Эволюционная генетика человека
Эволюционная генетика человека изучает, чем один человеческий геном отличается от другого, эволюционное прошлое, породившее человеческий геном, и его текущие последствия. Различия между геномами имеют антропологические , медицинские , исторические и судебно-медицинские последствия и применения. Генетические данные могут дать важную информацию об эволюции человека .
Происхождение обезьян
[ редактировать ]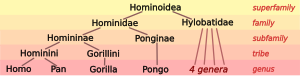
−10 — – −9 — – −8 — – −7 — – −6 — – −5 — – −4 — – −3 — – −2 — – −1 — – 0 — | ( О. praegens ) ( О. тугененсис ) ( Ар. кадабба ) ( Ар. ramidus ) |
| ||||||||||||||||||||||||||||
Биологи относят человека , наряду с лишь несколькими другими видами , к человекообразным обезьянам (видам семейства Hominidae ) . Современные гоминиды включают два различных вида шимпанзе ( бонобо , Pan paniscus и шимпанзе , Pan troglodytes ), два вида горилл ( западная горилла , Gorilla gorilla и восточная горилла , Gorilla graueri ), а также два вида орангутанов. ( борнейский орангутан и Pongo pygmaeus суматранский орангутан ) Pongo abelii . семейства Hylobatidae гиббонов Hominoidea образуют надсемейство человекообразных обезьян Человекообразные обезьяны .
Обезьяны, в свою очередь, относятся к отряду приматов (>400 видов) наряду с обезьянами Старого Света , обезьянами Нового Света и другими. Данные как митохондриальной ДНК (мтДНК), так и ядерной ДНК (нДНК) указывают на то, что приматы принадлежат к группе Euarchontoglires вместе с Rodentia , Lagomorpha , Dermoptera и Scandentia . [1] Это также подтверждается Alu-подобными короткими вкраплениями ядерных элементов (SINE), которые были обнаружены только у представителей Euarchontoglires. [2]
Филогенетика
[ редактировать ]Филогенетическое дерево обычно создается на основе ДНК или белков последовательностей популяций. Часто последовательности митохондриальной ДНК или Y-хромосомы используются для изучения демографических характеристик древнего человека. Эти однолокусные источники ДНК не рекомбинируют и почти всегда наследуются от одного родителя, за одним известным исключением в мтДНК. [3] Люди из более близких географических регионов обычно более похожи, чем люди из более отдаленных регионов. Расстояние на филогенетическом дереве можно использовать приблизительно для обозначения:
- Генетическая дистанция . Генетическая разница между человеком и шимпанзе составляет менее 2%. [4] или в три раза больше, чем вариация среди современных людей (по оценкам, 0,6%). [5]
- Временная удаленность самого недавнего общего предка. По оценкам, самый недавний общий предок современных людей, митохондриальный, жил примерно 160 000 лет назад. [6] последние общие предки человека и шимпанзе, жившие примерно 5–6 миллионов лет назад. [7]
Видообразование человека и африканских обезьян
[ редактировать ]Отделение человека от его ближайших родственников, нечеловеческих африканских обезьян (шимпанзе и горилл), интенсивно изучается уже более столетия. Были рассмотрены пять основных вопросов:
- Какие обезьяны являются нашими ближайшими предками?
- Когда произошло расставание?
- Какова была эффективная численность населения общего предка до разделения?
- Имеются ли следы популяционной структуры (субпопуляций), предшествующие видообразованию, или частичная примесь, следующая за ним?
- Каковы были конкретные события (включая слияние хромосом 2a и 2b) до и после разделения?
Общие замечания
[ редактировать ]Как обсуждалось ранее, разные части генома демонстрируют различное расхождение последовательностей у разных гоминоидов . Также было показано, что расхождение последовательностей ДНК человека и шимпанзе сильно различается. Например, расхождение последовательностей варьируется от 0% до 2,66% между некодирующими, неповторяющимися геномными областями человека и шимпанзе. [8] Процент нуклеотидов в геноме человека (hg38), которые имели точные совпадения с геномом шимпанзе (pantro6), составил 84,38%. Кроме того, генные деревья, созданные путем сравнительного анализа сегментов ДНК, не всегда соответствуют древу видов. Подводя итоги:
- Расхождение последовательностей значительно различается у людей, шимпанзе и горилл.
- По большинству последовательностей ДНК люди и шимпанзе кажутся наиболее близкими родственниками, но некоторые указывают на кладу человек-горилла или шимпанзе-горилла .
- Геном человека секвенирован, как и геном шимпанзе. У человека 23 пары хромосом, а у шимпанзе , горилл и орангутанов — 24. Человеческая хромосома 2 представляет собой слияние двух хромосом 2a и 2b, которые у других приматов остались отдельными. [9]
Время расхождения
[ редактировать ]Большой интерес представляет время отделения человека от других человекообразных обезьян. В одном из первых молекулярных исследований, опубликованном в 1967 году, были измерены иммунологические расстояния (ID) между разными приматами. [10] По сути, в исследовании измерялась сила иммунологического ответа, который антиген одного вида (человеческий альбумин) индуцирует в иммунной системе другого вида (человека, шимпанзе, гориллы и обезьян Старого Света ). Близкородственные виды должны иметь схожие антигены и, следовательно, более слабый иммунологический ответ на антигены друг друга. Иммунологический ответ вида на собственные антигены (например, человека на человека) был установлен равным 1.
