Генетическая история Пиренейского полуострова

Происхождение современных иберийцев (включая испанцев и португальцев ) соответствует географическому положению Пиренейского полуострова в юго-западной части Европы , демонстрируя характеристики, которые в значительной степени типичны для южных и западных европейцев. Как и в случае с большей частью остальной части Южной Европы, основным предком современных иберийцев являются ранние европейские земледельцы , прибывшие в эпоху неолита. Значительное преобладание гаплогруппы R1b Y-хромосомы, распространенной по всей Западной Европе , также является свидетельством значительного вклада различных волн (преимущественно мужчин) западных степных пастухов , которые возникли в Понтийско-Каспийской степи во время бронзового века . [2] [3]
Генетическое наследование современных иберийцев во многом происходит от доримских жителей Пиренейского полуострова:
- Доиндоевропейские и индоевропейскоязычные докельтские группы : ( иберы , лузитаны , веттоны , турдетаны , аквитаны , конии ). [4] [5] [6]
- Кельты ( галлеки , кельтиберы , турдули и кельты ), [7] [6] которые были латинизированы после завоевания региона древними римлянами . [8] [9]
Есть также незначительное генетическое влияние германских племен, прибывших в период раннего средневековья. [10] [11] [12] Благодаря своему положению на Средиземном море , как и другие страны Южной Европы , здесь также были контакты с другими средиземноморскими народами, такими как древние финикийцы , греки и карфагеняне, которые на короткое время поселились вдоль восточного и южного побережья Иберии, еврейская община сефардов , а также берберы и Арабы прибыли во времена Аль-Андалуса , и все они оставили после себя некоторое генетическое наследие Северной Африки и Ближнего Востока , особенно на юге и западе Пиренейского полуострова . [13] [14] [9] [15] [16] [17] [8] Подобно Сардинии , Иберия была защищена от заселения со стороны Ближнего Востока и Кавказа своим западным географическим положением и, таким образом, имеет более низкий уровень примеси Западной Азии и Ближнего Востока , чем Италия и Греция , большая часть которых, вероятно, прибыла в Иберию в исторический, а не исторический период. доисторические времена, особенно в римский период . [18] [19]
Популяционная генетика: методы и ограничения
[ редактировать ]Первым пионером изучения популяционной генетики был Луиджи Лука Кавалли-Сфорца . Кавалли-Сфорца использовал классические генетические маркеры для косвенного анализа ДНК. Этот метод изучает различия в частотах определенных аллельных признаков, а именно полиморфизмов белков, обнаруженных в крови человека (таких как группы крови АВО , антигены резус-крови, локусы HLA , иммуноглобулины , изоферменты G-6-PD и другие). Впоследствии его команда рассчитала генетические расстояния между популяциями, основываясь на том принципе, что две популяции, которые имеют схожие частоты признака, более тесно связаны, чем популяции, которые имеют более разные частоты этого признака. [20]
С тех пор популяционная генетика значительно продвинулась вперед, и исследования с использованием прямого анализа ДНК теперь многочисленны и могут использовать митохондриальную ДНК ( мтДНК ), нерекомбинирующую часть Y-хромосомы ( NRY ) или аутосомную ДНК. МтДНК и ДНК NRY имеют некоторые общие черты, которые сделали их особенно полезными в генетической антропологии. Эти свойства включают прямое, неизмененное наследование мтДНК и ДНК NRY от матери к потомству и от отца к сыну, соответственно, без эффектов «скремблирования» генетической рекомбинации . Мы также предполагаем, что на эти генетические локусы не влияет естественный отбор и что основным процессом, ответственным за изменения пар оснований, является мутация (которую можно вычислить). [21]
В то время как гаплогруппы Y-ДНК и мтДНК представляют собой лишь небольшой компонент пула ДНК человека, аутосомная ДНК имеет то преимущество, что содержит сотни и тысячи исследуемых генетических локусов, что дает более полную картину генетического состава. Родственные связи можно определить только на статистической основе, поскольку аутосомная ДНК подвергается рекомбинации. Одна хромосома может записать историю каждого гена. Аутосомные исследования гораздо более надежны для демонстрации взаимоотношений между существующими популяциями, но не предлагают возможностей для раскрытия их истории так, как обещают исследования мтДНК и ДНК NRY, несмотря на их многочисленные сложности. [ нужна ссылка ]
Анализы ядерной и древней ДНК
[ редактировать ]
Анализ ядерной ДНК показывает, что испанское и португальское население наиболее тесно связано с другими популяциями Западной Европы. [22] [23] [24] Существует ось значительной генетической дифференциации в направлении восток-запад, в отличие от замечательного генетического сходства в направлении север-юг. Примесь Северной Африки, связанная с исламским завоеванием , может быть датирована периодом между ок. 860–1120 гг. н.э. [25]
Исследование, опубликованное в 2019 году с использованием образцов 271 иберийца, охватывающих доисторические и исторические времена, предлагает следующие переломные точки в геномной истории Иберии: [26]
- Мезолит : охотники-собиратели из европейских степей Западной России, Грузии и Украины — первые люди, заселившие северо-запад Пиренейского полуострова.
- Неолит : неолитические фермеры заселяют весь Пиренейский полуостров из Анатолии .
- Эпоха энеолита : приток охотников-собирателей из Центральной Европы и некоторый приток генов в результате спорадических контактов с Северной Африкой .
- Бронзовый век : приток степей из Центральной Европы.
- Железный век генетический фонд басков остается в основном нетронутым. : дополнительный поток степных генов из Центральной Европы - с этого момента
- Римский период : генетический приток из Центрального и Восточного Средиземноморья . Некоторый дополнительный приток североафриканских генов обнаружен в Южной Иберии.
- Вестготский период : заметные притоки.
- Мусульманский период : приток из Северной Африки. После Реконкисты произошло дальнейшее генетическое сближение между Северной и Южной Иберией.
Влияние Северной Африки
[ редактировать ]
Ряд исследований был сосредоточен на выяснении генетического влияния исторических перемещений населения Северной Африки в Иберию на генетический состав современного испанского и португальского населения. Первоначальные исследования показали, что Гибралтарский пролив в доисторические времена действовал скорее как генетический барьер, чем мост. [27] [28] [29] в то время как другие исследования указывают на более высокий уровень недавней примеси из Северной Африки среди иберийцев, чем среди других европейских популяций, [30] [31] [32] [33] [34] [35] [36] хотя это является результатом более поздних миграционных движений, в частности мавританского вторжения в Иберию в 8 веке.
