в Восточной Африке . Однако самые старые субклады гаплогруппы А встречаются исключительно в Центральной и Северо-Западной Африке они (и, соответственно, отцовский предок , откуда, как полагают, и произошли современных людей). Оценки его временной глубины сильно различались: в отдельных исследованиях 2013 года они составляли либо около 190 тысяч лет назад, либо около 140 тысяч лет назад. [10] [13] и с включением ранее неизвестной гаплогруппы «A00» примерно в 270 тысяч лет назад в исследованиях 2015 года. [14] [15]
Клада также с заметной частотой наблюдалась у некоторых популяций в Эфиопии , а также у некоторых групп пигмеев в Центральной Африке и реже у говорящих на Нигере и Конго , которые в основном принадлежат к кладе E1b1a . гаплогруппа Е возникла в Северо-Восточной Африке. Считается, что [16] и позже был завезен в Западную Африку, откуда около 5000 лет назад распространился в Центральную, Южную и Юго-Восточную Африку с экспансией банту . [17] [18] По данным Вуда и др. (2005) и Роза и др. (2007), такие относительно недавние перемещения населения из Западной Африки изменили ранее существовавшее хромосомное разнообразие Y популяции в Центральной, Южной и Юго-Восточной Африке, заменив предыдущие гаплогруппы в этих областях ныне доминирующими линиями E1b1a. Однако сегодня в этих регионах можно наблюдать следы предковых жителей по наличию гаплогрупп Y-ДНК A-M91 и B-M60, которые распространены в некоторых реликтовых популяциях, таких как пигмеи мбути и койсаны . [19] [20] [21]
| Африка | ||
| Исследуемая популяция | Частота. (в %) | |
| [20] | Цумкве Сан (Намибия) | 66% |
| [20] | Нама (Намибия) | 64 |
| [22] | Динка (Судан) | 62 |
| [22] | Шиллук (Судан) | 53 |
| [22] | Нуба (Судан) | 46 |
| [23] | Хойсан | 44 |
| [24] [25] | Эфиопские евреи | 41 |
| [20] [24] | !Если /Секеле | ~40 |
| [22] | Боргу (Судан) | 35 |
| [22] | Нуэр (Судан) | 33 |
| [22] | Мех (Судан) | 31 |
| [20] | Масаи (Кения) | 27 |
| [26] | Нара (Эритрея) | 20 |
| [22] | Масалит (Судан) | 19 |
| [27] | эфиопы | 14 |
| [20] | Банту (Кения) | 14 |
| [22] | Мандара (Камерун) | 14 |
| [24] | Хауса (Судан) | 13 |
| [24] | Хве (Южная Африка) | 12 |
| [20] | Фульбе (Камерун) | 12 |
| [28] | Дама (Намибия) | 11 |
| [26] | Оромо (Эфиопия) | 10 |
| [20] | Кунама (Эритрея) | 10 |
| [27] | Южносемитский (Эфиопия) | 10 |
| Арабы (Египет) | 3 | |
В составной выборке из 3551 африканского мужчины частота гаплогруппы А составила 5,4%. [29] Самые высокие частоты гаплогруппы А зарегистрированы среди койсанцев Южной Африки, Бета-Израиля и нило-сахарцев из Судана.
Северная Америка
[ редактировать ]1 мужчина-афроамериканец из Лакросса, Висконсин, США, Мозес, Рамон, A00, A00-AF8
Африка
[ редактировать ]Северная Африка
[ редактировать ]В Северной Африке гаплогруппа А практически отсутствует. Его субклад A1 наблюдался на следовых частотах среди марокканцев.
Верхний Нил
[ редактировать ]Гаплогруппа A3b2-M13 распространена среди жителей Южного Судана (53%), [22] особенно суданцы динка (61,5%). [30] Гаплогруппа A3b2-M13 также наблюдалась в другой выборке населения Южного Судана с частотой 45% (18/40), включая 1/40 A3b2a-M171. [23]
Ниже по течению долины Нила субклад A3b2 также наблюдался с очень низкой частотой в выборке египетских мужчин (3%).
Западная Африка
[ редактировать ]Восемь мужчин из Гвинеи-Бисау , два мужчины из Нигера , один мужчина из Мали и один мужчина из Кабо-Верде несли гаплогруппу A1a . [31]
Центральная Африка
[ редактировать ]Гаплогруппа A3b2-M13 наблюдалась в популяциях северного Камеруна (2/9 = 22% Тупури , [20] 4/28 = 14% Мандары , [20] 2/17 = 12% фульбе [24] ) и восточной ДРК (2/9 = 22% Алура , [20] 1/18 = 6% Юг , [20] 1/47 = 2% Коза [20] ).
Гаплогруппа A-M91 (xA1a-M31, A2-M6/M14/P3/P4, A3-M32) наблюдалась у народа бакола на юге Камеруна (3/33 = 9%). [20]
Без тестирования на какой-либо субклад Y-ДНК гаплогруппы А наблюдалась в образцах нескольких популяций Габона , включая 9% (3/33) выборки Бака , 3% (1/36) выборки Ндуму , 2 % (1/46) выборки Думы , 2% (1/57) выборки Нзеби и 2% (1/60) выборки Цого . [18]
Восточная Африка
[ редактировать ]Великие африканские озера
[ редактировать ]Банту в Кении (14%, Луис и др., 2004 г.) и иракцы в Танзании (от 3/43 = 7,0% (Луис и др., 2004 г.) до 1/6 = 17% (Найт и др., 2003 г.)).
Африканский Рог
[ редактировать ]Гаплогруппа А встречается с низкой и умеренной частотой на Африканском Роге. Клада наблюдается с наибольшей частотой среди 41% выборки Бета-Израиля , встречаясь среди 41% одной выборки из этой популяции (Cruciani et al. 2002). В других частях региона гаплогруппа А обнаружена в 14,6% (7/48) выборки из Амхары . [28] 10,3% (8/78) образца оромо , [28] и 13,6% (12/88) другой выборки из Эфиопии. [23]
Южная Африка
[ редактировать ]Одно исследование 2005 года обнаружило гаплогруппу А в образцах различных койсаноязычных племен с частотой от 10% до 70%. [20] Эта конкретная гаплогруппа не была обнаружена у хадзабе из Танзании. [ нужна ссылка ] популяция, которую иногда считают остатком койсанидского населения позднего каменного века.