Идентификатор между людьми и гориллами был определен как 1,09, а между людьми и шимпанзе - 1,14. Однако расстояние до шести различных обезьян Старого Света составляло в среднем 2,46, что указывает на то, что африканские обезьяны более тесно связаны с людьми, чем с обезьянами. На основании ископаемых данных авторы считают, что время расхождения между обезьянами Старого Света и гоминоидами произошло 30 миллионов лет назад (MYA), а иммунологическое расстояние увеличивалось с постоянной скоростью. Они пришли к выводу, что время расхождения людей и африканских обезьян составляет примерно ~5 млн лет назад. Это был удивительный результат. Большинство учёных того времени считали, что люди и человекообразные обезьяны разошлись гораздо раньше (>15 млн лет назад).
С точки зрения идентификации горилла была ближе к человеку, чем к шимпанзе; однако разница была настолько незначительной, что трихотомию нельзя было определить с уверенностью. Более поздние исследования, основанные на молекулярной генетике, смогли разрешить трихотомию: шимпанзе филогенетически ближе к человеку, чем к гориллам. Однако некоторые времена дивергенции, оцененные позже (с использованием гораздо более сложных методов молекулярной генетики), существенно не отличаются от самой первой оценки 1967 года, но недавняя статья [11] оценивает его в 11–14 млн лет назад.
Время расхождения и предковая эффективная численность популяции
[ редактировать ]
Современные методы определения времени расхождения используют выравнивание последовательностей ДНК и молекулярные часы . Обычно молекулярные часы калибруются исходя из предположения, что орангутан отделился от африканских обезьян (включая человека) 12–16 млн лет назад. Некоторые исследования также включают некоторых обезьян Старого Света и устанавливают время их отделения от гоминоидов на 25-30 млн лет назад. Обе точки калибровки основаны на очень небольшом количестве данных об ископаемых и подверглись критике. [12]
Если эти даты будут пересмотрены, время расхождения, оцененное на основе молекулярных данных, также изменится. Однако относительное время дивергенции вряд ли изменится. Даже если мы не можем точно определить абсолютное время дивергенции, мы можем быть вполне уверены, что время дивергенции между шимпанзе и людьми примерно в шесть раз короче, чем между шимпанзе (или людьми) и обезьянами.
В одном исследовании (Takahata et al. , 1995) использовалось 15 последовательностей ДНК из разных областей генома человека и шимпанзе и 7 последовательностей ДНК человека, шимпанзе и гориллы . [13] Они определили, что шимпанзе более тесно связаны с людьми, чем гориллы. Используя различные статистические методы, они оценили время дивергенции человека и шимпанзе в 4,7 млн лет назад, а время дивергенции между гориллами и людьми (и шимпанзе) — в 7,2 млн лет назад.
Кроме того, они оценили эффективную численность популяции общего предка человека и шимпанзе примерно в 100 000 человек. Это было несколько удивительно, поскольку нынешняя эффективная численность людей оценивается всего в ~ 10 000 человек. Если это правда, это означает, что в ходе эволюции человеческая линия претерпела бы огромное уменьшение эффективного размера популяции (и, следовательно, генетического разнообразия). (см. теорию катастрофы Тоба )

Другое исследование (Chen & Li, 2001) секвенировало 53 неповторяющихся межгенных сегмента ДНК человека, шимпанзе , гориллы и орангутанга . [8] Когда последовательности ДНК были объединены в одну длинную последовательность, созданное дерево объединения соседей поддерживало кладу Homo - Pan со 100% бутстрапом (то есть люди и шимпанзе являются ближайшими родственными видами из четырех). Когда три вида довольно тесно связаны друг с другом (например, человек, шимпанзе и горилла), деревья, полученные на основе данных о последовательностях ДНК, могут не совпадать с деревом, которое представляет видообразование (дерево видов).
Чем короче междоузлия (T IN ), тем чаще встречаются неконгруэнтные генные деревья. Эффективный размер популяции (N e ) межузловой популяции определяет, как долго генетические линии сохраняются в популяции. Более высокий эффективный размер популяции приводит к большему количеству неконгруэнтных генных деревьев. Следовательно, если известен межузловой промежуток времени, можно рассчитать предковую эффективную численность популяции общего предка человека и шимпанзе.
Когда каждый сегмент анализировался индивидуально, 31 поддерживал кладу Человек - Пан , 10 поддерживали кладу Человек - Горилла и 12 поддерживали кладу Пан - Горилла . Используя молекулярные часы, авторы подсчитали, что гориллы разделились сначала 6,2–8,4 млн лет назад, а шимпанзе и люди разделились 1,6–2,2 миллиона лет спустя (межузловой промежуток времени) 4,6–6,2 млн лет назад. Межузловой временной интервал полезен для оценки предкового эффективного размера популяции общего предка человека и шимпанзе.
