Генетические исследования сингальцев
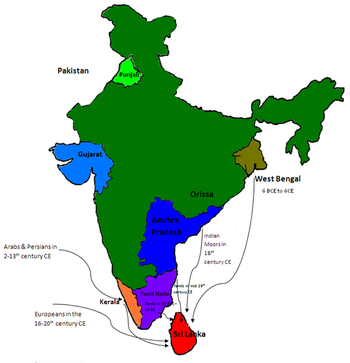
Генетические исследования сингальцев являются частью популяционной генетики, изучающей происхождение сингальского населения.
Все исследования сходятся во мнении, что существует значительная связь между сингальцами и бенгальцами и тамилами Южной Индии, а также что существует значительная генетическая связь между тамилами Шри-Ланки и сингальцами. Это также подтверждается исследованием генетического расстояния, которое показало низкие различия в генетическом расстоянии между сингальцами и бенгальцами , тамилами и кералитами- добровольцами. [1]
Отношения с бенгальцами
[ редактировать ]
Анализ Alu полиморфизма , проведенный Мастана С. (2007) с использованием сингальского, тамильского, бенгальского, гуджаратского ( Патель ) и пенджабского языков в качестве родительских популяций, выявил следующие пропорции генетического вклада: [2]
| Статистический метод | Бенгальский | тамильский | Северо-Западный |
|---|---|---|---|
| Оценка очков | 57.49% | 42.5% | - |
| Метод максимального правдоподобия | 88.07% | - | - |
| Использование тамильского, бенгальского и северо-западного населения в качестве родительского населения | 50-66% | 11-30% | 20-23% |
| Родительское население | Бенгальский | тамильский | Гуджарати | панджаби |
|---|---|---|---|---|
| Использование тамильского и бенгальского населения в качестве родительского населения | 70.03% | 29.97% | - | |
| Использование тамильского, бенгальского и гуджаратского языков в качестве родительского населения | 71.82% | 16.38% | 11.82% | |
| Использование бенгали, гуджарати и пенджаби в качестве родительского населения | 82.09% | - | 15.39% | 2.52% |
Анализ STR Х-хромосомы, проведенный Perera et al. (2021) обнаружили, что сингальцы (а также тамилы Шри-Ланки и мусульмане Шри-Ланки ) более тесно связаны с бенгальцами, чем с индийскими тамилами Шри-Ланки . [3]
Анализ генетических расстояний, проведенный Кирком (1976), показал, что сингальцы ближе к бенгальцам, чем к популяциям Гуджарата или Пенджаба. [4]
Частота аллеля D1S80 (популярный аллель для генетической дактилоскопии) также одинакова у сингальцев и бенгальцев, что позволяет предположить, что эти две группы тесно связаны. [5]
Сингальцы также имеют такие же частоты аллеля MTHFR 677T (13%), что и жители Западной Бенгалии (17%). [6] [7]
Отношения с индийскими тамилами
[ редактировать ]
Исследование генетической примеси, проведенное Кшатрией (1995), показало, что в сингальцах больше вклада индийских тамилов (69,86% +/- 0,61) по сравнению с бенгальцами (25,41% +/- 0,51). [8]
Генетический дистанционный анализ, проведенный Roychoudhury AK et al. (1985) предположили, что сингальцы более тесно связаны с населением Южной и Вест-Индии, чем бенгальцы. [9]
Анализ генетических расстояний, проведенный Кирком (1976), показал, что сингальцы ближе к тамилам и кералитам Южной Индии, чем к населению Гуджарата или Пенджаба. [4]

Исследование, проведенное Сингхом и др. в 2023 году с использованием маркеров с более высоким разрешением, чем предыдущие исследования, показало, что поток генов из Южной Индии к сингальцам был выше, чем из Северной Индии, при этом сингальцы имеют самую высокую идентичность по происхождению с тамилами , особенно с пирамалайскими калларами , по сравнению с другим изучаемым популяциям Индии. Исследование также выявило повышенное общение с индийскими маратхами , которого не было у тамильского и других южноиндийских популяций, что согласуется со следами принадлежности Северной Индии к сингальскому населению, что подтверждает лингвистическую гипотезу Лазаря Гейгера , Ральфа Лилли Тернера и Джорджа Ван. Дрим . Общий размер выборки сингальцев составил 9 человек. [10]
Отношения с индейцами Северо-Западной Индии
[ редактировать ]Анализ Alu полиморфизма , проведенный Мастана С. (2007), выявил незначительный вклад Северо-Западной Индии (20-23%). [11]
Анализ STR X-хромосомы, проведенный Перерой и др. (2011), показал, что сингальцы, тамилы Шри-Ланки, мавры и индийские тамилы Шри-Ланки имеют сходство с бхилами ( группой коренных народов) Северо-Западной Индии. [12]
Отношения с другими основными этническими группами Шри-Ланки