Азия
[ редактировать ]В Азии гаплогруппа А наблюдалась с низкими частотами в Малой Азии и на Ближнем Востоке среди эгейских турок, палестинцев, иорданцев, йеменцев. [32]
Европа
[ редактировать ]A3a2 (A-M13; ранее A3b2) наблюдался с очень низкой частотой на некоторых островах Средиземноморья. Без тестирования на какой-либо субклад гаплогруппа А была обнаружена в выборке греков из Митилини на острове Лесбос в Эгейском море. [32] и в образцах португальского языка из южной Португалии, центральной Португалии и Мадейры . [33] Авторы одного исследования сообщили об обнаружении гаплогруппы А у 3,1% (2/65) выборки киприотов . [34] хотя они окончательно не исключили возможность того, что любой из этих людей может принадлежать к редкому субкладу гаплогруппы BT , включая гаплогруппу CT .
Субклады
[ редактировать ]А00 (А00-AF6)
[ редактировать ]Мендес и др. (2013) объявили об открытии ранее неизвестной гаплогруппы, для которой предложили обозначение «А00». [35] «Генотипирование образца ДНК, который был отправлен в коммерческий центр генетического тестирования, показало, что Y-хромосома этого афроамериканца несет в себе предковое состояние всех известных SNP Y-хромосомы. Чтобы дополнительно охарактеризовать эту линию, которую мы назвали A00, [36] для предлагаемой номенклатуры)"; "Мы переименовали базальную ветвь в Cruciani et al. [2011] как A0 (ранее A1b), а указанную в настоящее время линию называют A00. Для глубоких ответвлений, обнаруженных в будущем, мы предлагаем продолжить номенклатуру А000 и так далее». [14] [15] так примерно современен известному внешнему виду самых ранних известных анатомически современных людей , таких как Джебель Ирхуд . [37] A00 также иногда называют «Y-хромосомой Перри» (или просто «Y Перри»). Эта ранее неизвестная гаплогруппа была обнаружена в 2012 году в Y-хромосоме афроамериканца, который предоставил свою ДНК для коммерческого генеалогического анализа. [38] Последующее обнаружение других самцов, принадлежащих к A00, привело к реклассификации Y Перри в A00a (A-L1149).
Позже исследователи обнаружили, что A00 принадлежал 11 мужчинам племени Мбо из Западного Камеруна (банту) (из выборки в 174 человека (6,32%). [39] Последующие исследования показали, что общий уровень A00 был даже выше среди Mbo, т.е., как позже выяснилось, 9,3% (8 из 86) попадали в категорию A00b (A-A4987).
Дальнейшие исследования, проведенные в 2015 году, показывают, что современное население с самой высокой концентрацией A00 — это бангва (или нве), группа Камеруна , говорящая на языке емба ( банту Грассфилдс ): 27 из 67 (40,3%) образцов были положительными на A00a. (Л1149). Один человек из бангва не вписывался ни в A00a, ни в A00b. [40]
Генетики секвенировали полногеномные данные ДНК четырех человек, похороненных на месте Шум-Лака в Камеруне между 8000–3000 лет назад, которые были наиболее генетически похожи на мбути пигмеев . Один человек был носителем глубоко дивергентной гаплогруппы А00 Y-хромосомы. [41]
А0 (А-В148)
[ редактировать ]Названия гаплогрупп «A-V148» и «A-CTS2809/L991» относятся к одной и той же гаплогруппе.
А0 встречается только у пигмеев Бакола (Южный Камерун ) в 8,3% и у берберов из Алжира в 1,5%. [10] Также встречается в Гане . [11] [ не удалось пройти проверку ]
А1а (А-М31)
[ редактировать ]Субклад A1a (M31) был обнаружен примерно в 2,8% (8/282) пула из семи выборок различных этнических групп Гвинеи-Бисау , особенно среди папель-манжако-манканья (5/64 = 7,8%). [19] В более раннем исследовании, опубликованном в 2003 году, Gonçalves et al. сообщили об обнаружении A1a-M31 в 5,1% (14/276) образцов из Гвинеи-Бисау и в 0,5% (1/201) пары образцов из Кабо-Верде . [42] Авторы другого исследования сообщили об обнаружении гаплогруппы A1a-M31 у 5% (2/39) выборки мандинка из Сенегамбии и 2% (1/55) выборки догонов из Мали . [20] Гаплогруппа A1a-M31 также была обнаружена у 3% (2/64) выборки берберов из Марокко. [24] и 2,3% (1/44) выборки неустановленной этнической принадлежности из Мали . [23]
В 2007 году семь мужчин из Йоркшира , Англия, носивших необычную фамилию Ревис, были идентифицированы как принадлежащие к субкладу A1a (M31). Было обнаружено, что у этих мужчин был общий предок по мужской линии из 18 века, но никакой предыдущей информации об африканском происхождении не было известно. [29]
В 2023 году Лакросс, Висконсин, 1 мужчина, A1a-M31, Моисей, Рамон. [43]
А1б1а1а (А-М6)
[ редактировать ]Субклад A1b1a1a (M6; ранее A2 и A1b1a1a-M6) обычно встречается среди койсанских народов. Авторы одного исследования сообщили об обнаружении гаплогруппы A-M6(xA-P28) у 28% (8/29) выборки Цумкве Сан и 16% (5/32) выборки !Кунг /Секеле, а также гаплогруппы A2b-P28 в 17% (5/29) образца Цумкве Сан, 9% (3/32) образца !Кунг /Секеле, 9% (1/11) образца Нама и 6% (1/18) образца Дамы . [20] Авторы другого исследования сообщили об обнаружении гаплогруппы A2 у 15,4% (6/39) выборки койсанских мужчин, включая 5/39 A2-M6/M14/M23/M29/M49/M71/M135/M141(xA2a-M114). ) и 1/39 А2а-М114. [23]
А1б1б (А-М32)
[ редактировать ]Клада A1b1b (M32; ранее A3) содержит наиболее густонаселенные ветви гаплогруппы A и в основном встречается в Восточной Африке и Южной Африке .
А1б1б1 (А-М28)
[ редактировать ]Субклад (считающийся отдельной гаплогруппой) A1b1b1 (M28; ранее A3a) лишь изредка наблюдался на Африканском Роге . В 5% (1/20) смешанной выборки носителей южносемитских языков из Эфиопии [20] 1,1% (1/88) выборки эфиопов, [23] и 0,5% (1/201) у сомалийцев . [16] это также наблюдалось в Восточной, Центральной и Южной Аравии. Текущие результаты, согласно FTDNA, позволяют предположить, что некоторые ветви, такие как A-V1127, возникли в Аравии. Кроме того, как предполагают эксперты, как видно из TMRCA в дереве Yfull, эта гаплогруппа, должно быть, пережила период «узкого места», когда люди, представляющие эту гаплогруппу, претерпели своего рода вымирание и их численность резко сократилась. Примечательно, что у несемитских носителей этой гаплогруппы нет ни у кои-сан, нилотов, ни у кушитов.