Скромный анализ показал , что 24 локуса поддерживают кладу Человек - Пан , 7 поддерживают кладу Человек - Горилла , 2 поддерживают кладу Пан - Горилла и 20 не дали никакого разрешения. Дополнительно они взяли из баз данных 35 локусов, кодирующих белки. Из них 12 поддержали кладу Гомо - Пан , 3 — кладу Гомо - Горилла , 4 — кладу Пан - Горилла и 16 не дали никакого решения. Следовательно, только ~70% из 52 локусов, давших разрешение (33 межгенных, 19 кодирующих белки), поддерживают «правильное» дерево видов. Исходя из доли локусов, не поддерживающих древо видов, и рассчитанного ими ранее межузлового интервала времени, эффективная популяция общего предка человека и шимпанзе оценивалась в пределах от ~52 000 до 96 000. Это значение не так велико, как это из первого исследования (Такахата), но все же намного выше, чем современная эффективная численность человеческой популяции.
В третьем исследовании (Янг, 2002) использовался тот же набор данных, что и Чен и Ли, но оценка предковой эффективной популяции составляла «всего» от ~ 12 000 до 21 000, используя другой статистический метод. [14]
Генетические различия между человеком и другими человекообразными обезьянами
[ редактировать ]Человек и шимпанзе идентичны на уровне кодирования на 99,1%, при сходстве на несинонимическом уровне 99,4% и на синонимическом уровне на 98,4%. [15] Выравниваемые последовательности в геномах человека и шимпанзе различаются примерно на 35 миллионов однонуклеотидных замен. Кроме того, около 3% полных геномов различаются делециями, вставками и дупликациями. [16]
Поскольку частота мутаций относительно постоянна, примерно половина этих изменений произошла в человеческом роду. Лишь очень малая часть этих фиксированных различий привела к появлению разных фенотипов человека и шимпанзе, и найти их — большая проблема. Подавляющее большинство различий нейтральны и не влияют на фенотип . [ нужна ссылка ]
Молекулярная эволюция может действовать по-разному: через эволюцию белков, потерю генов, дифференциальную регуляцию генов и эволюцию РНК. Считается, что все они сыграли определенную роль в эволюции человека.
Потеря гена
[ редактировать ]Множество различных мутаций могут инактивировать ген, но немногие из них изменят его функцию определенным образом. Таким образом, инактивационные мутации будут легко доступны для отбора и воздействия. Таким образом, потеря генов может быть распространенным механизмом эволюционной адаптации (гипотеза «меньше значит больше»). [17]
80 генов были потеряны в человеческой линии после отделения от последнего общего предка с шимпанзе. 36 из них относились к обонятельным рецепторам . Гены, участвующие в хеморецепции и иммунном ответе, представлены слишком широко. [18] Другое исследование показало, что 86 генов были потеряны. [19]
Ген кератина волос KRTHAP1
[ редактировать ]Ген кератина волос типа I был утерян в человеческой линии. Кератины являются основным компонентом волос. У людей все еще есть девять функциональных генов кератина волос типа I, но потеря этого конкретного гена могла вызвать истончение волос на теле человека. Основываясь на предположении о постоянных молекулярных часах, исследование предсказывает, что потеря генов произошла относительно недавно в эволюции человека — менее 240 000 лет назад, но и Виндия-Неандерталец, и денисовская последовательность с большим охватом содержат те же преждевременные стоп-кодоны, что и современные. людей и, следовательно, датировка должна быть больше, чем 750 000 лет назад. [20]
Ген миозина MYH16
[ редактировать ]Стедман и др. (2004) заявили, что потеря саркомерного миозина гена MYH16 у человека привела к уменьшению жевательных мышц . По их оценкам, мутация, которая привела к инактивации (удалению двух пар оснований), произошла 2,4 миллиона лет назад, до появления Homo ergaster /erectus в Африке. Последующий период был отмечен сильным увеличением объема черепа , что способствовало предположению, что потеря гена, возможно, устранила эволюционное ограничение на размер мозга у рода Homo . [21]
По другой оценке, потеря гена MYH16 произошла 5,3 миллиона лет назад, задолго до появления человека . [22]
Другой
[ редактировать ]- CASPASE12 — протеиназа цистеиниласпартата. Предполагается, что потеря этого гена снизила летальность бактериальной инфекции у людей. [18]
Добавление гена
[ редактировать ]Сегментарные дупликации (SD или LCR ) сыграли роль в создании новых генов приматов и формировании генетических вариаций человека.
Вставки ДНК, специфичные для человека
[ редактировать ]Когда геном человека сравнили с геномами пяти видов приматов сравнения, включая шимпанзе , гориллу , орангутанга , гиббона и макаку, было обнаружено, что существует около 20 000 специфичных для человека вставок, которые считаются регуляторными. Хотя большинство вставок кажутся нейтральными для фитнеса, небольшое их количество было идентифицировано в положительно отобранных генах, демонстрирующих ассоциации с нервными фенотипами, а некоторые связаны с фенотипами, связанными с зубным и сенсорным восприятием. Эти результаты намекают на, казалось бы, важную роль специфичных для человека вставок в недавней эволюции человека. [23]
Давление выбора
[ редактировать ]Ускоренные области человека — это области генома, которые различаются между людьми и шимпанзе в большей степени, чем это можно объяснить генетическим дрейфом с течением времени, поскольку эти два вида имели общего предка. Эти регионы демонстрируют признаки естественного отбора, что приводит к эволюции явно человеческих черт. Двумя примерами являются HAR1F , который, как полагают, связан с развитием мозга, и HAR2 (он же HACNS1 ), который, возможно, сыграл роль в развитии противопоставленного большого пальца .
