Гаплогруппа N-M231
Эта статья , возможно, содержит оригинальные исследования . ( Ноябрь 2021 г. ) |
| Гаплогруппа N | |
|---|---|
 | |
| Возможное время происхождения | 36 800 [95% ДИ 34 300–39 300] лет назад (YFull [ 1 ] ) 44 700 или 38 300 лет назад в зависимости от скорости мутации [ 2 ] 41,900 [95% CI 40,175-43,591] ybp[3] |
| Coalescence age | 21,700 [95% CI 19,500–23,900] ybp (YFull[1]) 25,313 [95% CI 21,722–28,956] ybp[3] |
| Possible place of origin | Northern East Asia[4][5] |
| Ancestor | NO |
| Defining mutations | M231 |
| Highest frequencies | Nganassan 58%[3]–94.1%,[6] Yakuts 81.8%[7]–94.6%,[8] Khakass (Shirinsky District) 90.2%,[9] Siberian Tatars 33.5% (Zabolotnie 89.5%, Iskero-Tobol 23.5%, Tatar-Bukharans 16.5%, Yalutorov 16.3%, Ishtyak-Tokuz 14.5%[10]), Ugrians 77.8%[3] (Khanty 64.3%[11]–89.3%,[12] Mansi 76%[12]), Udmurts 77.8%,[3] Khakas 41%[3]–65%,[11] Komi 33.3%[11]–79.5%,[6] Nenets 75%–92.9%[3] (Tundra Nenets 97.9%,[6] Forest Nenets 98.8%[6]), Vepsians 55%,[3] Finns 42.6% (West)[13]–70.9% (East)[13] or approx. 54%[3]–58.8%,[14] Tuvans 27.2–54.5%,[citation needed] Nanai 46.2%[3][15][16] (20% Hezhe in the PRC,[15] 44.6% Nanai in Russia,[3] 83.8% members of the Samar clan in the Gorin area of the Khabarovsk Territory[16]), Karelians 37.1%[5]–53.8%,[13] Arkhangelsk Russians 42.6% (Arkhangelsk 44.3%,[3] Pinega 40.8%[3]), Lithuanians 40.5%[3]–44.5%,[13] Latvians approx. 42% (41.6%,[13] 42.1%,[17] 43.0%[3]), Mari 41.2%,[3] Saami 40%, Chuvash 33.7%[14]–36%,[3] Buryats 34.5% (20.2%,[18] 25.0%,[19] 30.9%,[20] 48.0%[21]>), Koryaks 33.3%,[6] Estonians 30.6%[3]-33.9%,[13] Volga Tatars 27.8%,[3] Teleuts 25.0%,[6] Northern Altaians 21.8% (18.0%[11][22]–24.6%[23]), Pskov Russians 22.7%,[3] Russians 20%[24] Bashkirs 17.3%,[3] Sibe 17.1%[15]–18.0%,[25] Mordvins 12.5% (10%[3]–13.3%[3]), Mongols 11%,[26][20][15][19][27][28] Kalmyks 10.4% (Torguud 3.4%, Derbet 5.1%, Buzava 5.3%, Khoshut 38.2%),[29][28] Manchus 10% (5.8%,[20] 8.1%,[30] 9.1%,[25] 11.6%,[25] 12.5%,[25] 14.3%[15]), Belarusians 9.7%,[3] Central-Southern Russians 9.1% (Tver 13.2%,[31] Kursk 12.5%[31]–13.3%,[3] Belgorod 11.9%,[3] Kostroma 11.8%,[3] Smolensk 7.0%,[3] Voronezh 6.3%,[3] Oryol 5.5%[3]), Ukrainians 9.0%,[3] Southern Altaians 7.1% (4.2%[23]-9.7%[11]), Mulam 7.1%,[32] Sweden 6.8%[14] (0% Västra Götaland, Halland, Malmö, and Jönköping[33] - 19.5% Västerbotten[34]), Han Chinese 6.77% (0% to 21.4%),[25] Koreans 6.58% (4.41% to 12%) 12% Koreans,[35] 6.58% Koreans from KPGP(Korean Genome Project),[36] 6.9% Jeju[19] 6.4% Gochang [37] 6.3% Gangwon [19] 5.7% North Korean [38] 4.8% Gyeongsang,[19] 4.4% Jeolla,[19] 4.2% Chungcheong,[19] 4.0% Seoul,[39] 3.0% Daejeon,[39] 1.8% Seoul-Gyeonggi,[19] Ulchi 5.8%,[40] Tibetans 5.65%,[41] Kazakhs 5.33% [42] (Suan 0%, Qangly 0%, Oshaqty 0%, Jetyru 1.2%, Dulat 1.6%, Argyn 2.0%, Alimuly 2.5%, Ysty 3.5%, Baiuly 3.9%, Alban 4.3%, Qongyrat 7.4%, Qypshaq 10.3%, Jalair 10.9%, Qozha 16.7%, Syrgeli 65.6%), Northern Thai 5.2%,[43] Uyghurs 4.89% (2.8%,[44] 4.8%,[25] 4.99%,[45] 6.0%,[20] 8.6%[15]), Kyrgyz 3.9% (2.8% Kyzylsu,[46] 3.3% Kyzylsu,[47] 4.5% Kyrgyzstan,[27] 10% Urumqi[46]), Vietnamese 3.4%, Japanese 1.9% (0%,[2] 0.8%,[48] 0.9%,[49] 1.7%,[50] 2.5%,[19] 4.3%,[51] 4.8%,[20] 6.4%[15]) |
Гаплогруппа N (M231) представляет собой гаплогруппу ДНК Y-хромосомы, определяемую наличием маркера однонуклеотидного полиморфизма (SNP) M231. [ Филогенетика 1 ]
Чаще всего встречается у мужчин, происходящих из Северной Евразии . Это также наблюдалось с более низкой частотой у популяций, обитающих в других регионах, включая части Балкан , Центральной Азии , Восточной Азии и Юго-Восточной Азии .
Однако базальная парагруппа N* была обнаружена только у коренных популяций Китая и Камбоджи . [ 4 ] Субклады N-M231 были обнаружены в небольших количествах в Юго-Восточной Азии , на островах Тихого океана , Юго-Западной Азии и на Балканах . Эти факторы позволяют предположить, что он возник в Восточной или Юго-Восточной Азии.
Origins
[edit]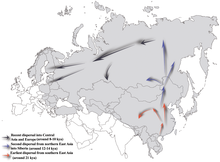
Haplogroup NO-M214 – its most recent common ancestor with its sibling, haplogroup O-M175 – is estimated to have existed about 36,800–44,700 years ago.[1][53][2]
It is generally considered that N-M231 arose in East Asia approximately 19,400 (±4,800) years ago and populated northern Eurasia after the Last Glacial Maximum. Males carrying the marker apparently moved northwards as the climate warmed in the Holocene, migrating in a counter-clockwise path, to eventually become concentrated in areas as far away as Fennoscandia and the Baltic.[5] The apparent dearth of haplogroup N-M231 amongst Native American peoples indicates that it spread after Beringia was submerged,[54] about 11,000 years ago.
Distribution
[edit]
Haplogroup N has a wide geographic distribution throughout northern Eurasia, and it also has been observed occasionally in other areas, including Central Asia and the Balkans.
It has been found with greatest frequency among indigenous peoples of Russia, including Finnic peoples, Mari, Udmurt, Komi, Khanty, Mansi, Nenets, Nganasans, Turkic peoples (Yakuts, Dolgans, Khakasses, Tuvans, Tatars, Chuvashes, etc.), Buryats, Tungusic peoples (Evenks, Evens, Negidals, Nanais, etc.), Yukaghirs, Luoravetlans (Chukchis, Koryaks), and Siberian Eskimos, but certain subclades are very common in Finland, Estonia, Latvia, and Lithuania, and other subclades are found at low frequency in China (Yi, Naxi, Lhoba, Han Chinese, etc.).[55] Especially in ethnic Finnic peoples and Baltic-speaking peoples of northern Europe, the Ob-Ugric-speaking and Northern Samoyed peoples of western Siberia, and Turkic-speaking peoples of Russia (especially Yakuts,[citation needed],but also Altaians, Shors, Khakas, Chuvashes, Tatars, and Bashkirs). Nearly all members of haplogroup N among these populations of northern Eurasia belong to subclades of either haplogroup N-CTS6128/M2048 or haplogroup N-P43.
Y-chromosomes belonging to N1b-F2930/M1881/V3743, or N1*-CTS11499/L735/M2291(xN1a-F1206/M2013/S11466), have been found in China (where they account for approximately 3.62% of all Y-DNA[56]) and sporadically throughout other parts of Eurasia. The N-CTS6128/M2048 and N-P43 subclades of N1a-F1206/M2013/S11466 are found in high numbers in Northern Eurasia; however, members of N1a-F1206(xCTS6128, P43) are currently found mainly in northern China and Korea.
N2-Y6503, the other primary subclade of haplogroup N, is extremely rare and is mainly represented among extant humans by a recently formed subclade that is virtually restricted to the countries making up the former Yugoslavia (Bosnia-Herzegovina, Croatia, Serbia, and Montenegro), Hungary and Austria. Other members of N2-Y6503 include a Hungarian with recent ancestry from Suceava in Bukovina, a Slovakian, a few British individuals, and an Altaian.[1]
N* (M231)
[edit]Y-chromosomes that display the M231 mutation that defines Haplogroup N-M231, but do not display the CTS11499, L735, M2291 mutations that define Haplogroup N1 are said to belong to paragroup N-M231*.[4]
N-M231* has been found at low levels in China.[4] Out of a sample of 165 Han males from China, two individuals (1.2%) were found to belong to N*.[57] One originated from Guangzhou and one from Xi'an.[citation needed]
Among the ancient samples from the Baikal Early Neolithic Kitoi culture, one of the Shamanka II samples (DA250), dated to c. 6500 BP, was analyzed as NO1-M214 in the original study.[58] However, this same specimen (DA250 or Shamanka 250) has subsequently been found to belong to N-FT210118, the same clade as the other haplogroup N specimens from the same site (besides DA247, who belongs to N-Y147969). N-FT210118 is derived from N-L666/N-F2199 but basal to N-CTS6380, this latter being the most recent common ancestor of present-day N-P43 (found mainly among Maris, Udmurts, Komis, Chuvashes, Tatars, Nenets, Nganasans, Khanty, Mansi, Khakas, Tuvans, etc.) and N-F1101 (found mainly among East Asians). Furthermore, N-FT210118 has not been found in any living individual who has had his Y-DNA tested to date, and the estimated TMRCA of N-CTS6380 exceeds the estimated date of deposition of any of the specimens from the Shamanka site associated with the Kitoi culture, so it appears that the representatives of the Kitoi culture at Shamanka (or at least their Y-DNA) have gone extinct rather than being direct ancestors of any living people.[59][60]
N1 (CTS11499, Z4762, CTS3750)
[edit]In 2014, there was a major change in the definition of subclade N1, when LLY22g was retired as the main defining SNP for N1 because of reports of LLY22g's unreliability. According to ISOGG, LLY22g is problematic because it is a "palindromic marker and can easily be misinterpreted."[4] Since then, the name N1 has been applied to a clade marked by a great number of SNPs, including CTS11499, Z4762, and CTS3750. N1 is the most recent common ancestor of all extant members of Haplogroup N-M231 except members of the rare N2-Y6503 (N2-B482) subclade. The TMRCA of N1 is estimated to be 18,000 years before present (16,300–19,700 BP; 95% CI).[1] Since the revision of 2014, the position of many examples of "N1-LLY22g" within haplogroup N have become unclear. Therefore, it is better to check yfull and ISOGG 2019 in order to understand the updated structure of N-M231.
However, in older studies, N-LLY22g has been reported to reach a frequency of up to 30% (13/43) among the Yi people of Butuo County, Sichuan in Southwest China.[20][61][62] It is also found in 34.6% of Lhoba people.[62] N1-LLY22g* has been found in samples of Han Chinese, but with widely varying frequency:
- 6.8% (3/44) Han from Xi'an[20][61]
- 6.7% (2/30) Han from Lanzhou[15]
- 3.6% (3/84) Taiwanese Han[20]
- 2.9% (1/34) Han from Chengdu[15]
- 2.9% (1/35) Han from Harbin[15]
- 2.9% (1/35) Han from Meixian District[15]
- 0% (0/32) Han from Yining City[15]
Other populations in which representatives of N1*-LLY22g have been found include:
- Hani people (4/34 = 11.8%)[15]
- Sibe people (4/41 = 9.8%)[15]
- Tujia people (2/49 = 4.1%)[20]
- Manchu people (2/52 = 3.8%[20] to 2/35 = 5.7%[15]
- Bit people (1/28 = 3.6%)[63]
- Uyghurs (2/70 = 2.9%[15] to 2/67 = 3.0%)[20]
- Tibetan people (3/105 = 2.9%[20] to 3/35 = 8.6%)[15]
- Koreans (0/106 = 0.0% – 2/25 = 8%[5][15][64]
- Vietnamese people (2/70 = 2.9%)[20]
- Japanese people (0/70 Tokushima – 2/26 = 7.7% Aomori)[20]
- Manchurian Evenks (0/26 = 0.0%[15] to 1/41 = 2.4%)[20]
- Altai people (0/50 Northern to 5/96 = 5.2% Southern, or 0/43 Beshpeltir to 5/46 = 10.9% Kulada),[20][22]
- Shors (2/23 = 8.7%)[5]
- Khakas people (5/181 = 2.8%)[5]
- Tuvans (5/311 = 1.6%)[5]
- Southern Borneo (1/40 = 2.5%)[5]
- Forest Nenets (1/89 = 1.1%)[5]
- Yakuts (0/215 – 1/121 = 0.8%)[5]
- Turkish people (1/523 = 0.2%)[5] In Turkey, the total of subclades of haplogroup N-M231 amounts to 4% of the male population.
- One individual who belongs either to N* or N1* has been found in a sample of 77 males from Kathmandu, Nepal (1/77 = 1.3% N-M231(xM128,P43,Tat))[65]
N1(xN1a,N1c) was found in ancient bones of Liao civilization:[66]
- Niuheliang (Hongshan Culture, 6500–5000 BP) 66.7%(=4/6)
- Halahaigou (Xiaoheyan Culture, 5000–4200 BP) 100.0%(=12/12)
- Dadianzi (Lower Xiajiadian culture, 4200–3600 BP) 60.0%(=3/5).
N-CTS4309: two people identified with this subgroup in Iraq. Very rare.
N1a (F1206/M2013/S11466)
[edit]The N1a2-F1008/L666 clade and N1a1-M46/Page70/Tat are estimated to share a most recent common ancestor in N1a-F1206/M2013/S11466 approximately 15,900 [95% CI 13,900 <-> 17,900] years before present[1] or 17,621 [95% CI 14,952 <-> 20,282] years before present.[3]
N1a1 (M46/Page70/Tat, L395/M2080)
[edit]All M46 in Yfull database are M178, being a quarter younger than separation from F1139.[67]
The mutations that define the subclade N-M46[Phylogenetics 2] are M46/Tat and P105. This is the most frequent subclade of N. It probably arose in a Northeast Asian population, because the oldest ancient samples comply with this genetic profile.[68] N has experienced serial bottlenecks in Siberia and secondary expansions in eastern Europe.[5] Haplogroup N-M46 is approximately 14,000 years old.