А1b1b2a (А-М51)
[ редактировать ]Субклад A1b1b2a (M51; ранее A3b1) наиболее часто встречается среди койсанских народов (6/11 = 55% нама , [20] 11/39 = 28% койсан, [23] 7/32 = 22% !Если /Секеле, [20] 6/29 = 21% Цумкве Сан, [20] 1/18 = 6% шанс [20] ). Однако это также было обнаружено с меньшей частотой среди народов банту Южной Африки , включая 2/28 = 7% сото-тсвана , [20] 3/53 = 6% некойсанских жителей Южной Африки, [23] 4/80 = 5% коса , [20] и 1/29 = 3% зулусов . [20]
А1b1b2b (А-М13)
[ редактировать ]Субклад A1b1b2b (M13; ранее A3b2) в основном распространен среди нилотских популяций в Восточной Африке и северном Камеруне. Он отличается от субклада А, обнаруженного в койсанских образцах и лишь отдаленно связанного с ними (на самом деле это лишь один из многих субкладов внутри гаплогруппы А). Это открытие предполагает древнее расхождение.
В Судане гаплогруппа A-M13 обнаружена у 28/53 = 52,8% южных суданцев , 13/28 = 46,4% нубийцев центрального Судана, 25/90 = 27,8% западных суданцев , 4/32 = 12,5% местных жителей хауса и 5/216 = 2,3% жителей Северного Судана. [44]
В Эфиопии одно исследование сообщило об обнаружении гаплогруппы A-M13 у 14,6% (7/48) выборки Амхары и 10,3% (8/78) выборки Оромо . [28] В другом исследовании сообщалось об обнаружении гаплогруппы A3b2b-M118 у 6,8% (6 из 88) и гаплогруппы A3b2*-M13(xA3b2a-M171, A3b2b-M118) у 5,7% (5 из 88) смешанной выборки эфиопов, что составляет всего 12,5% (11/88) А3б2-М13. [23]
Гаплогруппа A-M13 также иногда наблюдалась за пределами Центральной и Восточной Африки, например, в Эгейском регионе Турции . (2/30 = 6,7%) [45] ), йеменские евреи (1/20 = 5% [25] ), Египет (4/147 = 2,7%, [27] 3/92 = 3.3% [20] ), палестинские арабы (2/143 = 1,4% [46] ), Сардиния (1/77 = 1,3%, [47] 1/22 = 4.5% [23] ), столица Иордании Амман % (1/101=1 [48] ) и Оман (1/121 = 0,8% [27] ).
Гаплогруппа A-M13 была обнаружена среди трех окаменелостей периода неолита, раскопанных на стоянке Кадрука в Судане. [49]

Гаплогруппа A-M13 также была обнаружена у мужчины, ставшего жертвой извержения Везувия в Помпеях. [50]
Филогенетика
[ редактировать ]Филогенетическая история
[ редактировать ]До 2002 года в научной литературе существовало как минимум семь систем наименования филогенетического дерева Y-хромосомы. Это привело к значительной путанице. В 2002 году основные исследовательские группы объединились и сформировали Консорциум Y-хромосомы (YCC). Они опубликовали совместный документ, в котором было создано единое новое дерево, которое все согласились использовать. Позже группа гражданских ученых, интересующихся популяционной генетикой и генетической генеалогией, сформировала рабочую группу для создания любительского дерева, стремясь быть прежде всего своевременным. В таблице ниже собраны все эти работы, относящиеся к знаковому дереву YCC 2002 года. Это позволяет исследователю, просматривающему старую опубликованную литературу, быстро перемещаться между номенклатурами.
Первоначальное секвенирование Y-хромосомы человека позволило предположить, что первое разделение генеалогического древа Y-хромосомы произошло в результате мутаций, которые отделили гаплогруппу BT от Y-хромосомного Адама и гаплогруппы A в более широком смысле. [51] Впоследствии также стало известно о многих промежуточных расщеплениях между Y-хромосомным Адамом и БТ.
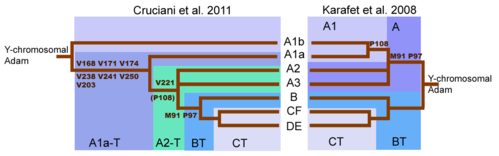
Серьезный сдвиг в понимании дерева Y-ДНК произошел с публикацией ( Cruciani 2011 ). Хотя маркер SNP M91 считался ключом к идентификации гаплогруппы BT, выяснилось, что область, окружающая M91, представляет собой горячую точку мутаций, склонную к повторяющимся обратным мутациям. Более того, участок 8T гаплогруппы A представлял собой наследственное состояние M91, а участок 9T гаплогруппы BT - производное состояние, возникшее после вставки 1T. Это объяснило, почему субклады A1b и A1a, самые глубокие ветви гаплогруппы A, оба обладали растяжением 8T. Точно так же маркер P97, который также использовался для идентификации гаплогруппы A, обладал наследственным состоянием в гаплогруппе A, но производным состоянием в гаплогруппе BT. [10] В конечном итоге склонность M91 к обратной мутации и (следовательно) его ненадежность привели к тому, что ISOGG в 2016 году отклонил M91 как определяющий SNP. [52] И наоборот, P97 был сохранен как определяющий маркер гаплогруппы BT.