[ редактировать ]Исследование, посвященное генетическим вариациям гена FUT2 в сингальском и тамильском населении Шри-Ланки, обнаружило схожие генетические корни для обеих этнических групп с небольшим генетическим потоком от других соседних азиатских групп населения. [13] Исследования также не обнаружили существенных различий в отношении группы крови , генетических маркеров крови (Саха, 1988) и однонуклеотидного полиморфизма между сингальцами и другими этническими группами Шри-Ланки. [14] [15] [16] Другое исследование также не выявило «никаких существенных генетических различий среди основных этнических групп Шри-Ланки». [17] Это также подтверждается исследованием, которое обнаружило очень схожие частоты аллелей MTHFR 677T , F2 20210A и F5 1691A у индийских тамилов, сингальцев, тамилов Шри-Ланки и мавров Шри-Ланки. [7]
Отношения с другими выходцами из Южной и Западной Азии
[ редактировать ]Исследование 1985 года, проведенное Ройчоудхури А.К. и Ней М., указывающее на значения генетической дистанции, показало, что сингальцы, наряду с популяциями четырех индийских субконтинентов из Пенджаба , Гуджарата , Андхра-Прадеша и Бангладеш , были ближе к афганцам и иранцам , чем к соседнему Востоку. Группы Юго-Восточной Азии, представленные бутанцами , малайцами , батаками на севере Суматры и китайцами . [9]
Отношения с выходцами из Восточной и Юго-Восточной Азии
[ редактировать ]Генетические маркеры иммуноглобулина среди сингальцев показывают высокую частоту afb1b3, который происходит из провинций Юньнань и Гуанси на юге Китая. [18] Он также часто встречается среди одиаса, некоторых непальских и северо-восточных индийских, южных ханьских китайцев, юго-восточной Азии и некоторых австронезийских популяций островов Тихого океана . [18] С более низкой частотой ab3st также встречается у сингальцев и обычно встречается с более высокой частотой среди северного ханьского китайца, тибетского, монгольского, корейского и японского населения. [18] Аллель трансферрина TF*Dchi, распространенный среди населения Восточной Азии и коренных американцев, также встречается у сингальцев. [9] HumDN1*4 и HumDN1*5 являются преобладающими генами ДНКазы I среди сингальцев, а также являются преобладающими генами среди этнических групп южного Китая и народа таманг в Непале. [19] Исследование 1988 года, проведенное Н. Саха, показало, что высокие частоты GC*1F и низкие GC*1S у сингальцев сопоставимы с таковыми у китайцев, японцев, корейцев, тайцев, малайцев, вьетнамцев, лаосцев и тибетцев. [20] Гемоглобин Е, вариант нормального гемоглобина, возникший и широко распространенный среди населения Юго-Восточной Азии , также распространен среди сингальцев и может достигать 40% в Шри-Ланке. [21]
Отцовская линия
[ редактировать ]Y-ДНК сингальского языка
[ редактировать ]Наиболее распространенными гаплогруппами ДНК Y-хромосомы, обнаруженными у сингальцев, являются гаплогруппа R2 , гаплогруппа L , гаплогруппа R1a и F в указанном порядке. [22]
| Население | н | С | И | Ф | Г | ЧАС | я | Дж | К | л | Н | ТО | П | вопрос | Р | Р1 | Р1а | Р1б | Р2 | Т | Другие | Ссылка |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| сингальский | 39 | 0 | 0 | 10.3% | 0 | 10.3% | 0 | 10.3% | 0 | 18% | 0 | 0 | 0 | 0 | 0 | 0 | 12.8% | 0 | 38.5% | 0 | Кивисилд2003 [22] |
Материнская линия
[ редактировать ]МтДНК сингальского языка
[ редактировать ]Ранвира и др. (2014) обнаружили, что наиболее распространенной гаплогруппой мтДНК у сингальцев являются гаплогруппа M , гаплогруппа U (U7a), гаплогруппа R (R30b) и гаплогруппа G (G3a1'2). [23] [24]
Гаплогруппа M представляет собой расселение современных людей около 60 000 лет назад вдоль южного побережья Азии по южному прибрежному маршруту через Аравию и Индию, чтобы вскоре достичь Австралии. [25]
Гаплогруппа U7 считается специфичной для Западной Евразии гаплогруппой мтДНК, которая, как полагают, возникла в районе Черного моря примерно 30 000 лет назад. В Южной Азии U7 встречается примерно у 12% в Гуджарате, тогда как по всей Индии его частота остается около 2% и 5% в Пакистане . У веддийского народа Шри-Ланки он достигает самой высокой частоты - 13,33% (субклад U7a). Предполагается, что крупномасштабная иммиграция привела эти митохондриальные гаплогруппы в Индию. [26]
Чаубей утверждает, что «значительное количество материнских линий Шри-Ланки является общим с Индией, точнее, с южной частью Индии ». [27]
Ссылки
[ редактировать ]- ^ Кирк, Р.Л. (июль 1976 г.). «Легенда о принце Виджае — исследование сингальского происхождения». Американский журнал физической антропологии . 45 (1): 91–99. дои : 10.1002/ajpa.1330450112 .