In Siberia, haplogroup N-M46 reaches a maximum frequency of approximately 90% among the Yakuts, a Turkic people who live mainly in the Sakha (Yakutia) Republic. However, N-M46 is present with much lower frequency among many of the Yakuts' neighbors, such as Evenks and Evens.[8] It also has been detected in 5.9% (3/51) of a sample of Hmong Daw from Laos,[63] 2.4% (2/85) of a sample from Seoul, South Korea,[26] and in 1.4% (1/70) of a sample from Tokushima, Japan.[20]
The haplogroup N-M46 has a low diversity among Yakuts suggestive of a population bottleneck or founder effect.[69] This was confirmed by a study of ancient DNA which traced the origins of the male Yakut lineages to a small group of horse-riders from the Cis-Baikal area.[70]
N-Tat has been observed with greatly varying frequency in samples from Sweden. Karlsson et al. (2006) found N-Tat in 44.7% (17/38) of a sample of Saami nomads from Jokkmokk, 19.5% (8/41) of a sample from Västerbotten, 14.5% (8/55) of a sample from Uppsala, 10.0% (4/40) of a sample from Gotland, 9.5% (4/42) of a sample from Värmland, 7.3% (3/41) of a sample from Östergötland/Jönköping, 2.4% (1/41) of a sample from Blekinge/Kristianstad, and 2.2% (1/45) of a sample from Skaraborg.[34]
Lappalainen et al. (2008) found N-Tat in 14.4% (23/160) of a sample from Sweden.[13]
Lappalainen et al. (2009) found N-Tat in 15.4% (4/26) of a sample from Södermanland, 12.5% (3/24) of a sample from Västmanland, 12.1% (4/33) of a sample from Uppsala, 7.8% (4/51) of a sample from Gothenburg, 7.0% (3/43) of a sample from Norrbotten, 6.8% (5/73) of a sample from Skåne, 6.6% (15/228) of a sample from Stockholm, 6.3% (3/48) of a sample from Sydnorrland, 6.3% (2/32) of a sample from Västerbotten, 6.3% (2/32) of a sample from Örebro, 5.9% (3/51) of a sample from Värmland/Dalarna, 5.4% (2/37) of a sample from Östra Götaland, and 5.1% (2/39) of a sample from southeastern Sweden (Kalmar, Gotland, Kronoberg, and Blekinge). They did not find any instance of N-Tat in their samples from Jönköping (0/28), Malmö (0/29), Halland (0/34), or Västra Götaland (0/75).[33]
N1a1a (M178)
[edit]The subclade N-M178[Phylogenetics 3] is defined by the presence of markers M178 and P298. N-M178* has higher average frequency in Northern Europe than in Siberia, reaching frequencies of approximately 60% among Finns and approximately 40% among Latvians, Lithuanians & 35% among Estonians.[71][13]
Miroslava Derenko and her colleagues noted that there are two subclusters within this haplogroup, both present in Siberia and Northern Europe, with different histories. The one that they labelled N3a1 first expanded in south Siberia and spread into Northern Europe. Meanwhile, the younger subcluster, which they labelled N3a2, originated in south Siberia (probably in the Baikal region).[71]
N-M178 was also found in two Na-Dené speaking Tłı̨chǫs in North America.[72]
Neolithic samples from Baikal area have yielded plenty of yDNA N specimens, and one sample from Fofonovo, Buryatia, 5000-4000 BC is among the first Tat samples in the ancient record.[68]
Earliest samples of N1a1a-L708 were found in Trans-Baikal (brn008, N1a1a1*-L708; brn003, N1a1a1a1*-M2126) between 8,000 and 6,000 YBP. Downstream samples were found in Yakutia (N4b2, N1a1a1a1a*-Z1979) and Krasnoyarsk Krai (kra001, N1a1a1a1a*-L392), between 5,000 and 4,000 YBP.[73][74]
N1a2 (F1008/L666)
[edit]N1a2a-M128 and N1a2b-B523/P43 are estimated to share a most recent common ancestor in N1a2-F1008/L666 approximately 8,600 [95% CI 7,500 <-> 9,800] years before present,[1] 9,200 years before present,[75] or 9,314 [95% CI 7,419 <-> 11,264] years before present.[3]
At least three of six tested male specimens from the Early Neolithic (ceramic-using hunter-gatherer of approximately 7200–6200 years ago) layer at the Shamanka archaeological site near the southern end of Lake Baikal have been found to belong to N1a2-L666.[58]
N1a2a-M128
[edit]| Haplogroup N-M128 | |
|---|---|
 | |
| Possible place of origin | Asia |
| Ancestor | N1a2 (F1008/L666) |
| Descendants | F710, F1998 |
| Defining mutations | M128 |
| Highest frequencies | Manchus about 1% (0/43 Bijie,[76] 0/30,[19] 30/2938 = 1.0% China total[77] 1/52 = 1.9% Liaoning,[20] 2/35 = 5.7% Xiuyan Manchu Autonomous County),[15] Koreans 0/43 South Korea,[15] 0/25 China,[15] 1/75 = 1.3% South Korea,[20] Xibe 1/41 = 2.4%,[15] Han 0.89% (Shanxi 1.83%, Henan 1.53%, Gansu 1.46%, Hebei 1.41%, Shaanxi 1.40%, Tianjin 1.38%, Shandong 1.17%, Beijing 1.14%, Inner Mongolia 1.13%, Heilungjiang 1.12%, Liaoning 1.12%, Fujian 1.12%, Anhui 0.93%, Jilin 0.86%, Ningxia 0.82%, Jiangsu 0.81%, Yunnan 0.79%, Hubei 0.77%, Zhejiang 0.73%, Taiwan 0.70%, Sichuan 0.54%, Hunan 0.52%, Chongqing 0.51%, Guizhou 0.51%, Guangdong 0.45%, Hainan 0.43%, Xinjiang 0.43%, Shanghai 0.36%, Jiangxi 0.34%, Guangxi 0.23%, Qinghai 0.18%), Mongolians 0.7% (Liaoning 1.9%, Horqin 0.3%, Ulaanbaatar 0.0%), Japanese 0.3% (Tokushima 3.5%, Ibaraki 2.0%, Hiroshima 1.3%, Nagoya 1.0%, Osaka 0.2%, Tokyo 0.0%, Shizuoka 0.0%, Yamaguchi 0.0%), Kazakhs 0.1% (Khoja 1.9%, Baiuly 0.6%, Uysun 0.4%) |
This subclade is defined by the presence of the marker M128.[Phylogenetics 4] N-M128 was first identified in a sample from Japan (1/23 = 4.3%) and in a sample from Central Asia and Siberia (1/184 = 0.5%) in a preliminary survey of worldwide Y-DNA variation.[51] Subsequently, it has been found with low frequency in some samples of the Manchu people, Sibe people, Evenks, Koreans, Han Chinese, Hui, Tibetans, Vietnamese, Bouyei people, Kazakhs, Uzbeks, Uyghurs, Salars, Tu, Mongols, the Buzava tribe of Kalmyks,[29] Khakas, and Komis.[5]
A number of a Han Chinese, an Ooled Mongol, a Qiang, and a Tibetan were found to belong to a sister branch (or branches) of N-M128 under paragroup N-F1154*.[78]
A neolithic sample brn002 (~5,940 BP) in Trans-Baikal was discovered to be an early offshoot upstream of N-M128.[79]
As a genetic testing result of Yelü clan, a royal family of the Liao Dynasty and Khitan descents, it was found to belong to N-F1998, a downstream of N-M128.
According to "The deep population history of northern East Asia from the Late Pleistocene to the Holocene",[80] the basal yDNA N-M128/mtDNA B5b2 HGDP01293 individual[81] occupied a position on the PCA between the Jiangsu Province’s and Anhui province’s specimens, but not far from the Shandong province’s mtDNA R11’B6>R11b specimen,[80] while a later descendant of the yDNA N-M128 clade, belonging to mtDNA R11’B6>R11b, was reported from the ancient DNA of the Western Zhou Cemetery, tomb M18.[82] Based on ancient DNA, the distribution of mtDNA B5b2 after 9500 years ago and prior to 4600 years ago in the direction of the Anhui and Jiangsu provinces from Shandong from the vicinity of the future Shandong Longshan Yinjiacheng site is shown in “Maternal genetic structure in ancient Shandong between 9500 and 1800 years ago”,[83] while the existence of the local Paleolithic Northern East Asian substratum, represented by individuals of the basal died-out yDNA O-M164*, separating from the Southern East Asian yDNA O-M188 and contributing to yDNA C2-M217 ancestors of Altaic and Korean representatives, was shown in "Human genetic history on the Tibetan Plateau in the past 5100 years".[84] Having occupied the position on the PCA between the Jiangsu Province’s and Anhui Province’s specimens, the basal yDNA N-M128/mtDNA B5b2 HGDP01293 individual became a participant of the uniform genetic cline, which spanned from Jiangsu and Anhui individuals to the Tai-speaking Dai people, and from Jiangsu and Anhui individuals to the ancient individual WGM20, belonging to mtDNA M11, of the Yangshao Wanggou site, dated to 5000-5500 years ago, and this ancient age also encompassed ancient yDNA pre-N-M128 Mazongshan individuals[85] and modern yDNA N-M128-affiliated Gansu Province’s individuals, who appeared to be included on the mentioned genetic cline closer to the ancient Henan province’s specimens of the Longshan period ca. 4000 years ago, than to the more genetically basal ancient individual WGM20 of the Yangshao Wanggou site, dated to 5000-5500 years ago.[80]
N1a2b (P43)
[edit]Haplogroup N-P43[Phylogenetics 5] is defined by the presence of the marker P43. Additionally, haplogroup N-P43 is defined by a marker Y3214, which is shared with a younger yDNA O1b2-K14, distributed in Japan (YFull). It has been estimated to be approximately 4,000 to 5,500 years old (TMRCA 4,510 years,[75] TMRCA 4,700 [95% CI 3,800 <-> 5,600] ybp,[1] or 4,727 [95% CI 3,824 <-> 5,693] years before present).[3] It has been found very frequently among Northern Samoyedic peoples, speakers of Ob-Ugric languages, and northern Khakassians, and it also has been observed with low to moderate frequency among speakers of some other Uralic languages, Turkic peoples, Mongolic peoples, Tungusic peoples, and Siberian Yupik people.
The highest frequencies of N-P43 are observed among north-west Siberian populations: 92% (35/38)[5] in a sample of Nganasan, 78% (7/9)[86][14] in a sample of Enets, 78% (21/27)[31] in a sample of Khants, 75% (44/59)[5] in a sample of Tundra Nenets, 69% (29/42)[3] in another sample of Nenets, 60% (15/25)[12] in a sample of Mansi, 57% (64/112)[11] in another sample of Khants, 54% (27/50)[3] in another sample of Nganasan, 45% (40/89)[5] in a sample of Forest Nenets, 38% (18/47)[87] in a third sample of Khants, and 25% (7/28)[12] in a fourth sample of Khants. In Europe, the N-P43 types have their highest frequency of 20% among Volga-Uralic populations. The extreme western border of the spread of N-P43 is Finland, where this haplogroup occurs only at marginal frequency – 0.4%. Yet N-P43 is quite frequent among Vepsas (17.9%), a small Finnic population living in immediate proximity to Finns, Karelians and Estonians.[5]
Haplogroup N-P43 also has been observed with very high frequency (26/29 = 89.7% of a sample from the settlement of Topanov and 19/22 = 86.4% of a sample from the settlement of Malyi Spirin) in samples of Kachins, a Turkic-speaking ethnic group or territorial subgroup of the Khakas people, from Shirinsky District of northern Khakassia.[9] There appears to be a cline through the Sagai (another Turkic-speaking ethnic group that is now considered to be a constituent of the Khakas people), with 46.2% (55/119) of Sagai sampled from Ust'–Es', Esino, Ust'–Chul', and Kyzlas settlements of Askizsky District of central Khakassia belonging to haplogroup N-P43 vs. only 13.6% (11/81) of Sagai sampled from Matur, Anchul', Bol'shaya Seya, and Butrakhty settlements of Tashtypsky District of southern Khakassia belonging to this haplogroup.[9] However, other researchers' samples of Khakas people have exhibited only moderate frequencies of N-P43 or potential N-P43. Derenko et al. (2006) examined a sample of Khakassians (n=53) collected in the settlements of Askiz, Shirinsk, Beisk and Ordzhonikidzevsk districts of Khakass Republic and found that 15 of them (28.3%) belonged to N-LLY22g(xTat).[18] Rootsi et al. (2007) examined a sample of Khakas (n=181) and found that 31 of them (17.1%) belonged to N-P43;[5] retested 174 of the individuals in this sample and found that 27 of them (15.5%) belonged to the N-B478 (Asian/northern Samoyedic) subclade of N-P43 and 2 of them (1.1%) belonged to the N-L1419 (European/Volga Finnic and Chuvash) subclade of N-P43 for a total of 29 (16.7%) N-P43.[3]
Haplogroup N-P43 forms two distinctive subclusters of STR haplotypes, Asian and European, the latter mostly distributed among Finno-Ugric-speaking peoples and related populations.[5]
N1a2b1-B478
