Генетические исследования марокканцев

Марокканская генетика охватывает генетическую историю народа Марокко и генетическое влияние этого происхождения на население мира. На это сильно повлияла география.
В доисторические времена пустыня Сахара на юге и Средиземное море на севере были важными географическими барьерами для притока мигрантов из стран Африки к югу от Сахары и Европы . Однако Западная Азия и Северная Африка образуют единый массив суши на Синае . Неолитическое население Западной Азии также могло быть привлечено к влажной Сахаре, прибывающей либо через Суэц , Баб-эль-Мандеб, либо через Средиземное море.
В результате этих географических влияний генетический профиль марокканского населения представляет собой сложную мозаику автохтонных магрибских линий, а также североафриканских , европейских , западноазиатских и западноафриканских элементов в различной степени. Хотя Северо-Западная Африка испытала поток генов из окружающих регионов, в некоторых частях она также пережила длительные периоды генетической изоляции. Это позволило развить отличительные генетические маркеры в некоторых популяциях Магриба, особенно в некоторых изолированных бербероязычных группах.
Население Марокко генетически неоднородно. [2] В основном это связано с миграцией арабов в Магриб , демографическим процессом, который в значительной степени повлек за собой поток генов и изменил генетическую структуру Магриба. [3] Хромосомный пул J-M267 возник не только в результате значительного ранней неолитической революции расселения в эпоху из Западной Азии , но и в гораздо большей степени в результате недавней экспансии арабских племен с Аравийского полуострова , во время которой к южным кахтанитам и северным аднанитам присоединились арабы. гетерогенный этнический плавильный котел. [4]
Предыстория и древность
[ редактировать ]Считалось, что территория современного Марокко была заселена со времен палеолита , где-то между 90 000 и 190 000 годами до нашей эры, но это уже не так после открытия 300 000-летнего человека разумного , и вместо этого теперь предполагается, что По тем же свидетельствам он был населен людьми с первобытных времен. [5] Во времена верхнего палеолита Магриб саванну был более плодородным, чем сегодня, и больше напоминал , чем сегодняшний засушливый ландшафт. [6] 22 000 лет назад атерийскую культуру, которая имела сходство с кроманьонскими культурами Евразии, сменила иберомаврская культура, которая имела сходство с иберийскими культурами. Сходство скелетов наблюдалось между иберомаврскими захоронениями Мечта-Афалу и останками европейских кроманьонцев . На смену иберомаврской индустрии в Марокко пришла капсийская культура .




Северная Африка и Марокко постепенно были втянуты в более широкий развивающийся средиземноморский мир западноазиатскими семитскими финикийцами , которые основали торговые колонии и поселения в ранний классический период. [7] Могадор был финикийской колонией еще в начале VI века до нашей эры. [8] [ нужна страница ]
Позже Марокко вошло в состав Финокийской Северо-Африканской и Иберийской империи со штаб-квартирой в Карфагене . Самым ранним известным независимым марокканским государством было берберское королевство Мавретания под властью короля Бокха I. Это королевство на севере Марокко, не путать с нынешним государством Мавритания , датируется как минимум 110 г. до н.э. [9]
Римская империя контролировала этот регион с I века до нашей эры, назвав его Мавретания Тингитана . Христианство было введено во 2 веке нашей эры и приобрело новообращенных в римских городах, среди рабов и некоторых берберских фермеров.
В V веке нашей эры, когда Римская империя пришла в упадок, в этот регион с севера вторглись сначала германские вандалы , а затем вестготы . В VI веке нашей эры север Марокко номинально входил в состав Восточной Римской или Византийской империи . Все это время берберские жители высоких гор внутреннего Марокко оставались непокоренными.
История
[ редактировать ]Ранняя арабская эпоха
[ редактировать ]В 670 году нашей эры первое арабское завоевание прибрежной равнины Северной Африки произошло под руководством Укбы ибн Нафи , генерала, служившего под началом Омейядов в Дамаске . Мусульмане Омейядов принесли в Марокко свой язык, систему правления и ислам. Многие берберы постепенно приняли ислам, в основном после того, как арабское правление отступило. Первым независимым берберским государством на территории современного Марокко было Королевство Некор , эмират в горах Риф . Он был основан Салихом I ибн Мансуром в 710 году как государство-клиент Халифата Рашидун . После начала Великого берберского восстания в 739 году берберы сформировали другие независимые государства, такие Микнаса Сиджилмаса как и Баргавата .
Идрис ибн Абдаллах бежал в Марокко после резни Аббасидами его племени в Ираке. Он убедил берберские племена Авраба отказаться от своей верности далеким халифам Аббасидов в Багдаде и основал династию Идрисидов в 788 году. Идрисиды сделали Фес своей столицей, а Марокко стало центром мусульманского образования и крупной региональной державой . Идриссиды были изгнаны в 927 году Фатимидским халифатом и их союзниками-микнасами. После того, как Микнаса разорвал отношения с Фатимидами в 932 году, они были отстранены от власти магравой Сиджилмасы в 980 году.
Берберские династии
[ редактировать ]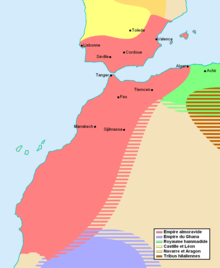
Начиная с XI века, ряд могущественных берберских [10] [11] [12] возникли династии. При династии Альморавидов [13] и династия Альмохадов доминировала в Магрибе, на большей части территории современной Испании и Португалии, а также в западном Средиземноморье. В XIII и XIV веках Мериниды удерживали власть в Марокко и стремились повторить успехи Альмохадов посредством военных кампаний в Алжире и Иберии. За ними последовали Ваттасиды . В 15 веке Реконкиста положила конец мусульманскому правлению в центральной и южной Иберии, и многие мусульмане и евреи бежали в Марокко. [14] Усилия Португалии по контролю над Атлантическим побережьем в 15 веке не сильно повлияли на внутренние районы Марокко. По словам Элизабет Алло Исичеи, «в 1520 году в Марокко был такой ужасный голод , что долгое время им датировали и другие события. Было высказано предположение, что население Марокко упало с 5 до менее 3 миллионов человек в период с начала шестнадцатого века и девятнадцатого века». [15]
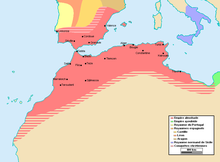
Арабские династии
[ редактировать ]В 1549 году регион перешел к сменяющим друг друга арабским династиям, заявлявшим о своем происхождении от исламского пророка Мухаммеда династия : сначала династия Саади , правившая с 1549 по 1659 год, а затем Алауитов , остававшаяся у власти с 17 века.