Что касается аутосомной ДНК, самое последнее исследование африканской примеси в иберийском населении было проведено в апреле 2013 года Botigué et al. используя полногеномные данные SNP для более чем 2000 человек из Европы, Магриба, Катара и стран к югу от Сахары, из которых 119 были испанцами и 117 португальцами, можно сделать вывод, что Испания и Португалия имеют значительный уровень североафриканского происхождения. По оценкам общего происхождения, в среднем от 4% в некоторых местах до 10% среди населения в целом; население Канарских островов имеет от 0% до 96% общего происхождения с выходцами из Северной Африки, хотя Канарские острова являются испанским эксклавом, расположенным на африканском континенте, и, таким образом, этот результат не репрезентативен для иберийского населения; эти же результаты не превышали 2% в других популяциях Западной и Южной Европы. [37] [38] [39] [40] Однако, в отличие от прошлых аутосомных исследований и того, что следует из частот Y-хромосомы и митохондриальных гаплотипов (см. ниже), он не обнаруживает значительных уровней происхождения из стран южнее Сахары ни в одной европейской популяции за пределами Канарских островов. Действительно, предыдущее аутосомное исследование 2011 года, проведенное Moorjani et al. обнаружил, что родословная из стран южнее Сахары во многих частях южной Европы находится в пределах 1-3%, «самая высокая доля африканского происхождения в Европе приходится на Иберию (Португалия 4,2 ± 0,3% и Испания 1,4 ± 0,3%), что согласуется с выводами, основанными на на митохондриальной ДНК и Y-хромосомах, а также наблюдение Auton et al., что в Европе жители Юго-Западной Европы имеют самый высокий уровень общих гаплотипов с жителями Северной Африки». [30] [34] [35]
Недавние исследования показывают незначительные связи между некоторыми иберийскими регионами и населением Северной Африки в результате исторического периода Аль-Андалуса , который в Португалии длился между 8 и 12 веками нашей эры, а на юге Испании - до конца 15 века нашей эры. Иберия - это европейский регион, в котором более заметно присутствие гаплогруппы E3b Y-хромосомы человека (E-M81). [41] гаплогруппы U (U6) и гаплотипа Va, и это может быть результатом некоторой первоначальной общей популяции западного Средиземноморья. В Португалии североафриканские гаплогруппы Y-хромосомы (особенно те, которые типично северо-западноафриканские) встречаются с частотой 7,1%. [42] Некоторые исследования митохондриальной ДНК также находят свидетельства существования североафриканской гаплогруппы U6, особенно в северной Португалии. [43] Хотя частота U6 низкая (4–6%), было подсчитано, что примерно 27% населения северной Португалии имели североафриканское происхождение, поскольку U6 также не является распространенной линией в Северной Африке. [44] Согласно некоторым исследованиям, североафриканские и арабские элементы в происхождении сегодняшних иберийцев более чем тривиальны по сравнению с основой доисламского происхождения, а Гибралтарский пролив, похоже, функционирует скорее как генетический мост, чем барьер. [45] [46] [47]
Однако исследование, в котором использовались различные генетические маркеры, пришло к другим выводам. В аутосомном исследовании Spínola et al. (2005), которые проанализировали человеческий лейкоцитарный антиген (гены HLA) (присущий всем предкам по прямой отцовской и материнской линии) у сотен людей из Португалии, показали, что португальское население находится под влиянием других европейцев и североафриканцев через многие древние миграции. По мнению авторов, Север и Юг Португалии демонстрируют большее сходство с жителями Северной Африки, в отличие от жителей центра страны, которые кажутся более близкими к другим европейцам, поскольку Север Португалии, по-видимому, сконцентрирован, конечно, из-за под давлением арабской экспансии, древнего генетического полюса, происходящего от многих североафриканцев и других европейцев, влиявшего на протяжении тысячелетий, [ нужны разъяснения ] в то время как южная Португалия демонстрирует генетическое влияние Северной Африки, вероятно, результат недавнего происхождения от амазигов , которые сопровождали арабскую экспансию. [48]

Согласно исследованию, опубликованному в Американском журнале генетики человека в декабре 2008 года, 30% современных португальцев (23,6% на севере и 36,3% на юге) имеют ДНК , которая показывает, что они имеют мужское сефардско-еврейское происхождение, а 14% (11,8% в Север и 16,1% на Юге) имеют мавританское происхождение. [49] Несмотря на возможные альтернативные источники происхождения, приписываемые сефардскому еврейскому происхождению, эти пропорции свидетельствовали о важности религиозного обращения (добровольного или принудительного), о чем свидетельствуют исторические эпизоды социальной и религиозной нетерпимости .

Что касается ДНК отцовской Y-хромосомы, недавние исследования совпадают в том, что в Иберии наблюдается наибольшее присутствие типичного для Северо-Западной Африки маркера гаплотипа Y-хромосомы E-M81 в Европе, в среднем 3%. [31] [32] а также гаплотип Va. [51] [33] Оценки происхождения Y-хромосомы различаются: исследование 2008 года, опубликованное в Американском журнале генетики человека, с использованием 1140 образцов со всего Пиренейского полуострова, дает 10,6% североафриканского происхождения. [34] [35] [36] к отцовскому составу иберов. Аналогичное исследование Y-хромосомы, проведенное в 2009 году с 659 образцами из Южной Португалии, 680 из Северной Испании, 37 образцами из Андалусии, 915 образцами из материковой Италии и 93 образцами из Сицилии, выявило значительно более высокие уровни мужского североафриканского происхождения в Португалии, Испании и других странах. Сицилия (7,1%, 7,7% и 7,5% соответственно), чем в полуостровной Италии (1,7%). [31]
Другие исследования иберийского генофонда выявили значительно более низкий уровень североафриканского происхождения. По данным Боша и др. 2000 «Население северо-западной Африки, возможно, содержало 7% иберийских Y-хромосом». [28] Широкомасштабное исследование Cruciani et al. В 2007 году, используя 6501 несвязанный образец Y-хромосомы из 81 популяции, было обнаружено, что: «Учитывая обе эти субгаплогруппы E-M78 (E-V12, E-V22, E-V65) и гаплогруппу E-M81, вклад североафриканских линии ко всему мужскому генофонду Иберии (за исключением Пасиегоса), континентальной Италии и Сицилии можно оценить как 5,6 процента, 3,6 процента и 6,6 процента соответственно». [52] Исследование 2007 года оценило вклад североафриканских линий во весь мужской генофонд Иберии в 5,6%. [53] В общих чертах, согласно (Bosch et al. 2007) «...происхождение иберийского пула Y-хромосом можно резюмировать следующим образом: 5% недавних северо-западных африканских, 78% верхнепалеолитических и более поздних местных производных (группа IX) и 10% неолит» (H58, H71). [54]
Исследования митохондриальной ДНК, проведенные в 2003 году, совпадают в том, что на Пиренейском полуострове наблюдаются более высокие уровни типично североафриканского гаплотипа U6, [35] [55] а также более высокие частоты африканской гаплогруппы L к югу от Сахары в Португалии. [56] [57] [58] [59] Высокие частоты в основном сконцентрированы на юге и юго-западе Пиренейского полуострова, поэтому общая частота выше в Португалии (7,8%), чем в Испании (1,9%), при средней частоте по всему полуострову 3,8%. На полуострове наблюдаются значительные географические расхождения, при этом высокие частоты наблюдаются в Западной Андалусии (14,6%). [59] и Кордова (8,3%), [56] Южная Португалия (10,7%), Юго-Западная Кастилия (8%). Адамс и др. и другие предыдущие публикации предполагают, что оккупация оставила несовершеннолетнего еврея мавританская Сакалибу [60] и некоторое арабско-берберское генетическое влияние, главным образом, в южных регионах Иберии. [61] [34]
Самые последние и всесторонние геномные исследования показывают, что североафриканское генетическое происхождение можно выявить на большей части Пиренейского полуострова в диапазоне от 0% до 11%, но оно наиболее высоко на юге и западе, а в Стране Басков оно отсутствует или почти отсутствует. и северо-восток. [62] [18] [19]
Текущие дебаты вращаются вокруг того, связано ли присутствие U6 с исламской экспансией на Пиренейский полуостров или с предшествующими перемещениями населения. [34] [35] [36] и связана ли гаплогруппа L с работорговлей или предыдущими перемещениями населения, связанными с исламской экспансией. Большинство линий гаплогруппы L в Иберии, происходящих из Северной Африки, указывают на последнее. [56] [58] [30] [35] [63] В 2015 году Эрнандес и др. пришел к выводу, что «предполагаемый вход североафриканских линий U6 в Иберию в 10 тысячелетии хорошо коррелирует с другими L-африканскими кладами, указывая на то, что некоторые линии U6 и L переместились вместе из Африки в Иберию в раннем голоцене , в то время как большинство из них были завезены в исторические времена. ." [64]
Гаплогруппы
[ редактировать ]
Гаплогруппы Y-хромосомы
[ редактировать ]Y-ДНК Как и у других жителей Западной Европы, среди испанцев и португальцев гаплогруппа R1b является наиболее частой и встречается более чем в 70% на большей части территории Испании. [65] R1b особенно доминирует в Стране Басков и Каталонии, где его частота составляет более 80%. В Иберии большинство мужчин с R1b принадлежат к субкладу R-P312 (R1b1a1a2a1a2; по состоянию на 2017 г.). Распространение гаплогрупп, отличных от R1b, широко варьируется от одного региона к другому.