[edit]The TMRCA of N-B478 has been estimated to be 3,007 [95% CI 2,171 <-> 3,970] years before present.[3] It is one of the most prevalent Y-DNA haplogroups among indigenous populations of northwestern Siberia: 69.0% (29/42) Nenets, 50.0% (25/50) Nganasan, 22.2% (12/54) Dolgan from Taymyr, 7.0% (3/43) Selkup, 1.6% (1/63) Ob-Ugrian. It is also quite prevalent among populations of Central Siberia, Southern Siberia, and Mongolia: 17.9% (17/95) Tuvan, 15.5% (27/174) Khakas, 13.0% (6/46) Tozhu Tuvans,[28] 8.7% (2/23) Shor, 8.3% (2/24) Even, 8.2% (5/61) Altaian, 5.3% (3/57) Evenk, 5.0% (19/381) Mongol, 4.9% (3/61) Sart-Kalmak (partial descendants of Oirat Mongols in Kyrgyzstan),[28] 4.2% (9/216) Yakut, 2.1% (1/47) Torgut (Mongolia),[28] 1.4% (1/69) Derbet (Kalmykia),[28] 0.9% (1/111) Buryat. A geographically outlying member has been found in a sample of Chuvash (1/114 = 0.88%).[3]
Karafet et al. (2018) have found N-P63, which appears to be roughly phylogenetically equivalent to N-B478, in 91.2% (31/34) Nganasan, 63.8% (30/47) Tundra Nenets, 42.7% (35/82) Forest Nenets, 14.0% (8/57) Dolgan, 7.0% (9/129) Selkup, 3.3% (3/91) Evenk, 2.7% (2/75) Mongol, 2.6% (2/78) Komi, 2.5% (2/80) Buryat, and 2.0% (2/98) Altai Kizhi.[6] This haplogroup was not observed in samples of Yukaghir (0/10), Koryak (0/11), Teleut (0/40), Ket (0/44), Yakut (0/62), or Khanty (0/165) populations.[6]
Kharkov et al. (2023) have found N-B478 in greatly differing percentages of samples of Khanty from two different villages of Khanty-Mansi Autonomous Okrug: 60.9% (39/64) of a sample of Khanty from the village of Russkinskaya, Surgut district and 14.8% (8/54) of a sample of Khanty from the village of Kazym, Beloyarsky district. Five of the eight members from the village of Kazym share a subclade marked by the B172 and Z35108 SNPs with all previously surveyed Nenets men from the Vanuito phratry belonging to the Vanuito, Puiko and Yaungat clans and the Purungui clan of Khanty origin.[88]
N1b (F2930)
[edit]Haplogroup N1b has been predominantly found in the Yi people, a Tibeto-Burman speaking ethnic group in southwestern China who originated from ancient Qiang tribes in northwestern China.[55] However, it also has been found in people all over China (where they account for approximately 3.62% of the country's male population and are mainly distributed in Shandong, Jiangsu, Zhejiang, Anhui, etc.[89]) and in some individuals from Spain,[59] Ecuador,[59] Poland,[59][1] Belarus,[59] Russia,[59][1] Iraq,[59] India,[59][1] Kazakhstan,[59] Korea,[59][1] Japan,[59][1] Bhutan,[59] Vietnam,[59][1] Cambodia,[59][1] Laos,[59] Thailand,[59][1] Malaysia,[1] and Singapore.[1]
N2 (Y6503)
[edit]N2 (Y6503/FGC28528; B482/FGC28394/Y6584) – a primary branch of haplogroup N-M231, is now represented mainly by a subclade, N-FGC28435, that has spread probably some time in the first half of the second millennium CE[90] and that has been found in individuals from Serbia, Croatia, Bosnia and Herzegovina, Montenegro, and Turkey (Istanbul).[91][90]
N-Y7310 (or N-F14667) subsumes N-FGC28435 and likewise probably descends from a common ancestor who has lived some time in the first half of the last millennium. However, members of N-Y7310(xFGC28435) exhibit a greater geographic range, including an individual from Rostov Oblast of Russia and a Romanian Hungarian individual with ancestry from Suceava, Bukovina.[1]
Other branches of N-P189 include members from Turkey,[1] Russia (Moscow Oblast),[1] France (Charente-Maritime),[1] and England (Devon).[1][59] The most recent common ancestor of all the aforementioned extant N-P189 lineages has been estimated to be 4,900 (95% CI 5,700 <-> 4,100) years before present.[1] An archaeological specimen attributed to the Botai culture of northern Kazakhstan and dated to the latter half of the fourth millennium BCE belongs to N-P189*, being basal to present-day European members of N-P189.[92][1]
Lineages that belong to N-Y6503(xP189) and are only distantly related (with a time to most recent common ancestor estimated to be greater than 10,000 years before present)[1] to the aforementioned members of N-P189 have been found in an individual from the present-day Altai Republic[1] and probably also in an archaeological specimen attributed to the Iron Age Mezőcsát culture of what is now Hungary (approx. 2,900 years before present)[93] and in an archaeological specimen attributed to the Kitoi culture of ceramic-using foragers of the area around Lake Baikal (approx. 6,700 years before present).[92]
Ancient peoples
[edit]A sample excavated at the Houtaomuga site in the Yonghe neighborhood of Honggangzi Township, Da'an, Jilin, China dating back to 7430–7320 years ago (Phase II of the Early Neolithic) has been found to belong to Y-DNA haplogroup N and mtDNA haplogroup B4c1a2. This sample is autosomally identical with the Neolithic Amur River Basin populations, of which Nivkh people are the closest modern representative. As the paper detected this ancestry in terminal Pleistocene USR1 specimen in Alaska, it is therefore, postulated that there was gene flow from Amur to America of a population belonging to a hypothetical Chukotko-Kamchatkan–Nivkh linguistic family.[citation needed]
N has also been found in many samples of Neolithic human remains exhumed from Liao civilization in northeastern China, and in the circum-Baikal area of southern Siberia.[66] It is suggested that yDNA N, reached southern Siberia from 12 to 14 kya. From there it reached southern Europe 8-10kya.[52]
Phylogeny
[edit]Phylogenetic tree
[edit]In the following tree the nomenclature of three sources is separated by slashes: ISOGG Tree 10 December 2017 (ver.12.317)
- NO-M214
- Н-М231/Пейдж91, М232/М2188
- N1-Z4762/CTS11499/L735/M2291
- N1a-L729
- Н-З1956
- N-Y149447 China (Shaanxi[1])
- Н1а1-М46/Страница70/Тат
- Н1а1а-М178
- N1a1a1-F1419
- N1a1a1a-L708
- N1a1a1a1-P298/M2126 China (Urumqi, Kashgar Prefecture, Turpan, Aksu Prefecture, Xianyang, Jincheng, Kaifeng, Qiqihar[75])
- N1a1a1a1a-L392
- N1a1a1a1a1-CTS10760
- N1a1a1a1a1a-CTS2929/VL29 Found with high frequency among Lithuanians, Latvians, Estonians, northwestern Russians, Swedish Saami, Karelians, Nenetses, Finns, and Maris, moderate frequency among other Russians, Belarusians, Ukrainians, and Poles, and low frequency among Komis, Mordva, Tatars, Chuvashes, Dolgans, Vepsa, Selkups, Karanogays, and Bashkirs[3]
- N1a1a1a1a1a1-Z4908
- N-Y46443
- N-Y46443* Russia (Moscow Oblast[1])
- N-BY33095 Russia (Samara Oblast[1])
- N1a1a1a1a1a1a-L550/S431
- N-L550* Sweden (Kronoberg County[1])
- N-Y9454 Sweden (Västra Götaland County,[1] Örebro County[1]), Finland,[1] Russia[1]
- N-Y20911 Finland (Western Finland Province,[1] Southern Finland Province[1])
- N-Y7795
- N-Y7795* Sweden (Norrbotten County,[1] Södermanland County[1])
- N-Y29766 Norway (Hedmark[1])
- N-Y20918 Sweden (Östergötland County,[1] Västerbotten County[1]), Finland (Western Finland Province[1])
- N-Y28771
- N-Y61225 Sweden (Södermanland County,[1] Uppsala County[1])
- N-Y30126
- N-Y30126* Sweden (Östergötland County[1])
- N-Y29764
- N-Y29764* Sweden (Stockholm County[1])
- N-Y30123 Sweden (Västerbotten County[1]), Finland[1]
- N-S9378
- N-S9378* Sweden (Västra Götaland County[1])
- N-S18447 Sweden (Södermanland County,[1] Uppsala County[1])
- N-Y36282
- N-Y36282* Finland (Southern Finland Province[1]), Estonia (Ida-Virumaa[1])
- N-BY21957 Poland (Kujawsko-pomorskie[1]), Sweden (Stockholm County[1]), Russia (Pskov Oblast[1]), Finland (Southern Finland Province[1])
- N-FGC14542
- N-Y4341
- N-BY21874 Sweden (Södermanland County[1]), Finland[1]
- N-Y4338
- N-Y4338*
- N-Y4339 Sweden (medieval Sigtuna[1])
- N-Y12104
- N-Y19111
- N-Y57577 Sweden (Skåne County[1])
- N-Y22774 Finland (Western Finland Province[1])
- N-Y5611
- N-Y5611* England[1]
- N-Y21546 Sweden (Stockholm County,[1] Västmanland County[1])
- N-F1983 Sweden (Gävleborg County[1]), Russia (Lipetsk Oblast[1])
- N-Y10932
- N-Y85136 Sweden (Uppsala County[1])
- N-Y10931 Russia[1]
- N1a1a1a1a1a1a1-L1025/B215 Highest frequency among Lithuanians, significant in Latvians and Estonians and lesser frequency in Belarusians, Ukrainians, South-West Russians, and Poles. With exception of Estonians, L1025 has highest share among N-M231 clades in previously mentioned populations. Also observed in Finland and Sweden, with sporadic instances in Norway, Germany, Netherlands, United Kingdom, the Azores, Czech Republic, and Slovakia.
- N-L1025* Russia (Kursk Oblast[1])
- N-BY30389 Sweden (Västernorrland County[1]), Finland (Western Finland Province[1])
- N-Y13982
- N-Y13982* Portugal (Azores[1])
- N-Y31236 Lithuania (Marijampolė County,[1] Vilnius County[1])
- N-A11940
- N-Y140872 Sweden (Södermanland County,[1] Norrbotten County[1]), Finland (Southern Finland Province[1])
- N-Y143451 Russia (Bashkortostan[1])
- N-Y5580
- N-Y93996 Lithuania (Šiauliai County[1]), Poland (Podlaskie[1])
- N-BY158 Russia (Voronezh Oblast,[1] Smolensk Oblast,[1] Tatarstan[1]), Belarus (Vitebsk Region[1]), Lithuania (Vilnius County,[1] Šiauliai County,[1] Panevėžys County[1]), Poland (Podlaskie[1]), Ukraine (Poltava Oblast[1]), Kazakhstan (Jambyl Region[1])
- N-Z16975
- N-VL69 Belarus (Minsk Region[1]), Russia (Bryansk Oblast[1]), Kazakhstan (Kostanay Region[1])
- N-Z16976
- N-Z16976* Belarus (Grodno Region[1]), Ukraine (Khmelnytskyi Oblast[1])
- N-Y13475 Poland,[1] Lithuania[1]
- N-Y21578 Lithuania (Kaunas County,[1] Telšiai County[1]), Russia (Smolensk Oblast[1])
- N-Y6129 Poland (Warmińsko-mazurskie[1]), Lithuania[1]
- N-Y19113 Poland (Warmińsko-mazurskie[1]), Lithuania (Tauragė County[1])
- N-Y134492 Belarus (Grodno Region,[1] Minsk Region,[1] Mogilev Region[1]), Lithuania,[1] United States (Illinois[1])
- N-L551
- N-L551* Lithuania (Vilnius County[1])
- N-Y15251 Lithuania (Alytus County[1]), Poland (Podlaskie[1])
- N-Y46313 Latvia[1]
- N-Y86578 Russia (Belgorod Oblast[1])
- N-Y14152
- N-BY21911 Poland,[1] Latvia (Krāslava District[1]), Ukraine (Kyiv Oblast[1]), Finland (Southern Finland Province[1])
- N-Y13979 Lithuania,[1] Russia (Ryazan Oblast[1]), Germany[1]
- N-Y4706
- N-Y4706* Sweden (Stockholm County,[1] Södermanland County,[1] Västra Götaland County[1]), Germany (Lower Saxony[1]), Finland (Southern Finland Province[1]), Russia (Tatarstan[1])
- N-A705 Sweden (Kalmar County,[1] Östergötland County[1])
- N-BY21893 Poland (Wielkopolskie[1])
- N-Y139030 Sweden (Västerbotten County[1]), Norway (Østfold[1])
- N-Y183040 Russia (Vologda Oblast,[1] Tatarstan[1])
- N-Y4707 Finland (Western Finland Province,[1] Åland,[1] Southern Finland Province,[1] Eastern Finland Province,[1] Oulu Province[1])
- N-Z16981
- N-A2358=N-BY14135 England,[1] Finland,[1] Latvia (Daugavpils apriņķis[1])
- N-CTS8173 Estonia (Lääne-Virumaa[1])
- N-FT213922 [59])
- N-BY32524 Estonia (Järvamaa[1]), Finland (Oulu Province[1])
- N-FGC39882 Lithuania (Telšiai County[1])
- N-FT6082 [59])
- N-A11470 Russia (Tyumen Oblast[1]), Lithuania (Panevėžys County[1]), Finland (Southern Finland Province[1]), Netherlands (South Holland,[1] North Brabant[1])
- N-FTC49461 Latvia (Jaunpils Parish, Tukuma apriņķis[59])
- N-FTE38475 Latvia (Jaunpils Parish, Tukuma apriņķis[59])
- N-FTE39526 Latvia (Jaunpils Parish, Tukuma apriņķis[59])
- N-FTE38475 Latvia (Jaunpils Parish, Tukuma apriņķis[59])
- N-BY14147 Finland, Belarus[59])
- N-BY52958 Latvia[59])
- N-FT109465 Latvia[59])
- N-FTD92343 Latvia, Ukraine[59])
- N-BY202983 Latvia[59])
- N-BY40639 Latvia, Russia[59])
- N-FTC49461 Latvia (Jaunpils Parish, Tukuma apriņķis[59])
- N-BY21926 Russia, Latvia, Belarus, Lithuania[59])
- N-A11470 Russia (Tyumen Oblast[1]), Lithuania (Panevėžys County[1]), Finland (Southern Finland Province[1]), Netherlands (South Holland,[1] North Brabant[1])
- N-Y15922=N-Z35238 Finland (Southern Finland Province[1]), Estonia (Ida-Virumaa,[1] Pärnumaa[1]), Latvia (Valmiera,[1] Liepāja,[1] Talsu apriņķis[1]), Russia (Pskov Oblast[1]), Poland (Pomorskie[1])