Была также выдвинута гипотеза, что большая часть различий между людьми и шимпанзе обусловлена регуляцией экспрессии генов, а не различиями в самих генах. Анализ консервативных некодирующих последовательностей , которые часто содержат функциональные и, следовательно, положительно выбранные регуляторные области, учитывает эту возможность. [24]
Расхождение последовательностей между людьми и обезьянами
[ редактировать ]проект последовательности генома обыкновенного шимпанзе ( Pan troglodytes ), 2400 миллионов оснований (из ~3160 миллионов оснований) были секвенированы и собраны достаточно хорошо, чтобы их можно было сравнить с геномом человека. Когда летом 2005 года был опубликован [16] 1,23% этой секвенации отличались одноосновательными заменами. Считалось, что из них 1,06% или меньше представляют собой фиксированные различия между видами, а остальные представляют собой варианты участков у людей или шимпанзе. Другой тип различий, называемый инделами (вставками/удалениями), обуславливает гораздо меньше различий (на 15% от их количества), но вносит ~1,5% уникальной последовательности в каждый геном, поскольку каждая вставка или делеция может включать от одного основания до миллионов базы. [16]
В сопутствующей статье исследовались сегментные дупликации в двух геномах. [25] вставка и делеция которого в геноме ответственны за большую часть последовательности индел. Они обнаружили, что в общей сложности 2,7% эухроматических последовательностей были дифференциально дублированы в той или иной линии.
| Локус | Человек-шимпанзе | Человек-Горилла | Человек-Орангутан |
|---|---|---|---|
| Алюминиевые элементы | 2 | - | - |
| Некодирующий (Chr. Y) | 1.68 ± 0.19 | 2.33 ± 0.2 | 5.63 ± 0.35 |
| Псевдогены (аутосомные) | 1.64 ± 0.10 | 1.87 ± 0.11 | - |
| Псевдоген (Хр. X) | 1.47 ± 0.17 | - | - |
| Некодирующий (аутосомный) | 1.24 ± 0.07 | 1.62 ± 0.08 | 3.08 ± 0.11 |
| Гены (K s ) | 1.11 | 1.48 | 2.98 |
| Интроны | 0.93 ± 0.08 | 1.23 ± 0.09 | - |
| Xq13.3 | 0.92 ± 0.10 | 1.42 ± 0.12 | 3.00 ± 0.18 |
| Итого по Х-хромосоме | 1.16 ± 0.07 | 1.47 ± 0.08 | - |
| Гены ( Ka ) | 0.8 | 0.93 | 1.96 |
Расхождение последовательностей обычно имеет следующую схему: Человек-шимпанзе < Человек-горилла << Человек-орангутанг, что подчеркивает близкое родство между людьми и африканскими обезьянами. Элементы Alu быстро расходятся из-за высокой частоты динуклеотидов CpG , которые мутируют примерно в 10 раз чаще, чем средний нуклеотид в геноме. Частота мутаций выше в мужской зародышевой линии , поэтому дивергенция в Y-хромосоме , которая наследуется исключительно от отца, выше, чем в аутосомах . наследуется Х-хромосома в два раза чаще по женской зародышевой линии, чем по мужской зародышевой линии, и поэтому имеет несколько меньшую дивергенцию последовательностей. Расхождение последовательностей области Xq13.3 у людей и шимпанзе удивительно низкое. [26]
Мутации, изменяющие аминокислотную последовательность белков (K a ), встречаются реже всего. Фактически ~29% всех ортологичных белков идентичны у человека и шимпанзе. Типичный белок отличается всего двумя аминокислотами. [16] Показатели расхождения последовательностей, показанные в таблице, учитывают только различия в замещении, например, от A ( аденин ) к G ( гуанин ). Однако последовательности ДНК также могут различаться вставками и делециями ( инделами ) оснований. Их обычно удаляют из выравниваний перед выполнением расчета расхождения последовательностей.
Генетические различия между современными людьми и неандертальцами
[ редактировать ]Международная группа ученых завершила проект последовательности генома неандертальца в мае 2010 года. Результаты указывают на некоторое размножение между современными людьми ( Homo sapiens ) и неандертальцами ( Homo neanderthalensis ) , поскольку в геномах неафриканских людей на 1–4% больше общего с неандертальцами, чем геномы африканцев к югу от Сахары. Неандертальцы и большинство современных людей имеют общий непереносимый к лактозе вариант гена лактазы , который кодирует фермент, который не способен расщеплять лактозу в молоке после отъема от груди. Современные люди и неандертальцы также имеют общий вариант гена FOXP2, связанный с развитием мозга и речью у современных людей, что указывает на то, что неандертальцы, возможно, были способны говорить. У шимпанзе есть два аминокислотных различия в FOXP2 по сравнению с FOXP2 человека и неандертальца. [27] [28] [29]
Генетические различия среди современных людей
[ редактировать ]Homo Sapiens Считается, что появился около 300 000 лет назад. Он распространился по всей Африке, а спустя 70 000 лет назад по всей Евразии и Океании.Исследование 2009 года выявило 14 «групп искового населения», самым отдаленным из которых является народ сан в Южной Африке. [30] [31]
Из-за их быстрого распространения в различных климатических зонах и особенно из-за наличия новых источников пищи с одомашниванием крупного рогатого скота и развитием сельского хозяйства , человеческие популяции с момента своего расселения подверглись значительному давлению отбора . Например, считается, что предки жителей Восточной Азии подверглись процессам отбора по ряду аллелей, включая варианты генов EDAR , ADH1B , ABCC1 и ALDH2 .