В Португалии в целом гаплогруппы R1b составляют 70%, а в некоторых регионах северо-запада - более 90%. [66]
Хотя R1b преобладает в большей части Западной Европы, ключевое различие обнаруживается в распространенности R-DF27 (R1b1a1a2a1a2a) в Иберии. Этот субклад встречается более чем у 60% мужского населения в Стране Басков и у 40-48% в Мадриде, Аликанте, Барселоне, Кантабрии, Андалусии, Астурии и Галисии. [67] R-DF27 составляет гораздо больше половины от общего количества R1b на Пиренейском полуострове. Последующая иммиграция представителей других гаплогрупп и субкладов R1b не повлияла на его общую распространенность, хотя она составляет лишь две трети от общего числа R1b в Валенсии и на побережье в целом. [65] R-DF27 также является важным субкладом R1b в некоторых частях Франции и Великобритании. R-S28/R-U152 (R1b1a1a2a1a2b) является преобладающим субкладом R1b в Северной Италии, Швейцарии и некоторых частях Франции, но он составляет менее 5,0% мужского населения Иберии. древние образцы центральноевропейской культуры колокольных кубков , гальштатской культуры и культуры курганов . К этому субкладу принадлежали [68] [69] [70] R-S28/R-U152 незначительно значим в Севилье , Барселоне , Португалии и Стране Басков и составляет 10-20% от общей численности населения, но он представлен с частотой всего 3,0% в Кантабрии и Сантандере , 2,0% в Кастилии и Леоне. , 6% в Валенсии и менее 1% в Андалусии . [65] Евреи-сефарды I1 0% I2*/I2a 1% I2 0% Гаплогруппа R 1a 5% R1b 13% G 15% Гаплогруппа J2 2 25% J */J1 22% E-M215 1b1b 9% T 6% Q 2% [71]
Гаплогруппа J , в основном субклады гаплогруппы J-M172 (J2), встречается на уровне более 20% в некоторых регионах, тогда как гаплогруппа E имеет общую частоту около 10%, хотя в некоторых областях пики превышают 30%. В целом, E-M78 (E1b1b1a1 в 2017 году) и E-M81 (E1b1b1b1a в 2017 году) составляют около 4,0% каждая, еще 1,0% приходится на гаплогруппу E-M123 (E1b1b1b2a1) и 1,0% - на неизвестные субклады E-M96. [34] (Широко считается, что E-M81 представляет собой относительно историческую миграцию из Северной Африки).
Митохондриальная ДНК
[ редактировать ]
В Европе был проведен ряд исследований гаплогрупп митохондриальной ДНК (мтДНК). В отличие от гаплогрупп Y-ДНК, гаплогруппы мтДНК не имели такого большого географического паттерна, но были более равномерно распространены. За исключением отдаленных саамов всех европейцев характерно преобладание гаплогрупп H , U и T. , для Отсутствие заметной географической структуризации мтДНК может быть связано с социокультурными факторами, а именно с патрилокальностью и отсутствием полиандрии . [73]
Субгаплогруппы H1 и H3 подверглись более детальному изучению и могут быть связаны с мадленской экспансией из Иберии c. 13 000 лет назад: [56]
- H1 охватывает важную часть западноевропейской мтДНК, достигая своего локального пика среди современных басков (27,8%) и с высокой частотой встречаясь среди других иберийцев и жителей Северной Африки . Его частота превышает 10% во многих других частях Европы ( Франция , Сардиния , Британские острова , Альпы, значительная часть Восточной Европы ) и выше 5% почти на всем континенте. Его субклад H1b наиболее распространен в Восточной Европе и северо-западе Сибири. [74] частота H1 — 61% — обнаружена среди туарегов региона Феццан На данный момент самая высокая в Ливии . [75]
- H3 представляет собой меньшую часть европейского генома, чем H1, но имеет несколько похожее распределение с пиком среди басков (13,9%), галисийцев (8,3%) и сардинцев (8,5%). Однако его частота снижается к северо-востоку от континента. Исследования показали, что гаплогруппа H3 обеспечивает высокую защиту от прогрессирования СПИДа. [76]
Общеевропейское исследование 2007 года, включающее испанских басков и валенсийских испанцев, показало, что иберийское население группируется дальше всего от других континентальных групп, подразумевая, что Иберия имеет самое древнее европейское происхождение. В этом исследовании было обнаружено, что наиболее заметная генетическая стратификация в Европе проходит с севера на юго-восток, в то время как другая важная ось дифференциации проходит через континент с востока на запад. Было также обнаружено, что, несмотря на различия, все европейцы тесно связаны между собой. [77]
Субрегионы
[ редактировать ]Испания
[ редактировать ]| Область | Размер выборки | С | И | Г | я | J2 | JxJ2 | Р1а | Р1б | Примечания |
|---|---|---|---|---|---|---|---|---|---|---|
| Арагон | 34 | 6% | 0% | 18% | 12% | 0% | 3% | 56% | ||
| Андалусия Восток | 95 | 4% | 3% | 6% | 9% | 3% | 1% | 72% | ||
| Андалусия Запад | 73 | 15% | 4% | 5% | 14% | 1% | 4% | 54% | ||
| Астурия | 20 | 15% | 5% | 10% | 15% | 0% | 0% | 50% | ||
| баски | 116 | 1% | 0% | 8% | 3% | 1% | 0% | 87% | ||
| Кастилия Ла Манча | 63 | 4% | 10% | 2% | 6% | 2% | 2% | 72% | ||
| Кастилия Северо-Восток | 31 | 9% | 3% | 3% | 3% | 0% | 0% | 77% | ||
| Кастилия Северо-Запад | 100 | 19% | 5% | 3% | 8% | 1% | 2% | 60% | ||
| Каталония | 80 | >0% [79] | 3% | 6% | 3% | 6% | 0% | 0% | 81% | |
| Эстремадура | 52 | 18% | 4% | 10% | 12% | 0% | 0% | 50% | ||
| Галисия | 88 | 17% | 6% | 10% | 7% | 1% | 0% | 57% | ||
| Валенсия | 73 | >0% [80] | 10% | 1% | 10% | 5% | 3% | 3% | 64% | |
| Майорка | 62 | 9% | 6% | 8% | 8% | 2% | 0% | 66% | ||
| Менорка | 37 | 19% | 0% | 3% | 3% | 0% | 3% | 73% | ||
| Ибица | 54 | 8% | 13% | 2% | 4% | 0% | 0% | 57% | ||
| Севилья | 155 | 7% | 4% | 12% | 8% | 3% | 1% | 60% | ||
| Уэльва | 22 | 14% | 0% | 9% | 14% | 0% | 0% | 59% | ||
| Кадис | 28 | 4% | 0% | 14% | 14% | 4% | 0% | 51% | ||
| Кордова | 27 | 11% | 0% | 15% | 15% | 0% | 0% | 56% | ||
| Малага | 26 | 31% | 4% | 0% | 15% | 0% | 8% | 43% | ||
| Леон | 60 | 10% | 7% | 3% | 5% | 2% | 7% | 62% | ||
| Кантабрия | 70 | 13% | 9% | 6% | 3% | 3% | 4% | 58% |
Португалия
[ редактировать ]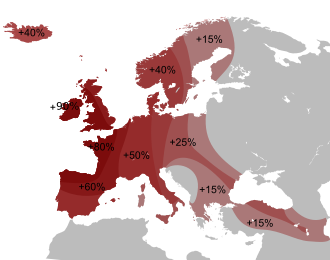
Выдержки из реферата опубликованного исследования [81] в 2015 году:
«[...] В случае с Португалией предыдущие исследования популяционной генетики уже выявили общий портрет разнообразия митохондрий HVS-I и HVS-II, и теперь становится важным обновить и расширить анализируемую митохондриальную область. Соответственно, в общей сложности было получено 292 полных последовательности контрольных регионов из континентальной Португалии в рамках строгого экспериментального плана, обеспечивающего качество данных посредством двойного секвенирования каждого целевого региона.* Кроме того, были изучены SNP H-специфических кодирующих регионов для детальной классификации гаплогрупп. и полные митогеномы были получены для всех последовательностей, принадлежащих гаплогруппам U4 и U5. В целом, типичная западноевропейская гаплогруппа или состав атлантического модального гаплотипа (АМГ) был обнаружен на материковой части Португалии, что связано с высоким уровнем генетического разнообразия митохондрий. Внутри страны никаких признаков субструктуры обнаружено не было. Типирование SNP дополнительных кодирующих областей обеспечило уточнение или подтверждение предыдущей классификации, полученной с помощью инструмента EMMA, в 96% случаев. Наконец, стало возможным расширить филогению гаплогруппы U за счет 28 новых митогеномов U4 и U5».
АМГ достигает самых высоких частот на Пиренейском полуострове, в Великобритании и Ирландии. На Пиренейском полуострове он достигает 70% в Португалии в целом, более 90% на северо-западе Португалии и почти 90% в Галисии (северо-запад Испании), а самый высокий показатель наблюдается у басков (северо-запад Испании).
Атлантический модальный гаплотип ) или гаплотип 15 представляет собой Y-хромосомы гаплотип микросателлитных вариаций Y-STR ( AMH , связанный с гаплогруппой R1b . Он был обнаружен раньше многих SNP, которые сейчас используются для идентификации субкладов R1b, и ссылки на него можно найти в некоторой старой литературе. Наиболее близко он соответствует субкладу R1b1a2a1a(1) [L11].