- N-Y6075 Poland (Małopolskie,[1] Mazowieckie[1]), Ukraine (Lviv Oblast[1]), Slovakia,[1] Czech Republic (Moravian-Silesian Region[1]), United States (New Jersey[1])
- N-Y11882
- N-Y11882* Russia (Tatarstan[1])
- N-ZS11617 Russia (Yaroslavl Oblast[1]), Lithuania[1]
- N-Y24601 Latvia[1]
- N-Y94659 Lithuania,[1] Belarus (Minsk Region[1])
- N-Y32725
- N-FT96305 Latvia (Krāslava District[1]), Estonia (Viljandimaa[1])
- N-Y33333 Ukraine (Sumy Oblast[1]), Belarus (Brest Region[1])
- N-FT213922 [59])
- N-Y46443
- N1a1a1a1a1a2-CTS9976
- N-L1022
- N-Y19098
- N-Y19098* Sweden (Västernorrland County[1])
- N-A12258 Финляндия (провинция Восточная Финляндия) [ 1 ] )
- N-BY117178 Финляндия (провинция Восточная Финляндия) [ 1 ] )
- N-Y5004
- Н-Y7300
- N-A17632
- N-Y128024 Швеция ( округ Эстергётланд) [ 1 ] )
- N-A16017
- N-Y7297 Финляндия (провинция Западная Финляндия, [ 1 ] Южная Финляндия, [ 1 ] Лапландия [ 1 ] )
- N-Y15813
- N-Y15813* Финляндия (провинция Западная Финляндия) [ 1 ] )
- N-Y49008 Финляндия (провинция Западная Финляндия, [ 1 ] Провинция Оулу [ 1 ] )
- N-Y46886 Финляндия (провинция Западная Финляндия) [ 1 ] )
- N-Y15812
- N-A14187 Финляндия (провинция Западная Финляндия) [ 1 ] ), Эстония ( Ляэнемаа [ 1 ] )
- N-Y24617 Финляндия (провинция Западная Финляндия) [ 1 ] ), Швеция ( округ Даларна [ 1 ] ), Россия ( Курская область [ 1 ] )
- N-Y23576
- N-Y23576* Финляндия (провинция Восточная Финляндия) [ 1 ] )
- N-Y15615
- N-A17632
- N-Y5005
- N-Y22106
- N-Y47789 Финляндия (провинция Западная Финляндия) [ 1 ] )
- N-Y79341 Финляндия (провинция Западная Финляндия) [ 1 ] ), Швеция ( округ Вестра-Гёталанд , [ 1 ] Лен Вермланд [ 1 ] )
- N-Y10756
- N-A13656 Финляндия (провинция Западная Финляндия, [ 1 ] Лапландия [ 1 ] ), Швеция ( округ Даларна [ 1 ] )
- Н-PH2196
- N-CTS11122 Финляндия (провинция Восточная Финляндия) [ 1 ] )
- N-A17277 Финляндия (Западная Финляндия, [ 1 ] Южная Финляндия [ 1 ] )
- N-PH547 Финляндия (Лапландия, [ 1 ] Восточная Финляндия, [ 1 ] Южная Финляндия [ 1 ] ), Швеция ( графство Вестерботтен , [ 1 ] Лен Норрботтен [ 1 ] )
- N-Y5003
- N-Y5003* Эстония ( Хийумаа [ 1 ] ), Финляндия (провинция Южная Финляндия [ 1 ] ), Великобритания ( Вестминстер [ 1 ] )
- N-BY22001 Эстония ( Ляэнемаа [ 1 ] ), Финляндия (провинция Западная Финляндия [ 1 ] ), Швеция (округ Норботтен [ 1 ] )
- N-BY6007 Россия ( Липецкая область [ 1 ] )
- N-Y132182 Швеция ( округ Вермланд) [ 1 ] ), Эстония ( Вильяндимаа [ 1 ] )
- N-Y20917 Швеция ( округ Кроноберг) [ 1 ] ), Финляндия (Западная Финляндия, [ 1 ] Южная Финляндия [ 1 ] )
- N-Y24502 Великобритания ( Шотландские границы , [ 1 ] Колдердейл , [ 1 ] Кирклис [ 1 ] )
- N-Y18420 Финляндия (провинция Южная Финляндия, [ 1 ] Западная Финляндия [ 1 ] )
- N-Z35267 Финляндия (Западная Финляндия, [ 1 ] Аландская провинция , [ 1 ] Южная Финляндия [ 1 ] ), Эстония ( Пярнумаа [ 1 ] )
- N-Y6599 Финляндия (провинция Восточная Финляндия, [ 1 ] Провинция Оулу, [ 1 ] Южная Финляндия [ 1 ] ), Эстония ( Йыгевамаа , [ 1 ] Хийумаа [ 1 ] ), Россия ( Тульская область [ 1 ] )
- Н-Y24000
- N-Y16503 Финляндия (провинция Западная Финляндия, [ 1 ] Южная Финляндия, [ 1 ] Восточная Финляндия, [ 1 ] Провинция Оулу [ 1 ] ), Швеция ( графство Евлеборг [ 1 ] )
- N-Y22106
- Н-Y7300
- N-Y19098
- N-BY6010 США ( Вирджиния) [ 1 ] )
- Н-CTS3451
- N-CTS3451* Русский [ 1 ]
- N-Y3667
- N-BY33087
- N-BY33087* Финляндия ( провинция Южная Финляндия) [ 1 ] )
- N-BY33088 Финляндия ( провинция Восточная Финляндия) [ 1 ] ), Россия ( Новгородская область [ 1 ] )
- Н-CTS657
- N-CTS657* Финляндия, [ 1 ] Россия ( Республика Карелия , [ 1 ] Новгородская область [ 1 ] )
- N-BY70437 Финляндия (провинция Восточная Финляндия) [ 1 ] )
- N-BY6024 Финляндия (провинция Восточная Финляндия) [ 1 ] )
- N-Y45925 Россия (Республика Карелия) [ 1 ] )
- N-Y26750
- N-Y26750* Russia ( Arkhangelsk Oblast [ 1 ] )
- N-A16653 Финляндия (провинция Восточная Финляндия) [ 1 ] )
- N-PH3568 Финляндия ( провинция Оулу , [ 1 ] Восточная Финляндия [ 1 ] )
- N-BY33087
- N-L1022
- N1a1a1a1a1a1-Z4908
- N1a1a1a1a1b-PH1266/Y28526/F4134
- N-Y46746 Россия [ 1 ]
- N-Y32732
- N-Y32732* Россия ( Санкт-Петербург [ 1 ] )
- N-Y192174
- N-Y32731 Россия ( Республика Коми [ 1 ] ), Швеция [ 1 ]
- N1a1a1a1a1c-B479 Нанай , [ 3 ] Ульчи [ 40 ]
- N1a1a1a1a1a-CTS2929/VL29 Found with high frequency among Lithuanians, Latvians, Estonians, northwestern Russians, Swedish Saami, Karelians, Nenetses, Finns, and Maris, moderate frequency among other Russians, Belarusians, Ukrainians, and Poles, and low frequency among Komis, Mordva, Tatars, Chuvashes, Dolgans, Vepsa, Selkups, Karanogays, and Bashkirs[3]
- N1a1a1a1a2-Z1936,CTS10082 Встречается с высокой частотой среди финнов , вепсов , карел , шведских саамов , северо-западных русских , башкир и волжских татар , умеренная частота среди других русских , коми , ненцев , обских угров , долган и сибирских татар , и низкая частота среди мордвы , нганасан , чувашей , эстонцев , латышей , украинцев и караногаев [ 3 ]
- N-Y13851
- N-BY199053 Франция, [ 59 ] ancient Western Siberia (Gornaya Bitiya, Iron Age Sargat culture , 300 - 100 BCE [ 59 ] )
- N-Y13852
- N1a1a1a1a2a1c-PH3340/Y13850
- N1a1a1a1a2a1c1-L1034
- N-Y28538
- N-Y28538* Россия ( Хантия-Мансия [ 1 ] )
- N-L1032 Россия (Хантия-Мансия [ 1 ] ), Казахстан ( Восточно-Казахстанская область [ 1 ] )
- Н-L1442
- Н-FT12605
- N-Y23732 Russia ( Bashkortostan [ 1 ] )
- N-Y24222
- N-Y24222* Венгрия [ 1 ]
- N-Y65329 Россия (Татарстан татарский , [ 1 ] Bashkortostan [ 1 ] )
- N-Y62987 Russia (Bashkortostan, [ 1 ] Самарская область , [ 1 ] Тюменская область Сибирский татарин [ 1 ] )
- N-Y28538
- N1a1a1a1a2a1c2-Y24361 (Y24372)
- N1a1a1a1a2a1c1-L1034
- N1a1a1a1a2a1c-PH3340/Y13850
- Н1а1а1а1а2а-З1934
- N-Y159520
- N-Y159520* Russia ( Tambov Oblast [ 1 ] )
- N-BY184755 Россия ( Новгородская область , [ 1 ] Воронежская область , [ 1 ] Москва [ 1 ] )
- N-Y18421
- N-Y18421* Russia (Tambov Oblast [ 1 ] )
- N-Y19110
- N-Y180247
- N-Y19108
- N-Y19108* Россия (Татарстан, [ 1 ] Kirov Oblast [ 1 ] )
- N-Y19453
- N-Y19453* Россия (Татарстан [ 1 ] )
- N-Y19451 Финляндия (провинция Западная Финляндия) [ 1 ] ), Россия ( Республика Карелия , [ 1 ] Tver Oblast [ 1 ] )
- Н-З1928/CTS2733
- N-YP6091
- N-YP6091* Россия ( Иркутская область [ 1 ] )
- N-YP6094
- N-Y129131
- N-Y129131* Russia ( Kostroma Oblast [ 1 ] )
- N-A25107 Россия ( Кировская область , [ 1 ] ВВП России [ 1 ] )
- N-YP6092
- N-YP6092* Russia ( Vologda Oblast [ 1 ] )
- Н-Б195
- N-B195* Russia ( Sverdlovsk Oblast [ 1 ] )
- N-Y62142 Россия ( Республика Карелия , [ 1 ] Архангельская область [ 1 ] )
- N-Y129131
- N1a1a1a1a2a-Z1925
- N-Z1925* Швеция ( округ Норрботтен) [ 1 ] )
- N-Y29767 Швеция ( округ Норрботтен) [ 1 ] )
- N-Y62904 Финляндия ( провинция Западная Финляндия , [ 1 ] Провинция Оулу [ 1 ] )
- N1a1a1a1a2a2a1a1-Z1926
- N-Y20920
- N-Y20920* Финляндия ( провинция Западная Финляндия) [ 1 ] ), Норвегия ( Норд-Трёнделаг [ 1 ] )
- N-Y21699 Финляндия ( провинция Западная Финляндия , [ 1 ] Южная Финляндия [ 1 ] )
- Н-З1927
- N-Y22108 Финляндия (провинция Восточная Финляндия, [ 1 ] Южная Финляндия, [ 1 ] Западная Финляндия [ 1 ] ), Швеция ( графство Стокгольм , [ 1 ] Округ Эребру [ 1 ] )
- Н-З1933
- Н-CTS4329
- N-Y22091
- N-Y22091* Норвегия ( Нордланд [ 1 ] ), Финляндия ( Провинция Лапландия [ 1 ] )
- N-Y47623
- N-Y31247
- N-Y31247* Russia ( Tver Oblast [ 1 ] )
- N-Y31249
- N-Y90283 Финляндия (провинция Лапландия) [ 1 ] )
- N-Y31244 Финляндия ( провинция Западная Финляндия , [ 1 ] Восточная Финляндия [ 1 ] )
- N-YP6269
- N-YP6269* Финляндия (провинция Западная Финляндия) [ 1 ] )
- N-Y151660
- N-Y151660* Финляндия (провинция Восточная Финляндия) [ 1 ] )
- N-BY149208 Швеция ( округ Норрботтен) [ 1 ] )
- Н-CTS8565
- N-BY18114 Финляндия (провинция Восточная Финляндия) [ 1 ] )
- N-BY30394 Финляндия (провинция Восточная Финляндия) [ 1 ] ), Россия ( Удмуртская Республика [ 1 ] )
- N-Y30513
- N-Y30513* Финляндия ( провинция Южная Финляндия) [ 1 ] )
- N-Y29759
- Н-Z4998
- N-Z4998* Финляндия (провинция Южная Финляндия) [ 1 ] )
- N-FGC65190 Финляндия (провинция Южная Финляндия, провинция Восточная Финляндия)
- N-Y18414 Финляндия (провинция Западная Финляндия)
- N-Y20910 Финляндия (провинция Западная Финляндия)
- N-BY194138 Финляндия (провинция Южная Финляндия)
- N-Y28547 Финляндия (провинция Восточная Финляндия)
- N-BY22141 Финляндия (провинция Западная Финляндия, провинция Восточная Финляндия), США (Вашингтон)
- N-FT5834 Финляндия (Западная Финляндия, Южная Финляндия)
- N-BY190112 Финляндия (провинция Западная Финляндия, провинция Южная Финляндия)
- N-Y19097 Финляндия (провинция Восточная Финляндия, провинция Южная Финляндия, провинция Западная Финляндия, провинция Оулу)
- N-CTS3223 Финляндия (Южная Финляндия, Восточная Финляндия, Западная Финляндия, Оулу, Лапландия), Швеция (Норрботтен, Вестерботтен, Даларна, Вермланд, Сконе), Россия ( Ленинградская область )
- N-Y22091
- Н-ВЛ62 Россия ( Алтайский край , [ 1 ] Костромская область [ 1 ] ), Эстония ( Харьюмаа [ 1 ] ), Финляндия [ 1 ]
- N-VL62* Russia ( Chelyabinsk Oblast [ 1 ] ), Финляндия [ 1 ]
- Н-ВЛ60
- N-VL60* Russia ( Kurgan Oblast [ 1 ] )
- N-Y63781 Россия ( Владимирская область , [ 1 ] Архангельская область [ 1 ] )
- Н-ВЛ72
- Н-ВЛ72* Россия ( Республика Карелия [ 1 ] )
- N-BY30470 Финляндия ( провинция Западная Финляндия) [ 1 ] )
- N-Y20915 Финляндия ( провинция Восточная Финляндия) [ 1 ] )
- Н-З1939
- N-Y132561
- Н-Z2445
- N-Z35031 Финляндия (провинция Восточная Финляндия, провинция Западная Финляндия), Швеция (лен Вестра-Гёталанд, округ Вестманланд)
- N-BY28931
- Н-CTS7189
- N-BY142665 Финляндия (провинция Восточная Финляндия) [ 1 ] )
- N-Y37149 Финляндия (провинция Восточная Финляндия) [ 1 ] )
- N-Y23568 Финляндия (провинция Восточная Финляндия) [ 1 ] )
- N-Y80226 Финляндия (провинция Восточная Финляндия) [ 1 ] )
- N-FT20730 Финляндия (провинция Восточная Финляндия) [ 1 ] ), Россия (Республика Карелия [ 1 ] )
- N-BY62666 Финляндия (провинция Восточная Финляндия) [ 1 ] ), Норвегия (Осло [ 1 ] ), США (Вирджиния, [ 1 ] Арканзас [ 1 ] )
- N-Y24218
- N-BY22038 Финляндия (провинция Восточная Финляндия, [ 1 ] Южная Финляндия, [ 1 ] Западная Финляндия [ 1 ] )
- N-Y13974 Финляндия (провинция Южная Финляндия, [ 1 ] Восточная Финляндия, [ 1 ] Провинция Оулу, [ 1 ] Западная Финляндия [ 1 ] )
- N-Y135981 Финляндия (провинция Восточная Финляндия, [ 1 ] Провинция Оулу, [ 1 ] Южная Финляндия, [ 1 ] Западная Финляндия [ 1 ] ), Норвегия ( Финнмарк [ 1 ] )
- Н-Z4747
- N-Y17790 Финляндия (провинция Восточная Финляндия, [ 1 ] Южная Финляндия, [ 1 ] Западная Финляндия [ 1 ] )
- Н-З1941
- N-BY22090 Финляндия (провинция Южная Финляндия, [ 1 ] Восточная Финляндия [ 1 ] )
- N-Y21575 Финляндия (провинция Западная Финляндия, [ 1 ] Южная Финляндия, [ 1 ] Восточная Финляндия, [ 1 ] Провинция Оулу, [ 1 ] Лапландия [ 1 ] ), Россия ( Ленинградская область , [ 1 ] Москва , [ 1 ] Татарстан [ 1 ] )
- N-Z1940 Финляндия (Западная Финляндия, [ 1 ] Южная Финляндия, [ 1 ] Восточная Финляндия, [ 1 ] Провинция Оулу, [ 1 ] Лапландия [ 1 ] ), Россия (Ленинградская область [ 1 ] ), Швеция (лен Вермланд [ 1 ] )
- Н-CTS4329
- N-Y20920
- N-YP6091
- N-Y159520
- N-Y13851
- N1a1a1a1a3-B197/Y16323
- N1a1a1a1a3a-F4205 Часто встречается среди бурят. [ 3 ] и Цаатаны , [ 28 ] умеренная частота среди караногаев , [ 3 ] Тувинцы , [ 3 ] тоджинс , [ 28 ] и монголы , [ 3 ] [ 28 ] и низкая частота среди алтайцев , [ 3 ] сибирские татары , [ 3 ] Казахи , [ 3 ] Эвенки , [ 3 ] крымские татары , [ 3 ] Каракалпаки , [ 3 ] Узбеки , [ 3 ] и украинцы [ 3 ]
- N-F4205* Китай (Внутренняя Монголия, Ганьсу, Синьцзян, Хэнань и т. д.) [ 75 ] )
- N-Y16221
- N-Y16221* Китай ( Внутренняя Монголия) [ 1 ] )
- N-Y16220
- N-Y16220* Украина ( Луганская область [ 1 ] )
- N-F22331 Турция ( Мерсин [ 1 ] ), Польша ( Малопольское воеводство [ 1 ] ), Kazakhstan ( Pavlodar Region ca. 1680 ybp [ 1 ] )