При династии Саади страна отразила османские вторжения и португальское вторжение в битве при Ксар-эль-Кебире в 1578 году. Правление Ахмада аль-Мансура принесло султанату новое богатство и престиж, а большая экспедиция в Западную Африку нанесла сокрушительный удар. поражение империи Сонгай в 1591 году. Однако управление территориями через Сахару оказалось слишком трудным. После смерти аль-Мансура страна была разделена между его сыновьями.
В 1666 году Марокко было воссоединено династией Алауитов , которая с тех пор является правящим домом Марокко. Марокко столкнулось с агрессией со стороны Испании, а Османская империя продвигалась на запад. Алауитам удалось стабилизировать свое положение, и хотя королевство было меньше предыдущих в регионе, оно оставалось довольно богатым. Несмотря на сопротивление местных племен Исмаил ибн Шариф (1672–1727) приступил к созданию единого государства. [16] Со своим «Джейш д'Аль ар-Риф» ( Рифианская армия) он захватил Танжер у англичан в 1684 году и изгнал испанцев из Лараша в 1689 году.
Марокко было первой страной, признавшей молодые Соединенные Штаты в качестве независимого государства в 1777 году. [17] [18] [19] [20] В начале американской революции американские торговые суда в Атлантическом океане подверглись нападению берберийских пиратов . 20 декабря 1777 года султан Марокко Мухаммед III заявил, что американские торговые суда будут находиться под защитой султаната и, таким образом, смогут пользоваться безопасным проходом. Мароккано -американский договор о дружбе , подписанный в 1786 году, является старейшим нерушимым договором о дружбе между США . [21] [22]
Арабский приток
[ редактировать ]Было несколько волн арабской миграции в Марокко. Самой значительной волной была миграция Бану Хилал , Бану Сулайм и Макил в 11 веке. Они переселились в район современного Марокко в XII веке, когда Альмохадов правитель Абд аль-Мумин переселил их туда в большом количестве и поселил на атлантических равнинах, в регионе, ранее населенном племенной группой баргавата . Этот регион был в значительной степени разрушен и обезлюден Альморавидами , затем снова обезлюден Альмохадами в 1149–1150 и 1197–1198 годах, прежде чем заменить обезлюдевшую территорию арабскими мигрантами-бедуинами. Альмохады помогли арабским племенам преодолеть барьеры Атласских гор и ускорили их экспансию в Марокко, чтобы завершить кочевое господство бедуинов над низменностями Магриба вплоть до прибрежных равнин Атлантического океана. [23] Появление арабов усложнило этническое население Марокко и внесло значительный неберберский элемент. Они играли все более важную роль в политике Империи Альмохадов. Правитель Альмохадов Абд аль-Мумин ожидал сопротивления со стороны Масмуды , для которого он был чужаком, поэтому он заручился поддержкой арабов, чтобы обеспечить преемственность своего сына. С упадком армии Альмохадов арабские бедуины стали самой могущественной силой на марокканских равнинах, и ни один правитель не смог бы удержать там власть без их поддержки. [23]
При Маринидах значение арабов в Марокко возросло. Из-за отсутствия сторонников Зената они приветствовали поддержку арабских кочевников, которые уже начали проникать в страну при Альмохадах. Зената были сильно ассимилированы с арабской культурой, а Маринид Махзан (правительство) состояло как из арабов, так и из Зената. Это привело к экспансии арабских племен в Марокко, где они поселились на равнинах, а многие группы берберов были арабизированы . При Маринидах арабский язык стал как общим, так и официальным языком. [24] Этот демографический процесс в значительной степени подразумевал поток генов и ремоделировал генетическую структуру. [3]
Генетические доказательства
[ редактировать ]В генетическом исследовании, опубликованном в январе 2012 года, говорится, что коренные жители Северо-Западной Африки, по-видимому, наиболее тесно связаны с популяциями за пределами Африки , но «расхождение между марокканцами и ближневосточными/ европейцами, вероятно, предшествует голоцену (> 12 000 лет назад) и палеолиту (> 40 000 лет назад). до нашей эры)». [25]
Более ранние исследования ясно показывают, что существенных генетических между арабоязычным марокканским населением и неарабоязычным марокканским населением не существует различий. Сами берберы составляют генетически разнообразные группы и обладают высокой степенью генетической гетерогенности. [3] человеческого лейкоцитарного антигена HLA Данные ДНК позволяют предположить, что большинство марокканцев , как неарабской этнолингвистической идентичности, так и арабской этнолингвистической идентичности, имеют берберское происхождение, и что генеалогические истинные арабы из Аравии, вторгшиеся в Северную Африку и некоторые части Южной Европы, действительно не вносят существенного вклада в генофонд. [26] [27] Однако результаты недавнего исследования 2017 года показали, что миграция арабов в Магриб была демографическим процессом, который подразумевал поток генов и изменил генетическую структуру Магриба, а не просто культурным процессом, как предполагалось ранее в более старых записях. . [3]
Согласно статье 2000 года в Европейском журнале генетики человека , марокканцы из Северо-Западной Африки, хотя и имеют множество различий, генетически ближе к южным европейцам , чем к африканцам южнее Сахары этнической принадлежности банту . [28]
Примерно у трети марокканцев присутствует значительный вклад ДНК стран Африки к югу от Сахары, при этом у большинства западноевразийских берберов вклад ДНК составляет в среднем 1–10% ДНК стран Африки к югу от Сахары. [29] Неберберское население продемонстрировало значительно больший вклад ДНК африканских стран к югу от Сахары (до 55%). Что касается вклада стран Африки к югу от Сахары, различия среди берберского населения не были значительными. Кудрей и др. (2009) и Эрнандес и др. (2015) показали повышенное представительство гаплогрупп мтДНК из стран Африки к югу от Сахары у фигуигских берберов (т.е. высокую распространенность линий, происходящих от L).