| YCC 2002/2008 (сокращенно) | (а) | (б) | (с) | (г) | (е) | (г) | (или) | YCC 2002 (от руки) | YCC 2005 (от руки) | YCC 2008 (от руки) | YCC 2010r (от руки) | ИСОГГ 2006 г. | ИСОГГ 2007 | ИСОГГ 2008 | ИСОГГ 2009 | ИСОГГ 2010 | ИЗОГГ 2011 | ИЗОГГ 2012 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| А-М31 | 7 | я | 1А | 1 | – | H1 | А | А1 | А1 | А1 | А1а | А1 | А1 | А1а | А1а | А1а | А1а | А1а |
| А-М6 | 27 | я | 2 | 3 | – | H1 | А | А2* | А2 | А2 | А2 | А2 | А2 | А2 | А2 | А2 | А2 | А1б1а1а |
| А-М114 | 27 | я | 2 | 3 | – | H1 | А | А2а | А2а | А2а | А2а | А2а | А2а | А2а | А2а | А2а | А2а | А1б1а1а1а |
| А-П28 | 27 | я | 2 | 4 | – | H1 | А | А2б | А2б | А2б | А2б | А2б | А2б | А2б | А2б | А2б | А2б | А1б1а1а1б |
| А-М32 | * | * | * | * | * | * | * | * | А3 | А3 | А3 | А3 | А3 | А3 | А3 | А3 | А3 | A1b1b |
| А-М28 | 7 | я | 1А | 1 | – | H1 | А | А3а | А3а | А3а | А3а | А3а | А3а | А3а | А3а | А3а | А3а | А1б1б1 |
| А-М51 | 7 | я | 1А | 1 | – | H1 | А | А3б1 | А3б1 | А3б1 | А3б1 | А3б1 | А3б1 | А3б1 | А3б1 | А3б1 | А3б1 | A1b1b2a |
| А-М13 | 7 | я | 1А | 2 | ЕС1 | H1 | А | A3b2* | А3б2 | А3б2 | А3б2 | А3б2 | А3б2 | А3б2 | А3б2 | А3б2 | А3б2 | A1b1b2b |
| А-М171 | 7 | я | 1А | 2 | ЕС1 | H1 | А | A3b2a | A3b2a | A3b2a | A3b2a | A3b2a | A3b2a | A3b2a | A3b2a | A3b2a | A3b2a | удаленный |
| А-М118 | 7 | я | 1А | 2 | ЕС1 | H1 | А | A3b2b | A3b2b | A3b2b | A3b2b | A3b2b | A3b2b | A3b2b | A3b2b | A3b2b | A3b2b | А1b1b2b1 |
Следующие исследовательские группы в соответствии с их публикациями были представлены в создании Дерева YCC.
Филогенетические деревья
[ редактировать ]Вышеупомянутое филогенетическое дерево основано на ISOGG , [17] ЮСС , [53] и последующие опубликованные исследования.
Y-хромосомный Адам
А00 (AF6/L1284)
- А00а (L1149, FGC25576, FGC26292, FGC26293, FGC27741)
- A00b (A4987/YP3666, A4981, A4982/YP2683, A4984/YP2995, A4985/YP3292, A4986, A4988/YP3731)
А0-Т (Л1085)
- A0 (CTS2809/L991) ранее A1b
- A1 (P305) ранее A1a-T, A0 и A1b
- А1а (М31)
- A1b (P108) ранее A2-T
- А1б1 (Л419/ПФ712)
- А1б1а (Л602, В50, В82, В198, В224)
- A1b1a1 (M14) ранее A2
- А1б1а1а (М6)
- A1b1a1a1 (P28) ранее A1b1a1a1b и A2b
- А1б1а1а (М6)
- A1b1a1 (M14) ранее A2
- A1b1b (M32) ранее A3
- A1b1b1 (M28) ранее A3a
- А1b1b2 (L427)
- A1b1b2a (M51/Страница 42) ранее A3b1
- А1b1b2a1 (P291)
- A1b1b2b (M13/PF1374) ранее A3b2
- А1б1б2б1 (М118)
- A1b1b2a (M51/Страница 42) ранее A3b1
- А1б1а (Л602, В50, В82, В198, В224)
- БТ (М91)
- А1б1 (Л419/ПФ712)
См. также
[ редактировать ]- Гаплогруппа ДНК Y-хромосомы человека
- Гаплогруппы Y-ДНК в населении стран Африки к югу от Сахары
- Гаплогруппы Y-ДНК по этническим группам
- Субклады Y-ДНК А
Ссылки
[ редактировать ]- ^ эквивалентно оценке возраста человеческого Y-MRCA (см. Там); включая линию A00, Karmin et al. (2015) и Тромбетта и др. (2015) оценивают возраст в 254 000 и 291 000 лет назад соответственно.
- ^ Кармин; и др. (2015). «Недавнее ограничение разнообразия Y-хромосомы совпадает с глобальными изменениями в культуре» . Геномные исследования . 25 (4): 459–66. дои : 10.1101/гр.186684.114 . ПМЦ 4381518 . ПМИД 25770088 . «Мы датируем самого последнего общего предка Y-хромосомы (MRCA) в Африке 254 (95% ДИ 192–307) тысячелетий назад и обнаруживаем кластер основных неафриканских гаплогрупп-основателей в узком временном интервале в 47–52 тысячелетий, что соответствует с моделью быстрой первоначальной колонизации Евразии и Океании после узкого места за пределами Африки. В отличие от демографических реконструкций, основанных на мтДНК, мы предполагаем второе сильное узкое место в линиях Y-хромосомы, датируемое последними 10 тысячелетиями. «Узкое место» вызвано культурными изменениями, влияющими на различия в репродуктивном успехе среди мужчин».
- ^ Мендес, Л.; и др. (2016). «Расхождение Y-хромосом неандертальца и современного человека» . Американский журнал генетики человека . 98 (4): 728–34. дои : 10.1016/j.ajhg.2016.02.023 . ПМЦ 4833433 . ПМИД 27058445 .
- ^ Липсон, Марк; Рибо, Изабель; Маллик, Свапан; Роланд, Надин; Олальде, Иниго; Адамски, Николь; Брумандхошбахт, Насрин; Лоусон, Энн Мари; Лопес, Сайоа; Оппенгеймер, Йонас; Стюардсон, Кристина; Коллекционер, Рэймонд Нейбор; Бочеренс, Эрве; Брэдман, Нил; Каллетон, Брендан Дж.; Корнелиссен, Элс; Кревкер, Изабель; Марет, Питер; Фомине, Форка Лейпей Мэтью; Прачечная, Филипп; Миндзи, Кристофер Мбида; Орбан, Розин; Савчук, Элизабет; Семаль, Патрик; Томас, Марк Г.; Ван Нир, Вим; Вирама, Кришна Р.; Кеннетт, Дуглас Дж.; Паттерсон, Ник; Хеллентал, Гарретт; Лалуэса-Фокс, Чарльз; Макихерн, Скотт; Прендергаст, Мэри Э.; Райх, Дэвид (30 января 2020 г.). «Древние собиратели Западной Африки в контексте истории населения Африки » Природа 577 (7792): 665–670. Bibcode : 2020Nature.577..665L дои : 10.1038/ s41586-020-1929-1 ПМЦ 8386425 . ПМИД 31969706 .
