Гаплогруппа N-M231
Эта статья , возможно, содержит оригинальные исследования . ( Ноябрь 2021 г. ) |
| Гаплогруппа N | |
|---|---|
 | |
| Возможное время происхождения | 36 800 [95% ДИ 34 300–39 300] лет назад (YFull [ 1 ] ) 44 700 или 38 300 лет назад в зависимости от скорости мутации [ 2 ] 41 900 [95% ДИ 40 175–43 591] лет назад [ 3 ] |
| Возраст слияния | 21 700 [95% ДИ 19 500–23 900] лет назад (YFull [ 1 ] ) 25 313 [95% ДИ 21 722–28 956] лет назад [ 3 ] |
| Возможное место происхождения | Северо- Восточная Азия [ 4 ] [ 5 ] |
| Предок | НЕТ |
| Определение мутаций | М231 |
| Самые высокие частоты | Нганассан 58% [ 3 ] –94.1%, [ 6 ] Якуты 81,8% [ 7 ] –94.6%, [ 8 ] Хакасия ( Ширинский район ) 90,2%, [ 9 ] Siberian Tatars 33.5% ( Zabolotnie 89.5%, Iskero-Tobol 23.5%, Tatar-Bukharans 16.5%, Yalutorov 16.3%, Ishtyak-Tokuz 14.5% [ 10 ] ), угры 77,8% [ 3 ] ( Khanty 64.3% [ 11 ] –89.3%, [ 12 ] Манси 76% [ 12 ] ), удмурты 77,8%, [ 3 ] Хакасы 41% [ 3 ] –65%, [ 11 ] Приходите 33,3% [ 11 ] –79.5%, [ 6 ] Ненцы 75–92,9% [ 3 ] (тундровые ненцы 97,9%, [ 6 ] Лесные ненцы 98,8% [ 6 ] ), вепсы 55%, [ 3 ] Доступно 42,6% (Запад) [ 13 ] –70,9% (Восток) [ 13 ] или ок. 54% [ 3 ] –58.8%, [ 14 ] Тувинцы 27,2–54,5%, [ нужна ссылка ] Нана 46,2% [ 3 ] [ 15 ] [ 16 ] (20% Хэжэ в КНР, [ 15 ] 44,6% нанайцы в России, [ 3 ] 83,8% члены рода Самар в Горинском районе Хабаровского края. [ 16 ] ), карелы 37,1% [ 5 ] –53.8%, [ 13 ] Архангельск Русские 42,6% (Архангельск 44,3%, [ 3 ] Пинега 40,8% [ 3 ] ), литовцы 40,5% [ 3 ] –44.5%, [ 13 ] латыши ок. 42% (41,6%, [ 13 ] 42.1%, [ 17 ] 43.0% [ 3 ] ), марийцы 41,2%, [ 3 ] Saami 40%, Chuvash 33.7% [ 14 ] –36%, [ 3 ] Буряты 34,5% (20,2%, [ 18 ] 25.0%, [ 19 ] 30.9%, [ 20 ] 48.0% [ 21 ] >), Koryaks 33.3%, [ 6 ] Эстонцы 30,6% [ 3 ] -33.9%, [ 13 ] Волжские татары 27,8%, [ 3 ] Телеуты 25,0%, [ 6 ] Северные Алтайцы 21,8% (18,0% [ 11 ] [ 22 ] –24.6% [ 23 ] ), псковские русские 22,7%, [ 3 ] Русские 20% [ 24 ] Башкиры 17,3%, [ 3 ] Нас было 17,1% [ 15 ] –18.0%, [ 25 ] Мордва 12,5% (10% [ 3 ] –13.3% [ 3 ] ), монголы 11%, [ 26 ] [ 20 ] [ 15 ] [ 19 ] [ 27 ] [ 28 ] Kalmyks 10.4% ( Torguud 3.4%, Derbet 5.1%, Buzava 5.3%, Khoshut 38.2%), [ 29 ] [ 28 ] Маньчжуры 10% (5,8%, [ 20 ] 8.1%, [ 30 ] 9.1%, [ 25 ] 11.6%, [ 25 ] 12.5%, [ 25 ] 14.3% [ 15 ] ), белорусы 9,7%, [ 3 ] Средне-южные россияне 9,1% ( Тверь 13,2%, [ 31 ] Курск 12,5% [ 31 ] –13.3%, [ 3 ] Белгород 11,9%, [ 3 ] Кострома 11,8%, [ 3 ] Смоленск 7,0%, [ 3 ] Воронеж 6,3%, [ 3 ] Орел 5,5% [ 3 ] ), украинцы 9,0%, [ 3 ] Южные Алтайцы 7,1% (4,2% [ 23 ] -9.7% [ 11 ] ), мул 7,1%, [ 32 ] Швеция 6,8% [ 14 ] (0% Вестра-Гёталанд , Халланд , Мальмё и Йёнчёпинг [ 33 ] - 19,5% Вестерботтен [ 34 ] ), ханьцы 6,77% (от 0% до 21,4%), [ 25 ] Корейцы 6,58% (от 4,41% до 12%) 12% корейцы, [ 35 ] 6,58% корейцы из KPGP (Корейский проект генома), [ 36 ] 6,9% Чеджу [ 19 ] 6,4% Гочан [ 37 ] 6,3% Канвондо [ 19 ] 5,7% Северокорейские [ 38 ] 4,8% Кёнсан, [ 19 ] 4,4% Чолла, [ 19 ] 4,2% Чхунчхон, [ 19 ] 4,0% Сеул, [ 39 ] 3,0% Тэджон, [ 39 ] 1,8% Сеул-Кёнги, [ 19 ] Ульчи 5,8%, [ 40 ] тибетцы 5,65%, [ 41 ] Казахи 5,33% [ 42 ] (Суан 0%, Канглы 0%, Ошакты 0%, Джетыру 1,2%, Дулат 1,6%, Аргын 2,0%, Алимулы 2,5%, Ысты 3,5%, Байулы 3,9%, Албан 4,3%, Конгырат 7,4%, Кыпшак 10,3%, Джалаир 10,9%, Кожа 16,7%, Сыргели 65,6%), Северный Тай 5,2%, [ 43 ] Уйгуры 4,89% (2,8%, [ 44 ] 4.8%, [ 25 ] 4.99%, [ 45 ] 6.0%, [ 20 ] 8.6% [ 15 ] ), Кыргызские 3,9% (2,8% Кызылсу , [ 46 ] 3,3% Кызылсу , [ 47 ] 4,5% Кыргызстан , [ 27 ] 10% Урумчи [ 46 ] ), вьетнамцы 3,4%, японцы 1,9% (0%, [ 2 ] 0.8%, [ 48 ] 0.9%, [ 49 ] 1.7%, [ 50 ] 2.5%, [ 19 ] 4.3%, [ 51 ] 4.8%, [ 20 ] 6.4% [ 15 ] ) |
Гаплогруппа N (M231) представляет собой гаплогруппу ДНК Y-хромосомы, определяемую наличием маркера однонуклеотидного полиморфизма (SNP) M231. [ Филогенетика 1 ]
Чаще всего встречается у мужчин, происходящих из Северной Евразии . Это также наблюдалось с более низкой частотой у популяций, обитающих в других регионах, включая части Балкан , Центральной Азии , Восточной Азии и Юго-Восточной Азии .
Однако базальная парагруппа N* была обнаружена только у коренных популяций Китая и Камбоджи . [ 4 ] Субклады N-M231 были обнаружены в небольших количествах в Юго-Восточной Азии , на островах Тихого океана , Юго-Западной Азии и на Балканах . Эти факторы позволяют предположить, что он возник в Восточной или Юго-Восточной Азии.
Происхождение
[ редактировать ]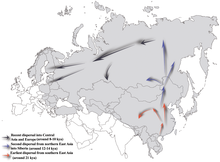
Гаплогруппа NO-M214 - ее последний общий предок с ее родным братом, гаплогруппой O-M175 - по оценкам, существовала около 36 800–44 700 лет назад. [ 1 ] [ 53 ] [ 2 ]
Принято считать, что N-M231 возник в Восточной Азии примерно 19 400 (±4 800) лет назад и заселил север Евразии после последнего ледникового максимума . Самцы, несущие маркер, по-видимому, двинулись на север по мере потепления климата в голоцене , мигрируя против часовой стрелки, и в конечном итоге концентрировались в таких далеких районах, как Фенноскандия и Балтика . [ 5 ] Очевидный недостаток гаплогруппы N-M231 среди коренных американцев указывает на то, что она распространилась после Берингии . затопления [ 54 ] около 11 000 лет назад.
Распределение
[ редактировать ]
Гаплогруппа N имеет широкое географическое распространение по всей северной Евразии, а также время от времени наблюдалась в других регионах, включая Центральную Азию и Балканы.
С наибольшей частотой он встречается у коренных народов России , в том числе у финских народов , марийцев , удмуртов , коми , хантов , манси , ненцев , нганасан , тюркских народов (якутов, долган, хакасов, тувинцев, татар, чувашей и др. ), Буряты , тунгусские народы ( эвенки , эвены , негидальцы , нанайцы ) и др. , юкагиры , луораветланы (чукчи, коряки) и сибирские эскимосы , но отдельные субклады очень распространены в Финляндии , Эстонии , Латвии и Литве , а другие субклады встречается с низкой частотой в Китае (И, Наси, Лхоба, ханьцы и т. д. ). [ 55 ] Особенно у этнических финских народов и балтоязычных народов Северной Европы, обско-угроязычных и северных самоедских народов Западной Сибири и тюркоязычных народов России (особенно якутов , [ нужна ссылка ] ,но также алтайцы , шорцы , хакасы , чуваши , татары и башкиры ). Почти все представители гаплогруппы N среди этих популяций северной Евразии принадлежат к субкладам либо гаплогруппы N-CTS6128/M2048, либо гаплогруппы N-P43.
Y-хромосомы, принадлежащие N1b-F2930/M1881/V3743 или N1*-CTS11499/L735/M2291(xN1a-F1206/M2013/S11466), обнаружены в Китае (где они составляют примерно 3,62% всей Y-ДНК). [ 56 ] ) и спорадически в других частях Евразии. Субклады N-CTS6128/M2048 и N-P43 N1a-F1206/M2013/S11466 в большом количестве встречаются в Северной Евразии; однако члены N1a-F1206(xCTS6128, P43) в настоящее время встречаются в основном в северном Китае и Корее.
N2-Y6503, другой первичный субклад гаплогруппы N, чрезвычайно редок и в основном представлен среди современных людей недавно сформировавшимся субкладом, который практически ограничен странами, составляющими бывшую Югославию (Босния-Герцеговина, Хорватия, Сербия и Черногория). ), Венгрия и Австрия. Среди других членов N2-Y6503 — венгр недавнего происхождения из Сучавы на Буковине , словак, несколько британцев и алтайец . [ 1 ]
Н* (М231)
[ редактировать ]Y-хромосомы, которые содержат мутацию M231, определяющую гаплогруппу N-M231, но не имеют мутаций CTS11499, L735, M2291, определяющих гаплогруппу N1, считаются принадлежащими к парагруппе N-M231*. [ 4 ]
N-M231* был обнаружен в Китае в небольших количествах. [ 4 ] Из выборки из 165 ханьцев из Китая два человека (1,2%) оказались принадлежащими к N*. [ 57 ] Один из них был родом из Гуанчжоу , другой — из Сианя . [ нужна ссылка ]
Среди древних образцов байкальской культуры раннего неолита Китой один из образцов Шаманка II (DA250), датированный ок. 6500 BP, в оригинальном исследовании был проанализирован как NO1-M214. [ 58 ] Однако впоследствии было обнаружено, что этот же образец (DA250 или Шаманка 250) принадлежит N-FT210118, той же кладе, что и другие образцы гаплогруппы N с того же сайта (кроме DA247, который принадлежит N-Y147969). N-FT210118 происходит от N-L666/N-F2199, но является базальным по отношению к N-CTS6380, причем последний является самым недавним общим предком современного N-P43 (в основном встречается среди марийцев, удмуртов, коми, чувашей, татар, ненцев). , нганасаны, ханты, манси, хакасы, тувинцы и др. ) и N-F1101 (встречается преимущественно среди выходцев из Восточной Азии). Более того, N-FT210118 не был обнаружен ни у одного живого человека, у которого на сегодняшний день было проверено его Y-ДНК, а предполагаемое TMRCA N-CTS6380 превышает предполагаемую дату осаждения любого из образцов с участка Шаманка, связанного с Китойская культура, поэтому получается, что представители китойской культуры в Шаманке (или, по крайней мере, их Y-ДНК) вымерли, а не являются прямыми предками каких-либо ныне живущих людей. [ 59 ] [ 60 ]
N1 (CTS11499, Z4762, CTS3750)
[ редактировать ]В 2014 году в определении субклада N1 произошли серьезные изменения, когда LLY22g был исключен из качестве основного определяющего SNP для N1 из-за сообщений о ненадежности LLY22g. Согласно ISOGG , LLY22g является проблематичным, поскольку является «палиндромным маркером и его легко неправильно интерпретировать». [ 4 ] С тех пор название N1 стало применяться к кладе, отмеченной большим количеством SNP, включая CTS11499, Z4762 и CTS3750. N1 является самым недавним общим предком всех существующих членов гаплогруппы N-M231, за исключением членов редкого субклада N2-Y6503 (N2-B482). По оценкам, TMRCA N1 возник за 18 000 лет до настоящего времени (16 300–19 700 лет назад; 95% ДИ). [ 1 ] После пересмотра 2014 года положение многих примеров «N1-LLY22g» в гаплогруппе N стало неясным. Поэтому лучше проверить yfull и ISOGG 2019 , чтобы понять обновленную структуру N-M231.
Однако в более ранних исследованиях сообщалось, что частота N-LLY22g достигает 30% (13/43) среди народа И из округа Буто , Сычуань на юго-западе Китая . [ 20 ] [ 61 ] [ 62 ] Он также встречается у 34,6% людей лхоба . [ 62 ] N1-LLY22g* был обнаружен в образцах ханьцев , но с разной частотой:
- 6,8% (3/44) Хан из Сианя [ 20 ] [ 61 ]
- 6,7% (2/30) Хан из Ланьчжоу [ 15 ]
- 3,6% (3/84) тайваньские хань [ 20 ]
- 2,9% (1/34) Мужчины из Чэнду [ 15 ]
- 2,9% (1/35) Хан из Харбина [ 15 ]
- 2,9% (1/35) Хан из района Мэйсянь [ 15 ]
- 0% (0/32) Хан из города Инин [ 15 ]
Другие популяции, в которых были обнаружены представители N1*-LLY22g, включают:
- Хани (4/34 = 11,8%) [ 15 ]
- Сибе (4/41 = 9,8%) [ 15 ]
- Люди туцзя (2/49 = 4,1%) [ 20 ]
- Маньчжуры (2/52 = 3,8% [ 20 ] до 2/35 = 5,7% [ 15 ]
- Битые люди (1/28 = 3,6%) [ 63 ]
- Уйгуры (2/70 = 2,9% [ 15 ] до 2/67 = 3,0%) [ 20 ]
- Тибетцы (3/105 = 2,9% [ 20 ] до 3/35 = 8,6%) [ 15 ]
- Корейцы (0/106 = 0,0% – 2/25 = 8% [ 5 ] [ 15 ] [ 64 ]
- Вьетнамцы (2/70 = 2,9%) [ 20 ]
- Японцы (0/70 Токусима - 2/26 = 7,7% Аомори ) [ 20 ]
- Маньчжурские эвенки (0/26 = 0,0% [ 15 ] до 1/41 = 2,4%) [ 20 ]
- Алтайцы (от 0/50 северных до 5/96 = 5,2% южных, или от 0/43 бешпельтир до 5/46 = 10,9% кулада), [ 20 ] [ 22 ]
- Шорцы (2/23 = 8,7%) [ 5 ]
- Хакасы (5/181 = 2,8%) [ 5 ]
- Тувинцы (5/311 = 1,6%) [ 5 ]
- Южный Борнео (1/40 = 2,5%) [ 5 ]
- Лесные ненцы (1/89 = 1,1%) [ 5 ]
- Якуты (0/215 – 1/121 = 0,8%) [ 5 ]
- Турки (1/523 = 0,2%) [ 5 ] В Турции общее количество субкладов гаплогруппы N-M231 составляет 4% мужского населения.