Различные изученные локусы выявили близкое сходство между берберами и другими североафриканскими группами, в основном с марокканскими арабоговорящими людьми, что согласуется с гипотезой о том, что нынешнее марокканское население имеет сильное берберское происхождение. [30] С другой стороны, различные исследования также показали, что население Марокко генетически гетерогенно и разнообразно. [2] и что даже сами берберы составляют генетически разнообразные группы. [3] Различные исследования популяционной генетики совместно с такими историками, как Габриэль Кэмпс и Шарль-Андре Жюльен, подтверждают идею о том, что основная часть генофонда современных африканцев Северо-Запада , независимо от языковой группы, имеет неолитическое происхождение. [31] Различные более поздние исследования выступили против этого и пришли к выводу, что пул хромосом J-M267 произошел не только от раннего неолита , но и в гораздо большей степени от недавнего расширения арабских племен с Аравийского полуострова . [4] что подразумевало поток генов и ремоделировало генетическую структуру Магриба. [3]
Марокканская Y-ДНК-хромосома
[ редактировать ]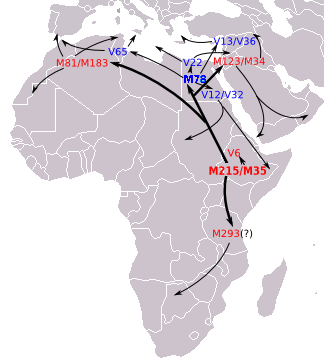
Э-М81
[ редактировать ]
E1b1b1b1 (E-M81) , ранее E1b1b1b , E3b1b и E3b2 , является наиболее распространенной гаплогруппой Y-хромосомы в Марокко , в которой доминирует ее субклад E-M183 . Считается, что он возник в Северной Африке 14 200 лет назад. [34]
его родительская клада E1b1b (E-M215) Считается, что впервые появилась на Африканском Роге около 42 600 лет назад. [35] Считается, что его доминирующий субклад E-M35 возник в Восточной Африке около 22 400 лет назад, а позже распространился в Северную Африку, а оттуда в Западную Азию. [36] [37] Считается, что все основные ответвления E-M35 возникли в той же области, что и родительская клада: в Северной Африке , на Африканском Роге или в близлежащих районах Ближнего Востока . Некоторые ветви E-M35 покинули Африку много тысяч лет назад. Например, Батталья и др. (2008) подсчитали, что E-M78 (названный в этой статье E1b1b1a1) существует в Европе уже более 10 000 лет. [38]
Эта гаплогруппа достигает средней частоты 85% в Северной Африке . Его частота снижается примерно с 80% и более в некоторых популяциях марокканских берберов, включая сахарцев, до примерно 28% к востоку от этого диапазона в Египте. [31] [37] [39]
Из-за распространенности клады среди этих берберских групп и других, таких как мозабиты , риффианцы , хлеухи , средний атлас и кабильцы , ее иногда называют генетическим берберским маркером .
Это филогенетическое древо субкладов берберских гаплогрупп основано на дереве YCC 2008 года и последующих опубликованных исследованиях, обобщенных ISOGG. [40] [41] [42]
- E1b1b1b (L19, V257)
- E1b1b1b1 (M81)
- E1b1b1b1a (M107) Underhill et al. (2000) .
- E1b1b1b1b (M183) Эта клада чрезвычайно доминирует в E-M81. Фактически, хотя Карафет и др. (2008) продолжают описывать это как субклад E-M81, а ISOGG ссылается на Karafet et al., все данные, по-видимому, подразумевают, что его действительно следует считать филогенетически эквивалентным M81. [ нужна ссылка ]
- E1b1b1b1b1 (M165) Underhill et al. (2000) .
- E1b1b1b1b2 (L351) Обнаружен у двух родственных участников проекта «Филогения E-M35» .
- E1b1b1b1 (M81)
Среднестатистические североафриканские марокканские берберы имеют частоту E3b3 +80%. Альварес и др. (2009) исследование показывает частоту E3b1b 28/33 или 84,8% у берберов из Марракеша . При остальных частотах 1/33=3% E3a*, 1/33=3% E3b*, 1/33 или 3% E3b1a и 1/33 или 3% E3b1c. [37]
E1b1b (M81) — протоберберский маркер . E1b1b1b1a1 (M107) — сокращенная протоберберская линия в Мали . [43]
J-P209
[ редактировать ]
Гаплогруппа J-P209 — вторая по распространенности Y-хромосома в Марокко, в которой доминирует субклад J-M267 (J1). Считается, что он возник примерно 31 700 лет назад в Юго-Западной Азии (31 700 ± 12 800 лет назад, согласно Semino et al. 2004 ).Самая высокая частота - 30–62,5% - наблюдалась среди арабского мусульманского населения на Ближнем Востоке. [4]
Гаплогруппа J-P209 встречается в наибольшей концентрации на юго-западе Аравийского полуострова . гаплогруппа J-P209 широко распространена в Северной Африке , являясь второй по распространенности гаплогруппой в Магрибе. Исследование 2021 года показало, что самая высокая частота ближневосточного компонента, когда-либо наблюдавшаяся в Северной Африке, наблюдалась у арабов Веслетии в Тунисе , у которых частота ближневосточного компонента составляла 71,8%. [44] J-P209 был обнаружен с частотой 35% в Алжире , 34% в Тунисе. [45] [46] и 20% в Египте ( Семино и др., 2004 ). В Марокко J-M267 встречается с частотой 26,4%, [47] 31.5%, [48] и 20,4%. [49]

Недавнее исследование показало, что пул хромосом J-M267 в Магрибе происходит не только от расселения раннего неолита , но и в гораздо большей степени от арабских миграций в Магриб , недавней экспансии арабских племен с Аравийского полуострова , во время которой оба Южные кахтаниты и северные аднанитские арабы пополнили гетерогенный этнический плавильный котел Магриба. [4] Это в значительной степени подразумевало поток генов и ремоделировало генетическую структуру стран Магриба, а не было простой культурной заменой, как утверждалось в более старых исследованиях. [3] Это увеличило генетическое сходство между жителями Северной Африки и Ближнего Востока. [50]
Э-М78
[ редактировать ]
Самая базальная и редкая парагруппа E-M78 * обнаружена с более низкими частотами у марокканских арабов. Субклад E-V65 встречается в больших количествах в регионах Магриба на крайнем севере Африки. Кручиани и др. (2007) сообщают об уровне около 20% среди ливийских арабов и около 30% среди марокканских арабов . Кажется, что это менее распространено среди берберов , но все же присутствует на уровне> 10%. Авторы предполагают североафриканское происхождение этой линии. В Европе лишь несколько особей были обнаружены в Италии и Греции. [ нужна ссылка ]
Капелли и др. (2009) изучали бета-кластер в Европе. Они обнаружили небольшие количества в Южной Италии, а также следы в Кантабрии, Португалии и Галисии, при этом в Кантабрии был самый высокий уровень в Европе по их исследованию - 3,1% (5 из 161 человека).