АМГ является наиболее часто встречающимся гаплотипом среди мужчин мужского пола в Атлантической Европе . Для него характерны следующие маркерные аллели :
- ДИС388 12
- ДИС390 24
- ДИС391 11
- ДИС392 13
- ДИС393 13
- DYS394 14 (также известный как DYS19)
См. также
[ редактировать ]- Генетическая история Европы
- Генетическая история Северной Африки
- Генетические исследования евреев
- Генетические исследования арабов
Ссылки
[ редактировать ]- ^ Пимента Дж., Лопес А.М., Карраседо А., Аренас М., Аморим А., Комас Д. (2019). «Пространственно-явный анализ выявляет сложные генетические градиенты человека на Пиренейском полуострове» . Научный представитель . 9 (1): 7825. Бибкод : 2019НатСР...9.7825П . дои : 10.1038/s41598-019-44121-6 . ПМЦ 6534591 . ПМИД 31127131 .
- ^ Нелис, Мэри; Эско, Тоне; Магия, Ридик; Зимприх, Фриц; Зимприч, Александр; Тончева, Драга; Карачанак, Сена; Пискачкова, Тереза; Балащак, Иван; Пелтонен, Лина; Джаккула, Эвелин; Ренстрём, Карола; Латроп, Марк; Хит, Саймон; Галан, Пилар; Шрайбер, Стефан; Мейтингер, Томас; Пфойфер, Арне; Вихманн, Х-Эрих; Мелег, Бела; Полгар, Ноэми; Тониоло, Даниэла; Гаспарини, Паоло; Д'Адамо, Пио; Кловинс, Дженис; Никитина-Заке, Лиене; Кучинская, Вайдути; Каснаускене, Юрате; Любински, Джон; Дебняк, Тадеуш; Лимборская, Светлана; Хрунин Андрей; Эстивилл, Ксавье; Рабионет, Ракель; Марсал, Сара; Хулия, Антонио; Антонаракис, Стилианос Э.; Дойч, Сэмюэл; Борель, Кристель; Аттар, Хома; Ганнебин, Мэрилин; Мацек, Милан; Кравчак, Майкл; Ремм, Майдо; Мецпалу, Андрей; Флейшер, Роберт К. (8 мая 2009 г.). «Генетическая структура европейцев: взгляд с северо-востока» . ПЛОС ОДИН . 4 (5): е5472. Бибкод : 2009PLoSO...4.5472N . дои : 10.1371/journal.pone.0005472 . ПМК 2675054 . ПМИД 19424496 .
- ^ Новембре, Джон; Джонсон, Тоби; Брик, Катажина; Куталик, Золтан; Бойко, Адам Р.; Аутон, Адам; Индап, Амит; Кинг, Карен С.; Бергманн, Свен; Нельсон, Мэтью Р.; Стивенс, Мэтью; Бустаманте, Карлос Д. (31 августа 2008 г.). «Гены отражают географию Европы» . Природа . 456 (7218): 98–101. Бибкод : 2008Natur.456...98N . дои : 10.1038/nature07331 . ПМК 2735096 . ПМИД 18758442 .
- ^ «Иберийцы — MSN Encarta» . Encarta.msn.com . Архивировано из оригинала 30 октября 2009 года . Проверено 12 января 2022 г.
- ^ Альварес-Санчис, Хесус (28 февраля 2005 г.). «Оппида и кельтское общество в западной Испании» . E-Keltoi: Журнал междисциплинарных кельтских исследований . 6 (1).
- ^ Перейти обратно: а б «Этнографическая карта доримской Иберии (около 200 г. до н. э.)» . Arqueotavira.com . Архивировано из оригинала 11 июня 2004 года . Проверено 12 января 2022 г.
- ^ «Испания – История» . Britannica.com .
- ^ Перейти обратно: а б Байкрофт, Клэр; Фернандес-Розадилья, Церера; Руис-Понте, Клара; Кинтела, Инес; Карраседо, Анхель; Доннелли, Питер; Майерс, Саймон (1 февраля 2019 г.). «Модели генетической дифференциации и следы исторических миграций на Пиренейском полуострове» . Природные коммуникации . 10 (1): 551. Бибкод : 2019NatCo..10..551B . дои : 10.1038/s41467-018-08272-w . ПМК 6358624 . ПМИД 30710075 .
- ^ Перейти обратно: а б Олальде, Иньиго; Маллик, Свапан; Паттерсон, Ник; Роланд, Надин; Вильяльба-Моуко, Ванесса; Сильва, Марина; Дулиас, Катарина; Эдвардс, Кейридвен Дж.; Гандини, Франческа; Пала, Мэри; Соарес, Педро; Феррандо-Берналь, Мануэль; Адамски, Николь; Брумандхошбахт, Насрин; Чероне, Оливия; Каллетон, Брендан Дж.; Фернандес, Даниэль; Лоусон, Энн Мари; Ма, Мэтью; Оппенгеймер, Йонас; Стюардсон, Кристин; Чжан, Чжао; Аренас, Хуан Мануэль Хименес; Мояно, Исидро Хорхе Торо; Саласар-Гарсия, Доминго К.; Кастанье, Пере; Сантос, Марта; Тремоледа, Иоахим; Лозано, Марина; и др. (15 марта 2019 г.). «Геномная история Пиренейского полуострова за последние 8000 лет» . Наука . 363 (6432): 1230–1234. Бибкод : 2019Sci...363.1230O . дои : 10.1126/science.aav4040 . ПМК 6436108 . ПМИД 30872528 .
- ^ «Вестготы в средневековой Португалии: современное состояние вопроса». Европа наследница вестготской Испании . Коллекция Дома Веласкеса. Книги.openedition.org. 23 января 2014. с. 326–339. ISBN 9788490960981 .
- ^ https://alpha.sib.uc.pt/?q=content/o-património-visigodo-da-l%C3%ADngua-portuguesa [ мертвая ссылка ]
- ^ Кирога, Хорхе Лопес (январь 2017 г.). «(PDF) IN TEMPORE SUEBORUM. Время свевов в Галлеции (411–585 гг. н.э.)» . Хорхе Лопес Кирога-Артемио М. Мартинес Техера (координатор): In Tempore Sueborum. Время свевов в Галлеции (411–585 гг. н.э.). Первое средневековое королевство Запада, Оренсе . Академия.edu . Проверено 21 января 2020 г.
- ^ Джеймс С. Амеланг. «Изгнание мориско: вопросов еще больше, чем ответов» (PDF) . Intransitduke.org . Автономный университет, Мадрид . Проверено 22 января 2022 г.
- ^ Йонссон 2007 , с. 195.
- ^ Адамс, Сьюзен М.; Босх, Елена; Балареск, Патрисия Л.; Баллеро, Стефан Ж.; Ли, Эндрю С.; Арройо, Эдуардо; Лопес-Парра, Ана М.; Алер, Мерседес; Грифо, Марина С. Гисберт; Брайон, Мэри; Карраседо, Анхель; Лавинья, Джон; Мартинес-Харрета, Бегонья; Кинтана-Мурси, Луис; Пикорнелл, Антония; Рамон, Мерси; Скорецкий, Карл; Бехар, Дорон М.; Калафель, Франческ; Джоблинг, Марк А. (12 декабря 2008 г.). «Генетическое наследие религиозного разнообразия и нетерпимости: отцовские линии христиан, евреев и мусульман на Пиренейском полуострове» . Американский журнал генетики человека . 83 (6): 725–736. дои : 10.1016/j.ajhg.2008.11.007 . ПМК 2668061 . ПМИД 19061982 .
- ^ Торрес, Габриэла (31 декабря 2008 г.). «В «чистом» испанском есть все» . Мир Би-би-си .
- ^ Виртуальный Сервантес: Арабское вторжение. Арабы и арабский элемент в испанском языке . Адамс С.М., Бош Э., Балареск П.Л., Баллеро С.Дж., Ли А.С., Арройо Е., Лопес-Парра А.М., Алер М., Грифо М.С., Брайон М., Карраседо А., Лавинья Дж., Мартинес-Харрета Б., Кинтана-Мурси Л., Пикорнелл А. , Рамон М., Скорецки К., Бехар Д.М., Калафель Ф., Джоблинг М.А. (декабрь 2008 г.). «Генетическое наследие религиозного разнообразия и нетерпимости: отцовские линии христиан, евреев и мусульман на Пиренейском полуострове» . Американский журнал генетики человека . 83 (6): 725–736. дои : 10.1016/j.ajhg.2008.11.007 . ПМК 2668061 . ПМИД 19061982 .