Восточноазиатские типы ADH1B, в частности, связаны с одомашниванием риса и, таким образом, могли возникнуть после развития выращивания риса примерно 10 000 лет назад. [32] Несколько фенотипических черт, характерных для жителей Восточной Азии, обусловлены единственной мутацией гена EDAR , датируемой c. 35 000 лет назад. [33]
По состоянию на 2017 год [update]База данных однонуклеотидных полиморфизмов ( dbSNP ), в которой перечислены SNP и другие варианты, перечислила в общей сложности 324 миллиона вариантов, обнаруженных в секвенированных геномах человека. [34] Нуклеотидное разнообразие , средняя доля нуклеотидов, которые различаются у двух людей, оценивается в пределах от 0,1% до 0,4% для современных людей (по сравнению с 2% между людьми и шимпанзе). [35] [36] Это соответствует различиям генома в нескольких миллионах сайтов; Проект «1000 геномов» обнаружил аналогичные результаты что «типичный [индивидуальный] геном отличается от эталонного генома человека на 4,1–5,0 миллионов сайтов… затрагивая 20 миллионов оснований последовательности». [37]
В феврале 2019 года ученые обнаружили доказательства, основанные на генетических исследованиях с использованием искусственного интеллекта (ИИ), которые предполагают существование неизвестного вида предков человека, а не неандертальца , денисовца или гибрида человека (например, (гибридного гоминина) ), в геноме Денни современные люди . [38] [39]
Исследования
[ редактировать ]В марте 2019 года китайские ученые сообщили о введении гена MCPH1, связанного с человеческим мозгом , лабораторным макакам-резусам , в результате чего трансгенные обезьяны показали себя лучше и быстрее ответили на «тесты на кратковременную память, включающие сопоставление цветов и форм», по сравнению с контрольными нетрансгенными обезьянами. обезьяны, по мнению исследователей. [40] [41]
В мае 2023 года ученые на основе генетических исследований сообщили о более сложном пути эволюции человека, чем предполагалось ранее. Согласно исследованиям, люди произошли в Африке из разных мест и времен, а не из одного места и периода времени. [42] [43]
31 августа 2023 года исследователи на основе генетических исследований сообщили, что предков человека узкое место в популяции произошло «около 930 000 и 813 000 лет назад… продолжалось около 117 000 лет и привело предков человека на грань исчезновения». [44] [45]
См. также
[ редактировать ]- Проект генома шимпанзе
- FOXP2 и эволюция человека
- Генетика и археогенетика Южной Азии
- Генетическая история Европы
- Генетическая история коренных народов Америки
- Генетическая история Италии
- Генетическая история Британских островов
- Генетическая история Ближнего Востока
- Люди
- Список гаплогрупп исторических и известных личностей
- Путешествие человека: генетическая одиссея
- Гаплогруппы Y-ДНК по этническим группам
- Раса и генетика
- Недавняя эволюция человека
Ссылки
[ редактировать ]- ^ Мерфи, Вашингтон; Эйзирик, Э.; О'Брайен, SJ; Мэдсен, О.; Скалли, М.; Дуади, CJ; Тилинг, Э.; Райдер, ОА; Стэнхоуп, MJ; де Йонг, WW; Спрингер, М.С. (2001). «Разрешение ранней плацентарной радиации млекопитающих с использованием байесовской филогенетики». Наука . 294 (5550): 2348–2351. Бибкод : 2001Sci...294.2348M . дои : 10.1126/science.1067179 . ПМИД 11743200 . S2CID 34367609 .
- ^ Кригс, Дж.О.; Чураков Г.; Кифманн, М.; Джордан, США; Брозиус, Дж.; Шмитц, Дж. (2006). «Ретропозированные элементы как архивы эволюционной истории плацентарных млекопитающих» . ПЛОС Биол . 4 (4): е91. doi : 10.1371/journal.pbio.0040091 . ПМЦ 1395351 . ПМИД 16515367 .
- ^ Шварц М., Виссинг Дж. (2002). «Отцовское наследование митохондриальной ДНК» . N Engl J Med . 347 (8): 576–580. дои : 10.1056/NEJMoa020350 . ПМИД 12192017 .
- ^ « «Человеческая хромосома 2» . PBS » . ПБС . Архивировано из оригинала 31 июля 2018 г. Проверено 31 августа 2017 г.
- ^ По состоянию на 2015 год типичная разница между геномами двух людей оценивалась в 20 миллионов пар оснований (или 0,6% от общего числа 3,2 миллиардов пар оснований):«Типичный [индивидуальный] геном отличается от эталонного человеческого генома на 4,1–5,0 миллионов сайтов [...], затрагивающих 20 миллионов оснований последовательности» Аутон А., Брукс Л.Д., Дурбин Р.М., Гаррисон Э.П., Канг Х.М., Корбел Д.О. и др. (октябрь 2015 г.). «Глобальный справочник по генетическим вариациям человека» . Природа . 526 (7571): 68–74. Бибкод : 2015Natur.526...68T . дои : 10.1038/nature15393 . ПМК 4750478 . ПМИД 26432245 .