- Одна особь, принадлежащая либо к N*, либо к N1*, была обнаружена в выборке из 77 мужчин из Катманду, Непал (1/77 = 1,3% N-M231(xM128,P43,Tat)) [ 65 ]
N1(xN1a,N1c) был найден в древних костях цивилизации Ляо : [ 66 ]
- Нюхэлян ( культура Хуншань , 6500–5000 лет назад) 66,7% (= 4/6)
- Халахайгоу ( культура Сяохэянь , 5000–4200 лет назад) 100,0% (= 12/12)
- Дадяньцзы ( культура Нижнего Сяцзядянь , 4200–3600 лет назад) 60,0% (=3/5).
N-CTS4309: два человека идентифицированы как представители этой подгруппы в Ираке. Очень редко.
Н1а (Ф1206/М2013/С11466)
[ редактировать ]По оценкам, клада N1a2-F1008/L666 и N1a1-M46/Page70/Tat имеют общего предка N1a-F1206/M2013/S11466 примерно за 15 900 [95% ДИ 13 900 <-> 17 900] лет до настоящего времени. [ 1 ] или 17 621 [95% ДИ 14 952 <-> 20 282] года назад. [ 3 ]
Н1а1 (М46/Страница70/Тат, L395/M2080)
[ редактировать ]Все M46 в базе данных Yfull — это M178 , что на четверть моложе, чем отделение от F1139. [ 67 ]
Мутации, определяющие субклад N-M46 [ Филогенетика 2 ] являются M46/Tat и P105. Это наиболее распространенный субклад N. Вероятно, он возник в популяции Северо-Восточной Азии, поскольку древнейшие древние образцы соответствуют этому генетическому профилю. [ 68 ] Компания N столкнулась с периодическими узкими местами в Сибири и вторичной экспансией в Восточной Европе. [ 5 ] Гаплогруппе N-M46 около 14 000 лет.
В Сибири гаплогруппа N-M46 достигает максимальной частоты примерно 90% у якутов — тюркского народа, проживающего преимущественно в Республике Саха (Якутия) . Однако Н-М46 с гораздо меньшей частотой присутствует у многих соседей якутов, таких как эвенки и эвены . [ 8 ] Он также был обнаружен у 5,9% (3/51) образца хмонг-до из Лаоса. [ 63 ] 2,4% (2/85) выборки из Сеула, Южная Корея, [ 26 ] и в 1,4% (1/70) выборки из Токусимы, Япония. [ 20 ]
Гаплогруппа N-M46 имеет низкое разнообразие среди якутов, что позволяет предположить наличие узкого места в популяции или эффекта основателя . [ 69 ] Это было подтверждено исследованием древней ДНК, которое проследило происхождение мужских якутских линий от небольшой группы всадников из Прибайкалья. [ 70 ]
N-Tat наблюдался с очень разной частотой в образцах из Швеции . Карлссон и др. (2006) обнаружили N-Tat у 44,7% (17/38) выборки саамских кочевников из Йокмокка , 19,5% (8/41) выборки из Вестерботтена, 14,5% (8/55) выборки из Уппсалы, 10,0% (4/40) образца из Готланда, 9,5% (4/42) образца из Вермланда, 7,3% (3/41) образца из Эстергётланда/Йёнчёпинга, 2,4% (1/41) образец из Блекинге/Кристианстад и 2,2% (1/45) образца из Скараборга. [ 34 ]
Лаппалайнен и др. (2008) обнаружили N-Tat в 14,4% (23/160) выборки из Швеции. [ 13 ]
Лаппалайнен и др. (2009) обнаружили N-Tat в 15,4% (4/26) образца из Седерманланда, в 12,5% (3/24) образца из Вестманланда, в 12,1% (4/33) образца из Уппсалы, 7,8 % (4/51) выборки из Гетеборга, 7,0% (3/43) выборки из Норботтена, 6,8% (5/73) выборки из Сконе, 6,6% (15/228) выборки из Стокгольма , 6,3 % (3/48) образца из Сидноррланда, 6,3 % (2/32) образца из Вестерботтена, 6,3 % (2/32) образца из Эребру, 5,9 % (3/51) образца из Вермланда/Даларны, 5,4% (2/37) выборки из Остра-Гёталанда и 5,1% (2/39) выборки из юго-восточной Швеции (Кальмар, Готланд, Кроноберг и Блекинге). Они не обнаружили ни одного экземпляра N-Tat в своих образцах из Йёнчепинга (0/28), Мальмё (0/29), Халланда (0/34) или Вестра-Гёталанда (0/75). [ 33 ]
Н1а1а (М178)
[ редактировать ]Субклад N-M178 [ Филогенетика 3 ] определяется наличием маркеров М178 и Р298. N-M178* имеет более высокую среднюю частоту в Северной Европе, чем в Сибири, достигая частоты примерно 60% среди финнов и примерно 40% среди латышей , литовцев и 35% среди эстонцев . [ 71 ] [ 13 ]
Мирослава Деренко и ее коллеги отметили, что внутри этой гаплогруппы есть два субкластера, присутствующие в Сибири и Северной Европе, с разной историей. Тот, который они назвали N3a1, сначала распространился на юге Сибири и распространился на Северную Европу. Между тем, более молодой субкластер, который они назвали N3a2, зародился на юге Сибири (вероятно, в Прибайкалье). [ 71 ]
N-M178 также был обнаружен у двух говорящих на языке тлычу в Северной Америке. [ 72 ]
Неолитические образцы из Прибайкалья дали множество образцов yДНК N, а один образец из Фофоново , Бурятия , 5000-4000 гг. до н.э., является одним из первых образцов Тат в древних записях. [ 68 ]
Самые ранние образцы N1a1a-L708 были обнаружены в Забайкалье (brn008, N1a1a1*-L708; brn003, N1a1a1a1*-M2126) между 8000 и 6000 YBP. Пробы ниже по течению были обнаружены в Якутии (N4b2, N1a1a1a1a*-Z1979) и Красноярском крае (kra001, N1a1a1a1a*-L392) на глубине от 5000 до 4000 YBP. [ 73 ] [ 74 ]
Н1а2 (F1008/L666)
[ редактировать ]По оценкам, N1a2a-M128 и N1a2b-B523/P43 имеют общего предка N1a2-F1008/L666 примерно за 8600 [95% ДИ 7500 <-> 9800] лет до настоящего времени, [ 1 ] 9200 лет назад, [ 75 ] или 9314 [95% ДИ 7419 <-> 11 264] лет назад. [ 3 ]
По крайней мере три из шести протестированных мужских экземпляров из слоя раннего неолита (использующего керамику охотников-собирателей примерно 7200–6200 лет назад) на археологическом памятнике Шаманка у южной оконечности озера Байкал оказались принадлежащими к N1a2-L666. [ 58 ]
Н1а2а-М128
[ редактировать ]| Гаплогруппа N-M128 | |
|---|---|
 | |
| Возможное место происхождения | Азия |
| Предок | Н1а2 (F1008/L666) |
| Потомки | Ф710, Ф1998 |
| Определение мутаций | М128 |
| Самые высокие частоты | Маньчжуры около 1% (0/43 Бицзе , [ 76 ] 0/30, [ 19 ] 30/2938 = 1,0% всего Китая [ 77 ] 1/52 = 1,9% Ляонин, [ 20 ] 2/35 = 5,7% Сюань Маньчжурский автономный округ ), [ 15 ] Корейцы 0/43 Южная Корея, [ 15 ] 0/25 Китай, [ 15 ] 1/75 = 1,3% Южная Корея, [ 20 ] Сибе 1/41 = 2,4%, [ 15 ] Хань 0,89% (Шаньси 1,83%, Хэнань 1,53%, Ганьсу 1,46%, Хэбэй 1,41%, Шэньси 1,40%, Тяньцзинь 1,38%, Шаньдун 1,17%, Пекин 1,14%, Внутренняя Монголия 1,13%, Хэйлунцзян 1,12%, Ляонин 1,12%, Фуцзянь 1,12%, Аньхой 0,93%, Цзилинь 0,86%, Нинся 0,82%, Цзянсу 0,81%, Юньнань 0,79%, Хубэй 0,77%, Чжэцзян 0,73%, Тайвань 0,70%, Сычуань 0,54%, Хунань 0,52%, Чунцин 0,51%, Гуйчжоу 0,5 1% , Гуандун 0,45%, Хайнань 0,43%, Синьцзян 0,43%, Шанхай 0,36%, Цзянси 0,34%, Гуанси 0,23%, Цинхай 0,18%), монголы 0,7% (Ляонин 1,9%, Хорцинь 0,3%, Улан-Батор 0,0%), японцы 0,3% (Токушима 3,5%, Ибараки 2,0%, Хиросима 1,3%, Нагоя 1,0%, Осака 0,2%, Токио 0,0%, Сидзуока 0,0%, Ямагути 0,0%), казахи 0,1% (Ходжа 1,9%, Байулы 0,6%, Уйсун 0,4%) |
Этот субклад определяется наличием маркера M128. [ Филогенетика 4 ] N-M128 был впервые идентифицирован в образце из Японии (1/23 = 4,3%) и в образце из Центральной Азии и Сибири (1/184 = 0,5%) в ходе предварительного исследования вариаций Y-ДНК во всем мире. [ 51 ] В дальнейшем он был обнаружен с низкой частотой у маньчжуров , сибэев , эвенков , корейцев , ханьцев , хуэйцев , тибетцев , вьетнамцев , буэйцев , казахов , узбеков , уйгуров , саларов , ту , монголов , бузавцев . племя калмыков , [ 29 ] Хакасы и коми . [ 5 ]
Было обнаружено , что ряд ханьских китайцев, олед- монголов , цянов и тибетцев принадлежали к сестринской ветви (или ветвям) N-M128 под парагруппой N-F1154*. [ 78 ]
Неолитический образец brn002 (~5940 л.н.) в Забайкалье оказался ранним ответвлением выше по течению от N-M128. [ 79 ]
В результате генетического тестирования клана Елюй , королевской семьи династии Ляо и потомков киданей , было обнаружено, что он принадлежит N-F1998, расположенному ниже по течению от N-M128.
Согласно «Глубокой истории населения северной части Восточной Азии от позднего плейстоцена до голоцена», [ 80 ] базальная yДНК N-M128/мтДНК B5b2 HGDP01293 индивидуума [ 81 ] занимал позицию на PCA между образцами из провинции Цзянсу и провинции Аньхой, но недалеко от образца мтДНК R11'B6>R11b провинции Шаньдун, [ 80 ] в то время как о более позднем потомке клады yDNA N-M128, принадлежащем мтДНК R11'B6>R11b, сообщалось из древней ДНК кладбища Западный Чжоу, гробницы M18. [ 82 ] На основе древней ДНК распространение мтДНК B5b2 после 9500 лет назад и до 4600 лет назад в направлении провинций Аньхой и Цзянсу от Шаньдуна из окрестностей будущего Шаньдунского городища Луншань Иньцзячэн показано в «Материнской генетической структуре в древних Шаньдун между 9500 и 1800 годами назад». [ 83 ] при этом существование локального палеолитического северо-восточноазиатского субстрата, представленного особями базально вымершей yДНК О-М164*, отделившейся от юго-восточноазиатской yДНК О-М188 и дающей вклад в yДНК C2-M217 предков алтайских и корейских представителей , было показано в «Генетической истории человека на Тибетском нагорье за последние 5100 лет». [ 84 ] Заняв позицию на PCA между образцами из провинции Цзянсу и провинции Аньхой, базальная особь yDNA N-M128/mtDNA B5b2 HGDP01293 стала участником единой генетической линии, которая простиралась от особей Цзянсу и Аньхой до тайскоязычного дай народа . , и от особей Цзянсу и Аньхой до древней особи WGM20, принадлежащей мтДНК M11, из стоянки Яншао Вангоу, датируемой 5000-5500 лет назад, и этот древний возраст также охватывал древнюю yDNA до N-M128 особей Мазонгшаня. [ 85 ] и современные особи провинции Ганьсу, связанные с yDNA N-M128, которые, по-видимому, были включены в упомянутую генетическую линию, более близкую к образцам древней провинции Хэнань периода Луншань ок. 4000 лет назад, чем более генетически базальная древняя особь WGM20 стоянки Яншао Вангоу, датируемая 5000-5500 лет назад. [ 80 ]
N1a2b(P43)
[ редактировать ]Гаплогруппа N-P43 [ Филогенетика 5 ] определяется наличием маркера P43. Кроме того, гаплогруппа N-P43 определяется маркером Y3214, который является общим с более молодой yDNA O1b2-K14, распространенной в Японии (YFull). Его возраст оценивается примерно от 4000 до 5500 лет (TMRCA 4510 лет, [ 75 ] TMRCA 4700 [95% ДИ 3800 <-> 5600] лет назад, [ 1 ] или 4727 [95% ДИ 3824 <-> 5693] лет назад). [ 3 ] Он очень часто встречается среди северных самодийских народов , носителей обско-угорских языков и северных хакасов , а также с низкой или умеренной частотой наблюдается среди носителей некоторых других уральских языков , тюркских народов , монгольских народов , тунгусских народов , и сибирские юпики .