- ^ Перейти обратно: а б Байкрофт, Клэр; и др. (2019). «Модели генетической дифференциации и следы исторических миграций на Пиренейском полуострове» . Природные коммуникации . 10 (1): 551. Бибкод : 2019NatCo..10..551B . дои : 10.1038/s41467-018-08272-w . ПМК 6358624 . ПМИД 30710075 .
- ^ Перейти обратно: а б Олальде, Иньиго; и др. (2019). «Геномная история Пиренейского полуострова за последние 8000 лет» . Наука . 363 (6432): 1230–1234. Бибкод : 2019Sci...363.1230O . дои : 10.1126/science.aav4040 . ПМК 6436108 . ПМИД 30872528 .
- ^ Кавалли-Сфорца Л.Л. , Меноцци П., Пьяцца А (1993). История и география человеческих генов . Издательство Принстонского университета. п. 51. ИСБН 978-0-691-08750-4 . Проверено 22 июля 2009 г.
- ^ Милисаускас С (2002). Европейская предыстория: обзор . Биркгаузер. п. 58. ИСБН 978-0-306-46793-6 .
- ^ Нелис, Мэри; Эско, Тоне; Магия, Ридик; Зимприх, Фриц; Зимприч, Александр; Тончева, Драга; Карачанак, Сена; Пискацкова, Тереза; Балашкак, Иван; Пелтонен, Л; Яккула, Э; Ренстрем, К; Латроп, М; Хит, С; Галан, П; Шрайбер, С; Мейтингер, Т; Пфейфер, А; Вихманн, HE; Мелег, Б; Полгар, Н.; Тониоло, Д; Гаспарини, П; д'Адамо, П; Кловинс, Дж; Никитина-Заке, Л; Кучинскас, В; Каснаускене, Ж; Любинский, Дж; Дебняк, Т (2009). Флейшер, Роберт К. (ред.). «Генетическая структура европейцев: взгляд с северо-востока» . ПЛОС ОДИН . 4 (5): е5472. Бибкод : 2009PLoSO...4.5472N . дои : 10.1371/journal.pone.0005472 . ПМК 2675054 . ПМИД 19424496 .
- ^ Уэйд, Николас (13 августа 2008 г.). «Генетическая карта Европы» . Нью-Йорк Таймс . Проверено 17 октября 2009 г.
- ^ Новембре, Джон; Джонсон, Тоби; Брик, Катажина; Куталик, Золтан; Бойко, Адам Р.; Аутон, Адам; Индап, Амит; Кинг, Карен С.; Бергманн, Свен; Нельсон, Мэтью Р.; Стивенс, Мэтью; Бустаманте, Карлос Д. (2008). «Гены отражают географию Европы» . Природа . 456 (7218): 98–101. Бибкод : 2008Natur.456...98N . дои : 10.1038/nature07331 . ПМК 2735096 . ПМИД 18758442 .
- «Снова генетическая карта Европы» . Экспрессия генов . 31 августа 2008 г.
- ^ Байкрофт, Клэр; Фернандес-Розадилья, Церера; Руис-Понте, Клара; Кинтела, Инес; Карраседо, Анхель; Доннелли, Питер; Майерс, Саймон (1 февраля 2019 г.). «Модели генетической дифференциации и следы исторических миграций на Пиренейском полуострове» . Природные коммуникации . 10 (1): 551. Бибкод : 2019NatCo..10..551B . дои : 10.1038/s41467-018-08272-w . ПМК 6358624 . ПМИД 30710075 .
- ^ Олальде, Иньиго; Маллик, Свапан; Паттерсон, Ник; Роланд, Надин; Вильяльба-Моуко, Ванесса; Сильва, Марина; Дулиас, Катарина; Эдвардс, Кейридвен Дж.; Гандини, Франческа; Пала, Мария; Соарес, Педро (15 марта 2019 г.). «Геномная история Пиренейского полуострова за последние 8000 лет» . Наука . 363 (6432): 1230–1234. Бибкод : 2019Sci...363.1230O . дои : 10.1126/science.aav4040 . ISSN 0036-8075 . ПМК 6436108 . ПМИД 30872528 .
- ^ Дюпанлуп, И.; Берторель, Дж; Чихи, Л; Барбуджани, Дж. (2004). «Оценка влияния доисторической примеси на геном европейцев» . Молекулярная биология и эволюция . 21 (7): 1361–72. дои : 10.1093/molbev/msh135 . ПМИД 15044595 . S2CID 17665038 .
- ^ Перейти обратно: а б Босх, Елена; Калафель, Франческ; Комас, Дэвид; Оефнер, Питер Дж.; Андерхилл, Питер А.; Бертранпети, Жауме (2001). «Анализ вариаций Y-хромосомы человека с высоким разрешением показывает резкий разрыв и ограниченный поток генов между Северо-Западной Африкой и Пиренейским полуостровом» . Американский журнал генетики человека . 68 (4): 1019–29. дои : 10.1086/319521 . ПМЦ 1275654 . ПМИД 11254456 .
- ^ Комас, Дэвид; Калафель, Франческ; Бенчемси, киНуфисса; Хелал, Ахмед; Лефранк, Жерар; Стоункинг, Марк; Батцер, Марк А.; Бертранпети, Жауме; Саджантила, Антти (2000). «Полиморфизмы вставки Alu в северо-западной части Африки и на Пиренейском полуострове: свидетельства сильной генетической границы через Гибралтарский пролив». Генетика человека . 107 (4): 312–9. дои : 10.1007/s004390000370 . ПМИД 11129330 . S2CID 9618737 .
- ^ Перейти обратно: а б с Мурджани П., Паттерсон Н., Хиршхорн Дж.Н., Кейнан А., Хао Л., Ацмон Г. и др. (2011). «История потока африканских генов в южных европейцев, левантийцев и евреев» . ПЛОС Генет . 7 (4): e1001373. дои : 10.1371/journal.pgen.1001373 . ПМК 3080861 . ПМИД 21533020 .
- ^ Перейти обратно: а б с Волосы, Кристиан; Онофри, Валерио; Бризигелли, Франческа; Боски, Илария; Скарниччи, Франческа; Масулло, Мара; Ферри, Джанмарко; Тофанелли, Серджио; Тальябраччи, Адриано; Гусмао, Леонор; Аморим, Антонио; Гатто, Франческо; Кирин, Мирна; Мерлитти, Давиде; Брайон, Мэри; Вереа, Алехандро Бланко; Романо, Валентино; Кали, Франческо; Паскали, Винченцо (2009). «Мавры и сарацины в Европе: оценка средневекового мужского наследия Северной Африки в южной Европе» . Европейский журнал генетики человека . 17 (6): 848–52. дои : 10.1038/ejhg.2008.258 . ПМЦ 2947089 . ПМИД 19156170 .
- ^ Перейти обратно: а б Семино, Орнелла; Магри, Кьяра; Бенуцци, Джорджия; Лин, Алиса А.; Аль-Захери, Надя; Батталья, Винченца; Макчиони, Лилиана; Триантафиллидис, Костас; Шен, Пейдун; Оефнер, Питер Дж.; Животовский Лев А.; Кинг, Рой; Торрони, Антонио; Кавалли-Сфорца, Л. Лука; Андерхилл, Питер А.; Сантакьяра-Бенерекетти, А. Сильвана (2004). «Происхождение, распространение и дифференциация гаплогрупп E и J Y-хромосомы: выводы о неолитизации Европы и более поздних миграционных событиях в Средиземноморье» . Американский журнал генетики человека . 74 (5): 1023–34. дои : 10.1086/386295 . ПМК 1181965 . ПМИД 15069642 .
- ^ Перейти обратно: а б Жерар, Натали; Берриш, Сала; Ауизерат, Анни; Дитерлен, Флоран; Люкотт, Жерар (2006). «Североафриканское берберское и арабское влияние в Западном Средиземноморье, выявленное гаплотипами ДНК Y-хромосомы». Биология человека . 78 (3): 307–16. дои : 10.1353/hub.2006.0045 . ПМИД 17216803 . S2CID 13347549 .