- N-Y63966 Казахстан, [ 1 ] Турция, [ 1 ] Узбекистан [ 1 ]
- N-Y226011
- N-Z35331 Китай ( город Ордос , Хайдун , Чэнду) [ 75 ] )
- Н-Б199 Россия (Бурятия) [ 1 ] ), Китай (Хуасай из Хулунбуира [ 75 ] )
- N-FT411781 Китай (Внутренняя Монголия) [ 1 ] )
- N-Y16312/F2288
- N-Z35331 Китай ( город Ордос , Хайдун , Чэнду) [ 75 ] )
- N1a1a1a1a3b-B202 Встречается с высокой частотой среди чукчей , [ 3 ] Коряки , [ 3 ] и сибирские эскимосы [ 3 ]
- N1a1a1a1a3a-F4205 Часто встречается среди бурят. [ 3 ] и Цаатаны , [ 28 ] умеренная частота среди караногаев , [ 3 ] Тувинцы , [ 3 ] тоджинс , [ 28 ] и монголы , [ 3 ] [ 28 ] и низкая частота среди алтайцев , [ 3 ] сибирские татары , [ 3 ] Казахи , [ 3 ] Эвенки , [ 3 ] крымские татары , [ 3 ] Каракалпаки , [ 3 ] Узбеки , [ 3 ] и украинцы [ 3 ]
- N1a1a1a1a1-CTS10760
- N1a1a1a1b-M2019/M2118 Эстония, [ 1 ] [ 94 ] Китай ( Хами , Яньань , Сиань , Люлян , Чанчжи , Лига Силингол , Чифэн , Цанчжоу , Сунъюань [ 75 ] )
- N-M2058 Китай (Шаньдун, Ляонин, Хэнань, Внутренняя Монголия, Ганьсу и т. д.) [ 75 ] [ 1 ] )
- N-M2016 Китай (особенно Синьцзян и Внутренняя Монголия). [ 75 ] )
- N1a1a1a1b2-A9408 Китай (Хэбэй, Шаньдун, Хэнань и др.) [ 75 ] ), Ливан [ 53 ] [ 3 ]
- N-Y70200 Китай [ 1 ] (Шэньси, Шаньси, Шаньдун и т. д. [ 75 ] )
- N-PH1612 Китай ( Юлинь , Шанцю , Дэчжоу , Таншань , Шэньян [ 75 ] )
- N-A9407/A9411 China ( Ili Kazakh Autonomous Prefecture [ 75 ] )
- N-A9416 Венгрия ( Сабольч-Сатмар-Берег [ 1 ] ), Хорватия ( Задарская жупания [ 1 ] ), Россия ( Чувашская Республика [ 1 ] )
- N-PH1896 Венгрия ( Боршод-Абауй-Земплен [ 1 ] ), Турция ( Хатай [ 1 ] [ 94 ] ), Китай (Синьцзян, Шаньдун, Аньхой, Хэйлунцзян, Хэбэй, Нинся, Гуйчжоу [ 75 ] )
- N-A9407/A9411 China ( Ili Kazakh Autonomous Prefecture [ 75 ] )
- N-M2058 Китай (Шаньдун, Ляонин, Хэнань, Внутренняя Монголия, Ганьсу и т. д.) [ 75 ] [ 1 ] )
- N1a1a1a1a-L392
- Н1а1а1а2-Б211 (Y9022) Удмуртский , [ 3 ] Приходить , [ 3 ] Чуваш , [ 3 ] Ob-Ugrians , [ 3 ] Большой , [ 3 ] Мордовия , [ 3 ] Алтай , [ 3 ] Белорусский , [ 3 ] Караногай , [ 3 ] карельский , [ 3 ] Башкирский , [ 3 ] татарин , [ 94 ] Русский , [ 3 ] [ 94 ] хакасы [ 3 ]
- N1a1a1a2a-B181
- N-Y182299 Россия ( Кировская область и др.) [ 1 ]
- N-Y23788 Russia ( Bashkortostan , Komi Republic , etc.) [ 1 ]
- N1a1a1a2b-Y23181
- N-Y23183 Russia ( Tatarstan , Mordovia , Penza Oblast ) [ 1 ]
- N-Y143277 Russia ( Tatarstan , Bashkortostan ) [ 1 ]
- N1a1a1a2a-B181
- N1a1a1a1-P298/M2126 China (Urumqi, Kashgar Prefecture, Turpan, Aksu Prefecture, Xianyang, Jincheng, Kaifeng, Qiqihar[75])
- N-Y24317
- N1a1a1a-L708
- N1a1a2-Y23747 Япония ( остров Садо [ 94 ] )
- N1a1a2a-Y23749 Япония [ 3 ] ( Айти , [ 1 ] Хоккайдо [ 1 ] )
- N1a1a2b-Y125664
- N-Y125664* Шанхай [ 1 ]
- N-MF16376
- N1a1a2b1-MF38607 Аньхой , [ 1 ] Себя [ 1 ]
- N1a1a2b2-F22150 Хэбэй , [ 1 ] Сучжоу [ 75 ]
- N-FT281705 Китай (Шаньдун, Ляонин и др.) [ 75 ] )
- N-MF15288/MF15344 Китай (особенно Цзянсу, но также Хэнань, Фуцзянь, Шанхай, Хэйлунцзян , [ 1 ] и т. д. [ 75 ] ), Южная Корея [ 1 ]
- N-MF55680 Китай (Хэбэй, Пекин, Хубэй и т. д.) [ 75 ] )
- N1a1a1-F1419
- Н1а1а-М178
- Н-F1360
- N-F1360* Китай ( Шэньси [ 1 ] )
- Н1а2-Ф1008/Л666
- Н-F1101
- N-F1101* Китай ( Шаньдун [ 1 ] )
- Н-F1154
- N-F1154* Китай ( Ляонин [ 1 ] )
- N-Y23741
- N-Y23741* Китай [ 1 ]
- Н1а2а-М128
- N-M128* Япония ( Фукуи ) [ 1 ]
- N-Y23738 Вьетнам [ 1 ]
- Н-CTS1350
- N1a2b-B523(P43)
- N1a2b-Y126204, ВЛ67/З35079, BY29083
- Н-ВЛ67/Z35079
- N1a2b1-B478 (P63) Ненцы, нганасаны, долганы, тувинцы, тоджинцы, [ 28 ] Хакасы, шорцы, эвены, алтайцы, селькупы, эвенки, монголы (сарт-калмак, [ 28 ] Торгут, [ 28 ] дерби, [ 28 ] Buryat), Yakuts, Ob-Ugrians, Chuvashes
- N1a2b1a-B168 Эвены
- N1a2b1b-B169
- N1a2b1b1-B170 Nenets , Khanty-Mansi Autonomous Okrug , Inner Mongolia
- N1a2b1b2-B175 Tuvinians , Mongols , Evenks , Yakuts , Tomsk Oblast
- N1a2b3-B525 Турция , татары , башкиры , казахи , монголы (Синьцзян-калмыки, [ 28 ] Монгольский Торгут [ 28 ] ), Словакия , Болгария , [ 1 ] украинцы , белорусы , русские
- N-B525* Афганистан , арабский
- N-BY173686 Россия ( Воронежская область [ 1 ] )
- N-VL83 Турция [ 1 ]
- N-Y136502 Россия ( Татарстан [ 1 ] ), Болгария ( Добрич [ 1 ] )
- N-Y37153
- N-Y37153* Россия ( Татарстан [ 1 ] )
- N-BY29131
- N-BY29131* Turkey ( Konya [ 1 ] ), Словакия ( Жилинский край [ 1 ] )
- N-BY30476 Россия ( Татарстан , [ 1 ] Пензенская область [ 1 ] )
- N1a2b1-B478 (P63) Ненцы, нганасаны, долганы, тувинцы, тоджинцы, [ 28 ] Хакасы, шорцы, эвены, алтайцы, селькупы, эвенки, монголы (сарт-калмак, [ 28 ] Торгут, [ 28 ] дерби, [ 28 ] Buryat), Yakuts, Ob-Ugrians, Chuvashes
- N1a2b2-FGC10872/Y3195
- N1a2b2a-FGC10847/Y3185 (L1419) Вепсы , марийцы , русские ( Архангельская область ), коми , Пермский край , Республика Коми , обские угры , чуваши , татары , башкиры , карелы , Западно-Финляндская губерния , тувинцы , буряты , хакасы , нганасаны , Азиатские эскимосы
- N1a2b2b-Y23786
- N1a2b2b* Оставайся
- N1a2b2b1-B528/Y24384 Удмурты , коми , ханты , татары , азиатские эскимосы, Кировская область , Пермский край , остров Медный
- Н-ВЛ67/Z35079
- N1a2b-Y126204, ВЛ67/З35079, BY29083
- Н-F1101
- Н-З1956
- N1b-F2905
- N1b1-CTS582 Китай (особенно Цзянсу, Юньнань, Хэбэй, Шаньдун, Шаньси, Хэйлунцзян [ 75 ] )
- N1b1a-Y6374/Z8029
- N1b1a1-CTS7324 Пекин [ 1 ]
- N1b1a2-L727
- N1b1a2*-L727 Пекин [ 1 ]
- N1b1a2a-L732 Беларусь [ 59 ]
- N1b1a2a1-F839 Китай (особенно Цзянсу, [ 75 ] Шаньдун, [ 75 ] Чжэцзян, [ 75 ] Шанхай, [ 1 ] [ 75 ] Хэнань [ 75 ] )
- N1b1a2a2-Y15972
- N1b1b-Y23789/CTS4309 Ирак [ 59 ]
- N1b1a-Y6374/Z8029
- N1b2-M1819/N-M1897/CTS12473/F1173 Китай, [ 59 ] Российская Федерация [ 59 ]
- N-M1897* древняя ДНК из Пинлянтай , Хэнань (ок. 4063 [95% ДИ 3974 <-> 4151] лет назад) [ 1 ]
- Н-М1845
- N-M1845* Китай (Сычуань, [ 1 ] Шанхай [ 1 ] )
- Н-М1928
- N-M1928* Сычуань ( Хань ) [ 1 ]
- N-Y125475
- N-Y63516 Китай ( Чунцин [ 1 ] ), Корея ( провинция Южный Пхеньян , [ 1 ] Провинция Южный Чхунчхон [ 1 ] )
- N-Y193396 Китай ( Нинся [ 1 ] ), Сингапур [ 1 ]
- N-CTS4714
- N1b1-CTS582 Китай (особенно Цзянсу, Юньнань, Хэбэй, Шаньдун, Шаньси, Хэйлунцзян [ 75 ] )
- N1a-L729
- N2-Y6503
- N2-Y6503* Республика Алтай [ 1 ]
- N2a-P189.2
- N2a1-Y6516
- N2a1-Y6516*
- N2a1a-Y7310
- N2a1a-Y7310* Румыния ( венгр из Сучавы ) [ 1 ]
- N2a1a1-Y7313
- N2a1a1-Y7313*
- N2a1a1a-BY35494 Россия ( Ростовская область ) [ 1 ]
- N2a1a1b-FGC28435
- N2a1a1b-FGC28435* Турция (Стамбул), Сербия, Черногория (Плужине), Босния и Герцеговина ( Республика Сербская ), Хорватия [ 1 ]
- N2a1a1b1-FGC28483 Сербия [ 1 ]
- N2a2-Y111068
- N2a2a-FT352925 Франция ( Приморская Шаранта [ 1 ] ), Турция [ 1 ]
- N2a2b-Y101945 Великобритания ( Девон [ 1 ] ), Россия ( Московская область [ 1 ] )
- N2a1-Y6516
- N1-Z4762/CTS11499/L735/M2291
- Н-М231/Пейдж91, М232/М2188
История филогенетической номенклатуры
[ редактировать ]До 2002 года в научной литературе существовало как минимум семь систем наименования филогенетического дерева Y-хромосомы. Это привело к значительной путанице. В 2002 году основные исследовательские группы объединились и сформировали Консорциум Y-хромосомы (YCC). Они опубликовали совместный документ, в котором было создано единое новое дерево, которое все согласились использовать. Позже группа гражданских ученых, интересующихся популяционной генетикой и генетической генеалогией, сформировала рабочую группу для создания любительского дерева, стремясь быть прежде всего своевременным. В таблице ниже собраны все эти работы, относящиеся к знаковому дереву YCC 2002 года. Это позволяет исследователю, просматривающему старую опубликованную литературу, быстро перемещаться между номенклатурами.
| YCC 2002/2008 (сокращенно) | (а) | (б) | (с) | (г) | (е) | (г) | (или) | YCC 2002 (от руки) | YCC 2005 (от руки) | YCC 2008 (от руки) | YCC 2010r (от руки) | ИСОГГ 2006 г. | ИСОГГ 2007 | ИСОГГ 2008 | ИСОГГ 2009 | ИСОГГ 2010 | ИЗОГГ 2011 | ИЗОГГ 2012 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Н-LLY22g | 12 | VIII | 1U | 25 | ЕС16 | Н5 | Ф | Н* | Н | N1 | N1 | - | - | - | - | - | - | - |
| Н-М128 | 12 | VIII | 1U | 25 | ЕС16 | Н5 | Ф | N1 | N1 | Н1а | Н1а | - | - | - | - | - | - | - |
| Н-П63 | 12 | VIII | 1U | 25 | ЕС16 | Н5 | Ф | Н2 | Н2а | N1b1 | N1b1 | - | - | - | - | - | - | - |
| Н-ТАТ | 12 | VIII | 1я | 26 | ЕС13 | Н5 | Ф | N3* | N3 | N1c | N1c | - | - | - | - | - | - | - |
| Н-М178 | 16 | VIII | 1я | 26 | ЕС14 | Н5 | Ф | Н3а* | М178 | N1c1 | N1c1 | - | - | - | - | - | - | - |
| Н-П21 | 16 | VIII | 1я | 26 | ЕС14 | Н5 | Ф | N3a1 | N3a1 | N1c1a | N1c1a | - | - | - | - | - | - | - |
Источники Следующие исследовательские группы в соответствии с их публикациями были представлены в создании Дерева YCC.
Недостоверные мутации (SNP и UEP)
Делеция b2/b3 в регионе AZFc Y-хромосомы, по-видимому, произошла независимо, по крайней мере, в четырех разных случаях. Следовательно, это удаление не следует воспринимать как уникальный полиморфизм событий, определяющий эту ветвь дерева Y-хромосомы ( ISOGG 2012 ).
Ссылки на концепции генетики
[ редактировать ]- генетическая генеалогия
- Генетическая история Европы
- Гаплогруппа
- Гаплотип
- Гаплогруппа ДНК Y-хромосомы человека
- молекулярная филогения
- Парагруппа
- Субклад
- Гаплогруппы Y-хромосомы в популяциях мира
- Гаплогруппы Y-ДНК по этническим группам
- Гаплогруппы Y-ДНК в популяциях Восточной и Юго-Восточной Азии
- Гаплогруппы Y-ДНК в населении Европы
Субклады Y-ДНК N
[ редактировать ]- Н-М231
Основополагающее дерево Y-ДНК
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также и аль являюсь а к ап ак с как в В из хорошо топор является тот нет бб до нашей эры др. быть парень бг чб с минет БК с бм млрд быть б.п. БК бр бс БТ этот бв б бх к бз что КБ копия компакт-диск Этот см. cg ч Там СиДжей ск кл см CN со КП cq кр CS КТ с резюме cw сх сай чешский и БД округ Колумбия дд из дф дг д Из диджей дк дл дм дн делать дп дк доктор дс дт из дв ДВ дх ты дз из Эб ЕС Эд да если например ага нет ej я он в в там эп. экв. является является И Евросоюз этот Вон тот бывший эй нет но ФБ ФК ФД фе фф фг фх быть фджей хз в фм фн фо фп fq фр. фс футы это было фв фу валютный курс мой фз га ГБ ГК Боже ге подруга gg хх ги джей ГК гл гм гн идти врач gq гр гс гт к гв гв gx гы гз ха чб хк HD он хф рт.ст. хх привет дрочка хк гл хм хн к л.с. штаб-квартира час хз хт ху что как хх он хз это а IC идентификатор т.е. если ig ih ii ij я il в в этот IP iq и является это я iv я ix запах iz и джб Джей Си Джей Ди является JF джг джх из Джей-Джей джк джл Джейм джин потому что Япония jq младший js ДжТ верно СП jw jx ты джз тот КБ кс кд тот кф кг х к кДж ок в км знать является КП кг ДКК кс КТ к кв кВт кх этот КЗ тот фунт ЖК лд тот лф LG левый что жж lk я буду лм Ин это лп лк лр лс лт ему лв лв лк ли лз и мб MC Мэриленд мне мф мг мх мне ЭмДжей мк мл мм минута для депутат кв.м. Мистер РС тонна в МВ МВт мх мой мз уже обратите внимание NC nd ne нф из нет в Нью-Джерси такой как Нидерланды нм пп нет например нк Нет. нс нт нет нв теперь нх тот Новая Зеландия из ob ок из ты из и ой привет ой хорошо ол о на и на белый или ты ot или ов ой бык ООО унция хорошо пб ПК ПД на пф стр. тел. пи пиджей ПК пожалуйста вечер пн po пп ПК пиар пс пт мог пв пш пикселей пи пз ка qb контроль качества qd ЧТО qf qg qh ци qj кк ql кв.м. qn qo qp qq qr вопросы qt что кв qw qx куй девочка дни РБ RC р-д ре рф рг резус ри РДжей рк рл РМ р-н ро рп rq рр РС рт ру фургон RW прием рай рз на сб СК SD с НФ сг ш и сж ск сл см зп так сп кв. сэр SS ул. являются св SW сх и нет лицом к лицу туберкулез ТК ТД тот ТС тг й из тиджей ТС ТЛ тм ТН к город tq тр тс тт что телевизор дважды Техас ty тс делать уб Калифорнийский университет вне уэ уф и Эм-м-м пользовательский интерфейс удж Великобритания ул один и друзья вверх UQ ты YПолная гаплогруппа YTree v6.05.11 от 25 сентября 2018 г.
