Гаплогруппа D-CTS3946
| Гаплогруппа D-CTS3946 | |
|---|---|
 | |
| Возможное время происхождения | 80 700–64 700 лет назад (ок. 72 700 лет назад) [ 1 ] |
| Возможное место происхождения | Африка [ 1 ] |
| Предок | ИЗ |
| Потомки | D-M174 (также известный как D1) D-A5580.2 (также известный как D2 и D0) |
| Определение мутаций | CTS3946, CTS4030/Z1605 |
Гаплогруппа D , также известная как D-CTS3946 , представляет собой гаплогруппу Y-хромосомы . Как и его относительный дальний брат, гаплогруппа E-M96 , D-CTS3946 обладает полиморфизмом уникальных событий YAP+ , который определяет их родительскую гаплогруппу DE . D-CTS3946 имеет две базальные ветви: D1 и D2 . D1 и D2 встречаются главным образом в Восточной Азии , с низкой частотой — в Центральной Азии и Юго-Восточной Азии и с очень низкой частотой — в Западной Африке и Западной Азии .
Происхождение
[ редактировать ]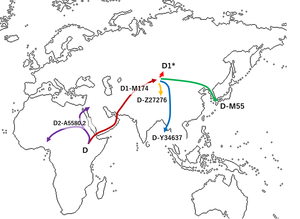
Гаплогруппа D ранее называлась линией D D-M174 . Существуют различные предположения относительно происхождения гаплогруппы DE , родительской гаплогруппы D, некоторые предполагают африканское происхождение. [ 2 ] и другие азиатского происхождения. [ 3 ] Но D-M174 считалось и обычно считается азиатского происхождения и встречается исключительно в Азии.
Хабер и др. (2019) идентифицировали гаплогруппу, названную «D0», у трех нигерийцев. Гаплогруппа «D0», определенная SNP A5580.2, находится за пределами M174, но принадлежит к линии D, разделяет с D-M174 7 SNP, которых нет у E, и было установлено, что она рано отошла от ветви D (около D-M174). /E разделить). Хабер и др. (2019) рассмотрели несколько возможностей, включая африканское и азиатское происхождение гаплогруппы, но отчасти из-за глубокого укоренения гаплогруппы «D0», а также недавно рассчитали время раннего расхождения для нее и ее родительской гаплогруппы DE. авторы делают вывод в пользу африканского происхождения D0 и DE, а также общего предка (теперь известного как D-CTS3946 или «D») D0 и D-M174. По данным Хабера и др. (2019), D0 — это ветвь линии DE, расположенная рядом с разделением D/E, но на ветви D, разошедшаяся около 71000 лет назад. Авторы находят время расхождения для DE*, E и D0, вероятно, в пределах периода около 76 000–71 000 лет назад, а вероятную дату выхода предков современных евразийцев из Африки (и последующей примеси неандертальцев) позже. , примерно 50 300–59 400 лет назад, что, по их мнению, также подтверждает африканское происхождение этих гаплогрупп. [ 1 ] Таким образом, предполагается, что D-CTS3946 распространился как внутри, так и за пределами Африки: одна ветвь дивергировала в D0 в Африке, а другая ветвь, покинувшая Африку, в конечном итоге дивергировала в D-M174 (т.е. с мутацией M174, позже возникшей из D-CTS3946). распространился в Азию).