- ^ Перейти обратно: а б с д и ж г час Адамс, Сьюзен М.; Босх, Елена; Балареск, Патрисия Л.; Баллеро, Стефан Ж.; Ли, Эндрю С.; Арройо, Эдуардо; Лопес-Парра, Ана М.; Алер, Мерседес; Грифо, Марина С. Гисберт; Брайон, Мэри; Карраседо, Анхель; Лавинья, Джон; Мартинес-Харрета, Бегонья; Кинтана-Мурси, Луис; Пикорнелл, Антония; Рамон, Мерси; Скорецкий, Карл; Бехар, Дорон М.; Калафель, Франческ; Джоблинг, Марк А. (2008). «Генетическое наследие религиозного разнообразия и нетерпимости: отцовские линии христиан, евреев и мусульман на Пиренейском полуострове» . Американский журнал генетики человека . 83 (6): 725–36. дои : 10.1016/j.ajhg.2008.11.007 . ПМК 2668061 . ПМИД 19061982 .
- Тина Хесман Сай (3 января 2009 г.). «Испанская инквизиция не смогла уничтожить мавританские еврейские гены» . Новости науки . Том. 175, нет. 1. Архивировано из оригинала 21 февраля 2009 г.
- ^ Перейти обратно: а б с д и ж Гонсалес, Ханна М.; Брем, Энтони; Перес, Джозеф А.; Мака-Мейер, Николь; Флауэрс, Чарльз; Кабрера, Винсент М. (апрель 2003 г.). «Сродство митохондриальной ДНК на атлантической окраине Европы». Американский журнал физической антропологии . 120 (4): 391–404. дои : 10.1002/ajpa.10168 . ПМИД 12627534 . S2CID 6430969 .
- ^ Перейти обратно: а б с Джакомо, Ф.; Лука, Ф.; Попа, Ло; Акар, Н.; Ананью, Н.; Банико Дж.; Брдичка, Р.; Барбуджани, Г.; и др. (2004). «Y-хромосомная гаплогруппа J как признак постнеолитической колонизации Европы». Генетика человека . 115 (5): 357–371. дои : 10.1007/s00439-004-1168-9 . ПМИД 15322918 . S2CID 18482536 .
- ^ Ботиге, LR; Хенн, Б.М.; Гравий, С; Мэйплс, Британская Колумбия; Жиньу, Чехия; Корона, Э; Ацмон, Г; Бернс, Э; Острер, Х; Флорес, К; Бертранпети, Дж; Комас, Д; Бустаманте, компакт-диск (2013). «Поток генов из Северной Африки способствует различному генетическому разнообразию человека в южной Европе» . Труды Национальной академии наук . 110 (29): 11791–11796. Бибкод : 2013PNAS..11011791B . дои : 10.1073/pnas.1306223110 . ПМК 3718088 . ПМИД 23733930 .
- ^ Оценка потока генов из Северной Африки в южную Европу. Архивировано 30 апреля 2015 г. на archive.today , Дэвид Комас, один из авторов исследования.
- ^ «Показатель в 20% встречается только на Канарских островах, для остальной части страны он колеблется от 10% до 12%», — объясняет Комас. Дэвид Комас, один из авторов исследования, « Мы, испанцы, — Европейцы с большим количеством североафриканских генов , Huffington Post, июнь 2013 г.
- ^ Цветы, Карлос; Вильяр, Хесус; Герра, Луиза; Эрнандес, Алексис; Басалдуа, Сантьяго; Корралес, Альмудена; Пино-Янес, Мария (30 марта 2011 г.). «Влияние Северной Африки и потенциальная предвзятость в исследованиях Ассоциации по контролю случаев заболевания среди испанского населения» . ПЛОС ОДИН . 6 (3): e18389. Бибкод : 2011PLoSO...618389P . дои : 10.1371/journal.pone.0018389 . ISSN 1932-6203 . ПМК 3068190 . ПМИД 21479138 .
- ^ Жерар, Натали; Берриш, Сала; Ауизерат, Анни; Дитерлен, Флоран; Люкотт, Жерар (2006). «Североафриканское берберское и арабское влияние в Западном Средиземноморье, выявленное гаплотипами ДНК Y-хромосомы» . Биология человека . 78 (3): 307–316. дои : 10.1353/hub.2006.0045 . ПМИД 17216803 . S2CID 13347549 .
- ^ Гонсалвес, Рита; Фрейтас, Ана; Бранко, Марта; Роза, Александра; Фернандес, Ана Т.; Животовский Лев А.; Андерхилл, Питер А.; Кивисилд, Тоомас; Брем, Антониу (2005). «Линии Y-хромосомы из Португалии, Мадейры и Асорских островов фиксируют элементы сефардского и берберского происхождения» . Анналы генетики человека . 69 (4): 443–454. дои : 10.1111/j.1529-8817.2005.00161.x . hdl : 10400.13/3018 . ПМИД 15996172 . S2CID 3229760 .
- ^ Перейра Л., Кунья К., Алвес К., Аморим А. (апрель 2005 г.). «Африканское женское наследие в Иберии: переоценка распределения линий мтДНК в настоящее время». Биология человека . 77 (2): 213–29. дои : 10.1353/hub.2005.0041 . hdl : 10216/109268 . ПМИД 16201138 . S2CID 20901589 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Гонсалес А.М., Брем А., Перес Х.А., Мака-Мейер Н., Флауэрс С., Кабрера В.М. (апрель 2003 г.). «Сродство митохондриальной ДНК на атлантической окраине Европы». Американский журнал физической антропологии . Том. 120, нет. 4. стр. 391–404. дои : 10.1002/ajpa.10168 . ПМИД 12627534 .
{{cite news}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Дюпанлуп И., Берторель Г., Чихи Л., Барбуджани Г. (июль 2004 г.). «Оценка влияния доисторической примеси на геном европейцев». Молекулярная биология и эволюция . Том. 21, нет. 7. стр. 1361–72. дои : 10.1093/molbev/msh135 . ПМИД 15044595 .
{{cite news}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Бош Э., Калафель Ф., Комас Д., Офнер П.Дж., Андерхилл П.А., Бертранпетит Дж. (апрель 2001 г.). «Анализ вариаций Y-хромосомы человека с высоким разрешением показывает резкий разрыв и ограниченный поток генов между северо-западной Африкой и Пиренейским полуостровом». Американский журнал генетики человека . Том. 68, нет. 4. С. 1019–29. дои : 10.1086/319521 . ПМЦ 1275654 . ПМИД 11254456 .
{{cite news}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Комас, Дэвид; Калафель, Франческ; Бенчемси, Нуфисса; Хелал, Ахмед; Лефранк, Жерар; Стоункинг, Марк; Батцер, Марк А.; Бертранпети, Жауме; Саджантила, Антти (2000). «Полиморфизмы вставок Alu в Северо-Западной Африке и на Пиренейском полуострове: свидетельства сильной генетической границы через Гибралтарский пролив» . Генетика человека . 107 (4): 312–319. дои : 10.1007/s004390000370 . ПМИД 11129330 . S2CID 9618737 .
- ^ Спинола, Х.; Миддлтон, Д.; Брем, А. (2005). «Гены HLA в Португалии, полученные в результате типирования на основе последовательностей: на перекрестке между Европой и Африкой» . Тканевые антигены . 66 (1): 26–36. дои : 10.1111/j.1399-0039.2005.00430.x . ПМИД 15982254 .
- ^ «Генетическое наследие религиозного разнообразия и нетерпимости: отцовские линии христиан, евреев и мусульман на Пиренейском полуострове» .
- ^ Эрнандес К.Л., Пита Г., Кавадас Б., Лопес С., Санчес-Мартинес Л.Дж., Дюгухон Х.М. и др. (2020). «Геномное разнообразие человека там, где Средиземное море соединяется с Атлантикой» . Мол Биол Эвол . 37 (4): 1041–1055. дои : 10.1093/molbev/msz288 . ПМК 7086172 . ПМИД 31816048 .
- ^ Люкотт, Жерар; Жерар, Натали; Мерсье, Жеральдин (5 апреля 2011 г.). «Североафриканские гены в Иберии, изученные с помощью гаплотипа 5 ДНК Y-хромосомы» . Биология человека . 73 (5).