О других частотах E1b1b1a1c (E-V22) сообщает Cruciani et al. (2007) включают марокканских арабов (7,27%, 55 человек) и марокканских евреев (8%, 50 человек).
Другие гаплогруппы
[ редактировать ]
Что касается E-M123, то без проверки SNP E-M34 встречается с небольшой частотой в Марокко. Низкий региональный процент E-M123 был зарегистрирован у марокканских берберов ( около 3%). E-M123 также известен как E1b1b1b2a (ISOGG 2012).
Евразийские гаплогруппы, такие как гаплогруппа J и гаплогруппа R1, также наблюдались с очень минимальной частотой. Тщательное исследование Cruciani et al. (2004), проанализировавшие население Марокко, пришли к выводу, что североафриканская модель вариаций Y-хромосомы (включая гаплогруппы J1 и R1b) в значительной степени имеет неолитическое происхождение, что позволяет предположить, что переход к неолиту в этой части мира сопровождался демической диффузией. бербероязычных скотоводов из Алжирской пустыни в Восточное Марокко, хотя более поздние документы предполагают, что эта дата могла произойти уже десять тысяч лет назад, с переходом от оранской к капсийской культуре в Северной Африке. [49] [51]
Гаплогруппы G и T редко встречаются в Марокко. В 147 образцах, взятых в Марокко , 1% оказался G. [52]
В другом исследовании 1% из 312 образцов в Марокко были G. [53]
В другом исследовании были собраны образцы только из деревень в долине Азгур в Марокко, где ни в одном из 33 образцов не было обнаружено G. [37] Эти деревни были выбраны потому, что по составу они считались типично берберскими .
Исследование 20 марокканских евреев показало, что 30% из них были Г. [37] Испытуемые тогда, очевидно, проживали в Израиле . Другое исследование еврейских мужчин показало, что 19,3% из 83 евреев-мужчин из Марокко принадлежали к гаплогруппе G. [54] Более G марокканские образцы, вероятно, положительны по гаплогруппе SNP G2a2b, она была идентифицирована в неолитических человеческих останках в Европе, датируемых между 5000 и 3000 годами до нашей эры.Более того, большинство всех мужских скелетов периода европейского неолита до сих пор содержали Y-ДНК, принадлежащую к этой гаплогруппе, как и мумифицированные останки Эци Ледяного человека . Национальное географическое общество полагает, что гаплогруппа G возникла на Ближнем Востоке 30 000 лет назад и предполагает, что люди, несущие гаплогруппу, принимали участие в распространении неолита в Африку, а затем и в Европу [55] Два процента арабов -марокканцев и от 0% до 8% берберов-марокканцев оазиса Асни также оказались представителями G. [56]
Гаплогруппа T встречается среди центральных берберов оазиса Асни недалеко от границ Алжира в 1,9% и наблюдается у марокканских евреев в 4%.