- ^ Тромбетта, Бениамино; д'Атанасио, Евгения; Массайя, Андреа; Майрес, Натали М.; Скоццари, Розария; Кручиани, Фульвио; Новеллетто, Андреа (2015). «Региональные различия в накоплении SNP в мужской части Y-хромосомы человека, повторяющей аутосомные паттерны: значение для генетического датирования» . ПЛОС ОДИН . 10 (7): e0134646. Бибкод : 2015PLoSO..1034646T . дои : 10.1371/journal.pone.0134646 . ПМК 4520482 . ПМИД 26226630 .
- ^ «A00 YДерево» .
- ^ По данным Кручиани и др. В 2011 году большинство базальных линий было обнаружено в Западной , Северо-Западной и Центральной Африке , что позволяет предположить вероятность того, что Y-MRCA обитает в общем регионе Северо-Центральной Африки».В выборке из 2204 африканских Y-хромосом 8 хромосом принадлежали либо к гаплогруппе A1b, либо к A1a. Гаплогруппа A1a была идентифицирована у двух марокканских берберов, одного фульбе и одного туарега из Нигера. Гаплогруппа A1b была идентифицирована у трех пигмеев бакола из Южного Камеруна и одного алжирского бербера. Кручиани, Фульвио; Тромбетта, Бениамино; Массайя, Андреа; Дестро-Бисоль, Джованни; Селитто, Даниэле; Скоццари, Розария (2011). «Пересмотренный корень филогенетического дерева Y-хромосомы человека: происхождение отцовского разнообразия в Африке» . Американский журнал генетики человека . 88 (6): 814–8. дои : 10.1016/j.ajhg.2011.05.002 . ПМК 3113241 . ПМИД 21601174 . Скоццари и др. (2012) согласились с вероятным размещением гаплогруппы A1b в «северо-западном квадранте Африканского континента»:«гипотеза о происхождении гаплогруппы A1b в северо-западном квадранте африканского континента и, вместе с недавними находками древних Y-линий в центрально-западной Африке, предоставляют новые доказательства относительно географического происхождения человеческого разнообразия MSY» . Скоццари Р; Массайя А; Д'Атанасио Э; Майрес, Нью-Мексико; Перего ЮА; и др. (2012). Карамелли, Дэвид (ред.). «Молекулярное рассечение базальных клад филогенетического дерева Y-хромосомы человека» . ПЛОС ОДИН . 7 (11): e49170. Бибкод : 2012PLoSO...749170S . дои : 10.1371/journal.pone.0049170 . ПМЦ 3492319 . ПМИД 23145109 .
- ^ Камин М., Сааг Л., Винсенте М. и др. (апрель 2015 г.). «Недавнее ограничение разнообразия Y-хромосомы совпадает с глобальными изменениями в культуре» . Геномные исследования . 25 (4): 459–466. дои : 10.1101/гр.186684.114 . ПМЦ 4381518 . ПМИД 25770088 .
- ^ Батини С., Ферри Г., Дестро-Бисол Г. и др. (сентябрь 2011 г.). «Признаки доземледельческих процессов заселения в Африке к югу от Сахары, выявленные с помощью филогеографии ранних линий Y-хромосомы» . Мол. Биол. Эвол . 28 (9): 2603–13. дои : 10.1093/molbev/msr089 . hdl : 10400.13/4486 . ПМИД 21478374 .
- ^ Перейти обратно: а б с д Кручиани Ф, Тромбетта Б, Массайя А, Дестро-Бисол Г, Селитто Д, Скоццари Р (июнь 2011 г.). «Пересмотренный корень филогенетического дерева Y-хромосомы человека: происхождение отцовского разнообразия в Африке» . Являюсь. Дж. Хум. Жене 88 (6): 814–8. дои : 10.1016/j.ajhg.2011.05.002 . ПМК 3113241 . ПМИД 21601174 .
- ^ Перейти обратно: а б Скоццари Р., Массайя А., Д'Атанасио Э. и др. (2012). «Молекулярное рассечение базальных клад филогенетического дерева Y-хромосомы человека» . ПЛОС ОДИН . 7 (11): e49170. Бибкод : 2012PLoSO...749170S . дои : 10.1371/journal.pone.0049170 . ПМЦ 3492319 . ПМИД 23145109 .
- ^ Карина М. Шлебуш; и др. (28 сентября 2017 г.). «Древние геномы Южной Африки оценивают дивергенцию современного человека от 350 000 до 260 000 лет назад» . Наука . 358 (6363): 652–655. Бибкод : 2017Sci...358..652S . дои : 10.1126/science.aao6266 . ПМИД 28971970 .
- ^ Франкалаччи П, Морелли Л, Ангиус А, Берутти Р, Рейньер Ф, Ацени Р, Пилу Р, Бусонеро Ф, Маскио А, Зара И, Санна Д, Усели А, Урру МФ, Марчелли М, Кусано Р, Оппо М, Золедзевска М , Питцалис М, Дейдда Ф, Порку Э, Подди Ф, Канг Х.М., Лайонс Р., Тарриер Б, Грешам Дж.Б., Ли Б, Тофанелли С., Алонсо С., Дей М., Лай С., Мулас А., Уэлен М.Б., Уззау С., Джонс С, Шлессингер Д., Абекасис Г.Р., Санна С., Сидоре С., Кукка Ф. (2013). «Низкопроходное секвенирование ДНК 1200 сардинцев реконструирует европейскую филогению Y-хромосомы» . Наука . 341 (6145): 565–569. Бибкод : 2013Sci...341..565F . дои : 10.1126/science.1237947 . ПМК 5500864 . ПМИД 23908240 . Позник Г.Д., Хенн Б.М., Йи MC, Сливерска Е, Оускирхен ГМ, Лин А.А., Снайдер М., Кинтана-Мурси Л., Кидд Дж.М., Андерхилл П.А., Бустаманте CD (2013). «Секвенирование Y-хромосомы устраняет несоответствие во времени общему предку мужчин и женщин» . Наука . 341 (6145): 562–565. Бибкод : 2013Sci...341..562P . дои : 10.1126/science.1237619 . ПМК 4032117 . ПМИД 23908239 . Кручиани и др. (2011) оценили в 142 тыс. лет назад.
- ^ Перейти обратно: а б «Недавнее ограничение разнообразия Y-хромосомы совпадает с глобальными изменениями в культуре» . Геномные исследования . 25 (4): 459–66. 2015. дои : 10.1101/гр.186684.114 . ПМЦ 4381518 . ПМИД 25770088 .
