Гаплогруппа D-M15
| Гаплогруппа D-M15 | |
|---|---|
| Предок | Д1а1 |
| Определение мутаций | М15 |
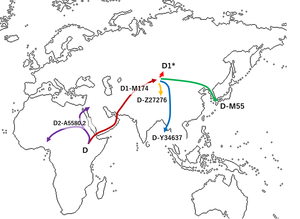

Его филогенетически ближайшие родственники встречаются у народов Японии , Средней Азии и Андаманских островов в Бенгальском заливе . Он более отдаленно связан с гаплогруппой D* , субклады которой распространены по всей Азии .
Распределение
[ редактировать ]Гаплогруппа D-M15 широко распространена среди популяций, населяющих северо-запад, север, северо-восток, восток и юго-восток от Гималаев . Он не встречается среди популяций Индии на юге и юго-западе. Распространение гаплогруппы D1 в Юго-Восточной Азии также весьма ограничено, поскольку она встречается там лишь с низкой частотой и только среди популяций, говорящих на тибето-бирманских или хмонг-миао языках, имеющих наследственные связи с севером.
Распространение гаплогруппы D-M15 гораздо более регулярно на севере, поскольку она встречается почти у всех популяций Центральной и Северо-Восточной Азии к югу от границы с Россией , хотя в целом с низкой частотой - 2% или меньше. Резкий скачок частоты гаплогруппы D-M15 происходит по мере приближения к Цинхай-Тибетскому нагорью в западном Китае : было обнаружено, что среди некоторых местных популяций в Цинхае она достигает 100%. Его частота постепенно снижается по мере продвижения на юг через территорию тибетских народов , поскольку гаплогруппа O3 , которая является наиболее распространенной гаплогруппой среди хмонг-миао, а также обычно встречается среди населения Юго-Восточной Азии , становится доминирующей. Гаплогруппа D-M15 по-прежнему встречается в целом очень редко среди ханьцев на востоке; однако есть некоторые признаки того, что частота D-M15 среди ханьцев может значительно различаться в зависимости от местности.
Что касается окончательного происхождения гаплогруппы D-M15, можно только предполагать, что она могла иметь общего недавнего предка с другими членами более крупной линии D-M174, которые не тестировались как часть другой ветви. Такие встречаются с низкой частотой среди современного населения Центральной Азии .
Филогенетика
[ редактировать ]Филогенетическая история
[ редактировать ]До 2002 года в научной литературе существовало как минимум семь систем наименования филогенетического дерева Y-хромосомы. Это привело к значительной путанице. В 2002 году основные исследовательские группы объединились и сформировали Консорциум Y-хромосомы (YCC). Они опубликовали совместный документ, в котором было создано единое новое дерево, которое все согласились использовать. Позже группа гражданских ученых, интересующихся популяционной генетикой и генетической генеалогией, сформировала рабочую группу для создания любительского дерева, стремясь быть, прежде всего, своевременным. В таблице ниже собраны все эти работы, относящиеся к знаковому дереву YCC 2002 года. Это позволяет исследователю, просматривающему старую опубликованную литературу, быстро перемещаться между номенклатурами.
| YCC 2002/2008 (сокращенно) | (а) | (б) | (с) | (г) | (е) | (г) | (или) | YCC 2002 (от руки) | YCC 2005 (от руки) | YCC 2008 (от руки) | YCC 2010r (от руки) | ИСОГГ 2006 г. | ИСОГГ 2007 | ИСОГГ 2008 | ИСОГГ 2009 | ИСОГГ 2010 | ИЗОГГ 2011 | ИЗОГГ 2012 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Д-М174 | * | * | * | * | * | * | * | * | Д | Д | Д | Д | Д | Д | Д | Д | Д | Д |
| Д-М15 | 4 | IV | 3G | 12 | ЕС5 | Н3 | Б | Д1 | Д1 | Д1 | Д1 | Д1 | Д1 | Д1 | Д1 | Д1 | Д1 | Д1 |
| Д-М55 | * | * | * | * | * | * | * | * | Д2 | Д2 | Д2 | Д2 | Д2 | Д2 | Д2 | Д2 | Д2 | Д2 |
| Д-П12 | 4 | IV | 3G | 11 | ЕС5 | Н2 | Б | Д2а | Д2а | Д2а1а1 | Д2а1а1 | Д2 | Д2 | Д2а1а1 | Д2а1а1 | Д2а1а1 | удаленный | удаленный |
| Д-М116.1 | 4 | IV | 3G | 11 | ЕС5 | Н2 | Б | Д2б* | Д2а | Д2а | Д2а | Д2а | Д2а | Д2а | Д2а | Д2а | удаленный | удаленный |
| Д-М125 | 4 | IV | 3G | 11 | ЕС5 | Н2 | Б | Д2б1 | Д2а1 | Д2а1 | Д2а1 | Д2а1 | Д2а1 | Д2а1 | Д2а1 | Д2а1 | Д2а1 | Д2а1 |
| Д-М151 | 4 | IV | 3G | 11 | ЕС5 | Н2 | Б | Д2б2 | Д2а1 | Д2а2 | Д2а2 | Д2а2 | Д2а2 | Д2а2 | Д2а2 | Д2а2 | Д2а2 | Д2а2 |
Научные публикации
[ редактировать ]Следующие исследовательские группы в соответствии с их публикациями были представлены при создании дерева YCC.