Самая базальная и редкая парагруппа E1a* была обнаружена на более низких частотах в образцах, полученных от марокканских берберов и сахарцев. датируется примерно 45 000 г. до н.э. Связан с задней евразийской миграцией с Ближнего Востока в Северную Африку вместе с E1b1b во времена палеолита. [56]
Марокканские гаплогруппы Y-ДНК
[ редактировать ]| Население | н | А/Б | Э-М33 | Э-В38 | Э-М35* | Э-М78 | Э-М81 | Э-М123 | Г | Дж | Р | Ссылка |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Марокко | 760 | 0.9 | 2.7 | 3.2 | 4.2 | 6.8 | 67.3 | 0.6 | 0.6 | 7.6 | 4.4 | Бекада и др. 2013 год [57] |
| Марокко | 87 | — | — | 9.2 | — | 5.7 | 52.8 | — | — | 26.4 | — | Фадлауи-Зид и др. 2013 год [47] |
| Марокко | 221 | — | 1.8 | 4.5 | 4 | 6.8 | 65 | — | — | 9 | 4 | Фрегель и др. 2009 год [58] |
| Марокко | 51 | 4 | 6 | — | 6 | 6 | 55 | — | — | 20 | 4 | Онофри и др. 2008 год [59] |
| Марокко | 176 | — | — | 6.3 | 5.1 | 6.3 | 63.6 | — | — | 13.6 | 2.8 | Бош и др. 2001 г. [60] |
| Арабы ( Марокко ) | 49 | — | — | — | — | 42.9 | 32.6 | — | — | 20.4 | — | Семино и др. 2004 г. [49] |
| Арабы ( Марокко ) | 44 | — | — | 6.8 | 2.2 | 11.3 | 52.2 | — | — | 15.9 | 6.8 | Бош и др. 2001 г. [60] |
| Арабы ( Марокко ) | 54 | — | — | — | — | 38.9 | 31.5 | — | — | — | — | Кручиани и др. 2004 г. [61] |
| Берберы ( Марокко ) | 64 | — | — | — | — | 10.9 | 68.7 | — | — | 6.3 | — | Семино и др. 2004 г. [49] |
| Берберы ( Марракеш ) | 29 | — | — | — | 3.4 | 6.9 | 72.4 | — | — | — | — | Кручиани и др. 2004 г. [62] |
| Берберы ( Средний Атлас ) | 69 | — | — | — | — | 10.1 | 71 | — | 4.3 | 5.8 | — | Кручиани и др. 2004 г. [62] |
| Берберы (Южное Марокко ) | 40 | — | — | 2.5 | 7.5 | 12.5 | 65 | — | — | 10 | — | Бош и др. 2001 г. [60] |
| Берберы (Северо-Центральный) | 63 | — | 3.1 | 9.5 | 7.9 | 1.5 | 65 | — | — | 11.1 | — | Бош и др. 2001 г. [60] |
| Берберы (Амизмиз) | 33 | 3 | — | 3 | 3 | 3 | 84.8 | 3 | — | — | — | Альварес и др. 2009 год [37] |
| Берберы (Асни) | 54 | — | — | — | 1.9 | 3.7 | 79.6 | — | — | 1.9 | 1.9 | Дюгужон и др. (2005) [63] |
| Берберы (Сиди Бухрия - Бени Знассен) | 67 | — | — | — | — | 1.5 | 77.6 | — | 6 | 1.5 | 6 | Дюгужон и др. (2005) [63] |
| Берберы (Северное Марокко) | 43 | — | — | — | — | — | 79.1 | — | — | — | — | Ахмед Региг и др. 2014 год [64] |
| Берберы (Южное Марокко) | 65 | — | — | — | — | — | 98.5 | — | — | — | — | Ахмед Региг и др. 2014 год [64] |
| Берберы (Центральное Марокко) | 187 | — | — | — | — | — | 89.8 | — | — | — | — | Ахмед Региг и др. 2014 год [64] |
| Марокканский сахарский | 189 | 0.5 | 5.2 | 6.8 | — | — | 55.5 | 11.1 | — | 13.2 | 7.2 | Бекада и др. 2013 год [57] |
| Марокканский сахарский | 89 | — | 8.9 | 11.2 | — | — | 59.5 | — | — | 20.2 | — | Фрегель и др. 2009 год [58] |
| Марокканский сахарский | 29 | — | 3.4 | 3.4 | — | — | 75.8 | — | — | 17.2 | — | Бош и др. 2001 г. [60] |
| Марокканские евреи | 19 | — | — | — | 21.1 | — | — | — | 26.3 | 31.5 | 10.5 | Франкалаччи и др. 2008 год [48] |
Распределение гаплогрупп в марокканском населении
[ редактировать ]Основные компоненты гаплогрупп Y-ДНК, присутствующие у марокканских берберов (E3b; 94%), являются общими с европейскими и соседними популяциями Северной Африки и Ближнего Востока. Незначительная доля гаплогрупп также включает те, которые относятся к африканцам Северо-Западной Африки (E1a, A1a; 1%), жителям Ближнего Востока (J, G, T; 2,4%), африканцам к югу от Сахары (E3a; 1,7%) и европейцам ( R1b, I1; 2%) сродство.
Некоторые из основных выявленных процентов были следующими:
- E1b1b : 56% [ нужна ссылка ] - Типично для афроазиатскоязычного населения.
- J-P209 : 20,4% ( Семино и др. 2004 ) — типичен для популяций Аравийского полуострова, Леванта и Кавказа , с умеренным распространением в Юго-Восточной Европе , Северной Африке, на Африканском Роге, в Средней Азии и Южной Азии .
- R1b : от 0,8% до 6,8% ( Bosch 2001 ) — типично для западноевропейцев , некоторых народов Западной Азии , суданских фулани и чадскоязычных народов Центральной Африки, а также некоторых народов Центральной Азии (таких как башкиры , туркмены и уйгуры ) . [65]
- Г : 0,4% — характерно для выходцев с Кавказа и в меньшей степени Ближнего Востока .
- E1a : 0,5% — редкая гаплогруппа, обнаруженная среди марокканских берберов , сахрави , южных европейцев и некоторых носителей чадского языка в Сахеле.
- E1b1a : 1,7% — типично для нигерско-конгоязычного населения.
- Т : 0,4% - Широко распространен в Западной Евразии.
- I : 0,4% - Его можно обнаружить у большинства современных европейских популяций, с пиком в Северной и Юго-Восточной Европе. Y-хромосомы гаплогруппы I1 также обнаружены у некоторых популяций Ближнего Востока , Кавказа, Северо-Восточной Африки и Центральной Сибири .