Самые высокие частоты N-P43 наблюдаются среди популяций Северо-Западной Сибири: 92% (35/38). [ 5 ] в выборке Нганасан 78% (7/9) [ 86 ] [ 14 ] в выборке энцев 78% (21/27) [ 31 ] в выборке хантов 75% (44/59) [ 5 ] в выборке тундровых ненцев - 69% (29/42) [ 3 ] в другой выборке ненцев - 60% (15/25) [ 12 ] в выборке манси - 57% (64/112) [ 11 ] в другой выборке хантов - 54% (27/50) [ 3 ] в другой выборке Нганасан — 45% (40/89) [ 5 ] в выборке лесных ненцев - 38% (18/47) [ 87 ] в третьей выборке хантов и 25% (7/28) [ 12 ] в четвертом образце хантов. В Европе типы N-P43 имеют самую высокую частоту - 20% среди популяций Волго-Уральского региона. Крайней западной границей распространения N-P43 является Финляндия, где эта гаплогруппа встречается лишь с предельной частотой – 0,4%. Тем не менее, N-P43 довольно часто встречается среди вепсов (17,9%), небольшой финской популяции, живущей в непосредственной близости от финнов, карел и эстонцев. [ 5 ]
Гаплогруппа N-P43 также с очень высокой частотой наблюдалась (26/29 = 89,7% выборки из пос. Топанов и 19/22 = 86,4% выборки из пос. Малый Спирин) в выборках Качинцев, Тюркоязычный этнос или территориальная подгруппа хакасов , из Ширинского района Северной Хакасии . [ 9 ] Судя по всему, существует линия через сагаев (еще одну тюркоязычную этническую группу, которая сейчас считается составной частью хакасов): 46,2% (55/119) сагаев были отобраны из Усть-Еса, Есино, Поселения Усть-Чуль и Кызлас Аскизского района Центральной Хакасии, принадлежащие к гаплогруппе N-P43, против всего 13,6% (11/81) сагаев, отобранных из поселений Матур, Анчуль, Большая Сея и Бутрахты Таштыпского Район южной Хакасии, принадлежащий к этой гаплогруппе. [ 9 ] Однако выборки хакасов, проведенные другими исследователями, показали лишь умеренную частоту N-P43 или потенциального N-P43. Деренко и др. (2006) исследовали выборку хакасов ( n =53), собранную в населенных пунктах Аскизского, Ширинского, Бейского и Орджоникидзевского районов Республики Хакасия, и установили, что 15 из них (28,3%) принадлежали к N-LLY22g(xTat). [ 18 ] Рутси и др. (2007) исследовали выборку хакасов ( n =181) и обнаружили, что 31 из них (17,1%) принадлежал к N-P43; [ 5 ] повторно протестировали 174 человека в этой выборке и обнаружили, что 27 из них (15,5%) принадлежали к субкладу N-B478 (азиатский/северный самодийский) субклада N-P43, а 2 из них (1,1%) принадлежали к N-L1419 ( Европейский/волжско-финский и чувашский) субклад N-P43, всего 29 (16,7%) N-P43. [ 3 ]
Гаплогруппа N-P43 образует два различных субкластера гаплотипов STR: азиатский и европейский, последний в основном распространен среди финно-угороязычных народов и родственных им популяций. [ 5 ]
N1a2b1-B478
[ редактировать ]По оценкам, TMRCA N-B478 составляет 3007 [95% ДИ 2171 <-> 3970] лет назад. [ 3 ] Это одна из наиболее распространенных гаплогрупп Y-ДНК среди коренного населения Северо-Западной Сибири: 69,0% (29/42) ненцы, 50,0% (25/50) нганасаны, 22,2% (12/54) долганы с Таймыра, 7,0% ( 3/43) Селькупы, 1,6% (1/63) обские угры. Достаточно распространена она и среди популяций Средней Сибири, Южной Сибири и Монголии: 17,9% (17/95) тувинцев, 15,5% (27/174) хакасов, 13,0% (6/46) тожуйских тувинцев , [ 28 ] 8,7% (2/23) шорцы, 8,3% (2/24) эвены, 8,2% (5/61) алтайцы, 5,3% (3/57) эвенки, 5,0% (19/381) монголы, 4,9% (3/ 61) Сарт-Калмак (частичные потомки ойратских монголов в Киргизии), [ 28 ] 4.2% (9/216) Yakut, 2.1% (1/47) Torgut (Mongolia), [ 28 ] 1,4% (1/69) Дербет (Калмыкия), [ 28 ] 0,9% (1/111) буряты. Географически удаленный член обнаружен в выборке чувашей (1/114 = 0,88%). [ 3 ]
По данным Карафета и др. (2018) обнаружили N-P63, который, по-видимому, примерно филогенетически эквивалентен N-B478, у 91,2% (31/34) рыб, 63,8% (30/47) тундровых ненцев и 42,7% (35/82) лесных. Ненцы, 14,0% (8/57) Долганы, 7,0% (9/129) Селькупы, 3,3% (3/91) Эвенкии, 2,7% (2/75) Монголы, 2,6% (2/78) Коми, 2,5% ( 2/80) бурятские и 2,0% (2/98) алтайские кижи. [ 6 ] Эта гаплогруппа не наблюдалась в выборках юкагиров (0/10), коряков (0/11), телеутов (0/40), кетов (0/44), якутов (0/62) или хантов (0/165). населения. [ 6 ]
Харьков и др. (2023) обнаружили N-B478 в сильно различающихся процентных долях выборок хантов из двух разных сел Ханты-Мансийского автономного округа : 60,9% (39/64) выборки хантов из станицы Русскинской Сургутского района и 14,8% (8/54) выборки хантов из села Казым Белоярского района. Пять из восьми представителей села Казым делят субклад, обозначенный SNP B172 и Z35108, со всеми ранее обследованными ненцами из фратрии Вануйто, принадлежащими к родам Вануито, Пуйко и Яунгат, а также клану Пурунги хантыйского происхождения. [ 88 ]
Н1б (Ф2930)
[ редактировать ]Гаплогруппа N1b преимущественно обнаружена у народа И, тибето-бирманской этнической группы на юго-западе Китая, которая произошла от древних племен Цян на северо-западе Китая. [ 55 ] Однако он также обнаружен у людей по всему Китаю (где они составляют примерно 3,62% мужского населения страны и в основном распространены в Шаньдуне, Цзянсу, Чжэцзяне, Аньхое и др.). [ 89 ] ) и у некоторых людей из Испании, [ 59 ] Эквадор, [ 59 ] Польша, [ 59 ] [ 1 ] Беларусь, [ 59 ] Россия, [ 59 ] [ 1 ] Ирак, [ 59 ] Индия, [ 59 ] [ 1 ] Казахстан, [ 59 ] Корея, [ 59 ] [ 1 ] Япония, [ 59 ] [ 1 ] Бутан, [ 59 ] Вьетнам, [ 59 ] [ 1 ] Камбоджа, [ 59 ] [ 1 ] Лаос, [ 59 ] Таиланд, [ 59 ] [ 1 ] Малайзия, [ 1 ] и Сингапур. [ 1 ]
Н2 (И6503)
[ редактировать ]N2 (Y6503/FGC28528; B482/FGC28394/Y6584) – первичная ветвь гаплогруппы N-M231, сейчас представлена в основном субкладом N-FGC28435, который распространился, вероятно, где-то в первой половине второго тысячелетия нашей эры. [ 90 ] и это было обнаружено у людей из Сербии , Хорватии , Боснии и Герцеговины , Черногории и Турции ( Стамбул ). [ 91 ] [ 90 ]
N-Y7310 (или N-F14667) включает в себя N-FGC28435 и также, вероятно, происходит от общего предка, жившего где-то в первой половине прошлого тысячелетия. Однако члены N-Y7310(xFGC28435) имеют более широкий географический диапазон, включая человека из Ростовской области России и румынского венгра, родом из Сучавы , Буковина . [ 1 ]
В другие отделения N-P189 входят члены из Турции, [ 1 ] Россия ( Московская область ), [ 1 ] Франция ( Приморская Шаранта ), [ 1 ] и Англия ( Девон ). [ 1 ] [ 59 ] По оценкам, самый последний общий предок всех вышеупомянутых существующих линий N-P189 возник 4900 (95% ДИ 5700 <-> 4100) лет назад. [ 1 ] Археологический образец, относящийся к ботайской культуре Северного Казахстана и датируемый второй половиной четвертого тысячелетия до нашей эры, принадлежит к N-P189*, являясь базальным для современных европейских представителей N-P189. [ 92 ] [ 1 ]
Линии, принадлежащие N-Y6503 (xP189) и имеющие лишь отдаленное родство (время до появления самого последнего общего предка оценивается более чем за 10 000 лет до настоящего времени) [ 1 ] вышеуказанным представителям N-P189 обнаружены у человека из современной Республики Алтай. [ 1 ] и, вероятно, также в археологическом образце, относящемся к культуре Мезочат железного века на территории современной Венгрии (примерно 2900 лет назад). [ 93 ] и в археологическом образце, относящемся к китойской культуре собирателей- фуражиров в районе озера Байкал (около 6700 лет до настоящего времени). [ 92 ]
Древние народы
[ редактировать ]Было обнаружено, что образец, раскопанный на стоянке Хоутаомуга в районе Юнхэ поселка Хуганцзы, Даань , Цзилинь , Китай, датируемый 7430–7320 годами назад (фаза II раннего неолита), принадлежит к гаплогруппе N и Y-ДНК. гаплогруппа мтДНК B4c1a2 . Эта выборка аутосомно идентична неолитическим популяциям бассейна реки Амур, наиболее близким современным представителем которых являются нивхи. Поскольку в статье обнаружено это происхождение у экземпляра USR1 конечного плейстоцена на Аляске, предполагается, что произошел поток генов из Амура в Америку популяции, принадлежащей к гипотетической чукотско-камчатско-нивхской языковой семье. [ нужна ссылка ]
N также был обнаружен во многих образцах неолитических человеческих останков, эксгумированных в цивилизации Ляо на северо-востоке Китая и в Прибайкалье на юге Сибири. [ 66 ] Предполагается, что yDNA N достигла юга Сибири от 12 до 14 тыс. лет назад. Оттуда он достиг Южной Европы 8-10 тыс. лет назад. [ 52 ]
Филогения
[ редактировать ]Филогенетическое дерево
[ редактировать ]В следующем дереве номенклатура трех источников разделена косой чертой: Дерево ISOGG, 10 декабря 2017 г. (версия 12.317).
- НЕТ-М214
- Н-М231/Пейдж91, М232/М2188
- N1-Z4762/CTS11499/L735/M2291
- N1a-L729
- Н-З1956
- N-Y149447 Китай ( Шэньси [ 1 ] )
- Н1а1-М46/Страница70/Тат
- Н1а1а-М178
- N1a1a1-F1419
- N1a1a1a-L708
- N1a1a1a1-P298/M2126 Китай ( Урумчи , префектура Кашгар , Турфан , префектура Аксу , Сяньян , Цзиньчэн , Кайфэн , Цицикар [ 75 ] )
- N1a1a1a1a-L392
- N1a1a1a1a1-CTS10760
- N1a1a1a1a1a-CTS2929/VL29 Встречается с высокой частотой среди литовцев , латышей , эстонцев северо-запада , русских , шведских саамов , карел , ненцев , финнов и марийцев , умеренной частотой среди других русских , белорусов , украинцев и поляков и низкой частотой среди коми , Мордва , татары , чуваши , долганы , вепсы , селькупы , караногайцы и башкиры. [ 3 ]
- N1a1a1a1a1a1-Z4908
- N-Y46443
- N-Y46443* Россия ( Московская область) [ 1 ] )
- N-BY33095 Россия ( Самарская область [ 1 ] )
- Н1а1а1а1а1а1а-Л550/С431
- N-L550* Швеция ( округ Кроноберг) [ 1 ] )
- N-Y9454 Швеция ( округ Вестра-Гёталанд , [ 1 ] Округ Эребру [ 1 ] ), Финляндия, [ 1 ] Россия [ 1 ]
- N-Y20911 Финляндия (провинция Западная Финляндия, [ 1 ] Южная Финляндия [ 1 ] )
- N-Y7795
- N-Y7795* Швеция ( округ Норрботтен , [ 1 ] Лен Сёдерманланд [ 1 ] )
- N-Y29766 Норвегия ( Хедмарк [ 1 ] )
- N-Y20918 Швеция ( округ Эстергётланд , [ 1 ] Лен Вестерботтен [ 1 ] ), Финляндия (провинция Западная Финляндия [ 1 ] )
- N-Y28771
- N-Y61225 Швеция ( округ Сёдерманланд , [ 1 ] Округ Уппсала [ 1 ] )
- N-Y30126
- N-Y30126* Швеция ( округ Эстергётланд) [ 1 ] )
- N-Y29764
- N-Y29764* Швеция ( округ Стокгольм) [ 1 ] )
- N-Y30123 Швеция ( округ Вестерботтен) [ 1 ] ), Финляндия [ 1 ]
- Н-S9378
- N-S9378* Швеция ( округ Вестра-Гёталанд) [ 1 ] )
- N-S18447 Швеция ( лен Сёдерманланд , [ 1 ] Округ Уппсала [ 1 ] )
- N-Y36282
- N-Y36282* Финляндия (провинция Южная Финляндия) [ 1 ] ), Эстония ( Ида-Вирумаа [ 1 ] )
- N-BY21957 Польша ( Куявско-Поморское воеводство) [ 1 ] ), Швеция ( округ Стокгольм [ 1 ] ), Россия ( Псковская область [ 1 ] ), Финляндия (провинция Южная Финляндия [ 1 ] )
- Н-FGC14542
- N-FGC14542* Финляндия (провинция Восточная Финляндия) [ 1 ] )
- N-BY21938 Норвегия ( Хедмарк [ 1 ] ), Швеция ( округ Даларна [ 1 ] )
- N-Y17113 Великобритания ( Саффолк , [ 1 ] Шотландия [ 1 ] ), Канада ( Манитоба [ 1 ] ), Норвегия ( Вестфолд , [ 1 ] Бускеруд [ 1 ] ), Швеция ( округ Вестра-Гёталанд) [ 1 ] )
- N-Y4341
- N-BY21874 Швеция ( лен Сёдерманланд [ 1 ] ), Финляндия [ 1 ]
- N-Y4338
- N-Y4338*
- N-Y4339 Швеция (средневековая Сигтуна [ 1 ] )
- N-Y12104
- N-Y19111
- N-Y57577 Швеция ( округ Сконе) [ 1 ] )
- N-Y22774 Финляндия (провинция Западная Финляндия) [ 1 ] )
- N-Y5611
- N-Y5611* Англия [ 1 ]
- N-Y21546 Швеция ( округ Стокгольм , [ 1 ] Лен Вестманланд [ 1 ] )
- N-F1983 Швеция ( графство Евлеборг) [ 1 ] ), Russia ( Lipetsk Oblast [ 1 ] )
- N-Y10932
- N-Y85136 Швеция ( округ Уппсала) [ 1 ] )
- N-Y10931 Россия [ 1 ]
- N1a1a1a1a1a1a1-L1025/B215 Самая высокая частота среди литовцев , значительная у латышей и эстонцев и меньшая частота у белорусов , украинцев , юго-западных русских и поляков . За исключением эстонцев , L1025 имеет самую высокую долю среди клад N-M231 в ранее упомянутых популяциях. Также наблюдается в Финляндии и Швеции , спорадические случаи – в Норвегии , Германии , Нидерландах , Великобритании , Азорских островах , Чехии и Словакии .