- ^ Перейти обратно: а б «Региональные различия в накоплении SNP в мужской части Y-хромосомы человека, повторяющей аутосомные паттерны: значение для генетического датирования» . ПЛОС ОДИН . 10 (7): e0134646. 2015. Бибкод : 2015PLoSO..1034646T . дои : 10.1371/journal.pone.0134646 . ПМК 4520482 . ПМИД 26226630 .
- ^ Перейти обратно: а б Абу-Амеро К.К., Хеллани А., Гонсалес А.М., Ларруга Х.М., Кабрера В.М., Андерхилл, Пенсильвания (2009). «Разнообразие Y-хромосомы Саудовской Аравии и его связь с близлежащими регионами» . БМК Генет . 10:59 . дои : 10.1186/1471-2156-10-59 . ПМК 2759955 . ПМИД 19772609 .
- ^ Перейти обратно: а б Международное общество генетической генеалогии. «Дерево гаплогрупп Y-ДНК» .
- ^ Перейти обратно: а б Берниелл-Ли Дж., Калафель Ф., Бош Э. и др. (июль 2009 г.). «Генетические и демографические последствия экспансии банту: данные об отцовских линиях человека» . Мол. Биол. Эвол . 26 (7): 1581–9. дои : 10.1093/molbev/msp069 . ПМИД 19369595 .
- ^ Перейти обратно: а б Роза А., Орнелас С., Джоблинг М.А., Брем А., Виллемс Р. (2007). «Y-хромосомное разнообразие населения Гвинеи-Бисау: многоэтническая перспектива» . БМК Эвол. Биол . 7 (1): 124. Бибкод : 2007BMCEE...7..124R . дои : 10.1186/1471-2148-7-124 . ЧВК 1976131 . ПМИД 17662131 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С Вуд Э.Т., Стовер Д.А., Эрет С. и др. (июль 2005 г.). «Контрастные закономерности вариаций Y-хромосомы и мтДНК в Африке: свидетельства демографических процессов с учетом пола» . Евро. Дж. Хум. Жене . 13 (7): 867–76. дои : 10.1038/sj.ejhg.5201408 . ПМИД 15856073 .
- ^ Андерхилл П.А., Пассарино Г., Лин А.А. и др. (январь 2001 г.). «Филогеография бинарных гаплотипов Y-хромосомы и происхождение современных человеческих популяций» . Энн. Хм. Жене . 65 (Часть 1): 43–62. дои : 10.1046/j.1469-1809.2001.6510043.x . ПМИД 11415522 . S2CID 9441236 .
- ^ Перейти обратно: а б с д и ж г час я 28/53 (Динка, Нуэр и Шиллук), Хасан Х.И., Андерхилл П.А., Кавалли-Сфорца Л.Л., Ибрагим М.Е. (ноябрь 2008 г.). «Вариации Y-хромосомы у суданцев: ограниченный поток генов, соответствие языку, географии и истории». Являюсь. Дж. Физ. Антрополь . 137 (3): 316–23. дои : 10.1002/ajpa.20876 . ПМИД 18618658 .
- ^ Перейти обратно: а б с д и ж г час я дж Андерхилл П.А., Шен П., Лин А.А. и др. (ноябрь 2000 г.). «Вариации последовательности Y-хромосомы и история человеческих популяций». Нат. Жене . 26 (3): 358–61. дои : 10.1038/81685 . ПМИД 11062480 . S2CID 12893406 .
- ^ Перейти обратно: а б с д и ж Кручиани Ф., Сантоламазца П., Шен П. и др. (май 2002 г.). «Обратная миграция из Азии в страны Африки к югу от Сахары подтверждается анализом гаплотипов Y-хромосомы человека с высоким разрешением» . Являюсь. Дж. Хум. Жене . 70 (5): 1197–214. дои : 10.1086/340257 . ПМЦ 447595 . ПМИД 11910562 .
- ^ Перейти обратно: а б Шен П., Лави Т., Кивисилд Т. и др. (сентябрь 2004 г.). «Реконструкция отцовских и матрилиний самаритян и других израильских популяций на основе вариаций последовательностей Y-хромосомы и митохондриальной ДНК». Хм. Мутат . 24 (3): 248–60. дои : 10.1002/humu.20077 . ПМИД 15300852 . S2CID 1571356 .
- ^ Перейти обратно: а б Кручиани Ф., Тромбетта Б., Селлито Д. и др. (июль 2010 г.). «Гаплогруппа R-V88 Y-хромосомы человека: отцовская генетическая запись транссахарских связей начала среднего голоцена и распространения чадских языков» . Евро. Дж. Хум. Жене . 18 (7): 800–7. дои : 10.1038/ejhg.2009.231 . ПМЦ 2987365 . ПМИД 20051990 .
- ^ Перейти обратно: а б с д Луис Дж.Р., Роуолд Д.Д., Регейро М. и др. (март 2004 г.). «Левант против Африканского Рога: свидетельства двунаправленных коридоров миграции людей» . Являюсь. Дж. Хум. Жене . 74 (3): 532–44. дои : 10.1086/382286 . ПМЦ 1182266 . ПМИД 14973781 .
- ^ Перейти обратно: а б с д Семино О, Сантакьяра-Бенерекетти А.С., Фаласки Ф, Кавалли-Сфорца ЛЛ, Андерхилл, Пенсильвания (январь 2002 г.). «Эфиопы и койсанцы разделяют самые глубокие клады филогении Y-хромосомы человека » Являюсь. Дж. Хум. Жене 70 (1): 265–8. дои : 10.1086/338306 . ПМЦ 384897 . ПМИД 11719903 .
- ^ Перейти обратно: а б Кинг Т.Э., Паркин Э.Дж., Суинфилд Г. и др. (март 2007 г.). «Африканцы в Йоркшире? Самая глубоко укоренившаяся клада филогении Y в английской генеалогии» . Евро. Дж. Хум. Жене . 15 (3): 288–93. дои : 10.1038/sj.ejhg.5201771 . ПМК 2590664 . ПМИД 17245408 .
Новостная статья: «Йоркширский клан связан с Африкой» . Новости Би-би-си . 24 января 2007 г. Проверено 27 января 2007 г. - ^ 16/26, Хасан и др. 2008 год
- ^ Батини, Кьяра; и др. (сентябрь 2011 г.). «Дополнительные данные: признаки доземледельческих процессов заселения в Африке к югу от Сахары, выявленные филогеографией ранних линий Y-хромосомы» . Молекулярная биология и эволюция . 28 (9): 2603–2613. дои : 10.1093/molbev/msr089 . hdl : 10400.13/4486 . ISSN 0737-4038 . OCLC 748733133 . ПМИД 21478374 . S2CID 11190055 .