Берберская генетическая идентичность марокканцев
[ редактировать ]
Доисторическое население Марокко, которое было предком берберов, было связано с более широкой группой палео-средиземноморских народов.Афроазиатская семья , вероятно, возникла в период мезолита, возможно, в контексте капсийской культуры . [66] [67] Анализ ДНК обнаружил сходство между берберским населением Марокко и населением саамов в Скандинавии, что указывает на связь, датируемую примерно 9000 лет назад. [68]
Около 5000 г. до н.э. население Северной Африки в основном происходило от создателей иберомаврской и капсийской культур, а более позднее вторжение было связано с неолитической революцией . [69] Протоберберские племена произошли от этих доисторических сообществ в период от поздней бронзы до раннего железного века. [70]
Генетические доисторические расширения
[ редактировать ]Данные ДНК показывают, что во время последнего ледникового максимума , периода между 25 000 и 19 000 лет назад, большие ледяные щиты толщиной более километра покрывали большую часть Северной Европы, делая этот регион непригодным для жизни людей. Считается, что человеческое население отступило на юг, в более теплые регионы вблизи Средиземноморья. Считается, что беженцы в этот период находились в Иберии , на Балканах и в Италии , и поэтому произошел некоторый поток генов из Северной Африки в Южную Европу . [71]
Считается, что после ледникового максимума, когда европейский климат потеплел, убежища стали источником повторного заселения Европы. Доисторические африканские племена, которые были завезены в Иберию в качестве беженцев, затем рассеялись по всей Европе с экспансией людей на север. Это могло бы объяснить наличие генетических линий в Восточной Европе и даже на севере России, которые, по-видимому, имеют доисторические связи с Северо-Западной Африкой, главным образом с Марокко (см. мтДНК ). [71] Считается также, что расширение человеческого населения из иберийских убежищ переместилось обратно в Марокко и Северо-Западную Африку . [72]
Неолит до конца доисторического периода
[ редактировать ]Переход от охоты и собирательства к сельскому хозяйству во время неолитической революции стал переломным моментом в мировой истории. Считается, что общества, которые первыми перешли к сельскому хозяйству, жили в Северной Африке и на Ближнем Востоке около 10 000 г. до н.э. Сельское хозяйство было завезено в Европу мигрирующими фермерами с Ближнего Востока. [73] Согласно модели демической диффузии , эти ближневосточные фермеры либо заменяли, либо скрещивались с местными популяциями охотников-собирателей, которые жили в Европе после миграции «из Африки». [74]
Было высказано предположение, что первые фермеры Ближнего Востока испытали Северной Африки, влияние главным образом, из-за капсийской культуры . [75] Были предположения, что некоторые генетические линии, найденные на Ближнем Востоке, прибыли туда в этот период. [76] Обычно считается, что первые сельскохозяйственные общества на Ближнем Востоке возникли из натуфийской культуры , существовавшей в Палестине с 12 000 до 10 000 лет до нашей эры. Важная миграция из Северо-Западной Африки, произошедшая иберо-маврикийцами из Марокко через Синай , по-видимому, произошла до образования натуфийцев. [77]
Генетическая преемственность в Марокко
[ редактировать ]В 2013 году скелеты, принадлежащие создателям эпипалеолитической иберомаврской культуры , которые были раскопаны на доисторических стоянках Тафоральт и Афалу , были проанализированы на предмет древней ДНК. Все экземпляры принадлежали к материнским кладам, связанным либо с Северной Африкой, либо с северным и южным побережьем Средиземноморья, что указывает на поток генов между этими областями со времен эпипалеолита. [78] Древние особи Тафоральта несли гаплогруппу Y-ДНК E1b1b и гаплогруппы мтДНК U6 , H , JT и V , что указывает на преемственность населения в регионе, начиная с иберомаврийского периода. [79] [80]
Согласно исследованию Фадлауи-Зида, проведенному в 2004 году, эта закономерность предполагает, что арабизация региона была в основном культурным процессом, а не демографической заменой берберского населения, населявшего регион, где происходила арабская экспансия. [81] Однако результаты более недавнего исследования 2017 года показали, что миграция арабов в Магриб была в основном демографическим процессом, который подразумевал поток генов и ремоделировал генетическую структуру, а не просто культурную замену, как предполагалось ранее историческими записями. [3]
Марокканская митохондриальная мтДНК
[ редактировать ]Марокканский митохондриальный пул по существу является берберским по своей структуре и характеризуется «в целом высокой частотой западноевразийских гаплогрупп». Представлен послепоследним ледниковым максимальным распространением от Иберии до Северной Африки, выявленным путем точной характеристики гаплогруппы HV мтДНК в Марокко. от 36% до 60%, несколько более низкая частота линий L к югу от Сахары и значительное (но дифференциальное) присутствие североафриканских гаплогрупп U6 и M1». [82] И по данным Черни и др. 2008 г. «Максимальное расширение после последнего ледникового периода, начавшееся в Иберии, привело не только к переселению Европы, но и Северной Африки». [83]
Евразийские (материнские) последовательности мтДНК были обнаружены с частотой 96% у марокканских берберов, 82% у алжирских берберов и 78% у неберберских марокканцев, по сравнению с только 4% в сенегальском населении. [ нужна ссылка ]
До недавнего времени в некоторых работах предполагалось, что распространение основных L-гаплогрупп в Марокко происходило главным образом за счет транссахарской работорговли. [84] Однако в сентябре 2010 года тщательное исследование берберской мтДНК, проведенное Сабе Фриги, пришло к выводу, что некоторые гаплогруппы, такие как L3*, были намного старше и занесены в древний африканский поток генов около 20 000 лет назад, в то время как другие, такие как L2a и L3b, имели более поздние гаплогруппы. происхождение. [85]
Марокканские северные берберы имеют только от 3% до 1% мтДНК SSA. Этот градиент с севера на юг во вкладе стран к югу от Сахары в генофонд подтверждается Esteban et al. , [86] остальные линии мтДНК в основном являются европеоидными/западноевразийскими, в то время как марокканские арабы имеют более повышенную материнскую примесь SSA - примерно от 21% до 36%. По данным последовательностей L-мтДНК, самые высокие частоты L-мтДНК наблюдаются у марокканских арабов из прилегающей территории. Эль -Джадида составляет 36%, и это в основном связано с работорговлей . [87]
Частоты (> 1%) L-мтДНК
| Страна | Этническая группа | Количество протестированных | Ссылка | L-мтДНК% |
| Марокко | Марокканец (евреи) | 149 | Бехар и др. (2008) | 1.34% |
| Марокко | Марокканский Северный (берберы) | 124 | Эстебан и др. (2004) | 1% |
| Марокко | Марокканец (арабы) | 81 | Харич и др. (2010) | 36% |
| Марокко | Марокканские арабы | 56 | Турчи и др. (2009) | 25.00% |
| Марокко | Марокканский Южный (берберы) | 64 | Турчи и др. (2009) | 26.00% |
См. также
[ редактировать ]- Демография Марокко
- Генетическая история Северной Африки
- Генетическая история Ближнего Востока
- Генетические исследования арабов
Ссылки
[ редактировать ]- ^ Y Гаплогруппы мира (PDF) Университет Иллинойса, Дж. Д. Макдональд, 2005 г.