- ^ Перейти обратно: а б с Позник Г.Д., Сюэ Ю., Мендес Ф.Л., Виллемс Т.Ф., Массайя А., Уилсон Сейрес М.А. и др. (июнь 2016 г.). «Периодические всплески демографии мужского пола, выявленные на основе 1244 последовательностей Y-хромосомы по всему миру» . Природная генетика . 48 (6): 593–599. дои : 10.1038/ng.3559 . ПМЦ 4884158 . ПМИД 27111036 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также и аль являюсь а к ап ак с как в В из хорошо топор является тот нет бб до нашей эры др. быть парень бг чб с минет БК с бм млрд быть б.п. БК бр бс БТ этот бв б бх к бз Илумяэ и др. 2016 год
- ^ Перейти обратно: а б с д и ISOGG, 2016, Гаплогруппа N Y-ДНК и ее субклады – 2016, 22 августа 2016 г.).
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с Роотси, Сиири [на эстонском языке] ; Животовский Лев А; Бальдович, Мариан; Кайзер, Манфред; и др. (06 декабря 2006 г.). «Северный маршрут гаплогруппы N Y-хромосомы против часовой стрелки из Юго-Восточной Азии в Европу» (PDF) . Европейский журнал генетики человека . 15 (4): 204–211. дои : 10.1038/sj.ejhg.5201748 . ПМИД 17149388 . S2CID 19265287 .
- ^ Перейти обратно: а б с д и ж г час Карафет Т.М., Осипова Л.П., Савина О.В., Холлмарк Б, Хаммер М.Ф. (ноябрь 2018). «Сибирское генетическое разнообразие раскрывает сложное происхождение самодийскоязычного населения» . Американский журнал биологии человека . 30 (6): e23194. дои : 10.1002/ajhb.23194 . ПМИД 30408262 .
- ^ Федорова С.А., Рейдла М, Метспалу Е, Метспалу М, Роотси С, Тамбетс К и др. (июнь 2013 г.). «Аутосомно-однородительские портреты коренного населения Саха (Якутии): значение для заселения Северо-Восточной Евразии» . Эволюционная биология BMC . 13 (13): 127. Бибкод : 2013BMCEE..13..127F . дои : 10.1186/1471-2148-13-127 . ПМЦ 3695835 . ПМИД 23782551 .
- ^ Перейти обратно: а б Дагган А.Т., Уиттен М., Вибе В., Кроуфорд М., Баттоф А., Спицын В. и др. (2013). «Исследование предыстории тунгусских народов Сибири и Амуро-Уссурийского региона с использованием полных последовательностей генома мтДНК и маркеров Y-хромосомы» . ПЛОС ОДИН . 8 (12): е83570. Бибкод : 2013PLoSO...883570D . дои : 10.1371/journal.pone.0083570 . ПМЦ 3861515 . ПМИД 24349531 .
- ^ Перейти обратно: а б с Харьков, В.Н.; Хамина, КВ; Медведева, О.Ф.; Штыгашева О.В.; Степанов, В.А. (2011). «Генетическое разнообразие хакасского генофонда: субэтническая дифференциация и структура гаплогрупп Y-хромосомы». Молекулярная биология . 45 (3): 404–416. дои : 10.1134/S0026893311020117 . ISSN 0026-8933 . S2CID 37140960 .
- ^ A. T. Agdzhoyan, E. V.Balanovska, A. D. Padyukova, D. O. Dolinina, M. A. Kuznetsova, V. V. Zaporozhchenko, R. A. Skhalyaho, S. M. Koshel, M. K. Zhabagin, Y. M. Yusupov, Kh. Kh. Mustafin, M. V. Ulyanova, Z. A. Tychinskih, M. B. Lavryashina, and O. P. Balanovsky (2016), "The Gene Pool of Siberian Tatars: Five Ways of Origin for the Five Subethnic Groups." МОЛЕКУЛЯРНАЯ БИОЛОГИЯ , Volume 50, No. 6, pp. 978–991. DOI: 10.7868/S0026898416060021
- ^ Перейти обратно: а б с д и ж , "СТРУКТУРА И ФИЛОГЕОГРАФИЯ ГЕНОФОНДА КОРЕННОГО НАСЕЛЕНИЯ СИБИРИ ПО МАРКЕРАМ Y-ХРОМОСОМЫ," Genetika 03.02.07 and "АВТОРЕФЕРАТ диссертации на соискание учёной степени доктора биологических наук, Tomsk 2012
- ^ Перейти обратно: а б с д Пименов В.Н., Комас Д., Пало Ю., Вершубский Г., Козлов А., Саянтила А. (октябрь 2008 г.). «Северо-западно-сибирские ханты и манси на стыке генофондов Западной и Восточной Евразии по данным однородительских маркеров» . Европейский журнал генетики человека . 16 (10): 1254–1264. дои : 10.1038/ejhg.2008.101 . ПМИД 18506205 . S2CID 19488203 .
- ^ Перейти обратно: а б с д и ж г час Лаппалайнен и др. 2008 год
- ^ Перейти обратно: а б с д Тамбетс К., Юнусбаев Б., Худжашов Г., Илумяэ А.М., Роотси С., Хонкола Т. и др. (сентябрь 2018 г.). «Гены обнаруживают следы общей недавней демографической истории для большинства уральскоязычного населения» . Геномная биология . 19 (1): 139. дои : 10.1186/s13059-018-1522-1 . ПМК 6151024 . ПМИД 30241495 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В Сюэ и др. 2006 г.
- ^ Перейти обратно: а б Богунов Ю.В.; Мальцева О.В.; Богунова А.А.; Балановская, Е.В. (2015). «НАНААЙСКИЙ КЛАН САМАР: СТРУКТУРА ГЕНОФОНДА НА ОСНОВЕ МАРКЕРОВ Y-ХРОМОСОМЫ1». Археология, этнология и антропология Евразии . 43 (2): 146–152. дои : 10.1016/j.aeae.2015.09.015 .
- ^ Плисс Л., Тимша Л., Роотси С., Тамбетс К., Пелнена И., Золе Е. и др. (ноябрь 2015 г.). «Y-хромосомные линии латышей в контексте генетической изменчивости Восточно-Балтийского региона». Анналы генетики человека . 79 (6): 418–430. дои : 10.1111/ahg.12130 . ПМИД 26411886 . S2CID 13050610 .
- ^ Перейти обратно: а б Деренко М., Малярчук Б., Денисова Г.А., Возняк М., Дамбуева И., Доржу С. и др. (январь 2006 г.). «Контрастные закономерности изменчивости Y-хромосомы в южносибирских популяциях Байкальского и Алтае-Саянского регионов». Генетика человека . 118 (5): 591–604. дои : 10.1007/s00439-005-0076-y . ПМИД 16261343 . S2CID 23011845 .
- ^ Перейти обратно: а б с д и ж г час я дж Ким Ш., Ким К.С., Шин DJ, Джин Х.Дж., Квак К.Д., Хан М.С. и др. (апрель 2011 г.). «Высокие частоты линий гаплогруппы Y-хромосомы O2b-SRY465 в Корее: генетический взгляд на заселение Кореи» . Исследовательская генетика . 2 (1): 10. дои : 10.1186/2041-2223-2-10 . ПМК 3087676 . ПМИД 21463511 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с Хаммер и др. 2006 г.
- ^ Kharkov, V. N.; Khamina, K. V.; Medvedeva, O. F.; Simonova, K. V.; Eremina, E. R.; Stepanov, V. A. (2014). "Gene pool of Buryats: Clinal variability and territorial subdivision based on data of Y-chromosome markers". Russian Journal of Genetics . 50 (2): 180–190. doi : 10.1134/S1022795413110082 . ISSN 1022-7954 . S2CID 15595963 .
- ^ Перейти обратно: а б Харьков и др. 2007 год
- ^ Перейти обратно: а б Дулик М.С., Жаданов С.И., Осипова Л.П., Аскапули А., Гау Л., Гоккумен О. и др. (февраль 2012 г.). «Вариации митохондриальной ДНК и Y-хромосомы свидетельствуют о недавнем общем происхождении коренных американцев и коренных алтайцев» . Американский журнал генетики человека . 90 (2): 229–246. дои : 10.1016/j.ajhg.2011.12.014 . ПМК 3276666 . ПМИД 22281367 .
- ^ Малярчук и др. 2004 г.
- ^ Перейти обратно: а б с д и ж Чжун Х., Ши Х., Ци С.Б., Дуань З.Ю., Тан П.П., Цзинь Л. и др. (январь 2011 г.). «Расширенное исследование Y-хромосомы позволяет предположить, что современные люди мигрировали в Восточную Азию северным путем после ледникового периода» . Молекулярная биология и эволюция . 28 (1): 717–727. дои : 10.1093/molbev/msq247 . ПМИД 20837606 .
- ^ Перейти обратно: а б Като и др. 2005 г.
- ^ Перейти обратно: а б Ди Кристофаро Дж., Пеннарун Э., Мазьер С., Майрес Н.М., Лин А.А., Темори С.А. и др. (2013). «Афганский Гиндукуш: место, где сходятся потоки генов Евразийского субконтинента» . ПЛОС ОДИН . 8 (10): е76748. Бибкод : 2013PLoSO...876748D . дои : 10.1371/journal.pone.0076748 . ПМЦ 3799995 . ПМИД 24204668 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п Балинова Н., Пост Х., Кушниаревич А., Флорес Р., Кармин М., Саакян Х. и др. (сентябрь 2019 г.). «Y-хромосомный анализ родовой структуры калмыков, единственного европейского монгольского народа, и их родство с ойратами-монголами Внутренней Азии» . Европейский журнал генетики человека . 27 (9): 1466–1474. дои : 10.1038/s41431-019-0399-0 . ПМК 6777519 . ПМИД 30976109 .
- ^ Перейти обратно: а б Малярчук Б, Деренко М, Денисова Г, Хойт С, Возняк М, Гжибовский Т, Захаров И (декабрь 2013 г.). «Y-хромосомное разнообразие у калмыков на этническом и племенном уровне» . Журнал генетики человека . 58 (12): 804–811. дои : 10.1038/jhg.2013.108 . ПМИД 24132124 .
- ^ Чжан X, Хэ Г, Ли В, Ван Ю, Ли Х, Чэнь Ю и др. (2021). «Геномный взгляд на историю смешения населения тунгусоязычных маньчжуров на северо-востоке Китая» . Границы генетики . 12 : 754492. doi : 10.3389/fgene.2021.754492 . ПМЦ 8515022 . ПМИД 34659368 .
- ^ Перейти обратно: а б с Мирабал С., Регейро М., Каденас А.М., Кавалли-Сфорца Л.Л., Андерхилл П.А., Вербенко Д.А. и др. (октябрь 2009 г.). «Распространение Y-хромосомы в геолингвистическом ландшафте северо-запада России» . Европейский журнал генетики человека . 17 (10): 1260–1273. дои : 10.1038/ejhg.2009.6 . ПМЦ 2986641 . ПМИД 19259129 .
- ^ Ван XQ, Ван CC, Дэн QY, Ли Х (февраль 2013 г.). «[Генетический анализ полиморфизма Y-хромосомы и митохондриальной ДНК этнической группы мулам в Гуанси, Китай]» . И Чуань = Эредитас (по-китайски). 35 (2): 168–74. дои : 10.3724/sp.j.1005.2013.00168 . ПМИД 23448929 .
- ^ Перейти обратно: а б Лаппалайнен Т., Ханнелиус У., Салмела Э., фон Дёбельн У., Линдгрен К.М., Хуопонен К. и др. (январь 2009 г.). «Структура населения в современной Швеции - анализ Y-хромосомной и митохондриальной ДНК». Анналы генетики человека . 73 (1): 61–73. дои : 10.1111/j.1469-1809.2008.00487.x . ПМИД 19040656 . S2CID 205598345 .
- ^ Перейти обратно: а б Карлссон А.О., Валлерстрем Т., Гётерстрем А., Холмлунд Г. (август 2006 г.). «Разнообразие Y-хромосомы в Швеции – долгосрочная перспектива» . Европейский журнал генетики человека . 14 (8): 963–970. дои : 10.1038/sj.ejhg.5201651 . ПМИД 16724001 . S2CID 23227271 .
- ^ Сон Сон Чхве, Кён Хва Пак, Да Ын Нам, Тэ Хун Кан, Ки Ва Чунг и др. , «Гаплогруппировка Y-хромосомы для азиатов с использованием целевого секвенирования Y-SNP».
- ^ Сунгвон Чон, Ёнджун Бхак, Ёнсон Чхве и др. , «Корейский проект генома: 1094 личных корейских генома с клинической информацией». Достижения науки 2020; 6: eaaz7835.
- ^ Сун Хи Ким; Бён Вон Чон; Чону Юнг; Брайан М. Кемп; и др. (июль 2005 г.). «Предварительное исследование происхождения корейцев на основе вариации Y-STR». Международный журнал юридической медицины . 32 (4): 353–359. дои : 10.1007/s13258-010-0030-9 . S2CID 7277981 .
- ^ К.С. Джонг; Х.Дж. Шин; С. Дж. Ли; Х.С. Ким; Дж. Я. Ким; М.С. Хан; Ю. Х. Ли; КВ Парк; Б.В. Чун (2018). «Генетические характеристики гаплотипов коротких тандемных повторов Y-хромосомы из образцов окурков, которые предположительно курили северокорейские мужчины». Международный журнал юридической медицины . 40 (8): 819–824. дои : 10.1007/s13258-018-0701-5 . ПМИД 30047114 . S2CID 256072306 .
- ^ Перейти обратно: а б Пак MJ, Ли ХИ, Ян В.И., Шин К.Дж. (июль 2012 г.). «Понимание вариаций Y-хромосомы в Корее - актуальность комбинированного анализа гаплогрупп и гаплотипов». Международный журнал юридической медицины . 126 (4): 589–599. дои : 10.1007/s00414-012-0703-9 . ПМИД 22569803 . S2CID 27644576 .