- ^ "134 по 188 ка": Фу К., Миттник А., Джонсон П.Л., Бос К., Лари М., Боллонгино Р., Сан С., Гимш Л., Шмитц Р., Бургер Дж., Рончителли А.М., Мартини Ф., Кремонези Р.Г., Свобода Дж., Бауэр П., Карамелли Д., Кастеллано С. , Райх Д., Паабо С., Краузе Дж. (21 марта 2013 г.). «Пересмотренная временная шкала эволюции человека, основанная на древних митохондриальных геномах» . Современная биология . 23 (7): 553–59. Бибкод : 2013CBio...23..553F . дои : 10.1016/j.cub.2013.02.044 . ПМК 5036973 . ПМИД 23523248 . .
- ^ Паттерсон Н., Рихтер Д.Д., Гнерр С., Ландер Э.С., Райх Д. (2006). «Генетические доказательства сложного видообразования человека и шимпанзе». Природа . 441 (7097): 1103–8. Бибкод : 2006Natur.441.1103P . дои : 10.1038/nature04789 . ПМИД 16710306 . S2CID 2325560 .
- ^ Jump up to: а б с Чен, ФК; Ли, WH (2001). «Геномные различия между людьми и другими гоминоидами и эффективный размер популяции общего предка человека и шимпанзе» . Ам Джей Хум Жене . 68 (2): 444–456. дои : 10.1086/318206 . ПМЦ 1235277 . ПМИД 11170892 .
- ^ Кен Миллер в стенограммах судебного разбирательства по делу Китцмиллер против Дувра .
- ^ Сарич, В.М.; Уилсон, AC (1967). «Иммунологическая шкала времени эволюции гоминид». Наука . 158 (3805): 1200–1203. Бибкод : 1967Sci...158.1200S . дои : 10.1126/science.158.3805.1200 . ПМИД 4964406 . S2CID 7349579 .
- ^ Венн, Оливер; Тернер, Исаак; Мэтисон, Иэн; де Гроот, Наташа; Бонтроп, Рональд; Маквин, Гил (июнь 2014 г.). «Сильная предвзятость к самцам приводит к мутациям зародышевой линии у шимпанзе» . Наука . 344 (6189): 1272–1275. Бибкод : 2014Sci...344.1272V . дои : 10.1126/science.344.6189.1272 . ПМЦ 4746749 . ПМИД 24926018 .
- ^ Йодер, AD; Ян, З. (1 июля 2000 г.). «Оценка дат видообразования приматов с использованием локальных молекулярных часов» . Мол Биол Эвол . 17 (7): 1081–1090. doi : 10.1093/oxfordjournals.molbev.a026389 . ПМИД 10889221 .
- ^ Такахата, Н.; Сатта, Ю.; Кляйн, Дж. (1995). «Время расхождения и размер популяции в линии, ведущей к современному человеку» . Теория народной биологии . 48 (2): 198–221. Бибкод : 1995TPBio..48..198T . дои : 10.1006/tpbi.1995.1026 . ПМИД 7482371 .
- ^ Ян, З. (1 декабря 2002 г.). «Оценка вероятности и Байеса размеров предковой популяции гоминоидов с использованием данных из нескольких локусов» (аннотация) . Генетика . 162 (4): 1811–1823. дои : 10.1093/генетика/162.4.1811 . ПМЦ 1462394 . ПМИД 12524351 . Архивировано из оригинала 16 февраля 2011 года . Проверено 25 августа 2006 г.
- ^ Уайлдман, Дерек Э.; Уддин, Моника; Лю, Гочжэнь; Гроссман, Лоуренс И.; Гудман, Моррис (10 июня 2003 г.). «Последствия естественного отбора в формировании 99,4% несинонимической идентичности ДНК между людьми и шимпанзе: увеличение рода Homo» . Труды Национальной академии наук . 100 (12): 7181–7188. Бибкод : 2003PNAS..100.7181W . дои : 10.1073/pnas.1232172100 . ISSN 0027-8424 . ПМК 165850 . ПМИД 12766228 .
- ^ Jump up to: а б с д Секвенирование шимпанзе; Аналитический консорциум (2005). «Исходная последовательность генома шимпанзе и сравнение с геномом человека» . Природа . 437 (7055): 69–87. Бибкод : 2005Natur.437...69. . дои : 10.1038/nature04072 . ПМИД 16136131 .
- ^ Олсон, М.В. (1999). «Когда меньше значит больше: потеря генов как двигатель эволюционных изменений» . Ам Джей Хум Жене . 64 (1): 18–23. дои : 10.1086/302219 . ПМЦ 1377697 . ПМИД 9915938 .
- ^ Jump up to: а б Ван, X.; Грус, МЫ; Чжан, Дж. (2006). «Потери генов во время происхождения человека» . ПЛОС Биол . 4 (3): е52. doi : 10.1371/journal.pbio.0040052 . ПМК 1361800 . ПМИД 16464126 .
- ^ Демут, Джеффри П.; Бие, Тейл Де; Стаич, Джейсон Э.; Кристианини, Нелло; Хан, Мэтью В. (декабрь 2006 г.). Боревиц, Джастин (ред.). «Эволюция генных семейств млекопитающих» . ПЛОС ОДИН . 1 (1): е85. Бибкод : 2006PLoSO...1...85D . дои : 10.1371/journal.pone.0000085 . ПМЦ 1762380 . ПМИД 17183716 .