- ^ Jump up to: а б Агути, И.; Баденс, К.; Абуюб, А.; Леви, Н.; Беннани, М. (декабрь 2008 г.). «Молекулярная основа бета-талассемии в Марокко: возможные причины молекулярной гетерогенности» . Генетическое тестирование . 12 (4): 563–568. дои : 10.1089/gte.2008.0058 . ПМИД 18976160 . S2CID 46000591 .
- ^ Jump up to: а б с д и ж г час Эвол, Мол Биол (февраль 2017 г.). «Недавние исторические миграции сформировали генофонд арабов и берберов в Северной Африке» . Молекулярная биология и эволюция . 34 (2): 318–329. дои : 10.1093/molbev/msw218 . ПМЦ 5644363 . ПМИД 27744413 .
- ^ Jump up to: а б с д Жене, Am J Hum (июнь 2002 г.). «Генетические доказательства экспансии арабских племен в Южный Левант и Северную Африку» . Американский журнал генетики человека . 70 (6): 1594–1596. дои : 10.1086/340669 . ПМК 379148 . ПМИД 11992266 .
- ^ Полевые проекты - Джебель Ирхуд. Архивировано 12 января 2017 г. в Wayback Machine . Отдел эволюции человека. Институт эволюционной антропологии Макса Планка
- ^ Краснуха, Д. (1984). «Экологизм и экономика палеолита Пи в Магрибе (ок. 20 000–5 000 лет назад)». В JD Clark & SA Brandt (ред.). От охотников до фермеров: причины и последствия производства продуктов питания в Африке . Беркли: Издательство Калифорнийского университета. стр. 41–56. ISBN 978-0520045743 .
- ^ Мегалитический портал; Карта Мегалита. «К. Майкл Хоган, Могадор: Форт Мыс , Мегалитический портал, под ред. Энди Бернэма» . Megalithic.co.uk . Проверено 2 июня 2010 г.
- ^ Москати, Сабатино (2001) Финикийцы , Телец, ISBN 1-85043-533-2
- ^ «Бокх I» . Проверено 27 сентября 2010 г.
- ^ Рамирес-Фариа, Карлос (1 января 2007 г.). Краткая энциклопедия всемирной истории . Atlantic Publishers & Dist. ISBN 978-81-269-0775-5 .
- ^ «Альморавиды» . Энциклопедия Универсалис .
- ^ «Династия Маринидов» . Британская энциклопедия .
- ^ «Магриб при Альморавидах и Альмохадах» . Britannica.com . Проверено 1 августа 2011 г.
- ^ «Марокко – История» . Britannica.com . Проверено 1 августа 2011 г.
- ^ Алло Исичеи, Элизабет (1997). История африканских обществ до 1870 года . Издательство Кембриджского университета. п. 264. ISBN 0-521-45599-5
- ^ « Марокко (страница 8 из 9) ». Интернет-энциклопедия Microsoft Encarta, 2009. 01.11.2009.
- ^ «Совместное заявление Соединенных Штатов Америки и Королевства Марокко» . Белый дом . 22 ноября 2013 г. – через Национальный архив .
- ^ Руководство по внешней политике и правительству Марокко . Международные деловые публикации США. 2004. ISBN 9780739760000 . [ постоянная мертвая ссылка ]
- ^ «Новости Минобороны» .
- ^ Козарин, Линда Д. «Коэн возобновляет американо-марокканские связи» . Министерство обороны США . Проверено 12 марта 2009 г.
- ^ Робертс, Присцилла Х. и Ричард С. Робертс, Томас Барклай (1728–1793): консул во Франции, дипломат в Барбари , Lehigh University Press, 2008, стр. 206–223 ISBN 093422398X .
- ^ «Вехи американской дипломатии, интересные исторические заметки и история Государственного департамента» . Государственный департамент США . Проверено 17 декабря 2007 г.
- ^ Jump up to: а б Фадж, JD; Оливер, Роланд (1975). Кембриджская история Африки . Издательство Кембриджского университета. п. 343. ИСБН 978-0-521-20981-6 .
- ^ Ниан, Джибрил Тамсир; Африка, Международный научный комитет ЮНЕСКО по составлению всеобщей истории (1 января 1984 г.). Африка с двенадцатого по шестнадцатый век . ЮНЕСКО. п. 89. ИСБН 978-92-3-101710-0 .
- ^ Хенн, Б.М.; Ботиге, LR; Гравий, С.; Ван, В.; Брисбин, А.; Бирнс, Дж. К.; Фадлауи-Зид, К.; Заллуа, Пенсильвания; Морено-Эстрада, А. (2012). Шируп, Миккель Х (ред.). «Геномное происхождение жителей Северной Африки поддерживает миграцию обратно в Африку» . ПЛОС Генетика . 8 (1): e1002397. дои : 10.1371/journal.pgen.1002397 . ПМК 3257290 . ПМИД 22253600 .
- ^ Балле, Жан-Луи (2000) «Глава 7: Завоевания и деградация земель в восточном Магрибе» В Баркере, Грэме и Гилбертсоне, Дэвиде (2000) Археология засушливых земель: жизнь на окраине Рутледж, Лондон, Том 1, Часть III – Сахара и Сахель, стр. 125–136, ISBN 978-0-415-23001-8
- ^ Гомес-Касадо, Э.; Дель Мораль, П.; Мартинес-Ласо, Дж.; Гарсия-Гомес, А.; Альенде, Л.; Сильвера-Редондо, К.; Лонгас, Дж.; Гонсалес-Хевилла, М.; Кандил, М.; и др. (2000). «Гены HLA у арабоязычных марокканцев: тесное родство с берберами и иберийцами». Тканевые антигены . 55 (3): 239–249. дои : 10.1034/j.1399-0039.2000.550307.x . ПМИД 10777099 .