- N-L1025* Россия ( Курская область [ 1 ] )
- N-BY30389 Швеция ( лен Вестерноррланд) [ 1 ] ), Финляндия (провинция Западная Финляндия [ 1 ] )
- N-Y13982
- N-Y13982* Португалия ( Азорские острова [ 1 ] )
- N-Y31236 Литва ( Мариямпольский уезд , [ 1 ] Вильнюсский уезд [ 1 ] )
- Н-А11940
- N-Y140872 Швеция ( округ Сёдерманланд , [ 1 ] Лен Норрботтен [ 1 ] ), Финляндия (провинция Южная Финляндия [ 1 ] )
- N-Y143451 Russia ( Bashkortostan [ 1 ] )
- N-Y5580
- N-Y93996 Литва ( Шяуляйский уезд [ 1 ] ), Польша ( Подляское воеводство [ 1 ] )
- N-BY158 Россия ( Воронежская область , [ 1 ] Смоленская область , [ 1 ] Татарстан [ 1 ] ), Беларусь ( Витебская область [ 1 ] ), Литва ( Вильнюсский уезд , [ 1 ] Шяуляйский уезд , [ 1 ] Паневежский уезд [ 1 ] ), Польша (Подляское воеводство [ 1 ] ), Украина ( Полтавская область [ 1 ] ), Казахстан ( Жамбылская область [ 1 ] )
- Н-Z16975
- Н-ВЛ69 Беларусь ( Минская область) [ 1 ] ), Russia ( Bryansk Oblast [ 1 ] ), Kazakhstan ( Kostanay Region [ 1 ] )
- Н-Z16976
- N-Z16976* Belarus ( Grodno Region [ 1 ] ), Украина ( Khmelnytskyi Область [ 1 ] )
- N-Y13475 Польша, [ 1 ] Литва [ 1 ]
- N-Y21578 Литва ( Каунасский уезд , [ 1 ] Тельшяйский уезд [ 1 ] ), Россия ( Смоленская область [ 1 ] )
- N-Y6129 Польша ( Варминско-Мазурское воеводство [ 1 ] ), Литва [ 1 ]
- N-Y19113 Польша ( Варминско-Мазурское воеводство [ 1 ] ), Литва ( Таурагский уезд [ 1 ] )
- N-Y134492 Belarus ( Grodno Region , [ 1 ] Минская область , [ 1 ] Mogilev Region [ 1 ] ), Литва, [ 1 ] США ( Иллинойс [ 1 ] )
- Н-Л551
- N-L551* Литва ( Вильнюсский уезд) [ 1 ] )
- N-Y15251 Литва ( Алитусский уезд [ 1 ] ), Польша ( Подляское воеводство [ 1 ] )
- N-Y46313 Латвия [ 1 ]
- N-Y86578 Russia ( Belgorod Oblast [ 1 ] )
- N-Y14152
- N-BY21911 Польша, [ 1 ] Латвия ( Краславский район [ 1 ] ), Украина ( Киевская область [ 1 ] ), Финляндия (провинция Южная Финляндия [ 1 ] )
- N-Y13979 Литва, [ 1 ] Russia ( Ryazan Oblast [ 1 ] ), Германия [ 1 ]
- N-Y4706
- N-Y4706* Швеция (округ Стокгольм, [ 1 ] Лен Сёдерманланд, [ 1 ] Лен Вестра-Гёталанд [ 1 ] ), Германия ( Нижняя Саксония [ 1 ] ), Финляндия (провинция Южная Финляндия [ 1 ] ), Россия ( Татарстан [ 1 ] )
- N-A705 Швеция ( округ Кальмар , [ 1 ] Лен Эстергётланд [ 1 ] )
- N-BY21893 Польша ( Велькопольское воеводство) [ 1 ] )
- N-Y139030 Швеция ( округ Вестерботтен) [ 1 ] ), Норвегия ( Эстфолд [ 1 ] )
- N-Y183040 Russia ( Vologda Oblast , [ 1 ] Татарстан [ 1 ] )
- N-Y4707 Финляндия (провинция Западная Финляндия, [ 1 ] Аланд , [ 1 ] Южная Финляндия, [ 1 ] Восточная Финляндия, [ 1 ] Провинция Оулу [ 1 ] )
- N-Z16981
- N-A2358=N-BY14135 Англия, [ 1 ] Финляндия, [ 1 ] Латвия ( Даугавпилсский район [ 1 ] )
- N-CTS8173 Эстония ( Ляэне-Вирумаа [ 1 ] )
- N-FT213922 [ 59 ] )
- N-BY32524 Эстония ( Ярвамаа) [ 1 ] ), Финляндия (провинция Оулу [ 1 ] )
- N-FGC39882 Литва ( Тельшяйский уезд [ 1 ] )
- Н-FT6082 [ 59 ] )
- N-A11470 Россия ( Тюменская область [ 1 ] ), Литва ( Паневежский уезд [ 1 ] ), Финляндия (провинция Южная Финляндия [ 1 ] ), Нидерланды ( Южная Голландия , [ 1 ] Северный Брабант [ 1 ] )
- N-FTC49461 Латвия ( Яунпилсская волость, Тукумский район) [ 59 ] )
- N-FTE38475 Латвия ( Яунпилсская волость, Тукумский район) [ 59 ] )
- N-FTE39526 Латвия ( Яунпилсская волость, Тукумский район) [ 59 ] )
- N-FTE38475 Латвия ( Яунпилсская волость, Тукумский район) [ 59 ] )
- N-BY14147 Финляндия, Беларусь [ 59 ] )
- N-BY52958 Латвия [ 59 ] )
- N-FT109465 Латвия [ 59 ] )
- N-FTD92343 Латвия, Украина [ 59 ] )
- N-BY202983 Латвия [ 59 ] )
- N-BY40639 Латвия, Россия [ 59 ] )
- N-FTC49461 Латвия ( Яунпилсская волость, Тукумский район) [ 59 ] )
- N-BY21926 Россия, Латвия, Беларусь, Литва [ 59 ] )
- N-A11470 Россия ( Тюменская область [ 1 ] ), Литва ( Паневежский уезд [ 1 ] ), Финляндия (провинция Южная Финляндия [ 1 ] ), Нидерланды ( Южная Голландия , [ 1 ] Северный Брабант [ 1 ] )
- N-Y15922=N-Z35238 Финляндия (провинция Южная Финляндия) [ 1 ] ), Эстония ( Ида-Вирумаа , [ 1 ] Пярнумаа [ 1 ] ), Латвия ( Валмиера , [ 1 ] Лиепая , [ 1 ] Талсинский уезд [ 1 ] ), Россия ( Псковская область [ 1 ] ), Польша ( Поморское воеводство [ 1 ] )
- N-Y6075 Польша ( Малопольское воеводство , [ 1 ] Мазовецкое воеводство [ 1 ] ), Украина ( Львовская область [ 1 ] ), Словакия, [ 1 ] Чехия ( Моравскосилезский край) [ 1 ] ), США ( Нью-Джерси [ 1 ] )
- N-Y11882
- N-Y11882* Россия (Татарстан [ 1 ] )
- N-ZS11617 Russia ( Yaroslavl Oblast [ 1 ] ), Литва [ 1 ]
- N-Y24601 Латвия [ 1 ]
- N-Y94659 Литва, [ 1 ] Беларусь ( Минская область [ 1 ] )
- N-Y32725
- N-FT96305 Латвия ( Краславский район [ 1 ] ), Эстония ( Вильяндимаа [ 1 ] )
- N-Y33333 Ukraine ( Sumy Oblast [ 1 ] ), Беларусь ( Брестская область [ 1 ] )
- N-FT213922 [ 59 ] )
- N-Y46443
- N1a1a1a1a1a2-CTS9976
- Н-L1022
- N-Y19098
- N-Y19098* Швеция ( округ Вестерноррланд) [ 1 ] )
- N-A12258 Финляндия (провинция Восточная Финляндия) [ 1 ] )
- N-BY117178 Финляндия (провинция Восточная Финляндия) [ 1 ] )
- N-Y5004
- Н-Y7300
- N-A17632
- N-Y128024 Швеция ( округ Эстергётланд) [ 1 ] )
- N-A16017
- N-Y7297 Финляндия (провинция Западная Финляндия, [ 1 ] Южная Финляндия, [ 1 ] Лапландия [ 1 ] )
- N-Y15813
- N-Y15813* Финляндия (провинция Западная Финляндия) [ 1 ] )
- N-Y49008 Финляндия (провинция Западная Финляндия, [ 1 ] Провинция Оулу [ 1 ] )
- N-Y46886 Финляндия (провинция Западная Финляндия) [ 1 ] )
- N-Y15812
- N-A14187 Финляндия (провинция Западная Финляндия) [ 1 ] ), Эстония ( Ляэнемаа [ 1 ] )
- N-Y24617 Финляндия (провинция Западная Финляндия) [ 1 ] ), Швеция ( округ Даларна [ 1 ] ), Россия ( Курская область [ 1 ] )
- N-Y23576
- N-Y23576* Финляндия (провинция Восточная Финляндия) [ 1 ] )
- N-Y15615
- N-A17632
- N-Y5005
- N-Y22106
- N-Y47789 Финляндия (провинция Западная Финляндия) [ 1 ] )
- N-Y79341 Финляндия (провинция Западная Финляндия) [ 1 ] ), Швеция ( округ Вестра-Гёталанд , [ 1 ] Лен Вермланд [ 1 ] )
- N-Y10756
- N-A13656 Финляндия (провинция Западная Финляндия, [ 1 ] Лапландия [ 1 ] ), Швеция ( округ Даларна [ 1 ] )
- Н-PH2196
- N-CTS11122 Финляндия (провинция Восточная Финляндия) [ 1 ] )
- N-A17277 Финляндия (Западная Финляндия, [ 1 ] Южная Финляндия [ 1 ] )
- N-PH547 Финляндия (Лапландия, [ 1 ] Восточная Финляндия, [ 1 ] Южная Финляндия [ 1 ] ), Швеция ( графство Вестерботтен , [ 1 ] Лен Норрботтен [ 1 ] )
- N-Y5003
- N-Y5003* Эстония ( Хийумаа [ 1 ] ), Финляндия (провинция Южная Финляндия [ 1 ] ), Великобритания ( Вестминстер [ 1 ] )
- N-BY22001 Эстония ( Ляэнемаа [ 1 ] ), Финляндия (провинция Западная Финляндия [ 1 ] ), Швеция (округ Норботтен [ 1 ] )
- N-BY6007 Россия ( Липецкая область [ 1 ] )
- N-Y132182 Швеция ( округ Вермланд) [ 1 ] ), Эстония ( Вильяндимаа [ 1 ] )
- N-Y20917 Швеция ( округ Кроноберг) [ 1 ] ), Финляндия (Западная Финляндия, [ 1 ] Южная Финляндия [ 1 ] )
- N-Y24502 Великобритания ( Шотландские границы , [ 1 ] Колдердейл , [ 1 ] Кирклис [ 1 ] )
- N-Y18420 Финляндия (провинция Южная Финляндия, [ 1 ] Западная Финляндия [ 1 ] )
- N-Z35267 Финляндия (Западная Финляндия, [ 1 ] Аландская провинция , [ 1 ] Южная Финляндия [ 1 ] ), Эстония ( Пярнумаа [ 1 ] )
- N-Y6599 Финляндия (провинция Восточная Финляндия, [ 1 ] Провинция Оулу, [ 1 ] Южная Финляндия [ 1 ] ), Эстония ( Йыгевамаа , [ 1 ] Хийумаа [ 1 ] ), Россия ( Тульская область [ 1 ] )
- Н-Y24000
- N-Y16503 Финляндия (провинция Западная Финляндия, [ 1 ] Южная Финляндия, [ 1 ] Восточная Финляндия, [ 1 ] Провинция Оулу [ 1 ] ), Швеция ( графство Евлеборг [ 1 ] )
- N-Y22106
- Н-Y7300
- N-Y19098
- N-BY6010 США ( Вирджиния) [ 1 ] )
- Н-CTS3451
- N-CTS3451* Русский [ 1 ]
- N-Y3667
- N-BY33087
- N-BY33087* Финляндия ( провинция Южная Финляндия) [ 1 ] )
- N-BY33088 Финляндия ( провинция Восточная Финляндия) [ 1 ] ), Россия ( Новгородская область [ 1 ] )
- Н-CTS657
- N-CTS657* Финляндия, [ 1 ] Россия ( Республика Карелия , [ 1 ] Новгородская область [ 1 ] )
- N-BY70437 Финляндия (провинция Восточная Финляндия) [ 1 ] )
- N-BY6024 Финляндия (провинция Восточная Финляндия) [ 1 ] )
- N-Y45925 Россия (Республика Карелия) [ 1 ] )
- N-Y26750
- N-Y26750* Russia ( Arkhangelsk Oblast [ 1 ] )
- N-A16653 Финляндия (провинция Восточная Финляндия) [ 1 ] )
- N-PH3568 Финляндия ( провинция Оулу , [ 1 ] Восточная Финляндия [ 1 ] )
- N-BY33087
- Н-L1022
- N1a1a1a1a1a1-Z4908
- N1a1a1a1a1b-PH1266/Y28526/F4134
- N-Y46746 Россия [ 1 ]
- N-Y32732
- N-Y32732* Россия ( Санкт-Петербург [ 1 ] )
- N-Y192174
- N-Y32731 Россия ( Республика Коми [ 1 ] ), Швеция [ 1 ]
- N1a1a1a1a1c-B479 Нанай , [ 3 ] Ульчи [ 40 ]
- N1a1a1a1a1a-CTS2929/VL29 Встречается с высокой частотой среди литовцев , латышей , эстонцев северо-запада , русских , шведских саамов , карел , ненцев , финнов и марийцев , умеренной частотой среди других русских , белорусов , украинцев и поляков и низкой частотой среди коми , Мордва , татары , чуваши , долганы , вепсы , селькупы , караногайцы и башкиры. [ 3 ]
- N1a1a1a1a2-Z1936,CTS10082 Встречается с высокой частотой среди финнов , вепсов , карел , шведских саамов , северо-западных русских , башкир и волжских татар , умеренная частота среди других русских , коми , ненцев , обских угров , долган и сибирских татар , и низкая частота среди мордвы , нганасан , чувашей , эстонцев , латышей , украинцев и караногаев [ 3 ]
- N-Y13851
- N-BY199053 Франция, [ 59 ] ancient Western Siberia (Gornaya Bitiya, Iron Age Sargat culture , 300 - 100 BCE [ 59 ] )
- N-Y13852
- N1a1a1a1a2a1c-PH3340/Y13850
- N1a1a1a1a2a1c1-L1034
- N-Y28538
- N-Y28538* Россия ( Хантия-Мансия [ 1 ] )
- N-L1032 Россия (Хантия-Мансия [ 1 ] ), Казахстан ( Восточно-Казахстанская область [ 1 ] )
- Н-L1442
- Н-FT12605
- N-Y23732 Russia ( Bashkortostan [ 1 ] )
- N-Y24222
- N-Y24222* Венгрия [ 1 ]
- N-Y65329 Россия (Татарстан татарский , [ 1 ] Bashkortostan [ 1 ] )
- N-Y62987 Russia (Bashkortostan, [ 1 ] Самарская область , [ 1 ] Тюменская область Сибирский татарин [ 1 ] )
- N-Y28538
- N1a1a1a1a2a1c2-Y24361 (Y24372)
- N1a1a1a1a2a1c1-L1034
- N1a1a1a1a2a1c-PH3340/Y13850
- Н1а1а1а1а2а-З1934
- N-Y159520
- N-Y159520* Russia ( Tambov Oblast [ 1 ] )
- N-BY184755 Россия ( Новгородская область , [ 1 ] Воронежская область , [ 1 ] Москва [ 1 ] )
- N-Y18421
- N-Y18421* Russia (Tambov Oblast [ 1 ] )
- N-Y19110
- N-Y180247
- N-Y19108
- N-Y19108* Россия (Татарстан, [ 1 ] Kirov Oblast [ 1 ] )
- N-Y19453
- N-Y19453* Россия (Татарстан [ 1 ] )
- N-Y19451 Финляндия (провинция Западная Финляндия) [ 1 ] ), Россия ( Республика Карелия , [ 1 ] Tver Oblast [ 1 ] )
- Н-З1928/CTS2733
- N-YP6091
- N-YP6091* Россия ( Иркутская область [ 1 ] )
- N-YP6094
- N-Y129131
- N-Y129131* Russia ( Kostroma Oblast [ 1 ] )
- N-A25107 Россия ( Кировская область , [ 1 ] ВВП России [ 1 ] )
- N-YP6092
- N-YP6092* Russia ( Vologda Oblast [ 1 ] )
- Н-Б195
- N-B195* Russia ( Sverdlovsk Oblast [ 1 ] )
- N-Y62142 Россия ( Республика Карелия , [ 1 ] Архангельская область [ 1 ] )
- N-Y129131
- N1a1a1a1a2a-Z1925
- N-Z1925* Швеция ( округ Норрботтен) [ 1 ] )
- N-Y29767 Швеция ( округ Норрботтен) [ 1 ] )
- N-Y62904 Финляндия ( провинция Западная Финляндия , [ 1 ] Провинция Оулу [ 1 ] )
- N1a1a1a1a2a2a1a1-Z1926
- N-Y20920
- N-Y20920* Финляндия ( провинция Западная Финляндия) [ 1 ] ), Норвегия ( Норд-Трёнделаг [ 1 ] )
- N-Y21699 Финляндия ( провинция Западная Финляндия , [ 1 ] Южная Финляндия [ 1 ] )
- Н-З1927
- N-Y22108 Финляндия (провинция Восточная Финляндия, [ 1 ] Южная Финляндия, [ 1 ] Западная Финляндия [ 1 ] ), Швеция ( графство Стокгольм , [ 1 ] Округ Эребру [ 1 ] )
- Н-З1933
- Н-CTS4329
- N-Y22091
- N-Y22091* Норвегия ( Нордланд [ 1 ] ), Финляндия ( Провинция Лапландия [ 1 ] )
- N-Y47623
- N-Y31247
- N-Y31247* Russia ( Tver Oblast [ 1 ] )
- N-Y31249
- N-Y90283 Финляндия (провинция Лапландия) [ 1 ] )
- N-Y31244 Финляндия ( провинция Западная Финляндия , [ 1 ] Восточная Финляндия [ 1 ] )
- N-YP6269
- N-YP6269* Финляндия (провинция Западная Финляндия) [ 1 ] )
- N-Y151660
- N-Y151660* Финляндия (провинция Восточная Финляндия) [ 1 ] )
- N-BY149208 Швеция ( округ Норрботтен) [ 1 ] )
- Н-CTS8565
- N-BY18114 Финляндия (провинция Восточная Финляндия) [ 1 ] )
- N-BY30394 Финляндия (провинция Восточная Финляндия) [ 1 ] ), Россия ( Удмуртская Республика [ 1 ] )
- N-Y30513
- N-Y30513* Финляндия ( провинция Южная Финляндия) [ 1 ] )
- N-Y29759
- Н-Z4998
- N-Z4998* Финляндия (провинция Южная Финляндия) [ 1 ] )
- N-FGC65190 Финляндия (провинция Южная Финляндия, провинция Восточная Финляндия)
- N-Y18414 Финляндия (провинция Западная Финляндия)