- ^ Зима, Х.; Лангбейн, Л.; Кравчак, М.; Купер, Д.Н.; Суарес, LFJ; Роджерс, Массачусетс; Прецель, С.; Хайдт, П.Дж.; Швейцер, Дж. (2001). «Псевдоген кератина волос типа I человека phihHaA имеет функциональные ортологи у шимпанзе и гориллы: свидетельства недавней инактивации человеческого гена после дивергенции Pan-Homo». Хум Жене . 108 (1): 37–42. дои : 10.1007/s004390000439 . ПМИД 11214905 . S2CID 21545865 .
- ^ Стедман, Х.Х.; Козяк, Б.В.; Нельсон, А.; Тезье, DM; Вс, LT; Низкий, ДВ; Бриджес, ЧР; Шрагер, Дж.Б.; Первис, Нью-Мексико; Митчелл, Массачусетс (2004). «Мутация гена миозина коррелирует с анатомическими изменениями в человеческом происхождении». Природа . 428 (6981): 415–418. Бибкод : 2004Natur.428..415S . дои : 10.1038/nature02358 . ПМИД 15042088 . S2CID 4304466 .
- ^ Перри, GH; Веррелли, Британская Колумбия; Стоун, AC (2005). «Сравнительный анализ раскрывает сложную историю молекулярной эволюции человека MYH16» . Мол Биол Эвол . 22 (3): 379–382. дои : 10.1093/molbev/msi004 . ПМИД 15470226 .
- ^ Хеллен, Элизабет Х.Б.; Керн, Эндрю Д. (01 апреля 2015 г.). «Роль вставок ДНК в фенотипической дифференциации человека и других приматов» . Геномная биология и эволюция . 7 (4): 1168–1178. дои : 10.1093/gbe/evv012 . ISSN 1759-6653 . ПМЦ 4419785 . ПМИД 25635043 .
- ^ Берд, Кристин П.; Лю, Морин; и др. (2007). «Быстро развивающиеся некодирующие последовательности в геноме человека» . Геномная биология . 8 (6): 118 р. дои : 10.1186/gb-2007-8-6-r118 . ПМК 2394770 . ПМИД 17578567 .
- ^ Ченг, З.; Вентура, М.; Она, Х.; Хаитович П.; Грейвс, Т.; Осоэгава, К.; Церковь, Д; Питер ДеДжонг, П.; Уилсон, РК; Паабо, С.; Рокки, М; Эйхлер, Э.Э. (2005). «Полногеномное сравнение недавних сегментных дупликаций шимпанзе и человека». Природа . 437 (1 сентября 2005 г.): 88–93. Бибкод : 2005Natur.437...88C . дои : 10.1038/nature04000 . ПМИД 16136132 . S2CID 4420359 .
- ^ Кассманн, Х.; Хейсиг, Ф.; фон Хэзелер, А.; Паабо, С. (1999). «Вариация последовательности ДНК в некодирующей области низкой рекомбинации на Х-хромосоме человека». Нат Жене . 22 (1): 78–81. дои : 10.1038/8785 . ПМИД 10319866 . S2CID 9153915 .
- ^ Сай, Тина Хесман (2009). «История первая: команда расшифровывает ДНК неандертальца: черновик генома может раскрыть секреты эволюции человека». Новости науки . 175 (6): 5–7. дои : 10.1002/scin.2009.5591750604 .
- ^ Грин, Ричард Э.; Краузе; Бриггс; Маричич; Стензель; Кирхер; Паттерсон; Ли; Чжай; Фриц; Хансен; Дюран; Маласпинас; Дженсен; Маркес Боне; алкан; экзаменатор; Мейер; Бурбано; Хороший; Шульц; Аксиму Петри; Баттхоф; Хёбер; Хёффнер; Зигемунд; Вейманн; грецкий орех; Страны; и др. (07 мая 2010 г.). «Проект последовательности генома неандертальца» . Наука . 328 (5979): 710–722. Бибкод : 2010Sci...328..710G . дои : 10.1126/science.1188021 . ПМК 5100745 . ПМИД 20448178 .
- ^ «НЕАНДЕРТАЛЬЦЫ ЖИВУТ!» . Блог Джона Хокса . 6 мая 2010 г. Архивировано из оригинала 16 декабря 2010 г. Проверено 31 декабря 2010 г.
- ^ Тишкофф, С.А.; Рид, штат Пенсильвания; Фридлендер, Франция; Эрет, К.; Ранчиаро, А.; Фромант, А.; Хирбо, Дж.Б.; Авомойи, А.А.; и др. (май 2009 г.). «Генетическая структура и история африканцев и афроамериканцев» . Наука . 324 (5930): 1035–44. Бибкод : 2009Sci...324.1035T . дои : 10.1126/science.1172257 . ПМЦ 2947357 . ПМИД 19407144 .
- ↑ BBC World News «Генетические секреты Африки раскрыты». Архивировано 1 июля 2009 г. в Wayback Machine , 1 мая 2009 г.
- ^ Пэн, Ю.; и др. (2010). «Полиморфизм ADH1B Arg47His в популяциях Восточной Азии и расширение одомашнивания риса в истории» . Эволюционная биология BMC . 10 (1): 15. Бибкод : 2010BMCEE..10...15P . дои : 10.1186/1471-2148-10-15 . ПМЦ 2823730 . ПМИД 20089146 .