Филогенетические деревья
[ редактировать ]По дереву ISOGG (Версия: 14.151). [ 1 ]
- ДЕЛАТЬ (ДЕЛАТЬ)
- Д (CTS3946)
- D1 (M174/Page30, IMS-JST021355, Гаплогруппа D-M174 )
- Д1а (CTS11577)
- Д1а1 (F6251/Z27276)
- D1a1a (M15) Тибет
- D1a1b (P99) Тибет , Монголия , Средняя Азия
- D1a2 (M64.1/Page44.1, M55) Япония ( Ямато 、 Рюкюань 、 Айны )
- D1a3 (Y34637) Андаманские острова ( народ онге , народ джарава )
- Д1а1 (F6251/Z27276)
- D1b (L1378) Филиппины [ 2 ]
- Д1а (CTS11577)
- D2 (A5580.2) Нигерия , Саудовская Аравия , Сирия
- D1 (M174/Page30, IMS-JST021355, Гаплогруппа D-M174 )
- Д (CTS3946)
См. также
[ редактировать ]Генетика
[ редактировать ]- Африканская примесь в Европе
- Генетическая генеалогия
- Гаплогруппа
- Гаплотип
- Гаплогруппа ДНК Y-хромосомы человека
- Молекулярная филогенетика
- Парагруппа
- Субклад
- Гаплогруппы Y-хромосомы в популяциях мира
- Гаплогруппы Y-ДНК по этническим группам
- Гаплогруппы Y-ДНК в популяциях Восточной и Юго-Восточной Азии
- Гаплогруппы Y-ДНК в популяциях Океании
Субклады Y-ДНК D
[ редактировать ]Основополагающее дерево Y-ДНК
[ редактировать ]Ссылки
[ редактировать ]Источники таблиц преобразования
[ редактировать ]- Капелли, Кристиан; Уилсон, Джеймс Ф.; Ричардс, Мартин; Штумпф, Майкл П.Х.; и др. (февраль 2001 г.). «Отцовское наследие преимущественно коренных народов австронезийскоязычных народов островной Юго-Восточной Азии и Океании» . Американский журнал генетики человека . 68 (2): 432–443. дои : 10.1086/318205 . ПМЦ 1235276 .
- Хаммер, Майкл Ф.; Карафет Татьяна М.; Редд, Алан Дж.; Джарджанази, Хамди; и др. (1 июля 2001 г.). «Иерархические закономерности глобального разнообразия Y-хромосом человека» . Молекулярная биология и эволюция . 18 (7): 1189–1203. doi : 10.1093/oxfordjournals.molbev.a003906 .
- Джоблинг, Марк А.; Тайлер-Смит, Крис (2000), «Новое использование новых гаплотипов», Trends in Genetics , 16 (8): 356–62, doi : 10.1016/S0168-9525(00)02057-6 , PMID 10904265
- Каладжиева, преп.; Калафель, Франческ; Джоблинг, Марк А; Ангеличева, Дора; и др. (февраль 2001 г.). «Закономерности меж- и внутригруппового генетического разнообразия у влакс-рома, выявленные с помощью Y-хромосомы и линий митохондриальной ДНК» . Европейский журнал генетики человека . 9 (2): 97–104. дои : 10.1038/sj.ejhg.5200597 .
- Карафет, Татьяна; Сюй, Липин; Ду, Руофу; Ван, Уильям; и др. (сентябрь 2001 г.). «История отцовского населения Восточной Азии: источники, закономерности и микроэволюционные процессы» . Американский журнал генетики человека . 69 (3): 615–628. дои : 10.1086/323299 . ПМЦ 1235490 .
- Семино, О.; Пассарино, Дж; Оефнер, П.Дж.; Лин, А.А.; и др. (2000), «Генетическое наследие палеолитического Homo sapiens sapiens у современных европейцев: хромосомная перспектива AY», Science , 290 (5494): 1155–9, Бибкод : 2000Sci...290.1155S , doi : 10.1126/science.290.5494. 1155 , PMID 11073453
- Су, Бинг; Сяо, Цзюньхуа; Андерхилл, Питер; Дека, Ранджан; и др. (декабрь 1999 г.). «Y-хромосома свидетельствует о миграции современных людей на север в Восточную Азию во время последнего ледникового периода» . Американский журнал генетики человека . 65 (6): 1718–1724. дои : 10.1086/302680 . ПМЦ 1288383 .
- Андерхилл, Питер А.; Шен, Пейдун; Лин, Алиса А.; Джин, Ли; и др. (ноябрь 2000 г.). «Вариации последовательности Y-хромосомы и история человеческих популяций». Природная генетика . 26 (3): 358–361. дои : 10.1038/81685 .