- ^ Мастана С (2007). «Молекулярная антропология: популяционные и судебно-генетические приложения» (PDF) . Специальный антрополог . 3 : 373–383.
- ^ Перера, Н., Галхена, Г. и Ранавака, Г., 2021. Генетические полиморфизмы на основе X-хромосомных STR и демографическая история этнических групп Шри-Ланки и их отношения с населением мира. Научные отчеты, 11(1), стр.1-12.
- ^ Перейти обратно: а б Кирк Р.Л. (июль 1976 г.). «Легенда о принце Виджае – исследование сингальского происхождения». Американский журнал физической антропологии . 45 (1): 91–99. дои : 10.1002/ajpa.1330450112 .
- ^ Суриндер Сингх Папиха (1999). Геномное разнообразие: применение в популяционной генетике человека. Лондон: Спрингер. 7.
- ^ Мукхопадьяй К., Дутта С., Дас Бхомик А. (январь 2007 г.). «Полиморфизмы гена MTHFR проанализированы в популяции из Калькутты, Западная Бенгалия» . Индийский журнал генетики человека . 13 (1): 38. doi : 10.4103/0971-6866.32035 (неактивно с 17 мая 2024 г.). ПМК 3168154 . ПМИД 21957342 .
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на май 2024 г. ( ссылка ) - ^ Перейти обратно: а б Диссанаяке В.Х., Вирасекера Л.Ю., Гаммулла К.Г., Джаясекара Р.В. (октябрь 2009 г.). «Распространенность генетических тромбофильных полиморфизмов среди населения Шри-Ланки - значение для разработки ассоциативных исследований и услуг клинического генетического тестирования». Экспериментальная и молекулярная патология . 87 (2): 159–62. дои : 10.1016/j.yexmp.2009.07.002 . ПМИД 19591822 .
- ^ Кшатрия ГК (декабрь 1995 г.). «Генетическое сходство населения Шри-Ланки». Биология человека . 67 (6). Американская ассоциация антропологической генетики: 843–66. ПМИД 8543296 .
- ^ Перейти обратно: а б с Ройчоудхури, Арун К.; Ней, Масатоши (2 сентября 2008 г.). «Генетические связи между индейцами и соседним населением» . Наследственность человека . 35 (4): 201–206. дои : 10.1159/000153545 . ISSN 0001-5652 . ПМИД 4029959 .
- ^ Праджджвал Пратап Сингх, Сачин Кумар, Нагарджуна Пасупулети, Нирадж Рай, Гьянешвер Чаубей, Р. Ранасингхе, «Реконструкция истории населения сингальцев, основной этнической группы в Шри-Ланке», iScience, 31 августа 2023 г., DOI: https:/ //doi.org/10.1016/j.isci.2023.107797 .
- ^ Мастана С (2007). «Молекулярная антропология: популяционные и судебно-генетические приложения» (PDF) . Специальный антрополог . 3 : 373–383.
- ^ Перера, Н., Галхена, Г. и Ранавака, Г., 2021. Генетические полиморфизмы на основе X-хромосомных STR и демографическая история этнических групп Шри-Ланки и их отношения с населением мира. Научные отчеты, 11(1), стр.1-12.
- ^ Соэдзима М., Кода Ю. (декабрь 2005 г.). «Денатурация генотипирования на основе высокоэффективной жидкостной хроматографии и генетических вариаций FUT2 в Шри-Ланке». Переливание . 45 (12): 1934–9. дои : 10.1111/j.1537-2995.2005.00651.x . ПМИД 16371047 . S2CID 10401001 .
- ^ Саха Н (1988). «Генетические маркеры крови в населении Шри-Ланки - переоценка легенды о принце Виджае». Американский журнал физической антропологии . 76 (2): 217–25. дои : 10.1002/ajpa.1330760210 . ПМИД 3166342 .
- ^ Робертс Д.Ф., Крин К.К., Абеяратне К.П. (1972). «Группы крови сингальцев». Мужчина . 7 (1): 122–127. дои : 10.2307/2799860 . JSTOR 2799860 .