- ^ Признаки, на которые влияет мутация, - это потовые железы, зубы, толщина волос и ткань молочной железы. Камберов; и др. (2013). «Моделирование недавней эволюции человека у мышей путем экспрессии выбранного варианта EDAR» . Клетка . 152 (4): 691–702. дои : 10.1016/j.cell.2013.01.016 . ПМК 3575602 . ПМИД 23415220 . Восточноазиатские физические черты, связанные с 35 000-летней мутацией. Архивировано 13 августа 2021 г. в Wayback Machine , NYT , 14 февраля 2013 г.
- ^ NCBI (08 мая 2017 г.). «Человеческая сборка dbSNP 150 удвоила количество записей RefSNP!» . Аналитика NCBI . Архивировано из оригинала 08 апреля 2020 г. Проверено 16 мая 2017 г.
- ^ Джорде, LB; Вудинг, СП (2004). «Генетическая изменчивость, классификация и «раса» » . Природная генетика . 36 (11с): С28–33. дои : 10.1038/ng1435 . ПМИД 15508000 .
- ^ Тишкофф, С.А.; Кидд, К.К. (2004). «Значение биогеографии человеческих популяций для расы и медицины» . Природная генетика . 36 (11с): С21–7. дои : 10.1038/ng1438 . ПМИД 15507999 .
- ^ Консорциум проекта «1000 геномов» (01 октября 2015 г.). «Глобальный справочник по генетическим вариациям человека» . Природа . 526 (7571): 68–74. Бибкод : 2015Natur.526...68T . дои : 10.1038/nature15393 . ISSN 0028-0836 . ПМК 4750478 . ПМИД 26432245 .
{{cite journal}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Мондал, Маюх; Бертранпедт, Жауме; Лео, Оскар (16 января 2019 г.). «Приблизительные байесовские вычисления с глубоким обучением подтверждают третью архаичную интрогрессию в Азии и Океании» . Природные коммуникации . 10 (246): 246. Бибкод : 2019NatCo..10..246M . дои : 10.1038/s41467-018-08089-7 . ПМК 6335398 . ПМИД 30651539 .
- ^ Докрилл, Питер (11 февраля 2019 г.). «Искусственный интеллект обнаружил в геноме человека неизвестного предка-призрака» . ScienceAlert.com . Архивировано из оригинала 23 апреля 2022 года . Проверено 11 февраля 2019 г.
- ^ Баррелл, Тил (29 декабря 2019 г.). «Ученые ввели ген человеческого интеллекта в обезьяну. Другие учёные обеспокоены» . Обнаружить . Архивировано из оригинала 30 декабря 2019 года . Проверено 30 декабря 2019 г.
- ^ Ши, Лей; и др. (27 марта 2019 г.). «Трансгенные макаки-резусы, несущие копии человеческого гена MCPH1, демонстрируют человеческую неотению развития мозга» . Китайский национальный научный обзор . 6 (3): 480–493. дои : 10.1093/nsr/nwz043 . ПМЦ 8291473 . ПМИД 34691896 . Архивировано из оригинала 4 октября 2022 года . Проверено 30 декабря 2019 г.
- ^ Циммер, Карл (17 мая 2023 г.). «Исследование предлагает новый взгляд на то, как развивались первые люди. Новый генетический анализ 290 человек предполагает, что люди появились в разное время и в разных местах Африки» . Нью-Йорк Таймс . Архивировано из оригинала 17 мая 2023 года . Проверено 18 мая 2023 г.
- ^ Рэгсдейл, Варон П.; и др. (17 мая 2023 г.). «Слабо структурированная основа происхождения человека в Африке» . Природа . 167 (7962): 755–763. Бибкод : 2023Natur.617..755R . дои : 10.1038/s41586-023-06055-y . ПМЦ 10208968 . ПМИД 37198480 .
- ^ Циммер, Карл (31 августа 2023 г.). «Предки человечества почти вымерли, как предполагает генетическое исследование. Численность населения сократилась в результате изменения климата около 930 000 лет назад, заключили ученые. Других экспертов этот анализ не убедил» . «Нью-Йорк Таймс» . Архивировано из оригинала 31 августа 2023 года . Проверено 2 сентября 2023 г.
- ^ Ху, Ванцзе; и др. (31 августа 2023 г.). «Геномный вывод о серьезном узком месте человека во время перехода от раннего к среднему плейстоцену» . Наука . 381 (6661): 979–984. Бибкод : 2023Sci...381..979H . дои : 10.1126/science.abq7487 . ПМИД 37651513 . S2CID 261396309 . Архивировано из оригинала 1 сентября 2023 года . Проверено 2 сентября 2023 г.
Дальнейшее чтение
[ редактировать ]- Джоблинг, Марк А.; Холлокс, Эдвард; Херлс, Мэтью; Кивисилд, Тоомас; Тайлер-Смит, Крис (2013). Эволюционная генетика человека . Нью-Йорк: Garland Science. ISBN 978-0-8153-4148-2 . OCLC 829099073 .
- Раннала Б., Ян З. (август 2003 г.). «Байесовская оценка времени дивергенции видов и размеров предковой популяции с использованием последовательностей ДНК из нескольких локусов» . Генетика . 164 (4): 1645–56. дои : 10.1093/генетика/164.4.1645 . ПМЦ 1462670 . ПМИД 12930768 .