- ^ Кручиани, Ф.; Ла Фратта, Р.; Тромбетта, Б.; Сантоламазца, П.; Селитто, Д.; Коломб, Э.Б.; Дюгужон, Ж.-М.; Кривелларо, Ф.; Бенинкаса, Т. (2007). «Отслеживание прошлых человеческих перемещений мужчин в Северной / Восточной Африке и Западной Евразии: новые подсказки по гаплогруппам Y-хромосомы E-M78 и J-M12» . Молекулярная биология и эволюция . 24 (6): 1300–1311. дои : 10.1093/molbev/msm049 . ПМИД 17351267 .
- ^ Кручиани, Ф.; Ла Фратта, Р.; Тромбетта, Б.; Сантоламазца, П.; Селитто, Д.; Коломб, Э.Б.; Дюгужон, Ж.-М.; Кривелларо, Ф.; Бенинкаса, Т.; Пасконе, Р.; Мораль, П.; Уотсон, Э.; Мелег, Б.; Барбуджани, Г.; Фюзелли, С.; Вона, Г.; Заградисник, Б.; Ассум, Г.; Брдичка, Р.; Козлов А.И.; Ефремов, Г.Д.; Коппа, А.; Новеллетто, А.; Скоццари, Р. (2007). «Отслеживание прошлых человеческих перемещений мужчин в Северной / Восточной Африке и Западной Евразии: новые подсказки по гаплогруппам Y-хромосомы E-M78 и J-M12» . Молекулярная биология и эволюция . 24 (6): 1300–11. дои : 10.1093/molbev/msm049 . ПМИД 17351267 .
- ^ Бош, Э; Калафель, Ф; Комас, Д; Оефнер, П.Дж.; Андерхилл, Пенсильвания; Бертранпети, Дж (апрель 2001 г.). «Анализ вариаций Y-хромосомы человека с высоким разрешением показывает резкий разрыв и ограниченный поток генов между северо-западной Африкой и Пиренейским полуостровом» . Являюсь. Дж. Хум. Жене . 68 (4): 1019–29. дои : 10.1086/319521 . ПМЦ 1275654 . ПМИД 11254456 .
- ^ Плаза, С.; Калафель, Ф.; Хелал, А.; Бузерна, Н.; Лефранк, Г.; Бертранпети, Ж.; Комас, Д. (2003). «Присоединение к Геркулесовым столпам: последовательности мтДНК показывают разнонаправленный поток генов в Западном Средиземноморье». Анналы генетики человека . 67 (Часть 4): 312–28. дои : 10.1046/j.1469-1809.2003.00039.x . ПМИД 12914566 . S2CID 11201992 .
- ^ Перейти обратно: а б с д Перейра, Луиза; Кунья, Карла; Алвес, Синтия; Аморим, Антонио (2005). «Африканское женское наследие в Иберии: переоценка распределения линий мтДНК в настоящее время». Биология человека . 77 (2): 213–29. дои : 10.1353/hub.2005.0041 . hdl : 10216/109268 . ПМИД 16201138 . S2CID 20901589 .
- ^ Сересо, М.; Ахилли, А.; Оливьери, А.; Перего, Украина; Гомес-Карбалла, А.; Бризигелли, Ф.; Лансиони, Х.; Вудворд, СР; Лопес-Сото, М.; Карраседо, А.; Капелли, К.; Торрони, А.; Салас, А. (27 марта 2012 г.). «Реконструкция древних связей митохондриальной ДНК между Африкой и Европой» . Геномные исследования . 22 (5): 821–826. дои : 10.1101/гр.134452.111 . ПМЦ 3337428 . ПМИД 22454235 .
- ^ Перейти обратно: а б Брем А., Перейра Л., Кивисилд Т., Аморим А. (декабрь 2003 г.). «Митохондриальные портреты архипелагов Мадейра и Асорские острова свидетельствуют о разных генетических пулах их поселенцев». Генетика человека . 114 (1): 77–86. дои : 10.1007/s00439-003-1024-3 . hdl : 10400.13/3046 . ПМИД 14513360 . S2CID 8870699 .
- ^ Перейти обратно: а б Эрнандес, Кандела Л; Реалес, Гильермо; Дюгужон, Жан-Мишель; Новеллетто, Андреа; Родригес, Хуан; Куэста, Педро; Кальдерон, Росарио (2014). «Человеческое материнское наследие в Андалусии (Испания): его состав демонстрирует высокую внутреннюю сложность и отличительное влияние гаплогрупп U6 и L мтДНК в западной и восточной части региона» . БМК Генетика . 15 (1): 11. дои : 10.1186/1471-2156-15-11 . ПМЦ 3905667 . ПМИД 24460736 .
- ^ Гордон, Мэтью; Хейн, Кэтрин А. (2017). Наложницы и куртизанки: женщины и рабство в истории ислама . Издательство Оксфордского университета. ISBN 9780190622183 .
- ^ «Отслеживание прошлых перемещений мужчин-мужчин в Северной / Восточной Африке и Западной Евразии» . Academic.oup.com . Проверено 21 января 2020 г.
- ^ https://reich.hms.harvard.edu/sites/reich.hms.harvard.edu/files/inline-files/2019_Olalde_Science_IberiaTransect_2.pdf [ пустой URL PDF ]
- ^ Альварес, Луис; Сантос, Кристина; Рамос, Аманда; Пратдесаба, Розер; Франкалаччи, Паоло; Алуха, Мария Пилар (1 февраля 2010 г.). «Образцы митохондриальной ДНК на северном иберийском плато: динамика населения и субструктура провинции Самора». Американский журнал физической антропологии . 142 (4): 531–539. дои : 10.1002/ajpa.21252 . ПМИД 20127843 .
- ^ Эрнандес, Кандела Л.; Соарес, Педро; Дюгужон, Жан М.; Новеллетто, Андреа; Родригес, Хуан Н.; Рито, Тереза; Оливейра, Мариса; Мелхауи, Мохаммед; Баали, Абдельлатиф; Перейра, Луиза; Кальдерон, Росарио; Ахилли, Алессандро (28 октября 2015 г.). «Ранние голоценовые и исторические африканские признаки мтДНК на Пиренейском полуострове: Андалузский регион как парадигма» . ПЛОС ОДИН . 10 (10): e0139784. Бибкод : 2015PLoSO..1039784H . дои : 10.1371/journal.pone.0139784 . ПМЦ 4624789 . ПМИД 26509580 .
- ^ Перейти обратно: а б с Майрес, Натали М; Роотси, Сиири; Лин, Алиса А; Ярве, Мари; Кинг, Рой Дж; Кутуев, Ильдус; Кабрера, Висенте М; Хуснутдинова, Эльза К; Пшеничнов Андрей; Юнусбаев, Баязит; Балановский Олег; Балановская, Елена; Рудан, Павао; Бальдович, Мариан; Эррера, Рене Дж; Кьярони, Жак; Ди Кристофаро, Джули; Виллемс, Ричард; Кивисилд, Тоомас; Андерхилл, Питер А. (25 августа 2010 г.). «Эффект основателя основной гаплогруппы Y-хромосомы R1b эпохи голоцена в Центральной и Западной Европе» . Европейский журнал генетики человека . 19 (1): 95–101. дои : 10.1038/ejhg.2010.146 . ПМК 3039512 . ПМИД 20736979 .
- ^ Маркес, София Л.; Гойос, Ана; Роча, Ана М.; Прата, Мария Жуан; Аморим, Антониу; Гужман, Леонор; Алвес, Синтия; Альварес, Луис (март 2015 г.). «Генетическое разнообразие португальской митохондриальной ДНК - обновление и филогенетический пересмотр». Международная судебно-медицинская экспертиза: Генетика . 15 :27–32. дои : 10.1016/j.fsigen.2014.10.004 . ПМИД 25457629 .
- ^ Вальверде, Лаура; Ильескас, Мария Хосе; Вильяскуза, Патрисия; Готор, Ампаро М.; Гарсиа, Айнара; Кардозо, Серджио; Альгорта, Хайме; Катарино, Сусана; Руо, Карен; Ферек, Клод; Хардиман, Орла; Заррабейтия, Майте; Хименес, Сусана; Пиньейру, Мария Фатима; Харрета, Бегонья М.; Олофссон, Джилл; Морлинг, Нильс; де Панкорбо, Мариан М. (март 2016 г.). «Новые ключи к разгадке эволюционной истории основной европейской отцовской линии M269: вскрытие Y-SNP S116 в Атлантической Европе и Иберии» . Европейский журнал генетики человека . 24 (3): 437–441. дои : 10.1038/ejhg.2015.114 . ПМЦ 4755366 . ПМИД 26081640 .