- ^ Диссанаяке В.Х., Джайлз В., Джаясекара Р.В., Сеневиратне Х.Р., Калшекер Н., Бротон Пипкин Ф., Морган Л. (апрель 2009 г.). «Исследование трех генов-кандидатов преэклампсии у сингальской популяции из Шри-Ланки» . Журнал исследований в области акушерства и гинекологии . 35 (2): 234–42. дои : 10.1111/j.1447-0756.2008.00926.x . ПМИД 19708171 . S2CID 24958292 .
- ^ Иллеперума Р.Дж., Мохотти С.Н., Де Сильва Т.М., Фернандопулле Н.Д., Ратнасурия В.Д. (июнь 2009 г.). «Генетический профиль 11 аутосомных локусов STR среди четырех основных этнических групп Шри-Ланки». Международная судебно-медицинская экспертиза. Генетика . 3 (3): е105-6. дои : 10.1016/j.fsigen.2008.10.002 . ПМИД 19414153 .
- ^ Перейти обратно: а б с Мацумото Х (2009). «Происхождение японской расы на основе генетических маркеров иммуноглобулина G» . Труды Японской академии. Серия Б, Физические и биологические науки . 85 (2): 69–82. Бибкод : 2009PJAB...85...69M . дои : 10.2183/pjab.85.69 . ПМЦ 3524296 . ПМИД 19212099 .
- ^ Фуджихара Дж., Ясуда Т., Иида Р., Уэки М., Сано Р., Коминато Ю. и др. (июль 2015 г.). «Глобальный анализ генетических вариаций вариативного числа полиморфизмов тандемных повторов длиной 56 п.н. в гене дезоксирибонуклеазы I человека». Юридическая медицина . 17 (4): 283–6. doi : 10.1016/j.legalmed.2015.01.005 . ПМИД 25771153 .
- ^ Малхотра Р. (1992). Антропология развития: памятный том в честь профессора И. П. Сингха . Публикации Миттала. ISBN 978-81-7099-328-5 . [ нужна страница ]
- ^ Кумар Д. (2012). Генетические заболевания Индийского субконтинента . Springer Science & Business Media. ISBN 978-1-4020-2231-9 . [ нужна страница ]
- ^ Перейти обратно: а б Кивисилд Т., Роотси С., Метспалу М., Мастана С., Калдма К., Парик Дж. и др. (февраль 2003 г.). «Генетическое наследие первых поселенцев сохраняется как в индийских племенах, так и в кастах» . Американский журнал генетики человека . 72 (2): 313–332. дои : 10.1086/346068 . ПМК 379225 . ПМИД 12536373 .
- ^ Ранавира, Ланка; Каевсутти, Супанни; Вин Тун, Аунг; Буньярит, Хатайханоке; Пулсуван, Самерчай; Лертрит, Патчари (январь 2014 г.). «История митохондриальной ДНК этнических народов Шри-Ланки: их отношения внутри острова и с индийским субконтинентальным населением» . Журнал генетики человека . 59 (1): 28–36. дои : 10.1038/jhg.2013.112 . ПМИД 24196378 .
- ^ Ранасингхе, Руванди; Теннекун, Камани Х.; Карунанаяке, Эрик Х.; Лембринг, Мария; Аллен, Мари (ноябрь 2015 г.). «Исследование генетических полиморфизмов в гипервариабельных областях I и II митохондриальной ДНК пяти основных этнических групп и населения Ведды в Шри-Ланке». Юридическая медицина . 17 (6): 539–546. doi : 10.1016/j.legalmed.2015.05.007 . ПМИД 26065620 .
- ^ Марреро, П.; Абу-Амеро, КК; Ларруга, Дж. М.; Кабрера, В.М. (2016). «Носители митохондриальной ДНК человека макрогаплогруппы М колонизировали Индию из Юго-Восточной Азии» . Эволюционная биология BMC . 16 (1): 246. дои : 10.1186/s12862-016-0816-8 . ПМК 5105315 . ПМИД 27832758 .
- ^ Мецпалу М., Кивисилд Т., Мецпалу Е., Парик Дж., Худжашов Г., Калдма К. и др. (август 2004 г.). «Большая часть существующих границ мтДНК на юге и юго-западе Азии, вероятно, сформировалась во время первоначального заселения Евразии анатомически современными людьми» . БМК Генетика . 5:26 . дои : 10.1186/1471-2156-5-26 . ПМК 516768 . ПМИД 15339343 .
- ^ Чаубей, Г. Языковые изоляты и их генетическая идентичность: комментарий к истории митохондриальной ДНК этнических народов Шри-Ланки: их отношения внутри острова и с индийским субконтинентальным населением. Дж Хум Жене 59, 61–63 (2014). https://doi.org/10.1038/jhg.2013.122
| Подтемы | |
|---|---|
| Генетическая история по регионам | |
| Популяционная генетика по группе |
|