- N-Y20910 Финляндия (провинция Западная Финляндия)
- N-BY194138 Финляндия (провинция Южная Финляндия)
- N-Y28547 Финляндия (провинция Восточная Финляндия)
- N-BY22141 Финляндия (провинция Западная Финляндия, провинция Восточная Финляндия), США (Вашингтон)
- N-FT5834 Финляндия (Западная Финляндия, Южная Финляндия)
- N-BY190112 Финляндия (провинция Западная Финляндия, провинция Южная Финляндия)
- N-Y19097 Финляндия (провинция Восточная Финляндия, провинция Южная Финляндия, провинция Западная Финляндия, провинция Оулу)
- N-CTS3223 Финляндия (Южная Финляндия, Восточная Финляндия, Западная Финляндия, Оулу, Лапландия), Швеция (Норрботтен, Вестерботтен, Даларна, Вермланд, Сконе), Россия ( Ленинградская область )
- N-Y22091
- Н-ВЛ62 Россия ( Алтайский край , [ 1 ] Костромская область [ 1 ] ), Эстония ( Харьюмаа [ 1 ] ), Финляндия [ 1 ]
- N-VL62* Russia ( Chelyabinsk Oblast [ 1 ] ), Финляндия [ 1 ]
- Н-ВЛ60
- N-VL60* Russia ( Kurgan Oblast [ 1 ] )
- N-Y63781 Россия ( Владимирская область , [ 1 ] Архангельская область [ 1 ] )
- Н-ВЛ72
- Н-ВЛ72* Россия ( Республика Карелия [ 1 ] )
- N-BY30470 Финляндия ( провинция Западная Финляндия) [ 1 ] )
- N-Y20915 Финляндия ( провинция Восточная Финляндия) [ 1 ] )
- Н-З1939
- N-Y132561
- Н-Z2445
- N-Z35031 Финляндия (провинция Восточная Финляндия, провинция Западная Финляндия), Швеция (лен Вестра-Гёталанд, округ Вестманланд)
- N-BY28931
- Н-CTS7189
- N-BY142665 Финляндия (провинция Восточная Финляндия) [ 1 ] )
- N-Y37149 Финляндия (провинция Восточная Финляндия) [ 1 ] )
- N-Y23568 Финляндия (провинция Восточная Финляндия) [ 1 ] )
- N-Y80226 Финляндия (провинция Восточная Финляндия) [ 1 ] )
- N-FT20730 Финляндия (провинция Восточная Финляндия) [ 1 ] ), Россия (Республика Карелия [ 1 ] )
- N-BY62666 Финляндия (провинция Восточная Финляндия) [ 1 ] ), Норвегия (Осло [ 1 ] ), США (Вирджиния, [ 1 ] Арканзас [ 1 ] )
- N-Y24218
- N-BY22038 Финляндия (провинция Восточная Финляндия, [ 1 ] Южная Финляндия, [ 1 ] Западная Финляндия [ 1 ] )
- N-Y13974 Финляндия (провинция Южная Финляндия, [ 1 ] Восточная Финляндия, [ 1 ] Провинция Оулу, [ 1 ] Западная Финляндия [ 1 ] )
- N-Y135981 Финляндия (провинция Восточная Финляндия, [ 1 ] Провинция Оулу, [ 1 ] Южная Финляндия, [ 1 ] Западная Финляндия [ 1 ] ), Норвегия ( Финнмарк [ 1 ] )
- Н-Z4747
- N-Y17790 Финляндия (провинция Восточная Финляндия, [ 1 ] Южная Финляндия, [ 1 ] Западная Финляндия [ 1 ] )
- Н-З1941
- N-BY22090 Финляндия (провинция Южная Финляндия, [ 1 ] Восточная Финляндия [ 1 ] )
- N-Y21575 Финляндия (провинция Западная Финляндия, [ 1 ] Южная Финляндия, [ 1 ] Восточная Финляндия, [ 1 ] Провинция Оулу, [ 1 ] Лапландия [ 1 ] ), Россия ( Ленинградская область , [ 1 ] Москва , [ 1 ] Татарстан [ 1 ] )
- N-Z1940 Финляндия (Западная Финляндия, [ 1 ] Южная Финляндия, [ 1 ] Восточная Финляндия, [ 1 ] Провинция Оулу, [ 1 ] Лапландия [ 1 ] ), Россия (Ленинградская область [ 1 ] ), Швеция (лен Вермланд [ 1 ] )
- Н-CTS4329
- N-Y20920
- N-YP6091
- N-Y159520
- N-Y13851
- N1a1a1a1a3-B197/Y16323
- N1a1a1a1a3a-F4205 Часто встречается среди бурят. [ 3 ] и Цаатаны , [ 28 ] умеренная частота среди караногаев , [ 3 ] Тувинцы , [ 3 ] тоджинс , [ 28 ] и монголы , [ 3 ] [ 28 ] и низкая частота среди алтайцев , [ 3 ] сибирские татары , [ 3 ] Казахи , [ 3 ] Эвенки , [ 3 ] крымские татары , [ 3 ] Каракалпаки , [ 3 ] Узбеки , [ 3 ] и украинцы [ 3 ]
- N-F4205* Китай (Внутренняя Монголия, Ганьсу, Синьцзян, Хэнань и т. д.) [ 75 ] )
- N-Y16221
- N-Y16221* Китай ( Внутренняя Монголия) [ 1 ] )
- N-Y16220
- N-Y16220* Украина ( Луганская область [ 1 ] )
- N-F22331 Турция ( Мерсин [ 1 ] ), Польша ( Малопольское воеводство [ 1 ] ), Kazakhstan ( Pavlodar Region ca. 1680 ybp [ 1 ] )
- N-Y63966 Казахстан, [ 1 ] Турция, [ 1 ] Узбекистан [ 1 ]
- N-Y226011
- N-Z35331 Китай ( город Ордос , Хайдун , Чэнду) [ 75 ] )
- Н-Б199 Россия (Бурятия) [ 1 ] ), Китай (Хуасай из Хулунбуира [ 75 ] )
- N-FT411781 Китай (Внутренняя Монголия) [ 1 ] )
- N-Y16312/F2288
- N-Z35331 Китай ( город Ордос , Хайдун , Чэнду) [ 75 ] )
- N1a1a1a1a3b-B202 Встречается с высокой частотой среди чукчей , [ 3 ] Коряки , [ 3 ] и сибирские эскимосы [ 3 ]
- N1a1a1a1a3a-F4205 Часто встречается среди бурят. [ 3 ] и Цаатаны , [ 28 ] умеренная частота среди караногаев , [ 3 ] Тувинцы , [ 3 ] тоджинс , [ 28 ] и монголы , [ 3 ] [ 28 ] и низкая частота среди алтайцев , [ 3 ] сибирские татары , [ 3 ] Казахи , [ 3 ] Эвенки , [ 3 ] крымские татары , [ 3 ] Каракалпаки , [ 3 ] Узбеки , [ 3 ] и украинцы [ 3 ]
- N1a1a1a1a1-CTS10760
- N1a1a1a1b-M2019/M2118 Эстония, [ 1 ] [ 94 ] Китай ( Хами , Яньань , Сиань , Люлян , Чанчжи , Лига Силингол , Чифэн , Цанчжоу , Сунъюань [ 75 ] )
- N-M2058 Китай (Шаньдун, Ляонин, Хэнань, Внутренняя Монголия, Ганьсу и т. д.) [ 75 ] [ 1 ] )
- N-M2016 Китай (особенно Синьцзян и Внутренняя Монголия). [ 75 ] )
- N1a1a1a1b2-A9408 Китай (Хэбэй, Шаньдун, Хэнань и др.) [ 75 ] ), Ливан [ 53 ] [ 3 ]
- N-Y70200 Китай [ 1 ] (Шэньси, Шаньси, Шаньдун и т. д. [ 75 ] )
- N-PH1612 Китай ( Юлинь , Шанцю , Дэчжоу , Таншань , Шэньян [ 75 ] )
- N-A9407/A9411 China ( Ili Kazakh Autonomous Prefecture [ 75 ] )
- N-A9416 Венгрия ( Сабольч-Сатмар-Берег [ 1 ] ), Хорватия ( Задарская жупания [ 1 ] ), Россия ( Чувашская Республика [ 1 ] )
- N-PH1896 Венгрия ( Боршод-Абауй-Земплен [ 1 ] ), Турция ( Хатай [ 1 ] [ 94 ] ), Китай (Синьцзян, Шаньдун, Аньхой, Хэйлунцзян, Хэбэй, Нинся, Гуйчжоу [ 75 ] )
- N-A9407/A9411 China ( Ili Kazakh Autonomous Prefecture [ 75 ] )
- N-M2058 Китай (Шаньдун, Ляонин, Хэнань, Внутренняя Монголия, Ганьсу и т. д.) [ 75 ] [ 1 ] )
- N1a1a1a1a-L392
- Н1а1а1а2-Б211 (Y9022) Удмуртский , [ 3 ] Приходить , [ 3 ] Чуваш , [ 3 ] Ob-Ugrians , [ 3 ] Большой , [ 3 ] Мордовия , [ 3 ] Алтай , [ 3 ] Белорусский , [ 3 ] Караногай , [ 3 ] карельский , [ 3 ] Башкирский , [ 3 ] татарин , [ 94 ] Русский , [ 3 ] [ 94 ] хакасы [ 3 ]
- N1a1a1a2a-B181
- N-Y182299 Россия ( Кировская область и др.) [ 1 ]
- N-Y23788 Russia ( Bashkortostan , Komi Republic , etc.) [ 1 ]
- N1a1a1a2b-Y23181
- N-Y23183 Russia ( Tatarstan , Mordovia , Penza Oblast ) [ 1 ]
- N-Y143277 Russia ( Tatarstan , Bashkortostan ) [ 1 ]
- N1a1a1a2a-B181
- N1a1a1a1-P298/M2126 Китай ( Урумчи , префектура Кашгар , Турфан , префектура Аксу , Сяньян , Цзиньчэн , Кайфэн , Цицикар [ 75 ] )
- N-Y24317
- N1a1a1a-L708
- N1a1a2-Y23747 Япония ( остров Садо [ 94 ] )
- N1a1a2a-Y23749 Япония [ 3 ] ( Айти , [ 1 ] Хоккайдо [ 1 ] )
- N1a1a2b-Y125664
- N-Y125664* Шанхай [ 1 ]
- N-MF16376
- N1a1a2b1-MF38607 Аньхой , [ 1 ] Себя [ 1 ]
- N1a1a2b2-F22150 Хэбэй , [ 1 ] Сучжоу [ 75 ]
- N-FT281705 Китай (Шаньдун, Ляонин и др.) [ 75 ] )
- N-MF15288/MF15344 Китай (особенно Цзянсу, но также Хэнань, Фуцзянь, Шанхай, Хэйлунцзян , [ 1 ] и т. д. [ 75 ] ), Южная Корея [ 1 ]
- N-MF55680 Китай (Хэбэй, Пекин, Хубэй и т. д.) [ 75 ] )
- N1a1a1-F1419
- Н1а1а-М178
- Н-F1360
- N-F1360* Китай ( Шэньси [ 1 ] )
- Н1а2-Ф1008/Л666
- Н-F1101
- N-F1101* Китай ( Шаньдун [ 1 ] )
- Н-F1154
- N-F1154* Китай ( Ляонин [ 1 ] )
- N-Y23741
- N-Y23741* Китай [ 1 ]
- Н1а2а-М128
- N-M128* Япония ( Фукуи ) [ 1 ]
- N-Y23738 Вьетнам [ 1 ]
- Н-CTS1350
- N1a2b-B523(P43)
- N1a2b-Y126204, ВЛ67/З35079, BY29083
- Н-ВЛ67/Z35079
- N1a2b1-B478 (P63) Ненцы, нганасаны, долганы, тувинцы, тоджинцы, [ 28 ] Хакасы, шорцы, эвены, алтайцы, селькупы, эвенки, монголы (сарт-калмак, [ 28 ] Торгут, [ 28 ] дерби, [ 28 ] Buryat), Yakuts, Ob-Ugrians, Chuvashes
- N1a2b1a-B168 Эвены
- N1a2b1b-B169
- N1a2b1b1-B170 Nenets , Khanty-Mansi Autonomous Okrug , Inner Mongolia
- N1a2b1b2-B175 Tuvinians , Mongols , Evenks , Yakuts , Tomsk Oblast
- N1a2b3-B525 Турция , татары , башкиры , казахи , монголы (Синьцзян-калмыки, [ 28 ] Монгольский Торгут [ 28 ] ), Словакия , Болгария , [ 1 ] украинцы , белорусы , русские
- N-B525* Афганистан , арабский
- N-BY173686 Россия ( Воронежская область [ 1 ] )
- N-VL83 Турция [ 1 ]
- N-Y136502 Россия ( Татарстан [ 1 ] ), Болгария ( Добрич [ 1 ] )
- N-Y37153
- N-Y37153* Россия ( Татарстан [ 1 ] )
- N-BY29131
- N-BY29131* Turkey ( Konya [ 1 ] ), Словакия ( Жилинский край [ 1 ] )
- N-BY30476 Россия ( Татарстан , [ 1 ] Пензенская область [ 1 ] )
- N1a2b1-B478 (P63) Ненцы, нганасаны, долганы, тувинцы, тоджинцы, [ 28 ] Хакасы, шорцы, эвены, алтайцы, селькупы, эвенки, монголы (сарт-калмак, [ 28 ] Торгут, [ 28 ] дерби, [ 28 ] Buryat), Yakuts, Ob-Ugrians, Chuvashes
- N1a2b2-FGC10872/Y3195
- N1a2b2a-FGC10847/Y3185 (L1419) Вепсы , марийцы , русские ( Архангельская область ), коми , Пермский край , Республика Коми , обские угры , чуваши , татары , башкиры , карелы , Западно-Финляндская губерния , тувинцы , буряты , хакасы , нганасаны , Азиатские эскимосы
- N1a2b2b-Y23786
- N1a2b2b* Оставайся
- N1a2b2b1-B528/Y24384 Удмурты , коми , ханты , татары , азиатские эскимосы, Кировская область , Пермский край , остров Медный
- Н-ВЛ67/Z35079
- N1a2b-Y126204, ВЛ67/З35079, BY29083
- Н-F1101
- Н-З1956
- N1b-F2905
- N1b1-CTS582 Китай (особенно Цзянсу, Юньнань, Хэбэй, Шаньдун, Шаньси, Хэйлунцзян [ 75 ] )
- N1b1a-Y6374/Z8029
- N1b1a1-CTS7324 Пекин [ 1 ]
- N1b1a2-L727
- N1b1a2*-L727 Пекин [ 1 ]
- N1b1a2a-L732 Беларусь [ 59 ]
- N1b1a2a1-F839 Китай (особенно Цзянсу, [ 75 ] Шаньдун, [ 75 ] Чжэцзян, [ 75 ] Шанхай, [ 1 ] [ 75 ] Хэнань [ 75 ] )
- N1b1a2a2-Y15972
- N1b1b-Y23789/CTS4309 Ирак [ 59 ]
- N1b1a-Y6374/Z8029
- N1b2-M1819/N-M1897/CTS12473/F1173 Китай, [ 59 ] Российская Федерация [ 59 ]
- N-M1897* древняя ДНК из Пинлянтай , Хэнань (ок. 4063 [95% ДИ 3974 <-> 4151] лет назад) [ 1 ]
- Н-М1845
- N-M1845* Китай (Сычуань, [ 1 ] Шанхай [ 1 ] )
- Н-М1928
- N-M1928* Сычуань ( Хань ) [ 1 ]
- N-Y125475
- N-Y63516 Китай ( Чунцин [ 1 ] ), Корея ( провинция Южный Пхеньян , [ 1 ] Провинция Южный Чхунчхон [ 1 ] )
- N-Y193396 Китай ( Нинся [ 1 ] ), Сингапур [ 1 ]
- N-CTS4714
- N1b1-CTS582 Китай (особенно Цзянсу, Юньнань, Хэбэй, Шаньдун, Шаньси, Хэйлунцзян [ 75 ] )
- N1a-L729
- N2-Y6503
- N2-Y6503* Республика Алтай [ 1 ]
- N2a-P189.2
- N2a1-Y6516
- N2a1-Y6516*
- N2a1a-Y7310
- N2a1a-Y7310* Румыния ( венгр из Сучавы ) [ 1 ]
- N2a1a1-Y7313
- N2a1a1-Y7313*
- N2a1a1a-BY35494 Россия ( Ростовская область ) [ 1 ]
- N2a1a1b-FGC28435
- N2a1a1b-FGC28435* Турция (Стамбул), Сербия, Черногория (Плужине), Босния и Герцеговина ( Республика Сербская ), Хорватия [ 1 ]
- N2a1a1b1-FGC28483 Сербия [ 1 ]
- N2a2-Y111068
- N2a2a-FT352925 Франция ( Приморская Шаранта [ 1 ] ), Турция [ 1 ]
- N2a2b-Y101945 Великобритания ( Девон [ 1 ] ), Россия ( Московская область [ 1 ] )
- N2a1-Y6516
- N1-Z4762/CTS11499/L735/M2291
- Н-М231/Пейдж91, М232/М2188
История филогенетической номенклатуры
[ редактировать ]До 2002 года в научной литературе существовало как минимум семь систем наименования филогенетического дерева Y-хромосомы. Это привело к значительной путанице. В 2002 году основные исследовательские группы объединились и сформировали Консорциум Y-хромосомы (YCC). Они опубликовали совместный документ, в котором было создано единое новое дерево, которое все согласились использовать. Позже группа гражданских ученых, интересующихся популяционной генетикой и генетической генеалогией, сформировала рабочую группу для создания любительского дерева, стремясь быть прежде всего своевременным. В таблице ниже собраны все эти работы, относящиеся к знаковому дереву YCC 2002 года. Это позволяет исследователю, просматривающему старую опубликованную литературу, быстро перемещаться между номенклатурами.
| YCC 2002/2008 (сокращенно) | (а) | (б) | (с) | (г) | (е) | (г) | (или) | YCC 2002 (от руки) | YCC 2005 (от руки) | YCC 2008 (от руки) | YCC 2010r (от руки) | ИСОГГ 2006 г. | ИСОГГ 2007 | ИСОГГ 2008 | ИСОГГ 2009 | ИСОГГ 2010 | ИЗОГГ 2011 | ИЗОГГ 2012 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Н-LLY22g | 12 | VIII | 1U | 25 | ЕС16 | Н5 | Ф | Н* | Н | N1 | N1 | - | - | - | - | - | - | - |
| Н-М128 | 12 | VIII | 1U | 25 | ЕС16 | Н5 | Ф | N1 | N1 | Н1а | Н1а | - | - | - | - | - | - | - |
| Н-П63 | 12 | VIII | 1U | 25 | ЕС16 | Н5 | Ф | Н2 | Н2а | N1b1 | N1b1 | - | - | - | - | - | - | - |
| Н-ТАТ | 12 | VIII | 1я | 26 | ЕС13 | Н5 | Ф | N3* | N3 | N1c | N1c | - | - | - | - | - | - | - |
| Н-М178 | 16 | VIII | 1я | 26 | ЕС14 | Н5 | Ф | Н3а* | М178 | N1c1 | N1c1 | - | - | - | - | - | - | - |
| Н-П21 | 16 | VIII | 1я | 26 | ЕС14 | Н5 | Ф | N3a1 | N3a1 | N1c1a | N1c1a | - | - | - | - | - | - | - |
Источники Следующие исследовательские группы в соответствии с их публикациями были представлены в создании Дерева YCC.