Халласт и др. (2020) о древних и современных гаплогруппах с использованием филогенетического анализа последовательностей гаплогрупп C, D и FT, включая очень редкие линии с глубокими корнями (такие как D0/D2, отобранные Хабером и др., 2019), утверждает, принимая «редкие глубокие -rooted D0» учитывает, что первоначальные расколы внутри гаплогруппы CT (предка DE) произошли в Африке. Они также утверждают, что все филогеографические анализы древних и современных неафриканских Y-хромосом указывают на Восточную/Юго-Восточную Азию как на источник 55 000–50 000 лет назад всех известных сохранившихся неафриканских мужских линий (кроме недавних мигрантов) вскоре после этого. первоначальная миграция базальной гаплогруппы D и других базальных Y-линий из Африки, произошедшая 70–55 000 лет назад. Они утверждают, что эти линии затем быстро распространились по Евразии, позже диверсифицировались в Юго-Восточной Азии, а затем распространились на запад около 55–50 000 лет назад, заменив другие местные линии в Евразии, и пришли к выводу, что гаплогруппа D (как D-M174) затем подверглась быстрой экспансии внутри Евразии. Восточно-евразийское население и состоит из 5 различных ветвей, сформировавшихся около 45 000 лет назад. Они обнаружили, что эти гаплогруппы в настоящее время имеют наибольшее разнообразие в Восточной Евразии (восток/юго-восточная Азия). Было обнаружено, что тибето-бирманское население Восточной и Юго-Восточной Азии отличается наибольшим разнообразием. [ 4 ]
Распределение
[ редактировать ]
FTDNA также обнаружила три других образца D2 (также в 2019 году): два в Аль-Ваджхе на западном побережье Саудовской Аравии. [ 5 ] [ 6 ] и еще один на сирийском (он же D2b-FT51782). [ 7 ] [ 5 ] Образец D2b-FT51782 также был найден в Аль-Кирби из Байды. [ 8 ] в Йемене, который, как оказалось, на несколько тысяч лет был связан с сирийским. [ 7 ] есть еще один человек (фамилия неизвестна) В соседней провинции Шабва с TMRCA 1700 г. назад с первым. [ 9 ] В 2020 году Майкл Сейгер из FTDNA объявил, что еще один образец был найден у афроамериканца. [ 10 ] и один в другом афроамериканец. [ 10 ] [ 11 ] Образцы, обнаруженные у сирийского потомка и у одного из афроамериканцев, на сегодняшний день являются наиболее базальными образцами D2. [ 10 ] [ 5 ] [ 6 ] [ 11 ] Другая афроамериканская выборка делит ветвь с тремя нигерийцами. [ 10 ] Недавние данные (как также предположили Хабер и др.) предполагают, что D2 представляет собой сильно дивергентную гаплогруппу, близкую к расколу DE, но на ветви D и лишенную мутации M174, которой обладают другие известные линии D (принадлежащие ее родственному брату D- М174). [ 5 ]
Субклады
[ редактировать ]Д1
[ редактировать ]Гаплогруппа D1 является субкладом гаплогруппы D-CTS3946.
Д2
[ редактировать ]Гаплогруппа D2 представляет собой субклад верхнего палеолита гаплогруппы D-CTS3946. В связи с этим открытием D0 был переименован в D2, а D-M174 был переименован в D1. [ 5 ] [ 6 ]
Филогенетические деревья
[ редактировать ]ISOGG (версия: 14.151). [ 12 ]
- DE (YAP) Карибские острова , Гвинея-Бисау , Тибет , Нигерия
- Д (CTS3946)
- D1 (M174/Страница30, IMS-JST021355)
- Д1а (CTS11577)
- Д1а1 (F6251/Z27276)
- D1a1a (M15) Китай (особенно Мяо , И , тибетцы ) и др. , Юго-Восточная Азия , Монголия , Средняя Азия
- D1a1b (P99) Часто встречается в Тибете и изредка в других частях Китая , Монголии , [ 12 ] Средняя Азия , [ 13 ] и Республика Алтай
- Д1а2 (Z3660)
- D1a2a (M64.1/Page44.1, M55) Япония [ 13 ]
- Д1а2а1 (М116.1)
- Д1а2а2 (CTS131)
- Д1а2а2а (CTS220)
- D1a2a2b (CTS68) Япония (остров Ребун)
- D1a2b (Y34637) Андаманские острова [ 14 ]
- D1a2a (M64.1/Page44.1, M55) Япония [ 13 ]
- Д1а1 (F6251/Z27276)
- D1b (L1378) Филиппины [ 15 ]
- Д1а (CTS11577)
- D2 (A5580.2) Нигерия , Саудовская Аравия , Сирия , США , Йемен
- D1 (M174/Страница30, IMS-JST021355)
- Д (CTS3946)
Ссылки
[ редактировать ]- ^ Jump up to: а б с Хабер М., Джонс А.Л., Коннелл Б.А., Арсьеро Э., Ян Х., Томас М.Г. и др. (август 2019 г.). «Редкая глубоко укоренившаяся африканская гаплогруппа Y-хромосомы D0 и ее значение для расселения современного человека за пределы Африки» . Генетика . 212 (4): 1421–1428. дои : 10.1534/genetics.119.302368 . ПМК 6707464 . ПМИД 31196864 .