- ^ Перейти обратно: а б Ди Джакомо Ф., Лука Ф., Ананью Н. и др. (сентябрь 2003 г.). «Клинические закономерности разнообразия Y-хромосомы человека в континентальной Италии и Греции определяются эффектами дрейфа и основателя». Мол. Филогенет. Эвол . 28 (3): 387–95. дои : 10.1016/S1055-7903(03)00016-2 . ПМИД 12927125 .
- ^ Гонсалвес Р., Фрейтас А., Бранко М. и др. (июль 2005 г.). «Линии Y-хромосомы из Португалии, Мадейры и Азорских островов фиксируют элементы сефардского и берберского происхождения». Энн. Хм. Жене . 69 (Часть 4): 443–54. дои : 10.1111/j.1529-8817.2005.00161.x . hdl : 10400.13/3018 . ПМИД 15996172 . S2CID 3229760 .
- ^ Капелли С., Редхед Н., Романо В. и др. (март 2006 г.). «Структура населения в бассейне Средиземного моря: перспектива Y-хромосомы». Энн. Хм. Жене . 70 (Часть 2): 207–25. дои : 10.1111/j.1529-8817.2005.00224.x . hdl : 2108/37090 . ПМИД 16626331 . S2CID 25536759 .
- ^ Мендес, Фернандо Л.; Кран, Томас; Шрак, Бонни; Кран, Астрид-Мария; Вирама, Кришна Р.; Вернер, Август Э.; Фомине, Форка Лейпей Мэтью; Брэдман, Нил; Томас, Марк Г.; Карафет Татьяна М.; Хаммер, Майкл Ф. (март 2013 г.). «Афроамериканская отцовская линия добавляет чрезвычайно древний корень к филогенетическому дереву Y-хромосомы человека» . Американский журнал генетики человека . 92 (3): 454–459. дои : 10.1016/j.ajhg.2013.02.002 . ПМК 3591855 . ПМИД 23453668 .
- ^ Мендес, Фернандо Л.; Кран, Томас; Шрак, Бонни; Кран, Астрид-Мария; Вирама, Кришна Р.; Вернер, Август Э.; Фомине, Форка Лейпей Мэтью; Брэдман, Нил; Томас, Марк Г.; Карафет Татьяна М.; Хаммер, Майкл Ф. (март 2013 г.). «Афроамериканская отцовская линия добавляет чрезвычайно древний корень к филогенетическому дереву Y-хромосомы человека» . Американский журнал генетики человека . 92 (3): 454–459. дои : 10.1016/j.ajhg.2013.02.002 . ПМК 3591855 . ПМИД 23453668 .
- ^ Сначала через Mendez et al. (2013), это было объявлено «чрезвычайно древним» (95% доверительный интервал 237–581 тысяч лет назад для возраста Y-MRCA, включая происхождение этой постулируемой гаплогруппы).
- ^ Альберт Перри, раб , родившийся в Соединенных Штатах между ок. 1819–1827, жил в округе Йорк , Южная Каролина . См. FamilyTreeDNA, диаграмму гаплогруппы A.
- ^ Мендес и др. (2013), с. 455. Цитата: «После поиска в большой панафриканской базе данных, состоящей из 5648 образцов из десяти стран [...] мы идентифицировали 11 Y-хромосом, которые были инвариантными и идентичными хромосоме A00 в пяти из шести Y-STR (2 из 11 хромосом несли DYS19-16, тогда как остальные несли DYS19-15. Все эти 11 хромосом были обнаружены в выборке из 174 (~6,3%) особей Mbo из западного Камеруна (рис. 2). для дальнейшего тестирования, и генотипы оказались идентичными в 37 из 39 SNP, которые, как известно, произошли от хромосомы A00 (т.е. два из этих генотипированных SNP были предковыми в образцах Mbo)».
- ^ У кого из народов Камеруна есть члены гаплогруппы A00? // Обновление финансируемого исследования на сайте Experiment.com (Schrack/Fomine Forka) доступно в Интернете. [ самостоятельно опубликованный источник? ] Цитаты: Теперь мы можем ясно видеть, что с 40% А00 Бангва представляет собой эпицентр А00 в этом регионе и, вполне возможно, во всем мире. Как я рассказал в последней лабораторной заметке, мы обнаружили, что на данный момент существуют две основные подгруппы A00, определяемые различными мутациями Y-SNP, которые, естественно, делятся по этническому признаку: A00a среди бангва и A00b среди мбо. Мы также нашли один образец бангва, который не принадлежал ни к одной из подгрупп».
- ^ Липсон Марк и др. ДНК древнего человека из Шум-Лаки (Камерун) в контексте истории населения Африки // SAA 2019
- ^ Гонсалвес Р., Роза А., Фрейтас А. и др. (ноябрь 2003 г.). «Линии Y-хромосомы на островах Кабо-Верде свидетельствуют о разнообразном географическом происхождении первых поселенцев-мужчин». Хм. Жене . 113 (6): 467–72. дои : 10.1007/s00439-003-1007-4 . hdl : 10400.13/3047 . ПМИД 12942365 . S2CID 63381583 .
- ^ 23andme необработанные данные
- ^ Хишам Ю. Хасан и др. (2008). «Южный Судан» включает 26 динка, 15 шиллуков и 12 нуэров. «Западный Судан» включает 26 боргу, 32 масалита и 32 фура. «Северный Судан» включает 39 нубийцев, 42 беджа, 33 копта, 50 гаалиен, 28 месерий и 24 аракиена.
- ^ Чинниоглу С, Кинг Р, Кивисилд Т; и др. (январь 2004 г.). «Раскопки слоев гаплотипов Y-хромосомы в Анатолии». Хм. Жене . 114 (2): 127–48. дои : 10.1007/s00439-003-1031-4 . ПМИД 14586639 . S2CID 10763736 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Небель А., Филон Д., Бринкманн Б., Маджумдер П.П., Фаерман М., Оппенгейм А. (ноябрь 2001 г.). «Пул Y-хромосомы евреев как часть генетического ландшафта Ближнего Востока» . Являюсь. Дж. Хум. Жене . 69 (5): 1095–112. дои : 10.1086/324070 . ПМЦ 1274378 . ПМИД 11573163 .