Недостоверные мутации (SNP и UEP)
Делеция b2/b3 в регионе AZFc Y-хромосомы, по-видимому, произошла независимо, по крайней мере, в четырех разных случаях. Следовательно, это удаление не следует воспринимать как уникальный полиморфизм событий, определяющий эту ветвь дерева Y-хромосомы ( ISOGG 2012 ).
Ссылки на концепции генетики
[ редактировать ]- генетическая генеалогия
- Генетическая история Европы
- Гаплогруппа
- Гаплотип
- Гаплогруппа ДНК Y-хромосомы человека
- молекулярная филогения
- Парагруппа
- Субклад
- Гаплогруппы Y-хромосомы в популяциях мира
- Гаплогруппы Y-ДНК по этническим группам
- Гаплогруппы Y-ДНК в популяциях Восточной и Юго-Восточной Азии
- Гаплогруппы Y-ДНК в населении Европы
Субклады Y-ДНК N
[ редактировать ]- Н-М231
Основополагающее дерево Y-ДНК
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также и аль являюсь а к ап ак с как в В из хорошо топор является тот нет бб до нашей эры др. быть парень бг чб с минет БК с бм млрд быть б.п. БК бр бс БТ этот бв б бх к бз что КБ копия компакт-диск Этот см. cg ч Там СиДжей ск кл см CN со КП cq кр CS КТ с резюме cw сх сай чешский и БД округ Колумбия дд из дф дг д Из диджей дк дл дм дн делать дп дк доктор дс дт из дв ДВ дх ты дз из Эб ЕС Эд да если например ага нет ej я он в в там эп. экв. является является И Евросоюз этот Вон тот бывший эй нет но ФБ ФК ФД фе фф фг фх быть фджей хз в фм фн фо фп fq фр. фс футы это было фв фу валюта мой фз га ГБ ГК Боже ге подруга gg хх ги джей ГК гл гм гн идти врач gq гр гс гт к гв гв gx гы гз ха чб хк HD он хф рт.ст. хх привет дрочка хк гл хм хн к л.с. штаб-квартира час хз хт ху что как хх он хз это а IC идентификатор т.е. если ig ih ii ij я il в в этот IP iq и является это я iv я ix запах iz и джб Джей Си Джей Ди является JF джг джх из Джей-Джей джк джл Джейм джин потому что Япония jq младший js ДжТ верно СП jw jx ты джз тот КБ кс кд тот кф кг х к кДж ок в км знать является КП кг ДКК кс КТ к кв кВт кх этот КЗ тот фунт ЖК лд тот лф LG левый что жж lk я буду лм Ин это лп лк лр лс лт ему лв лв лк ли лз и мб MC Мэриленд мне мф мг мх мне ЭмДжей мк мл мм минута для депутат кв.м. Мистер РС тонна в МВ МВт мх мой мз уже обратите внимание NC nd ne нф из нет в Нью-Джерси такой как Нидерланды нм пп нет например нк Нет. нс нт нет нв теперь нх тот Новая Зеландия из ob ок из ты из и ой привет ой хорошо ол о на и на белый или ты ot или ов ой бык ООО унция хорошо пб ПК ПД на пф стр. тел. пи пиджей ПК пожалуйста вечер пн po пп ПК пиар пс пт мог пв пш пикселей пи пз ка qb контроль качества qd ЧТО qf qg qh ци qj кк ql кв.м. qn qo qp qq qr вопросы qt что кв qw qx куй девочка день РБ RC р-д ре рф рг резус ри РДжей рк рл РМ р-н ро рп rq рр РС рт ру фургон RW прием рай рз на сб СК SD с НФ сг ш и сж ск сл см зп так сп кв. сэр SS ул. являются св SW сх и нет лицом к лицу туберкулез ТК ТД тот ТС тг й из тиджей ТС ТЛ тм ТН к город tq тр тс тт что телевизор дважды Техас ty тс делать уб Калифорнийский университет вне уэ уф и Эм-м-м пользовательский интерфейс удж Великобритания ул один и друзья вверх UQ ты YПолная гаплогруппа YTree v6.05.11 от 25 сентября 2018 г.
- ^ Перейти обратно: а б с Позник Г.Д., Сюэ Ю., Мендес Ф.Л., Виллемс Т.Ф., Массайя А., Уилсон Сейрес М.А. и др. (июнь 2016 г.). «Периодические всплески демографии мужского пола, выявленные на основе 1244 последовательностей Y-хромосомы по всему миру» . Природная генетика . 48 (6): 593–599. дои : 10.1038/ng.3559 . ПМЦ 4884158 . ПМИД 27111036 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также и аль являюсь а к ап ак с как в В из хорошо топор является тот нет бб до нашей эры др. быть парень бг чб с минет БК с бм млрд быть б.п. БК бр бс БТ этот бв б бх к бз Илумяэ и др. 2016 год
- ^ Перейти обратно: а б с д и ISOGG, 2016, Гаплогруппа N Y-ДНК и ее субклады – 2016, 22 августа 2016 г.).
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с Роотси, Сиири [на эстонском языке] ; Животовский Лев А; Бальдович, Мариан; Кайзер, Манфред; и др. (06 декабря 2006 г.). «Северный маршрут гаплогруппы N Y-хромосомы против часовой стрелки из Юго-Восточной Азии в Европу» (PDF) . Европейский журнал генетики человека . 15 (4): 204–211. дои : 10.1038/sj.ejhg.5201748 . ПМИД 17149388 . S2CID 19265287 .
- ^ Перейти обратно: а б с д и ж г час Карафет Т.М., Осипова Л.П., Савина О.В., Холлмарк Б, Хаммер М.Ф. (ноябрь 2018). «Сибирское генетическое разнообразие раскрывает сложное происхождение самодийскоязычного населения» . Американский журнал биологии человека . 30 (6): e23194. дои : 10.1002/ajhb.23194 . ПМИД 30408262 .
- ^ Федорова С.А., Рейдла М, Метспалу Е, Метспалу М, Роотси С, Тамбетс К и др. (июнь 2013 г.). «Аутосомно-однородительские портреты коренного населения Саха (Якутии): значение для заселения Северо-Восточной Евразии» . Эволюционная биология BMC . 13 (13): 127. Бибкод : 2013BMCEE..13..127F . дои : 10.1186/1471-2148-13-127 . ПМЦ 3695835 . ПМИД 23782551 .
- ^ Перейти обратно: а б Дагган А.Т., Уиттен М., Вибе В., Кроуфорд М., Баттоф А., Спицын В. и др. (2013). «Исследование предыстории тунгусских народов Сибири и Амуро-Уссурийского региона с использованием полных последовательностей генома мтДНК и маркеров Y-хромосомы» . ПЛОС ОДИН . 8 (12): е83570. Бибкод : 2013PLoSO...883570D . дои : 10.1371/journal.pone.0083570 . ПМЦ 3861515 . ПМИД 24349531 .
- ^ Перейти обратно: а б с Харьков, В.Н.; Хамина, КВ; Медведева, О.Ф.; Штыгашева О.В.; Степанов, В.А. (2011). «Генетическое разнообразие хакасского генофонда: субэтническая дифференциация и структура гаплогрупп Y-хромосомы». Молекулярная биология . 45 (3): 404–416. дои : 10.1134/S0026893311020117 . ISSN 0026-8933 . S2CID 37140960 .
- ^ A. T. Agdzhoyan, E. V.Balanovska, A. D. Padyukova, D. O. Dolinina, M. A. Kuznetsova, V. V. Zaporozhchenko, R. A. Skhalyaho, S. M. Koshel, M. K. Zhabagin, Y. M. Yusupov, Kh. Kh. Mustafin, M. V. Ulyanova, Z. A. Tychinskih, M. B. Lavryashina, and O. P. Balanovsky (2016), "The Gene Pool of Siberian Tatars: Five Ways of Origin for the Five Subethnic Groups." МОЛЕКУЛЯРНАЯ БИОЛОГИЯ , Volume 50, No. 6, pp. 978–991. DOI: 10.7868/S0026898416060021
- ^ Перейти обратно: а б с д и ж , "СТРУКТУРА И ФИЛОГЕОГРАФИЯ ГЕНОФОНДА КОРЕННОГО НАСЕЛЕНИЯ СИБИРИ ПО МАРКЕРАМ Y-ХРОМОСОМЫ," Genetika 03.02.07 and "АВТОРЕФЕРАТ диссертации на соискание учёной степени доктора биологических наук, Tomsk 2012
- ^ Перейти обратно: а б с д Пименов В.Н., Комас Д., Пало Ю., Вершубский Г., Козлов А., Саянтила А. (октябрь 2008 г.). «Северо-западно-сибирские ханты и манси на стыке генофондов Западной и Восточной Евразии по данным однородительских маркеров» . Европейский журнал генетики человека . 16 (10): 1254–1264. дои : 10.1038/ejhg.2008.101 . ПМИД 18506205 . S2CID 19488203 .
- ^ Перейти обратно: а б с д и ж г час Лаппалайнен и др. 2008 год
- ^ Перейти обратно: а б с д Тамбетс К., Юнусбаев Б., Худжашов Г., Илумяэ А.М., Роотси С., Хонкола Т. и др. (сентябрь 2018 г.). «Гены обнаруживают следы общей недавней демографической истории для большинства уральскоязычного населения» . Геномная биология . 19 (1): 139. дои : 10.1186/s13059-018-1522-1 . ПМК 6151024 . ПМИД 30241495 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В Сюэ и др. 2006 г.
- ^ Перейти обратно: а б Богунов Ю.В.; Мальцева О.В.; Богунова А.А.; Балановская, Е.В. (2015). «НАНААЙСКИЙ КЛАН САМАР: СТРУКТУРА ГЕНОФОНДА НА ОСНОВЕ МАРКЕРОВ Y-ХРОМОСОМЫ1». Археология, этнология и антропология Евразии . 43 (2): 146–152. дои : 10.1016/j.aeae.2015.09.015 .
- ^ Плисс Л., Тимша Л., Роотси С., Тамбетс К., Пелнена И., Золе Е. и др. (ноябрь 2015 г.). «Y-хромосомные линии латышей в контексте генетической изменчивости Восточно-Балтийского региона». Анналы генетики человека . 79 (6): 418–430. дои : 10.1111/ahg.12130 . ПМИД 26411886 . S2CID 13050610 .
- ^ Перейти обратно: а б Деренко М., Малярчук Б., Денисова Г.А., Возняк М., Дамбуева И., Доржу С. и др. (январь 2006 г.). «Контрастные закономерности изменчивости Y-хромосомы в южносибирских популяциях Байкальского и Алтае-Саянского регионов». Генетика человека . 118 (5): 591–604. дои : 10.1007/s00439-005-0076-y . ПМИД 16261343 . S2CID 23011845 .
- ^ Перейти обратно: а б с д и ж г час я дж Ким Ш., Ким К.С., Шин DJ, Джин Х.Дж., Квак К.Д., Хан М.С. и др. (апрель 2011 г.). «Высокие частоты линий гаплогруппы Y-хромосомы O2b-SRY465 в Корее: генетический взгляд на заселение Кореи» . Исследовательская генетика . 2 (1): 10. дои : 10.1186/2041-2223-2-10 . ПМК 3087676 . ПМИД 21463511 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с Хаммер и др. 2006 г.
- ^ Kharkov, V. N.; Khamina, K. V.; Medvedeva, O. F.; Simonova, K. V.; Eremina, E. R.; Stepanov, V. A. (2014). "Gene pool of Buryats: Clinal variability and territorial subdivision based on data of Y-chromosome markers". Russian Journal of Genetics . 50 (2): 180–190. doi : 10.1134/S1022795413110082 . ISSN 1022-7954 . S2CID 15595963 .
- ^ Перейти обратно: а б Харьков и др. 2007 год
- ^ Перейти обратно: а б Дулик М.С., Жаданов С.И., Осипова Л.П., Аскапули А., Гау Л., Гоккумен О. и др. (февраль 2012 г.). «Вариации митохондриальной ДНК и Y-хромосомы свидетельствуют о недавнем общем происхождении коренных американцев и коренных алтайцев» . Американский журнал генетики человека . 90 (2): 229–246. дои : 10.1016/j.ajhg.2011.12.014 . ПМК 3276666 . ПМИД 22281367 .
- ^ Малярчук и др. 2004 г.
- ^ Перейти обратно: а б с д и ж Чжун Х., Ши Х., Ци С.Б., Дуань З.Ю., Тан П.П., Цзинь Л. и др. (январь 2011 г.). «Расширенное исследование Y-хромосомы позволяет предположить, что современные люди мигрировали в Восточную Азию северным путем после ледникового периода» . Молекулярная биология и эволюция . 28 (1): 717–727. дои : 10.1093/molbev/msq247 . ПМИД 20837606 .
- ^ Перейти обратно: а б Като и др. 2005 г.
- ^ Перейти обратно: а б Ди Кристофаро Дж., Пеннарун Э., Мазьер С., Майрес Н.М., Лин А.А., Темори С.А. и др. (2013). «Афганский Гиндукуш: место, где сходятся потоки генов Евразийского субконтинента» . ПЛОС ОДИН . 8 (10): е76748. Бибкод : 2013PLoSO...876748D . дои : 10.1371/journal.pone.0076748 . ПМЦ 3799995 . ПМИД 24204668 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п Балинова Н., Пост Х., Кушниаревич А., Флорес Р., Кармин М., Саакян Х. и др. (сентябрь 2019 г.). «Y-хромосомный анализ родовой структуры калмыков, единственного европейского монгольского народа, и их родство с ойратами-монголами Внутренней Азии» . Европейский журнал генетики человека . 27 (9): 1466–1474. дои : 10.1038/s41431-019-0399-0 . ПМК 6777519 . ПМИД 30976109 .
- ^ Перейти обратно: а б Малярчук Б, Деренко М, Денисова Г, Хойт С, Возняк М, Гжибовский Т, Захаров И (декабрь 2013 г.). «Y-хромосомное разнообразие у калмыков на этническом и племенном уровне» . Журнал генетики человека . 58 (12): 804–811. дои : 10.1038/jhg.2013.108 . ПМИД 24132124 .
- ^ Чжан X, Хэ Г, Ли В, Ван Ю, Ли Х, Чэнь Ю и др. (2021). «Геномный взгляд на историю смешения населения тунгусоязычных маньчжуров на северо-востоке Китая» . Границы генетики . 12 : 754492. doi : 10.3389/fgene.2021.754492 . ПМЦ 8515022 . ПМИД 34659368 .
- ^ Перейти обратно: а б с Мирабал С., Регейро М., Каденас А.М., Кавалли-Сфорца Л.Л., Андерхилл П.А., Вербенко Д.А. и др. (октябрь 2009 г.). «Распространение Y-хромосомы в геолингвистическом ландшафте северо-запада России» . Европейский журнал генетики человека . 17 (10): 1260–1273. дои : 10.1038/ejhg.2009.6 . ПМЦ 2986641 . ПМИД 19259129 .
- ^ Ван XQ, Ван CC, Дэн QY, Ли Х (февраль 2013 г.). «[Генетический анализ полиморфизма Y-хромосомы и митохондриальной ДНК этнической группы мулам в Гуанси, Китай]» . И Чуань = Эредитас (по-китайски). 35 (2): 168–74. дои : 10.3724/sp.j.1005.2013.00168 . ПМИД 23448929 .
- ^ Перейти обратно: а б Лаппалайнен Т., Ханнелиус У., Салмела Э., фон Дёбельн У., Линдгрен К.М., Хуопонен К. и др. (январь 2009 г.). «Структура населения в современной Швеции - анализ Y-хромосомной и митохондриальной ДНК». Анналы генетики человека . 73 (1): 61–73. дои : 10.1111/j.1469-1809.2008.00487.x . ПМИД 19040656 . S2CID 205598345 .