- ^ Андерхилл, Пенсильвания, Кивисилд Т (2007). «Использование структуры популяции Y-хромосомы и митохондриальной ДНК для отслеживания миграций человека». Ежегодный обзор генетики . 41 : 539–64. дои : 10.1146/annurev.genet.41.110306.130407 . ПМИД 18076332 .
- ^ Кабрера В.М. (2017). «Носители основных линий макрогаплогруппы L3 митохондриальной ДНК мигрировали обратно в Африку из Азии около 70 000 лет назад». bioRxiv 10.1101/233502 .
- ^ Халласт П., Агджоян А., Балановский О., Сюэ Ю., Тайлер-Смит С. (февраль 2021 г.). «Происхождение современных неафриканских Y-хромосом человека из Юго-Восточной Азии» . Генетика человека . 140 (2): 299–307. дои : 10.1007/s00439-020-02204-9 . ПМЦ 7864842 . ПМИД 32666166 .
Принимая во внимание редкую африканскую линию D0 и временные рамки, изложенные выше, мы пришли к выводу (Haber et al. 2019), что первоначальные расколы внутри CT, вероятно, произошли в Африке до выхода, и что три линии, C, D, и FT, осуществлялись предками современных неафриканцев. Каждая из этих трех линий впоследствии расширилась: C и D умеренно, а FT массово». лет назад, за которым следовала смесь неандертальцев, составлявшая ~ 2% всех неафриканцев. Однако детали этой первоначальной миграции остаются плохо изученными, поскольку анализы древней ДНК за этот ключевой период времени недоступны, а интерпретация современных аутосомных данных затруднена из-за последующих перемещений/изменений популяции. Однако один локус действительно сохраняет информацию, специфичную для самцов этого раннего периода: Y-хромосома, где была построена детальная калиброванная филогения. В результате первоначальной миграции были перенесены три современные линии Y: редкая гаплогруппа D, умеренно редкая C и очень распространенная линия FT, которая сейчас доминирует в большинстве неафриканских популяций.
 В эту статью включен текст из этого источника, доступного по лицензии CC BY 4.0 .
В эту статью включен текст из этого источника, доступного по лицензии CC BY 4.0 .
- ^ Jump up to: а б с д и Эстес, Роберта (21 июня 2019 г.). «Захватывающие новые открытия гаплогруппы D ДНК Y!» . DNAeXplained – Генетическая генеалогия . Проверено 06 июля 2019 г.
- ^ Jump up to: а б с "Изображение" .
- ^ Jump up to: а б «FamilyTreeDNA – Проект гаплогруппы Y-ДНК D» .
- ^ "Д YДерево" .
- ^ «D-Y330435 YДерево» . Архивировано из оригинала 1 мая 2023 г.
- ^ Jump up to: а б с д Сагер, Майкл (февраль 2020 г.). Древо человечества от FTDNA (Майк Сагер) .
- ^ Jump up to: а б Эстес, Роберта (10 декабря 2020 г.). «Новые открытия проливают свет на теорию выхода из Африки и за ее пределы» . DNAeXplained – Генетическая генеалогия .
- ^ Jump up to: а б Гаплогруппа D Y-ДНК и ее субклады – 2019 г.
- ^ Jump up to: а б с Хаммер М.Ф., Карафет Т.М., Парк Х. и др. (2006). «Двойное происхождение японцев: общая основа для Y-хромосом охотников-собирателей и фермеров». Дж. Хум. Жене. 51 (1): 47–58. doi:10.1007/s10038-005-0322-0. ПМИД 16328082 .
- ^ "Д YДерево" . www.yfull.com . Проверено 3 сентября 2019 г.
- ^ Гаплогруппа D Y-ДНК и ее субклады - 2014 г.