- ^ Перейти обратно: а б Балановская Е.В., Богунов Ю.В., Каменщикова Е.Н., Балаганская О.А., Агджоян А.Т., Богунова А.А. и др. (октябрь 2018 г.). «Демографические и генетические портреты населения ульчей». Российский генетический журнал . 54 (10): 1245–1253. дои : 10.1134/S1022795418100046 . S2CID 53085396 .
- ^ Ци X, Цуй С, Пэн Y, Чжан X, Ян Z, Чжун Х и др. (август 2013 г.). «Генетические свидетельства палеолитической колонизации и неолитической экспансии современного человека на Тибетское плато» . Молекулярная биология и эволюция . 30 (8): 1761–1778. дои : 10.1093/molbev/mst093 . ПМИД 23682168 .
- ^ Аширбеков Е.Е., Ботбаев Д.М., Белкожаев А.М., Абайлдаев А.О., Неупокоева А.С., Мухатаев Ж.Е. и др. (2017). «Распространение гаплогрупп Y-хромосомы казахов Южно-Казахстанской, Жамбылской и Алматинской областей». Доклады Национальной академии наук Республики Казахстан . 6 (316): 85–95.
- ^ Брунелли А., Кампуансай Дж., Сейлстад М., Ломтайсонг К., Кангванпонг Д., Гиротто С., Кутанан В. (2017). «Y-хромосомные данные о происхождении жителей северного Таиланда» . ПЛОС ОДИН . 12 (7): e0181935. Бибкод : 2017PLoSO..1281935B . дои : 10.1371/journal.pone.0181935 . ПМК 5524406 . ПМИД 28742125 .
- ^ Шуху Л.И., Илихаму Н.И., Баке Р.А., Бупатима А.Б., Матюсуп Д.О. (март 2018 г.). «Исследование генетического разнообразия трех изолированных популяций в Синьцзяне с использованием Y-SNP». Acta Anthropologica Sinica . 37 (1): 146–156.
- ^ Ян Л. (2011). Западном Китае Генетическая смесь популяций в (докторская диссертация) (на китайском языке: Университет Фудань, стр. 1–84).
- ^ Перейти обратно: а б Вэнь С.К., Ду ПХ, Сунь С., Цуй В., Сюй Ю.Р., Мэн Х.Л. и др. (март 2022 г.). «Двойное происхождение северо-западных китайских кыргызов: смесь сибирских Y-хромосом бронзового века и средневековых нируун-монгольских Y-хромосом». Журнал генетики человека . 67 (3): 175–180. дои : 10.1038/s10038-021-00979-x . ПМИД 34531527 . S2CID 237546416 .
- ^ Го Ю, Ся З, Цуй В, Чэнь С, Цзинь Икс, Чжу Б (май 2020 г.). «Совместный генетический анализ молекулярных маркеров митохондрий и Y-хромосомы для населения Северо-Западного Китая» . Гены . 11 (5): 564. doi : 10.3390/genes11050564 . ПМК 7290686 . ПМИД 32443545 .
- ^ Нонака, я; Минагучи, К; Такезаки, Н. (июль 2007 г.). «Бинарные гаплогруппы Y-хромосомы в японском населении и их связь с 16 полиморфизмами Y-STR». Анналы генетики человека . 71 (4): 480–495. дои : 10.1111/j.1469-1809.2006.00343.x . hdl : 10130/491 . ПМИД 17274803 . S2CID 1041367 .
- ^ Хараяма, Юта; Камей, Саяко; Сато, Норико; Хаяси, Токутаро; и др. (январь 2014 г.). «Анализ гаплогрупп Y-хромосомы у японского населения с использованием коротких ампликонов и его применение в судебно-медицинской экспертизе» (PDF) . Юридическая медицина . 16 (1): 20–25. doi : 10.1016/j.legalmed.2013.10.005 . hdl : 10091/17954 . ПМИД 24262653 . S2CID 19534157 .
- ^ Отиай, Эрико; Минагучи, Киёси; Намбияр, Пхрабхакаран; Какимото, Ю; и др. (сентябрь 2016 г.). «Оценка гаплогруппировки SNP Y-хромосомы в панели идентификации HID-Ion AmpliSeq™». Юридическая медицина . 22 : 58–61. doi : 10.1016/j.legalmed.2016.08.001 . ПМИД 27591541 . S2CID 43668117 .
- ^ Перейти обратно: а б Андерхилл и др. 2000 .
- ^ Перейти обратно: а б с Ши Х, Ци Х, Чжун Х, Пэн Ю, Чжан Х, Ма РЗ, Су Б (2013). «Генетические доказательства восточноазиатского происхождения и палеолитической миграции на север гаплогруппы N Y-хромосомы» . ПЛОС ОДИН . 8 (6): е66102. Бибкод : 2013PLoSO...866102S . дои : 10.1371/journal.pone.0066102 . ПМЦ 3688714 . ПМИД 23840409 .
- ^ Перейти обратно: а б с Кармин М., Сааг Л., Висенте М., Уилсон Сейрес М.А., Ярве М., Талас У.Г. и др. (апрель 2015 г.). «Недавнее ограничение разнообразия Y-хромосомы совпадает с глобальными изменениями в культуре» . Геномные исследования . 25 (4): 459–466. дои : 10.1101/гр.186684.114 . ПМЦ 4381518 . ПМИД 25770088 .
- ^ Кьярони, Андерхилл и Кавалли-Сфорца, 2009 г.
- ^ Перейти обратно: а б Ху и др. 2015 год
- ^ https://www.23mofang.com/ancestry/ytree/N-F2930/detail
- ^ Карафет и др. 2010 год
- ^ Перейти обратно: а б де Баррос Дамгаард П., Мартиниано Р., Камм Дж., Морено-Майар Дж.В., Кроонен Г., Пейро М. и др. (июнь 2018 г.). «Первые пастухи и влияние степной экспансии раннего бронзового века в Азию» . Наука . 360 (6396): eaar7711. дои : 10.1126/science.aar7711 . ПМК 6748862 . ПМИД 29743352 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также и аль являюсь а к ап ак Дерево гаплогруппы Y-ДНК N-M231 в Family Tree DNA Discover
- ^ "Haplotree.info - Ancientdna.info. Карта на основе All Ancient DNA v. 2.07.26" .
- ^ Перейти обратно: а б с Карафет и др. 2001 г.
- ^ Перейти обратно: а б Вен и др. 2004 г.
- ^ Перейти обратно: а б Цай и др. 2011 год
- ^ Ким и др. 2007 год
- ^ Гайден и др. 2007 год
- ^ Перейти обратно: а б Цуй, Иньцю; Ли, Хунцзе; Нин, Чао; Чжан, Е; и др. (30 сентября 2013 г.). «Анализ Y-хромосомы доисторических человеческих популяций в долине реки Западный Ляо, Северо-Восточный Китай» . Эволюционная биология BMC . 13 (1): 216. Бибкод : 2013BMCEE..13..216C . дои : 10.1186/1471-2148-13-216 . ПМЦ 3850526 . ПМИД 24079706 . S2CID 13544780 .
- ^ «N-Z1956 YДерево» .
- ^ Перейти обратно: а б Сирак и др. 2020 .
- ^ Пакендорф и др. 2002 г.
- ^ Крубези и др. 2010 год
- ^ Перейти обратно: а б Деренко и др. 2007 год
- ^ Дулик М.К., Оуингс А.С., Гайески Дж.Б., Вилар М.Г., Андре А., Ленни С. и др. (май 2012 г.). «Анализ Y-хромосомы выявляет генетическое расхождение и появление новых коренных родословных в атапасских и эскимосских популяциях» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (22): 8471–8476. Бибкод : 2012PNAS..109.8471D . дои : 10.1073/pnas.1118760109 . ПМК 3365193 . ПМИД 22586127 .
- ^ Кылинч Г.М., Кашуба Н., Коптекин Д., Бергфельдт Н., Донерташ Х.М., Родригес-Варела Р. и др. (январь 2021 г.). «Динамика человеческого населения и Yersinia pestis в древней Северо-Восточной Азии» . Достижения науки . 7 (2): eabc4587. Бибкод : 2021SciA....7.4587K . дои : 10.1126/sciadv.abc4587 . ПМЦ 7787494 . ПМИД 33523963 .
- ^ «N-L708 YДерево» . Проверено 28 ноября 2022 г.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также и аль являюсь а к ап ак с как в В из хорошо топор является тот нет бб до нашей эры др. быть парень бг чб с Филогенетическое древо гаплогруппы N на 23 мофанге
- ^ Чэнь Дж, Хэ Г, Жэнь З, Ван Ц, Лю Ю, Чжан Х, Ян М, Чжан Х, Цзи Дж, Чжао Дж, Го Дж, Чжу К, Ян Икс, Ван Р, Ма Х, Ван CC и Хуан J (2021), «Геномный взгляд на историю примеси монголо- и тунгусоязычного населения Юго-Западной Восточной Азии». Передний. Жене. 12:685285. дои: 10.3389/fgene.2021.685285
- ^ «Zhōngguó manzú qúntì fùxì dan bei qún ji minzú xiě tŏng gòuchéng qíngkuàng jiânjiè» Краткое введение в отцовскую гаплогруппу и состав этнического происхождения маньчжуров в Китае. [Краткое введение в отцовскую гаплогруппу и состав этнического происхождения маньчжурского народа в Китае]. 23мофан (на китайском языке). 21 января 2021 г. Проверено 11 марта 2024 г.
- ^ Ху и др. 2015 .
- ^ Ма, Пэнчэн; Ян, Сюань; Ян, Ши; Ли, Чуньсян; и др. (декабрь 2021 г.). «Древняя Y-ДНК с реконструированной филогенией дает представление о демографической истории отцовской гаплогруппы N1a2-F1360» . Журнал генетики и геномики . 48 (12): 1130–1133. дои : 10.1016/j.jgg.2021.07.018 . ПМИД 34425243 . S2CID 237281094 .
- ^ Перейти обратно: а б с Мао, Сяовэй; Чжан, Хуцай; Цяо, Шию; Лю, Ичэнь; и др. (июнь 2021 г.). «Глубокая история населения северной части Восточной Азии от позднего плейстоцена до голоцена» . Клетка . 184 (12): 3256–3266.e13. дои : 10.1016/j.cell.2021.04.040 . ISSN 0092-8674 . ПМИД 34048699 .
- ^ «Гаплогруппа Theytree YTree (N-Y24206)» . Проверено 10 марта 2024 г.
- ^ «Гаплогруппа Theytree YTree (N-F710)» . Проверено 10 марта 2024 г.
- ^ Лю, Цзюнсен; Цзэн, Вэнь; Сунь, Бо; Мао, Сяовэй; и др. (29 мая 2021 г.). «Материнская генетическая структура в древнем Шаньдуне между 9500 и 1800 годами назад» . Научный вестник . 66 (11): 1129–1135. Бибкод : 2021SciBu..66.1129L . дои : 10.1016/j.scib.2021.01.029 . ПМИД 36654346 .
- ^ Ван, Хунру; Ян, Мелинда А.; Вангду, Шарган; Лу, Хунлян; и др. (15 марта 2023 г.). «Генетическая история человека на Тибетском нагорье за последние 5100 лет» . Достижения науки . 9 (11): eadd5582. Бибкод : 2023SciA....9D5582W . дои : 10.1126/sciadv.add5582 . ISSN 2375-2548 . ПМЦ 10022901 . ПМИД 36930720 .
- ^ «Гаплогруппа Theytree YTree (N-CTS2959)» . Проверено 10 марта 2024 г.
- ^ Карафет Татьяна М.; Осипова Людмила П; Губина Марина А; Посух, Ольга Л; и др. (декабрь 2002 г.). «Высокий уровень дифференциации Y-хромосомы среди коренного сибирского населения и генетическая подпись бореального образа жизни охотников-собирателей» . Биология человека . 74 (6): 761–789. дои : 10.1353/hub.2003.0006 . ПМИД 12617488 . S2CID 9443804 .
- ^ Тамбетс, Кристина; Роотси, Сирия [на эстонском языке] ; Кивисилд, Томас ; Помогите, Ад; и др. (апрель 2004 г.). «Западные и восточные корни саамов – история генетических «выбросов», рассказанная митохондриальной ДНК и Y-хромосомами» . Американский журнал генетики человека . 74 (4): 661–682. дои : 10.1086/383203 . ПМК 1181943 . ПМИД 15024688 . S2CID 18395985 .
- ^ Харьков, ВН; Колесников Н.А.; Валихова Л.В.; Зарубин А.А.; и др. (март 2023 г.). «Связь генофонда хантов с народами Западной Сибири, Приуралья и Алтае-Саянского региона по данным о полиморфизме аутосомного локуса и Y-хромосомы » Вавиловский журнал генетики и селекции . 27 (1): 46–54. дои : 10.18699/VJGB-23-07 . eISSN 2500-3259 . ПМЦ 10009483 . ПМИД 36923476 . S2CID 257478780 .
- ^ «Подробности о гаплогруппе N-F2930» .
- ^ Перейти обратно: а б N-P189.2 YПолный, 2018 г. (24 июня 2018 г.).
- ^ Гаплогруппа N Y-ДНК и ее субклады - 2018 , ISOGG , 2018, (24 июня 2018 г.).
- ^ Перейти обратно: а б де Баррос Дамгаард, Питер; Марки, Нина; Расмуссен, Саймон; Пейро, Микаэль; и др. (9 мая 2018 г.). «137 древних геномов человека со всех концов Евразийских степей» . Природа . 557 (7705): 369–374. Бибкод : 2018Natur.557..369D . дои : 10.1038/s41586-018-0094-2 . hdl : 1887/3202709 . ПМИД 29743675 . S2CID 256769352 .
- ^ Гамба, Кристина; Джонс, Эппи Р.; Тисдейл, Мэтью Д.; Маклафлин, Рассел Л.; и др. (21 октября 2014 г.). «Поток и застой генома на протяжении пяти тысячелетий европейской предыстории» (PDF) . Природные коммуникации . 5 : 5257. Бибкод : 2014NatCo...5.5257G . дои : 10.1038/ncomms6257 . ПМЦ 4218962 . ПМИД 25334030 . S2CID 19587039 .
- ^ Перейти обратно: а б с д и ж Гаплогруппа N Североевразийский проект YDNA в генеалогическом древе ДНК
- ^ Липпольд, Себастьян; Сюй, Хунъян; Ко, Альберт; Ли, Минкун; и др. (24 сентября 2014 г.). «Демографические истории отца и матери человека: данные из Y-хромосомы высокого разрешения и последовательностей мтДНК» (PDF) . Исследовательская генетика . 5:13 . дои : 10.1186/2041-2223-5-13 . ПМК 4174254 . ПМИД 25254093 . S2CID 16464327 .
- ^ Ван, Лин-Сян; Лу, Ян; Чжан, Чао; Вэй, Лан-Хай; и др. (октябрь 2018 г.). «Реконструкция филогении Y-хромосомы выявляет две неолитические экспансии тибето-бирманского населения». Молекулярная генетика и геномика . 293 (5): 1293–1300. дои : 10.1007/s00438-018-1461-2 . ПМИД 29923068 . S2CID 49311699 .
- ^ Перейти обратно: а б Ли, Ивонн Ю; Чанг, Грейс Тай; Луи, Вивиан, Вайоминг; То, Ка-Фай; и др. (18 января 2017 г.). «Секвенирование экзома и генома рака носоглотки выявляет мутации, активирующие путь NF-κB» (PDF) . Природные коммуникации . 8 : 14121. Бибкод : 2017NatCo...814121L . дои : 10.1038/ncomms14121 . ПМЦ 5253631 . ПМИД 28098136 . S2CID 15387583 .