- ^ Перейти обратно: а б Карлссон А.О., Валлерстрем Т., Гётерстрем А., Холмлунд Г. (август 2006 г.). «Разнообразие Y-хромосомы в Швеции – долгосрочная перспектива» . Европейский журнал генетики человека . 14 (8): 963–970. дои : 10.1038/sj.ejhg.5201651 . ПМИД 16724001 . S2CID 23227271 .
- ^ Сон Сон Чхве, Кён Хва Пак, Да Ын Нам, Тэ Хун Кан, Ки Ва Чунг и др. , «Гаплогруппировка Y-хромосомы для азиатов с использованием целевого секвенирования Y-SNP».
- ^ Сунгвон Чон, Ёнджун Бхак, Ёнсон Чхве и др. , «Корейский проект генома: 1094 личных корейских генома с клинической информацией». Достижения науки 2020; 6: eaaz7835.
- ^ Сун Хи Ким; Бён Вон Чон; Чону Юнг; Брайан М. Кемп; и др. (июль 2005 г.). «Предварительное исследование происхождения корейцев на основе вариации Y-STR». Международный журнал юридической медицины . 32 (4): 353–359. дои : 10.1007/s13258-010-0030-9 . S2CID 7277981 .
- ^ К.С. Джонг; Х.Дж. Шин; С. Дж. Ли; Х.С. Ким; Дж. Я. Ким; М.С. Хан; Ю. Х. Ли; КВ Парк; Б.В. Чун (2018). «Генетические характеристики гаплотипов коротких тандемных повторов Y-хромосомы из образцов окурков, которые предположительно курили северокорейские мужчины». Международный журнал юридической медицины . 40 (8): 819–824. дои : 10.1007/s13258-018-0701-5 . ПМИД 30047114 . S2CID 256072306 .
- ^ Перейти обратно: а б Пак MJ, Ли ХИ, Ян В.И., Шин К.Дж. (июль 2012 г.). «Понимание вариаций Y-хромосомы в Корее - актуальность комбинированного анализа гаплогрупп и гаплотипов». Международный журнал юридической медицины . 126 (4): 589–599. дои : 10.1007/s00414-012-0703-9 . ПМИД 22569803 . S2CID 27644576 .
- ^ Перейти обратно: а б Балановская Е.В., Богунов Ю.В., Каменщикова Е.Н., Балаганская О.А., Агджоян А.Т., Богунова А.А. и др. (октябрь 2018 г.). «Демографические и генетические портреты населения ульчей». Российский генетический журнал . 54 (10): 1245–1253. дои : 10.1134/S1022795418100046 . S2CID 53085396 .
- ^ Ци X, Цуй С, Пэн Y, Чжан X, Ян Z, Чжун Х и др. (август 2013 г.). «Генетические свидетельства палеолитической колонизации и неолитической экспансии современного человека на Тибетское плато» . Молекулярная биология и эволюция . 30 (8): 1761–1778. дои : 10.1093/molbev/mst093 . ПМИД 23682168 .
- ^ Аширбеков Е.Е., Ботбаев Д.М., Белкожаев А.М., Абайлдаев А.О., Неупокоева А.С., Мухатаев Ж.Е. и др. (2017). «Распространение гаплогрупп Y-хромосомы казахов Южно-Казахстанской, Жамбылской и Алматинской областей». Доклады Национальной академии наук Республики Казахстан . 6 (316): 85–95.
- ^ Брунелли А., Кампуансай Дж., Сейлстад М., Ломтайсонг К., Кангванпонг Д., Гиротто С., Кутанан В. (2017). «Y-хромосомные данные о происхождении жителей северного Таиланда» . ПЛОС ОДИН . 12 (7): e0181935. Бибкод : 2017PLoSO..1281935B . дои : 10.1371/journal.pone.0181935 . ПМК 5524406 . ПМИД 28742125 .
- ^ Шуху Л.И., Илихаму Н.И., Баке Р.А., Бупатима А.Б., Матюсуп Д.О. (март 2018 г.). «Исследование генетического разнообразия трех изолированных популяций в Синьцзяне с использованием Y-SNP». Acta Anthropologica Sinica . 37 (1): 146–156.
- ^ Ян Л. (2011). Западном Китае Генетическая смесь популяций в (докторская диссертация) (на китайском языке: Университет Фудань, стр. 1–84).
- ^ Перейти обратно: а б Вэнь С.К., Ду ПХ, Сунь С., Цуй В., Сюй Ю.Р., Мэн Х.Л. и др. (март 2022 г.). «Двойное происхождение северо-западных китайских кыргызов: смесь сибирских Y-хромосом бронзового века и средневековых нируун-монгольских Y-хромосом». Журнал генетики человека . 67 (3): 175–180. дои : 10.1038/s10038-021-00979-x . ПМИД 34531527 . S2CID 237546416 .
- ^ Го Ю, Ся З, Цуй В, Чэнь С, Цзинь Икс, Чжу Б (май 2020 г.). «Совместный генетический анализ молекулярных маркеров митохондрий и Y-хромосомы для населения Северо-Западного Китая» . Гены . 11 (5): 564. doi : 10.3390/genes11050564 . ПМК 7290686 . ПМИД 32443545 .
- ^ Нонака, я; Минагучи, К; Такезаки, Н. (июль 2007 г.). «Бинарные гаплогруппы Y-хромосомы в японском населении и их связь с 16 полиморфизмами Y-STR». Анналы генетики человека . 71 (4): 480–495. дои : 10.1111/j.1469-1809.2006.00343.x . hdl : 10130/491 . ПМИД 17274803 . S2CID 1041367 .
- ^ Хараяма, Юта; Камей, Саяко; Сато, Норико; Хаяси, Токутаро; и др. (январь 2014 г.). «Анализ гаплогрупп Y-хромосомы у японского населения с использованием коротких ампликонов и его применение в судебно-медицинской экспертизе» (PDF) . Юридическая медицина . 16 (1): 20–25. doi : 10.1016/j.legalmed.2013.10.005 . hdl : 10091/17954 . ПМИД 24262653 . S2CID 19534157 .
- ^ Отиай, Эрико; Минагучи, Киёси; Намбияр, Пхрабхакаран; Какимото, Ю; и др. (сентябрь 2016 г.). «Оценка гаплогруппировки SNP Y-хромосомы в панели идентификации HID-Ion AmpliSeq™». Юридическая медицина . 22 : 58–61. doi : 10.1016/j.legalmed.2016.08.001 . ПМИД 27591541 . S2CID 43668117 .
- ^ Перейти обратно: а б Андерхилл и др. 2000 .
- ^ Перейти обратно: а б с Ши Х, Ци Х, Чжун Х, Пэн Ю, Чжан Х, Ма РЗ, Су Б (2013). «Генетические доказательства восточноазиатского происхождения и палеолитической миграции на север гаплогруппы N Y-хромосомы» . ПЛОС ОДИН . 8 (6): е66102. Бибкод : 2013PLoSO...866102S . дои : 10.1371/journal.pone.0066102 . ПМЦ 3688714 . ПМИД 23840409 .
- ^ Перейти обратно: а б с Кармин М., Сааг Л., Висенте М., Уилсон Сейрес М.А., Ярве М., Талас У.Г. и др. (апрель 2015 г.). «Недавнее ограничение разнообразия Y-хромосомы совпадает с глобальными изменениями в культуре» . Геномные исследования . 25 (4): 459–466. дои : 10.1101/гр.186684.114 . ПМЦ 4381518 . ПМИД 25770088 .
- ^ Кьярони, Андерхилл и Кавалли-Сфорца, 2009 г.
- ^ Перейти обратно: а б Ху и др. 2015 год
- ^ https://www.23mofang.com/ancestry/ytree/N-F2930/detail
- ^ Карафет и др. 2010 год
- ^ Перейти обратно: а б де Баррос Дамгаард П., Мартиниано Р., Камм Дж., Морено-Майар Дж.В., Кроонен Г., Пейро М. и др. (июнь 2018 г.). «Первые пастухи и влияние степной экспансии раннего бронзового века в Азию» . Наука . 360 (6396): eaar7711. дои : 10.1126/science.aar7711 . ПМК 6748862 . ПМИД 29743352 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также и аль являюсь а к ап ак Дерево гаплогруппы Y-ДНК N-M231 в Family Tree DNA Discover
- ^ "Haplotree.info - Ancientdna.info. Карта на основе All Ancient DNA v. 2.07.26" .
- ^ Перейти обратно: а б с Карафет и др. 2001 г.
- ^ Перейти обратно: а б Вен и др. 2004 г.
- ^ Перейти обратно: а б Цай и др. 2011 год
- ^ Ким и др. 2007 год
- ^ Гайден и др. 2007 год
- ^ Перейти обратно: а б Цуй, Иньцю; Ли, Хунцзе; Нин, Чао; Чжан, Е; и др. (30 сентября 2013 г.). «Анализ Y-хромосомы доисторических человеческих популяций в долине реки Западный Ляо, Северо-Восточный Китай» . Эволюционная биология BMC . 13 (1): 216. Бибкод : 2013BMCEE..13..216C . дои : 10.1186/1471-2148-13-216 . ПМЦ 3850526 . ПМИД 24079706 . S2CID 13544780 .
- ^ «N-Z1956 YДерево» .
- ^ Перейти обратно: а б Сирак и др. 2020 .
- ^ Пакендорф и др. 2002 г.
- ^ Крубези и др. 2010 год
- ^ Перейти обратно: а б Деренко и др. 2007 год
- ^ Дулик М.К., Оуингс А.С., Гайески Дж.Б., Вилар М.Г., Андре А., Ленни С. и др. (май 2012 г.). «Анализ Y-хромосомы выявляет генетическое расхождение и появление новых коренных родословных в атапасских и эскимосских популяциях» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (22): 8471–8476. Бибкод : 2012PNAS..109.8471D . дои : 10.1073/pnas.1118760109 . ПМК 3365193 . ПМИД 22586127 .
- ^ Кылинч Г.М., Кашуба Н., Коптекин Д., Бергфельдт Н., Донерташ Х.М., Родригес-Варела Р. и др. (январь 2021 г.). «Динамика человеческого населения и Yersinia pestis в древней Северо-Восточной Азии» . Достижения науки . 7 (2): eabc4587. Бибкод : 2021SciA....7.4587K . дои : 10.1126/sciadv.abc4587 . ПМЦ 7787494 . ПМИД 33523963 .
- ^ «N-L708 YДерево» . Проверено 28 ноября 2022 г.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также и аль являюсь а к ап ак с как в В из хорошо топор является тот нет бб до нашей эры др. быть парень бг чб с Филогенетическое древо гаплогруппы N на 23 мофанге
- ^ Чэнь Дж, Хэ Г, Жэнь З, Ван Ц, Лю Ю, Чжан Х, Ян М, Чжан Х, Цзи Дж, Чжао Дж, Го Дж, Чжу К, Ян Икс, Ван Р, Ма Х, Ван CC и Хуан J (2021), «Геномный взгляд на историю примеси монголо- и тунгусоязычного населения Юго-Западной Восточной Азии». Передний. Жене. 12:685285. дои: 10.3389/fgene.2021.685285
- ^ «Zhōngguó manzú qúntì fùxì dan bei qún ji minzú xiě tŏng gòuchéng qíngkuàng jiânjiè» Краткое введение в отцовскую гаплогруппу и состав этнического происхождения маньчжуров в Китае. [Краткое введение в отцовскую гаплогруппу и состав этнического происхождения маньчжурского народа в Китае]. 23мофан (на китайском языке). 21 января 2021 г. Проверено 11 марта 2024 г.
- ^ Ху и др. 2015 .
- ^ Ма, Пэнчэн; Ян, Сюань; Ян, Ши; Ли, Чуньсян; и др. (декабрь 2021 г.). «Древняя Y-ДНК с реконструированной филогенией дает представление о демографической истории отцовской гаплогруппы N1a2-F1360» . Журнал генетики и геномики . 48 (12): 1130–1133. дои : 10.1016/j.jgg.2021.07.018 . ПМИД 34425243 . S2CID 237281094 .
- ^ Перейти обратно: а б с Мао, Сяовэй; Чжан, Хуцай; Цяо, Шию; Лю, Ичэнь; и др. (июнь 2021 г.). «Глубокая история населения северной части Восточной Азии от позднего плейстоцена до голоцена» . Клетка . 184 (12): 3256–3266.e13. дои : 10.1016/j.cell.2021.04.040 . ISSN 0092-8674 . ПМИД 34048699 .
- ^ «Гаплогруппа Theytree YTree (N-Y24206)» . Проверено 10 марта 2024 г.
- ^ «Гаплогруппа Theytree YTree (N-F710)» . Проверено 10 марта 2024 г.
- ^ Лю, Цзюнсен; Цзэн, Вэнь; Сунь, Бо; Мао, Сяовэй; и др. (29 мая 2021 г.). «Материнская генетическая структура в древнем Шаньдуне между 9500 и 1800 годами назад» . Научный вестник . 66 (11): 1129–1135. Бибкод : 2021SciBu..66.1129L . дои : 10.1016/j.scib.2021.01.029 . ПМИД 36654346 .
- ^ Ван, Хунру; Ян, Мелинда А.; Вангду, Шарган; Лу, Хунлян; и др. (15 марта 2023 г.). «Генетическая история человека на Тибетском нагорье за последние 5100 лет» . Достижения науки . 9 (11): eadd5582. Бибкод : 2023SciA....9D5582W . дои : 10.1126/sciadv.add5582 . ISSN 2375-2548 . ПМЦ 10022901 . ПМИД 36930720 .
- ^ «Гаплогруппа Theytree YTree (N-CTS2959)» . Проверено 10 марта 2024 г.
- ^ Карафет Татьяна М.; Осипова Людмила П; Губина Марина А; Посух, Ольга Л; и др. (декабрь 2002 г.). «Высокий уровень дифференциации Y-хромосомы среди коренного сибирского населения и генетическая подпись бореального образа жизни охотников-собирателей» . Биология человека . 74 (6): 761–789. дои : 10.1353/hub.2003.0006 . ПМИД 12617488 . S2CID 9443804 .
- ^ Тамбетс, Кристина; Роотси, Сирия [на эстонском языке] ; Кивисилд, Томас ; Помогите, Ад; и др. (апрель 2004 г.). «Западные и восточные корни саамов – история генетических «выбросов», рассказанная митохондриальной ДНК и Y-хромосомами» . Американский журнал генетики человека . 74 (4): 661–682. дои : 10.1086/383203 . ПМК 1181943 . ПМИД 15024688 . S2CID 18395985 .
- ^ Харьков, ВН; Колесников Н.А.; Валихова Л.В.; Зарубин А.А.; и др. (март 2023 г.). «Связь генофонда хантов с народами Западной Сибири, Приуралья и Алтае-Саянского региона по данным о полиморфизме аутосомного локуса и Y-хромосомы » Вавиловский журнал генетики и селекции . 27 (1): 46–54. дои : 10.18699/VJGB-23-07 . eISSN 2500-3259 . ПМЦ 10009483 . ПМИД 36923476 . S2CID 257478780 .
- ^ «Подробности о гаплогруппе N-F2930» .
- ^ Перейти обратно: а б N-P189.2 YПолный, 2018 г. (24 июня 2018 г.).
- ^ Гаплогруппа N Y-ДНК и ее субклады - 2018 , ISOGG , 2018, (24 июня 2018 г.).
- ^ Перейти обратно: а б де Баррос Дамгаард, Питер; Марки, Нина; Расмуссен, Саймон; Пейро, Микаэль; и др. (9 мая 2018 г.). «137 древних геномов человека со всех евразийских степей» . Природа . 557 (7705): 369–374. Бибкод : 2018Natur.557..369D . дои : 10.1038/s41586-018-0094-2 . hdl : 1887/3202709 . ПМИД 29743675 . S2CID 256769352 .
- ^ Гамба, Кристина; Джонс, Эппи Р.; Тисдейл, Мэтью Д.; Маклафлин, Рассел Л.; и др. (21 октября 2014 г.). «Поток и застой генома на протяжении пяти тысячелетий европейской предыстории» (PDF) . Природные коммуникации . 5 : 5257. Бибкод : 2014NatCo...5.5257G . дои : 10.1038/ncomms6257 . ПМЦ 4218962 . ПМИД 25334030 . S2CID 19587039 .
- ^ Перейти обратно: а б с д и ж Гаплогруппа N Североевразийский проект YDNA в генеалогическом древе ДНК
- ^ Липпольд, Себастьян; Сюй, Хунъян; Ко, Альберт; Ли, Минкун; и др. (24 сентября 2014 г.). «Демографические истории отца и матери человека: данные из Y-хромосомы высокого разрешения и последовательностей мтДНК» (PDF) . Исследовательская генетика . 5:13 . дои : 10.1186/2041-2223-5-13 . ПМК 4174254 . ПМИД 25254093 . S2CID 16464327 .