- ^ Семино О., Пассарино Г., Офнер П.Дж. и др. (ноябрь 2000 г.). «Генетическое наследие палеолитического Homo sapiens sapiens у современных европейцев: взгляд на Y-хромосому». Наука . 290 (5494): 1155–9. Бибкод : 2000Sci...290.1155S . дои : 10.1126/science.290.5494.1155 . ПМИД 11073453 .
- ^ Флорес С., Мака-Мейер Н., Ларруга Дж.М., Кабрера В.М., Карадше Н., Гонсалес А.М. (2005). «Изоляты в коридоре миграций: анализ вариаций Y-хромосомы с высоким разрешением в Иордании» . Дж. Хум. Жене . 50 (9): 435–41. дои : 10.1007/s10038-005-0274-4 . ПМИД 16142507 .
- ^ Юсиф, Хишам; Эльтайеб, Мунтасер (июль 2009 г.). Генетические закономерности вариаций Y-хромосомы и митохондриальной ДНК, имеющие значение для заселения Судана (Диссертация). Архивировано из оригинала 25 августа 2022 г. Проверено 25 августа 2022 г.
- ^ Скоррано, Габриэле; Да здравствует, Серена; Пинотти, Томаз; Фаббри, Пьер Франческо; Рикардс, Ольга; Маккарди, Фабио (26 мая 2022 г.). «Биоархеологический и палеогеномный портрет двух помпейцев, погибших во время извержения Везувия в 79 году нашей эры» . Научные отчеты . 12 (1): 6468. Бибкод : 2022NatSR..12.6468S . дои : 10.1038/s41598-022-10899-1 . ПМЦ 9135728 . ПМИД 35618734 .
- ^ Карафет Т.М., Мендес Ф.Л., Мейлерман М.Б., Андерхилл П.А., Зегура С.Л., Хаммер М.Ф. (2008). «Новые бинарные полиморфизмы изменяют форму и увеличивают разрешение дерева гаплогруппы Y-хромосомы человека» . Геномные исследования . 18 (5): 830–8. дои : 10.1101/гр.7172008 . ПМК 2336805 . ПМИД 18385274 .
- ^ ISOGG, 2016, Дерево гаплогрупп Y-ДНК, 2016 . (Доступ: 29 августа 2017 г.)
- ^ Кран, Томас. «Дерево YCC» . ФТДНА. Архивировано из оригинала 26 июля 2011 г.
Библиография
[ редактировать ]- Мендес Ф.Л., Кран Т., Шрак Б. и др. (март 2013 г.). «Афроамериканская линия по отцовской линии добавляет чрезвычайно древний корень к филогенетическому дереву Y-хромосомы человека» . Являюсь. Дж. Хум. Жене . 92 (3): 454–9. дои : 10.1016/j.ajhg.2013.02.002 . ПМК 3591855 . ПМИД 23453668 . в формате PDF. Архивировано 24 сентября 2019 г. на Wayback Machine.
- «Y-гаплогруппа А Филогенетическое древо» . Март 2013 г. Архивировано из оригинала 18 августа 2018 г. Проверено 30 марта 2013 г. (диаграмма, показывающая новые ветви, добавленные в филодерево A в марте 2013 г.)
Источники таблиц преобразования
[ редактировать ]- Капелли, Кристиан; Уилсон, Джеймс Ф.; Ричардс, Мартин; Штумпф, Майкл П.Х.; и др. (февраль 2001 г.). «Отцовское наследие преимущественно коренных народов австронезийскоязычных народов островной Юго-Восточной Азии и Океании» . Американский журнал генетики человека . 68 (2): 432–443. дои : 10.1086/318205 . ПМЦ 1235276 . ПМИД 11170891 .
- Хаммер, Майкл Ф.; Карафет Татьяна М.; Редд, Алан Дж.; Джарджанази, Хамди; и др. (1 июля 2001 г.). «Иерархические закономерности глобального разнообразия Y-хромосом человека» . Молекулярная биология и эволюция . 18 (7): 1189–1203. doi : 10.1093/oxfordjournals.molbev.a003906 . ПМИД 11420360 .
- Джоблинг, Марк А.; Тайлер-Смит, Крис (2000), «Новое использование новых гаплотипов», Trends in Genetics , 16 (8): 356–62, doi : 10.1016/S0168-9525(00)02057-6 , PMID 10904265
- Каладжиева, преп.; Калафель, Франческ; Джоблинг, Марк А; Ангеличева, Дора; и др. (февраль 2001 г.). «Закономерности меж- и внутригруппового генетического разнообразия у влакс-рома, выявленные с помощью Y-хромосомы и линий митохондриальной ДНК» . Европейский журнал генетики человека . 9 (2): 97–104. дои : 10.1038/sj.ejhg.5200597 . ПМИД 11313742 .
- Карафет, Татьяна; Сюй, Липин; Ду, Руофу; Ван, Уильям; и др. (сентябрь 2001 г.). «История отцовского населения Восточной Азии: источники, закономерности и микроэволюционные процессы» . Американский журнал генетики человека . 69 (3): 615–628. дои : 10.1086/323299 . ПМЦ 1235490 . ПМИД 11481588 .
- Семино, О.; Пассарино, Дж; Оефнер, П.Дж.; Лин, А.А.; и др. (2000), «Генетическое наследие палеолитического Homo sapiens sapiens у современных европейцев: хромосомная перспектива AY», Science , 290 (5494): 1155–9, Бибкод : 2000Sci...290.1155S , doi : 10.1126/science.290.5494. 1155 , ПМИД 11073453
- Су, Бинг; Сяо, Цзюньхуа; Андерхилл, Питер; Дека, Ранджан; и др. (декабрь 1999 г.). «Y-хромосома свидетельствует о миграции современных людей на север в Восточную Азию во время последнего ледникового периода» . Американский журнал генетики человека . 65 (6): 1718–1724. дои : 10.1086/302680 . ПМЦ 1288383 . ПМИД 10577926 .
- Андерхилл, Питер А.; Шен, Пейдун; Лин, Алиса А.; Джин, Ли; и др. (ноябрь 2000 г.). «Вариации последовательности Y-хромосомы и история человеческих популяций». Природная генетика . 26 (3): 358–361. дои : 10.1038/81685 . ПМИД 11062480 . S2CID 12893406 .
Внешние ссылки
[ редактировать ]- ДНК генеалогического древа — проект Y-гаплогруппы А
- Проект африканской гаплогруппы в FTDNA
- Распространение гаплогруппы А , от National Geographic