- ^ Биобанк Института медицинских исследований Кориелла.
- ^ Джоблинг и Тайлер-Смит, 2000 г.
- ^ Каладжиева и др. 2001 г.
- ^ Андерхилл и др. 2000 г.
- ^ Хаммер и др. 2001 г.
- ^ Семино и др. 2000 г.
- ^ Су и др. 1999 год
- ^ Капелли и др. 2001 г.
Библиография
[ редактировать ]- Цай, Сяоюнь; Цинь, Чжэндун; Вэнь, Бо; Сюй, Шухуа; и др. (31 августа 2011 г.). О'Рурк, Деннис (ред.). «Миграция человека через узкие места из Юго-Восточной Азии в Восточную Азию во время последнего ледникового максимума, выявленная Y-хромосомами» . ПЛОС Один . 6 (8). Генографический консорциум: e24282. Бибкод : 2011PLoSO...624282C . дои : 10.1371/journal.pone.0024282 . ПМК 3164178 . ПМИД 21904623 . S2CID 2731184 .
- Кьярони, Жак; Андерхилл, Питер А; Кавалли-Сфорца, Лука Л. (17 ноября 2009 г.). «Разнообразие Y-хромосомы, человеческая экспансия, дрейф и культурная эволюция» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (48): 20174–20179. Бибкод : 2009PNAS..10620174C . дои : 10.1073/pnas.0910803106 . ПМЦ 2787129 . ПМИД 19920170 . S2CID 32040326 .
- Крубези, Эрик; Амори, Сильвен; Кейзер, Кристина; Буаказе, Кэролайн; и др. (25 января 2010 г.). «Эволюция человека в Сибири: от замороженных тел к древней ДНК» (PDF) . Эволюционная биология BMC . 10 (1): 25. Бибкод : 2010BMCEE..10...25C . дои : 10.1186/1471-2148-10-25 . ПМК 2829035 . ПМИД 20100333 . S2CID 16326890 .
- Деренко, Мирослава; Венок, Борис; Денисова Галина; Возняк, Марцин; и др. (17 августа 2007 г.). «Распространение гаплогруппы N Y-хромосомы из южной Сибири в Европу» (PDF) . Журнал генетики человека . 52 (9): 763–770. дои : 10.1007/s10038-007-0179-5 . ПМИД 17703276 . S2CID 25599501 .
- Гайден, Тензин; Каденас, Алисия М; Регейру, Мария; Сингх, Нанда Б; и др. (4 апреля 2007 г.). «Гималаи как направленный барьер на пути потока генов» (PDF) . Американский журнал генетики человека . 80 (5): 884–894. дои : 10.1086/516757 . ПМЦ 1852741 . ПМИД 17436243 . S2CID 25312596 .
- Хаммер, Майкл Ф; Карафет Татьяна М; Пак, Хваён; Омото, Кэйичи [на японском языке] ; и др. (2006). «Двойное происхождение японцев: общая основа Y-хромосом охотников-собирателей и фермеров» (PDF) . Журнал генетики человека . 51 (1): 47–58. дои : 10.1007/s10038-005-0322-0 . ПМИД 16328082 . S2CID 6559289 .
- Илумяэ, Анне-Май; Рейдла, Маере [на эстонском языке] ; Чухряева Марина; Ярве, Мари [на эстонском языке] ; и др. (07.07.2016). «Гаплогруппа Y-хромосомы человека: нетривиальная филогеография с временным разрешением, охватывающая языковые семьи» (PDF) . Американский журнал генетики человека . 99 (1): 163–173. дои : 10.1016/j.ajhg.2016.05.025 . ПМЦ 500549 . ПМИД 27392075 . S2CID 20536258 .
- Ху, Канг; Ян, Ши; Лю, Кай; Нин, Чао; и др. (24 апреля 2015 г.). «Дихотомическая структура гаплогруппы N Y-хромосомы». arXiv : 1504.06463 [ q-bio.PE ].
- Карафет, Татьяна; Сюй, Липин; Ду, Руофу; Ван, Уильям; и др. (30 июля 2001 г.). «История отцовского населения Восточной Азии: источники, закономерности и микроэволюционные процессы» (PDF) . Американский журнал генетики человека . 69 (3): 615–628. дои : 10.1086/323299 . ПМЦ 1235490 . ПМИД 11481588 . S2CID 27785973 .
В этой статье образец «Южная Хань», полученный исследовательской группой Карафета и Хаммера, описывается как происходящий из провинции Гуандун, а образец «Северная Хань» описывается как происходящий из провинции Шэньси.
- Карафет Татьяна М; Мендес, Фернандо Л; Мейлерман, Моника Б; Андерхилл, Питер А; и др. (2 апреля 2008 г.). «Новые бинарные полиморфизмы изменяют форму и увеличивают разрешение дерева гаплогруппы Y-хромосомы человека» . Геномные исследования . 18 (5): 80–838. дои : 10.1101/гр.7172008 . ПМК 2336805 . ПМИД 18385274 . S2CID 12995903 .
- Карафет Татьяна М; Холлмарк, Брайан; Кокс, Мюррей П; Судойо, Херавати [на индонезийском языке] ; и др. (05 марта 2010 г.). «Основное разделение Восток-Запад лежит в основе стратификации Y-хромосомы в Индонезии» . Молекулярная биология и эволюция . 27 (8): 1833–1844. дои : 10.1093/molbev/msq063 . ПМИД 20207712 . S2CID 4819475 .
- Като, Тору; Мунхбат, Батмунх; Тунаи, Кеничи; Мано, Шухей; и др. (14 февраля 2005 г.). «Генетические особенности монгольских этносов, выявленные с помощью Y-хромосомного анализа». Джин . 346 : 63–70. дои : 10.1016/j.gene.2004.10.023 . ПМИД 15716011 . S2CID 28132496 .
- Харьков В.Н., Степанов В.А., Медведева О.Ф., Спиридонова М.Г., Воевода М.И., Тадинова В.Н. и др. (май 2007 г.). «Различия генофонда северных и южных Алтайцев, выведенные из данных по гаплогруппам Y-хромосомы» (PDF) . Российский генетический журнал . 43 (5): 551–562. дои : 10.1134/S1022795407050110 . ПМИД 17633562 . S2CID 566825 .
- Ким В., Ю Т.К., Ким С.Дж., Шин Дж.Д., Тайлер-Смит С., Джин Х.Дж. и др. (январь 2007 г.). Благосклонный М (ред.). «Отсутствие связи между гаплогруппами Y-хромосомы и раком простаты у корейского населения» . ПЛОС ОДИН . 2 (1): е172. Бибкод : 2007PLoSO...2..172K . дои : 10.1371/journal.pone.0000172 . ПМЦ 1766463 . ПМИД 17245448 .
- Лаппалайнен Т., Лайтинен В., Салмела Э., Андерсен П., Хуопонен К., Савонтаус М.Л., Лаэрмо П. (май 2008 г.). «Миграционные волны в регион Балтийского моря» . Анналы генетики человека . 72 (Часть 3): 337–348. дои : 10.1111/j.1469-1809.2007.00429.x . ПМИД 18294359 . S2CID 32079904 .
- Малярчук Б, Деренко М, Гжибовский Т, Лунькина А, Чарный Ю, Рычков С и др. (декабрь 2004 г.). «Дифференциация митохондриальной ДНК и Y-хромосом в российских популяциях». Биология человека . 76 (6): 877–900. дои : 10.1353/hub.2005.0021 . ПМИД 15974299 . S2CID 17385503 .
- Пакендорф Б., Морар Б., Тарская Л.А., Кайзер М., Судьял Х., Родевальд А., Стоункинг М. (февраль 2002 г.). «Y-хромосомные доказательства сильного сокращения численности мужского населения якутов». Генетика человека . 110 (2): 198–200. дои : 10.1007/s00439-001-0664-4 . ПМИД 11935328 . S2CID 30671159 .
- Сирак К., Фернандес Д., Черонет О., Харни Э., Мах М., Маллик С. и др. (март 2020 г.). «Слуховые косточки человека как альтернативный оптимальный источник древней ДНК» . Геномные исследования . 30 (3): 427–436. дои : 10.1101/гр.260141.119 . ПМК 7111520 . ПМИД 32098773 .
- Вэнь Б, Се Х, Гао С, Ли Х, Ши Х, Сун Х и др. (май 2004 г.). «Анализ генетической структуры тибето-бирманских популяций выявил примесь по признаку пола у южных тибето-бирманцев» . Американский журнал генетики человека . 74 (5): 856–865. дои : 10.1086/386292 . ПМК 1181980 . ПМИД 15042512 .
- Сюэ Ю, Зержал Т, Бао В, Чжу С, Шу Ц, Сюй Дж и др. (апрель 2006 г.). «Мужская демография в Восточной Азии: контраст между севером и югом во времени роста населения» . Генетика . 172 (4): 2431–2439. doi : 10.1534/genetics.105.054270 . ПМЦ 1456369 . ПМИД 16489223 .
Веб-сайты
[ редактировать ]- ИЗОГГ (2012). «Дерево гаплогрупп Y-ДНК 2012» .
- Макдональд, доктор юридических наук (2004). «Гаплогруппы Макдональда Y мира» (PDF) . Архивировано из оригинала (PDF) 28 июля 2004 г. Проверено 29 августа 2009 г.
Источники таблиц преобразования
[ редактировать ]- Капелли, Кристиан; Уилсон, Джеймс Ф.; Ричардс, Мартин; Штумпф, Майкл П.Х.; и др. (февраль 2001 г.). «Отцовское наследие преимущественно коренных народов австронезийскоязычных народов островной Юго-Восточной Азии и Океании» . Американский журнал генетики человека . 68 (2): 432–443. дои : 10.1086/318205 . ПМЦ 1235276 . ПМИД 11170891 .
- Хаммер, Майкл Ф.; Карафет Татьяна М.; Редд, Алан Дж.; Джарджанази, Хамди; и др. (1 июля 2001 г.). «Иерархические закономерности глобального разнообразия Y-хромосом человека» . Молекулярная биология и эволюция . 18 (7): 1189–1203. doi : 10.1093/oxfordjournals.molbev.a003906 . ПМИД 11420360 .
- Джоблинг, Марк А.; Тайлер-Смит, Крис (2000), «Новое использование новых гаплотипов», Trends in Genetics , 16 (8): 356–62, doi : 10.1016/S0168-9525(00)02057-6 , PMID 10904265
- Каладжиева, преп.; Калафель, Франческ; Джоблинг, Марк А; Ангеличева, Дора; и др. (февраль 2001 г.). «Закономерности меж- и внутригруппового генетического разнообразия у Vlax Roma, выявленные с помощью Y-хромосомы и линий митохондриальной ДНК» . Европейский журнал генетики человека . 9 (2): 97–104. дои : 10.1038/sj.ejhg.5200597 . ПМИД 11313742 .
- Семино О., Пассарино Г., Офнер П.Дж., Лин А.А. и др. (2000), «Генетическое наследие палеолитического Homo sapiens sapiens у современных европейцев: хромосомная перспектива AY», Science , 290 (5494): 1155–1159, Бибкод : 2000Sci...290.1155S , doi : 10.1126/science.290.5494. 1155 , ПМИД 11073453
- Су, Бинг; Сяо, Цзюньхуа; Андерхилл, Питер; Дека, Ранджан; и др. (декабрь 1999 г.). «Y-хромосома свидетельствует о миграции современных людей на север в Восточную Азию во время последнего ледникового периода» . Американский журнал генетики человека . 65 (6): 1718–1724. дои : 10.1086/302680 . ПМЦ 1288383 . ПМИД 10577926 .
- Андерхилл П.А., Шен П., Лин А.А., Джин Л. и др. (ноябрь 2000 г.). «Вариации последовательности Y-хромосомы и история человеческих популяций». Природная генетика . 26 (3): 358–361. дои : 10.1038/81685 . ПМИД 11062480 . S2CID 12893406 .
Дальнейшее чтение
[ редактировать ]Этот раздел нуждается в расширении . Вы можете помочь, добавив к нему . ( декабрь 2012 г. ) |
- ИЗОГГ (2006). «Дерево гаплогрупп Y-ДНК 2006» .
- ИЗОГГ (2007). «Дерево гаплогрупп Y-ДНК 2007» .
- ИЗОГГ (2008). «Дерево гаплогрупп Y-ДНК 2008» .
- ИЗОГГ (2009). «Дерево гаплогрупп Y-ДНК 2009» .
- ИЗОГГ (2010). «Дерево гаплогрупп Y-ДНК 2010» .
- ИЗОГГ (2011). «Дерево гаплогрупп Y-ДНК 2011» .
- ИЗОГГ (2018). «Гаплогруппа N Y-ДНК и ее субклады» .
- YПолный. «Полное экспериментальное YTree» .
- ИЗОГГ (2019–2020 гг.). «Гаплогруппа N Y-ДНК и ее субклады» .
Филогенетика
[ редактировать ]- ^ Делеция b2/b3 в регионе AZFc Y-хромосомы человека является характеристикой гаплотипов гаплогруппы N-M231. Однако это удаление, по-видимому, происходило независимо в четырех разных случаях. Поэтому эту делецию не следует рассматривать как уникальный полиморфизм событий, способствующий определению этой ветви дерева Y-хромосомы ( ISOGG 2012 ).
- ^ В этой таблице показаны исторические названия N-M46 (AKA N-Tat) из рецензируемой литературы.
YCC 2002/2008 (сокращенно) Н-М46/Н-ТАТ Джоблинг и Тайлер-Смит, 2000 г. 12 Андерхилл 2000 VIII Хаммер 2001 1я Карафет 2001 26 Семино 2000 ЕС13 Они в 1999 году Н5 Волосы 2001 Ф YCC 2002 (от руки) N3* YCC 2005 (от руки) N3 YCC 2008 (от руки) N1c YCC 2010r (от руки) N1c - ^ В этой таблице показаны исторические названия N-M178 из рецензируемой литературы.
YCC 2002/2008 (сокращенно) Н-М178 Джоблинг и Тайлер-Смит, 2000 г. 16 Андерхилл 2000 VIII Хаммер 2001 1я Карафет 2001 26 Семино 2000 ЕС14 Они в 1999 году Н5 Волосы 2001 Ф YCC 2002 (от руки) Н3а* YCC 2005 (от руки) М178 YCC 2008 (от руки) N1c1 YCC 2010r (от руки) N1c1 - ^ В этой таблице показаны исторические названия N-M128 из рецензируемой литературы.
YCC 2002/2008 (сокращенно) Н-М128 Джоблинг и Тайлер-Смит, 2000 г. 12 Андерхилл 2000 VIII Хаммер 2001 1U Карафет 2001 25 Семино 2000 ЕС16 Они в 1999 году Н5 Волосы 2001 Ф YCC 2002 (от руки) N1 YCC 2005 (от руки) N1 YCC 2008 (от руки) Н1а YCC 2010r (от руки) Н1а - ^ Эту ветвь на ранних деревьях иногда называют N1b.
Внешние ссылки
[ редактировать ]- Распространение гаплогруппы N , из Генографического проекта , National Geographic.
- Северо-евразийский проект YDNA в FamilyTreeDNA
- Проект гаплогруппы N Y-ДНК в FamilyTreeDNA
- Проект гаплогруппы N1c1 Y-ДНК в FamilyTreeDNA
- Распространение гаплогруппы N Y-хромосомы из Южной Сибири в Европу
- Проект ДНК династии Рюриковичей в FamilyTreeDNA
- Проект ДНК русского дворянства в FamilyTreeDNA