- ^ Ван, Лин-Сян; Лу, Ян; Чжан, Чао; Вэй, Лан-Хай; и др. (октябрь 2018 г.). «Реконструкция филогении Y-хромосомы выявляет две неолитические экспансии тибето-бирманского населения». Молекулярная генетика и геномика . 293 (5): 1293–1300. дои : 10.1007/s00438-018-1461-2 . ПМИД 29923068 . S2CID 49311699 .
- ^ Перейти обратно: а б Ли, Ивонн Ю; Чанг, Грейс Тай; Луи, Вивиан, Вайоминг; То, Ка-Фай; и др. (18 января 2017 г.). «Секвенирование экзома и генома рака носоглотки выявляет мутации, активирующие путь NF-κB» (PDF) . Природные коммуникации . 8 : 14121. Бибкод : 2017NatCo...814121L . дои : 10.1038/ncomms14121 . ПМЦ 5253631 . ПМИД 28098136 . S2CID 15387583 .
- ^ Биобанк Института медицинских исследований Кориелла.
- ^ Джоблинг и Тайлер-Смит, 2000 г.
- ^ Каладжиева и др. 2001 г.
- ^ Андерхилл и др. 2000 г.
- ^ Хаммер и др. 2001 г.
- ^ Семино и др. 2000 г.
- ^ Су и др. 1999 год
- ^ Капелли и др. 2001 г.
Библиография
[ редактировать ]- Цай, Сяоюнь; Цинь, Чжэндун; Вэнь, Бо; Сюй, Шухуа; и др. (31 августа 2011 г.). О'Рурк, Деннис (ред.). «Миграция человека через узкие места из Юго-Восточной Азии в Восточную Азию во время последнего ледникового максимума, выявленная Y-хромосомами» . ПЛОС Один . 6 (8). Генографический консорциум: e24282. Бибкод : 2011PLoSO...624282C . дои : 10.1371/journal.pone.0024282 . ПМК 3164178 . ПМИД 21904623 . S2CID 2731184 .
- Кьярони, Жак; Андерхилл, Питер А; Кавалли-Сфорца, Лука Л. (17 ноября 2009 г.). «Разнообразие Y-хромосомы, человеческая экспансия, дрейф и культурная эволюция» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (48): 20174–20179. Бибкод : 2009PNAS..10620174C . дои : 10.1073/pnas.0910803106 . ПМЦ 2787129 . ПМИД 19920170 . S2CID 32040326 .
- Крубези, Эрик; Амори, Сильвен; Кейзер, Кристина; Буаказе, Кэролайн; и др. (25 января 2010 г.). «Эволюция человека в Сибири: от замороженных тел к древней ДНК» (PDF) . Эволюционная биология BMC . 10 (1): 25. Бибкод : 2010BMCEE..10...25C . дои : 10.1186/1471-2148-10-25 . ПМК 2829035 . ПМИД 20100333 . S2CID 16326890 .
- Деренко, Мирослава; Венок, Борис; Денисова Галина; Возняк, Марцин; и др. (17 августа 2007 г.). «Распространение гаплогруппы N Y-хромосомы из южной Сибири в Европу» (PDF) . Журнал генетики человека . 52 (9): 763–770. дои : 10.1007/s10038-007-0179-5 . ПМИД 17703276 . S2CID 25599501 .
- Гайден, Тензин; Каденас, Алисия М; Регейру, Мария; Сингх, Нанда Б; и др. (4 апреля 2007 г.). «Гималаи как направленный барьер на пути потока генов» (PDF) . Американский журнал генетики человека . 80 (5): 884–894. дои : 10.1086/516757 . ПМЦ 1852741 . ПМИД 17436243 . S2CID 25312596 .
- Хаммер, Майкл Ф; Карафет Татьяна М; Пак, Хваён; Омото, Кэйичи [на японском языке] ; и др. (2006). «Двойное происхождение японцев: общая основа Y-хромосом охотников-собирателей и фермеров» (PDF) . Журнал генетики человека . 51 (1): 47–58. дои : 10.1007/s10038-005-0322-0 . ПМИД 16328082 . S2CID 6559289 .
- Илумяэ, Анне-Май; Рейдла, Маере [на эстонском языке] ; Чухряева Марина; Ярве, Мари [на эстонском языке] ; и др. (07.07.2016). «Гаплогруппа Y-хромосомы человека: нетривиальная филогеография с временным разрешением, охватывающая языковые семьи» (PDF) . Американский журнал генетики человека . 99 (1): 163–173. дои : 10.1016/j.ajhg.2016.05.025 . ПМЦ 500549 . ПМИД 27392075 . S2CID 20536258 .
- Ху, Канг; Ян, Ши; Лю, Кай; Нин, Чао; и др. (24 апреля 2015 г.). «Дихотомическая структура гаплогруппы N Y-хромосомы». arXiv : 1504.06463 [ q-bio.PE ].
- Карафет, Татьяна; Сюй, Липин; Ду, Руофу; Ван, Уильям; и др. (30 июля 2001 г.). «История отцовского населения Восточной Азии: источники, закономерности и микроэволюционные процессы» (PDF) . Американский журнал генетики человека . 69 (3): 615–628. дои : 10.1086/323299 . ПМЦ 1235490 . ПМИД 11481588 . S2CID 27785973 .
В этой статье образец «Южная Хань», полученный исследовательской группой Карафета и Хаммера, описывается как происходящий из провинции Гуандун, а образец «Северная Хань» описывается как происходящий из провинции Шэньси.
- Карафет Татьяна М; Мендес, Фернандо Л; Мейлерман, Моника Б; Андерхилл, Питер А; и др. (2 апреля 2008 г.). «Новые бинарные полиморфизмы изменяют форму и увеличивают разрешение дерева гаплогруппы Y-хромосомы человека» . Геномные исследования . 18 (5): 80–838. дои : 10.1101/гр.7172008 . ПМК 2336805 . ПМИД 18385274 . S2CID 12995903 .
- Карафет Татьяна М; Холлмарк, Брайан; Кокс, Мюррей П; Судойо, Херавати [на индонезийском языке] ; и др. (05 марта 2010 г.). «Основное разделение Восток-Запад лежит в основе стратификации Y-хромосомы в Индонезии» . Молекулярная биология и эволюция . 27 (8): 1833–1844. дои : 10.1093/molbev/msq063 . ПМИД 20207712 . S2CID 4819475 .
- Като, Тору; Мунхбат, Батмунх; Тунаи, Кеничи; Мано, Шухей; и др. (14 февраля 2005 г.). «Генетические особенности монгольских этносов, выявленные с помощью Y-хромосомного анализа». Джин . 346 : 63–70. дои : 10.1016/j.gene.2004.10.023 . ПМИД 15716011 . S2CID 28132496 .
- Харьков В.Н., Степанов В.А., Медведева О.Ф., Спиридонова М.Г., Воевода М.И., Тадинова В.Н. и др. (май 2007 г.). «Различия генофонда северных и южных Алтайцев, выведенные из данных по гаплогруппам Y-хромосомы» (PDF) . Российский генетический журнал . 43 (5): 551–562. дои : 10.1134/S1022795407050110 . ПМИД 17633562 . S2CID 566825 .
- Ким В., Ю ТК, Ким С.Дж., Шин DJ, Тайлер-Смит С., Джин Х.Дж. и др. (январь 2007 г.). Благосклонный М (ред.). «Отсутствие связи между гаплогруппами Y-хромосомы и раком простаты у корейского населения» . ПЛОС ОДИН . 2 (1): е172. Бибкод : 2007PLoSO...2..172K . дои : 10.1371/journal.pone.0000172 . ПМЦ 1766463 . ПМИД 17245448 .
- Лаппалайнен Т., Лайтинен В., Салмела Э., Андерсен П., Хуопонен К., Савонтаус М.Л., Лаэрмо П. (май 2008 г.). «Миграционные волны в регион Балтийского моря» . Анналы генетики человека . 72 (Часть 3): 337–348. дои : 10.1111/j.1469-1809.2007.00429.x . ПМИД 18294359 . S2CID 32079904 .
- Малярчук Б, Деренко М, Гжибовский Т, Лунькина А, Чарный Ю, Рычков С и др. (декабрь 2004 г.). «Дифференциация митохондриальной ДНК и Y-хромосом в российских популяциях». Биология человека . 76 (6): 877–900. дои : 10.1353/hub.2005.0021 . ПМИД 15974299 . S2CID 17385503 .
- Пакендорф Б., Морар Б., Тарская Л.А., Кайзер М., Судьял Х., Родевальд А., Стоункинг М. (февраль 2002 г.). «Y-хромосомные доказательства сильного сокращения численности мужского населения якутов». Генетика человека . 110 (2): 198–200. дои : 10.1007/s00439-001-0664-4 . ПМИД 11935328 . S2CID 30671159 .
- Сирак К., Фернандес Д., Черонет О., Харни Э., Мах М., Маллик С. и др. (март 2020 г.). «Слуховые косточки человека как альтернативный оптимальный источник древней ДНК» . Геномные исследования . 30 (3): 427–436. дои : 10.1101/гр.260141.119 . ПМК 7111520 . ПМИД 32098773 .
- Вэнь Б, Се Х, Гао С, Ли Х, Ши Х, Сун Х и др. (май 2004 г.). «Анализ генетической структуры тибето-бирманских популяций выявил примесь по признаку пола у южных тибето-бирманцев» . Американский журнал генетики человека . 74 (5): 856–865. дои : 10.1086/386292 . ПМК 1181980 . ПМИД 15042512 .
- Сюэ Ю, Зержал Т, Бао В, Чжу С, Шу Ц, Сюй Дж и др. (апрель 2006 г.). «Мужская демография в Восточной Азии: контраст между севером и югом во времени роста населения» . Генетика . 172 (4): 2431–2439. doi : 10.1534/genetics.105.054270 . ПМЦ 1456369 . ПМИД 16489223 .
Веб-сайты
[ редактировать ]- ИЗОГГ (2012). «Дерево гаплогрупп Y-ДНК 2012» .
- Макдональд, доктор юридических наук (2004). «Гаплогруппы Макдональда Y мира» (PDF) . Архивировано из оригинала (PDF) 28 июля 2004 г. Проверено 29 августа 2009 г.
Источники таблиц преобразования
[ редактировать ]- Капелли, Кристиан; Уилсон, Джеймс Ф.; Ричардс, Мартин; Штумпф, Майкл П.Х.; и др. (февраль 2001 г.). «Отцовское наследие преимущественно коренных народов австронезийскоязычных народов островной Юго-Восточной Азии и Океании» . Американский журнал генетики человека . 68 (2): 432–443. дои : 10.1086/318205 . ПМЦ 1235276 . ПМИД 11170891 .
- Хаммер, Майкл Ф.; Карафет Татьяна М.; Редд, Алан Дж.; Джарджанази, Хамди; и др. (1 июля 2001 г.). «Иерархические закономерности глобального разнообразия Y-хромосом человека» . Молекулярная биология и эволюция . 18 (7): 1189–1203. doi : 10.1093/oxfordjournals.molbev.a003906 . ПМИД 11420360 .
- Джоблинг, Марк А.; Тайлер-Смит, Крис (2000), «Новое использование новых гаплотипов», Trends in Genetics , 16 (8): 356–62, doi : 10.1016/S0168-9525(00)02057-6 , PMID 10904265
- Каладжиева, преп.; Калафель, Франческ; Джоблинг, Марк А; Ангеличева, Дора; и др. (февраль 2001 г.). «Закономерности меж- и внутригруппового генетического разнообразия у Vlax Roma, выявленные с помощью Y-хромосомы и линий митохондриальной ДНК» . Европейский журнал генетики человека . 9 (2): 97–104. дои : 10.1038/sj.ejhg.5200597 . ПМИД 11313742 .
- Семино О., Пассарино Г., Офнер П.Дж., Лин А.А. и др. (2000), «Генетическое наследие палеолитического Homo sapiens sapiens у современных европейцев: хромосомная перспектива AY», Science , 290 (5494): 1155–1159, Бибкод : 2000Sci...290.1155S , doi : 10.1126/science.290.5494. 1155 , ПМИД 11073453
- Су, Бинг; Сяо, Цзюньхуа; Андерхилл, Питер; Дека, Ранджан; и др. (декабрь 1999 г.). «Y-хромосома свидетельствует о миграции современных людей на север в Восточную Азию во время последнего ледникового периода» . Американский журнал генетики человека . 65 (6): 1718–1724. дои : 10.1086/302680 . ПМЦ 1288383 . ПМИД 10577926 .
- Андерхилл П.А., Шен П., Лин А.А., Джин Л. и др. (ноябрь 2000 г.). «Вариации последовательности Y-хромосомы и история человеческих популяций». Природная генетика . 26 (3): 358–361. дои : 10.1038/81685 . ПМИД 11062480 . S2CID 12893406 .
Дальнейшее чтение
[ редактировать ]Этот раздел нуждается в расширении . Вы можете помочь, добавив к нему . ( декабрь 2012 г. ) |
- ИЗОГГ (2006). «Дерево гаплогрупп Y-ДНК 2006» .
- ИЗОГГ (2007). «Дерево гаплогрупп Y-ДНК 2007» .
- ИЗОГГ (2008). «Дерево гаплогрупп Y-ДНК 2008» .
- ИЗОГГ (2009). «Дерево гаплогрупп Y-ДНК 2009» .
- ИЗОГГ (2010). «Дерево гаплогрупп Y-ДНК 2010» .
- ИЗОГГ (2011). «Дерево гаплогрупп Y-ДНК 2011» .
- ИЗОГГ (2018). «Гаплогруппа N Y-ДНК и ее субклады» .
- YПолный. «Полное экспериментальное YTree» .
- ИЗОГГ (2019–2020 гг.). «Гаплогруппа N Y-ДНК и ее субклады» .
Филогенетика
[ редактировать ]- ^ Делеция b2/b3 в регионе AZFc Y-хромосомы человека является характеристикой гаплотипов гаплогруппы N-M231. Однако это удаление, по-видимому, происходило независимо в четырех разных случаях. Поэтому эту делецию не следует рассматривать как уникальный полиморфизм событий, способствующий определению этой ветви дерева Y-хромосомы ( ISOGG 2012 ).
- ^ В этой таблице показаны исторические названия N-M46 (AKA N-Tat) из рецензируемой литературы.
YCC 2002/2008 (сокращенно) Н-М46/Н-ТАТ Джоблинг и Тайлер-Смит, 2000 г. 12 Андерхилл 2000 VIII Хаммер 2001 1я Карафет 2001 26 Семино 2000 ЕС13 Они в 1999 году Н5 Волосы 2001 Ф YCC 2002 (от руки) N3* YCC 2005 (от руки) N3 YCC 2008 (от руки) N1c YCC 2010r (от руки) N1c - ^ В этой таблице показаны исторические названия N-M178 из рецензируемой литературы.
YCC 2002/2008 (сокращенно) Н-М178 Джоблинг и Тайлер-Смит, 2000 г. 16 Андерхилл 2000 VIII Хаммер 2001 1я Карафет 2001 26 Семино 2000 ЕС14 Они в 1999 году Н5 Волосы 2001 Ф YCC 2002 (от руки) Н3а* YCC 2005 (от руки) М178 YCC 2008 (от руки) N1c1 YCC 2010r (от руки) N1c1 - ^ В этой таблице показаны исторические названия N-M128 из рецензируемой литературы.
YCC 2002/2008 (сокращенно) Н-М128 Джоблинг и Тайлер-Смит, 2000 г. 12 Андерхилл 2000 VIII Хаммер 2001 1U Карафет 2001 25 Семино 2000 ЕС16 Они в 1999 году Н5 Волосы 2001 Ф YCC 2002 (от руки) N1 YCC 2005 (от руки) N1 YCC 2008 (от руки) Н1а YCC 2010r (от руки) Н1а - ^ Эту ветвь на ранних деревьях иногда называют N1b.
Внешние ссылки
[ редактировать ]- Распространение гаплогруппы N , из Генографического проекта , National Geographic.
- Северо-евразийский проект YDNA в FamilyTreeDNA
- Проект гаплогруппы N Y-ДНК в FamilyTreeDNA
- Проект гаплогруппы N1c1 Y-ДНК в FamilyTreeDNA
- Распространение гаплогруппы N Y-хромосомы из Южной Сибири в Европу
- Проект ДНК династии Рюриковичей в FamilyTreeDNA
- Проект ДНК русского дворянства в FamilyTreeDNA