- ^ Олальде, Иньиго; Брейс, Селина; Аллентофт, Мортен Э.; Армит, Ян; Кристиансен, Кристиан; Бут, Томас; Роланд, Надин; Маллик, Свапан; Сечени-Надь, Анна; Миттник, Алисса; Альтена, Эвелин; Липсон, Марк; Лазаридис, Джозеф; Харпер, Томас К.; Паттерсон, Ник; Брумандхошбахт, Насрин; Дикманн, Джоан; Фалтыскова, Зузана; Фернандес, Даниэль; Ферри, Мэтью; Харни, Эдаоин; де Книфф, Питер; Мишель, Меган; Оппенгеймер, Йонас; Стюардсон, Кристин; Барклай, Алистер; Альт, Курт Вернер; Лизау, Корина; Риверс, Патрисия; и др. (март 2018 г.). «Феномен стакана и геномная трансформация северо-западной Европы» . Природа . 555 (7695): 190–196. Бибкод : 2018Natur.555..190O . дои : 10.1038/nature25738 . ПМЦ 5973796 . ПМИД 29466337 .
- ^ Аллентофт, Мортен Э.; Сикора, Мартин; Шегрен, Карл-Йоран; Расмуссен, Саймон; Расмуссен, Мортен; Стендерап, Йеспер; Дамгаард, Питер Б.; Шредер, Ханнес; Альстрем, Торбьёрн; Победитель, Лассе; Маласпинас, Анна-Сапфо; Маргарян, Ашот; Хайэм, Том; Чивалл, Дэвид; Линнер, Нильс; Харвиг, Средняя школа; барон Юстина; Каса, Филипп Делла; Домбровский, Павел; Даффи, Пол Р.; Эбель, Александр В.; Епимахов Андрей; Фрей, Карин; Фурманек, Мирослав; Гралак, Томас; Громов, Андрей; Гронкевич, Станислав; Групе, Гизела; Хайду, Томас; Яриш, Радослав; Хартанович, Валери; Хохлов, Александр; Поцелуй, Виктория; Коларж, Ян; Крийска, Ивар; Ласак, Ирена; Лонги, Кристина; МакГлинн, Джордж; Меркявичюс, Альгимантас; Мерките, Инга; Мецпалу, Мейт; Мкртчян, Рузан; Моисеев Вячеслав; Пайя, Ласло; Палфи, Дьердь; Покутта, Далия; Поторопился, Люк; Прайс, Т. Дуглас; Сааг, Лехти; Саблин, Михаил; Шишлина, Наталья; Смрчка, Вацлав; Соенов Василий Иванович; Севереньи, Вайк; Тот, Густав; Трифанова Сынару В.; Варул, Леви; Вице, Магдольна; Член епископальной церкви Левон; Житенев Владислав; Орландо, Людовик; Зихеритц-Понтен, Томас; Брунак, Сорен; Нильсен, Расмус; Кристиансен, Кристиан; Виллерслев, Эске (июнь 2015 г.). «Популяционная геномика Евразии бронзового века» . Природа . 522 (7555): 167–172. Бибкод : 2015Natur.522..167A . дои : 10.1038/nature14507 . ПМИД 26062507 . S2CID 4399103 .
- ^ Питер де Баррос, Дамгаард (9 мая 2018 г.). «137 древних геномов человека со всех евразийских степей». Природа . 557 (7705): 369–374. Бибкод : 2018Natur.557..369D . дои : 10.1038/s41586-018-0094-2 . hdl : 1887/3202709 . ПМИД 29743675 . S2CID 13670282 .
- ^ «Эупедия» .
- ^ Баррал-Арка Р., Пишедда С., Гомес-Карбалья А., Пасториса А., Москера-Мигель А., Лопес-Сото М. и др. (2016). «Метаанализ вариаций митохондриальной ДНК на Пиренейском полуострове» . ПЛОС ОДИН . 11 (7): e0159735. Бибкод : 2016PLoSO..1159735B . дои : 10.1371/journal.pone.0159735 . ПМЦ 4956223 . ПМИД 27441366 .
- ^ Россер и др. (2000)
- ^ Лоогвяли Э.Л., Роосталу У., Малярчук Б.А. и др. (ноябрь 2004 г.). «Разъединяющее единообразие: пестрая кладистическая картина гаплогруппы H мтДНК в Евразии» . Молекулярная биология и эволюция . 21 (11): 2012–21. дои : 10.1093/molbev/msh209 . ПМИД 15254257 .
- ^ Оттони, Клаудио; Примативо, Джузеппина; Хушиар Кашани, Бахарак; Ахилли, Алессандро; Мартинес-Лабарга, Кристина; Бионди, Джанфранко; Торрони, Антонио; Рикардс, Ольга; Кайзер, Манфред (21 октября 2010 г.). «Митохондриальная гаплогруппа H1 в Северной Африке: прибытие из Иберии в раннем голоцене» . ПЛОС ОДИН . 5 (10): е13378. Бибкод : 2010PLoSO...513378O . дои : 10.1371/journal.pone.0013378 . ПМЦ 2958834 . ПМИД 20975840 .
- ^ Хендриксон С.Л., Хатчесон Х.Б., Руис-Песини Э. и др. (ноябрь 2008 г.). «Гаплогруппы митохондриальной ДНК влияют на прогрессирование СПИДа» . СПИД . 22 (18): 2429–39. дои : 10.1097/QAD.0b013e32831940bb . ПМЦ 2699618 . ПМИД 19005266 .
- ^ Боше, М; МакЭвой, Б; Пирсон, Л.Н.; Куиллен, Э.Э.; Саркисян Т; Оганесян, К; Дека, Р; Брэдли, генеральный директор; Шрайвер, доктор медицины (2007). «Измерение стратификации населения Европы с помощью данных генотипа микрочипов» . Американский журнал генетики человека . 80 (5): 948–56. дои : 10.1086/513477 . ПМЦ 1852743 . ПМИД 17436249 .
- ^ Флорес, Карлос; Мака-Мейер, Николь; Гонсалес, Ана М.; Оефнер, Питер Дж.; Шен, Пейдун; Перес, Хосе А.; Рохас, Антонио; Ларруга, Хосе М.; Андерхилл, Питер А. (октябрь 2004 г.). «Редуцированная генетическая структура Пиренейского полуострова, выявленная с помощью анализа Y-хромосомы: последствия для демографии населения» . Европейский журнал генетики человека . 12 (10): 855–863. дои : 10.1038/sj.ejhg.5201225 . ПМИД 15280900 .
- ^ Гаплогруппа C* (C-M130) была обнаружена среди мужчин с фамилией Ллах, происходящих из Гаррочи , Каталония , Испания . Он не был обнаружен среди мужчин с такой же фамилией из других регионов или мужчин с другими фамилиями каталонского происхождения ( Cognoms Catalans, nd, Resultat ; доступ 15 сентября 2015 г.). Проект Cognoms Catalans, который исследует «генетические фамилии» в Каталонии, Валенсии и на Балеарских островах, базируется в Университете Помпеу Фабра в Барселоне.
- ^ Гаплогруппа C - Классическая таблица Y-ДНК (21 января 2017 г.).
- ^ Маркес, София Л; Гойос, Ана; Роча, Ана М; Прата, Мария Дж; Аморим, Антониу; Гужман, Леонор; Алвес, Синтия; Альварес, Луис (март 2015 г.). «Генетическое разнообразие португальской митохондриальной ДНК - обновление и филогенетический пересмотр». ФСИ Генетика . 15 :27–32. дои : 10.1016/j.fsigen.2014.10.004 . ПМИД 25457629 .
Цитируемые работы
[ редактировать ]- Йонссон, Мар (2007). «Изгнание мориско из Испании в 1609–1614 годах: разрушение исламской периферии». Журнал глобальной истории . 2 (2): 195–212. дои : 10.1017/S1740022807002252 . S2CID 154793596 .
- Россер, З; Зержал, Т; Херлс, М; Адохаан, М; Алавантик, Д; Аморим, А; Амос, В; Арментерос, М; и др. (2000), «Разнообразие Y-хромосомы в Европе является клиническим и зависит в первую очередь от географии, а не от языка» , Американский журнал генетики человека , 67 (6): 1526–1543, doi : 10.1086/316890 , PMC 1287948 , PMID 11078479 , заархивировано из оригинала 6 мая 2008 г.
| Подтемы | |
|---|---|
| Генетическая история по регионам | |
| Популяционная генетика по группе |
|