- ^ Бош, Э.; Калафель, Ф.; Перес-Лецион, А.; Кларимон, Дж.; Комас, Д.; Мэтью, Э.; Мартинес-Ариас, Р.; Морера, Б.; Бракес З.; и др. (2000). «Генетическая структура северо-западной Африки, выявленная с помощью STR-анализа» . Европейский журнал генетики человека . 8 (5): 360–366. дои : 10.1038/sj.ejhg.5200464 . ПМИД 10854096 .
- ^ Кальдерон, Росарио; Перейра, Луиза; Куэста, Педро; Новеллетто, Андреа; Дюгужон, Жан-Мишель; Санчес-Мартинес, Луис Х.; Лопес, Сайоа; Кавадас, Бруно; Пита, Гильермо; Эрнандес, Кандела Л. (2020). «Геномное разнообразие человека там, где Средиземное море соединяется с Атлантикой» . Молекулярная биология и эволюция . стр. 1041–1055. дои : 10.1093/molbev/msz288 . ПМК 7086172 . ПМИД 31816048 .
- ^ Харич, Н.; Эстебан, Э.; Чафик, А.; Лопес-Аломар, А.; Вона, Г.; Мораль, П. (2002). «Классические полиморфизмы у берберов из Мойен Атлас (Марокко): генетика, география и исторические свидетельства средиземноморских народов». Анналы биологии человека . 29 (5): 473–487. дои : 10.1080/03014460110104393 . ПМИД 12396367 . S2CID 34025436 .
- ^ Jump up to: а б Арреди, Б.; Полони, Э.; Параккини, С.; Зержал, Т.; Фаталлах, Д.; Макрелуф, М.; Паскали, В.; Новеллетто, А.; Тайлер-Смит, К. (2004). «Преимущественно неолитическое происхождение вариаций ДНК Y-хромосомы в Северной Африке» . Американский журнал генетики человека . 75 (2): 338–345. дои : 10.1086/423147 . ПМК 1216069 . ПМИД 15202071 .
- ^ Хенн и др. (2008) .
- ^ Хасан и др. (2008) .
- ^ «E-M81 YTree» . yfull.com . Проверено 10 июля 2016 г.
- ^ «Е-М215 YДерево» . yfull.com . Проверено 10 июля 2016 г.
- ^ Кручиани, Ф.; Ла Фратта, Р.; Сантоламазца, П.; Селитто, Д.; Пасконе, Р.; Мораль, П.; Уотсон, Э.; Гуида, В.; Коломб, Э.Б. (2004). «Филогеографический анализ гаплогруппы E3b (E-M215) Y-хромосом выявляет множественные миграционные события внутри Африки и из нее» . Американский журнал генетики человека . 74 (5): 1014–1022. дои : 10.1086/386294 . ПМК 1181964 . ПМИД 15042509 .
- ^ Jump up to: а б с д и ж Альварес, Л.; Сантос, К.; Монтьель, Р.; Каейро, Б.; Баали, А.; Дюгужон, Ж.-М.; Алуя, депутат (2009). «Вариации Y-хромосомы в Южной Иберии: взгляд на вклад Северной Африки» . Американский журнал биологии человека . 21 (3): 407–409. дои : 10.1002/ajhb.20888 . ПМИД 19213004 . S2CID 7041905 .
- ^ Кейта и Бойс (2005) .
- ^ Куянова, Мартина; Перейра, Луиза; Фернандес, Вероника; Перейра, Джоана Б.; Черны, Виктор (октябрь 2009 г.). «Генетический вклад ближневосточного неолита в небольшой оазис Западной египетской пустыни». Американский журнал физической антропологии . 140 (2): 336–346. дои : 10.1002/ajpa.21078 . ISSN 1096-8644 . ПМИД 19425100 .
- ^ ИЗОГГ (2008) .
- ^ Карафет, ТМ; Мендес, Флорида; Мейлерман, МБ; Андерхилл, Пенсильвания; Зегура, СЛ; Хаммер, МФ (2008). «Новые бинарные полиморфизмы изменяют форму и увеличивают разрешение дерева гаплогрупп Y-хромосомы человека» . Геномные исследования . 18 (5): 830–838. дои : 10.1101/гр.7172008 . ПМК 2336805 . ПМИД 18385274 .
- ^ Консорциум, TYC (2002 г.). «Система номенклатуры дерева бинарных гаплогрупп Y-хромосомы человека» . Геномные исследования . 12 (2): 339–348. дои : 10.1101/гр.217602 . ПМК 155271 . ПМИД 11827954 .
- ^ «Генетический атлас - история мутаций E1b1b Meditid» . thegeneticatlas.com . Проверено 1 октября 2015 г.
- ^ Член палаты представителей (август 2021 г.). «Взгляд на ближневосточный отцовский генетический фонд в Тунисе: высокая распространенность гаплогруппы Т-М70 в арабском населении» . Научные отчеты . 11 (1): 15728. Бибкод : 2021NatSR..1115728E . дои : 10.1038/s41598-021-95144-x . ПМЦ 8333252 . ПМИД 34344940 .
- ^ Жене, Am J Hum (май 2004 г.). «Происхождение, распространение и дифференциация гаплогрупп E и J Y-хромосомы: выводы о неолитизации Европы и более поздних миграционных событиях в Средиземноморье» . Американский журнал генетики человека . 74 (5): 1023–1034. дои : 10.1086/386295 . ПМК 1181965 . ПМИД 15069642 .
- ^ 31% основано на объединенных данных.
- Семино и др. 2004 : 30%
- Арреди и др. 2004 : 32%
- ^ Jump up to: а б Фадлауи-Зид и др. 2013, Общегеномное и отцовское разнообразие раскрывают недавнее происхождение человеческих популяций в Северной Африке.
- ^ Jump up to: а б Франкалаччи и др. 2008, История и география Y-хромосомы человека в Европе: взгляд на SNP.
- ^ Jump up to: а б с д Семино, О.; Магри, К.; Бенуцци, Г.; Лин, А.А.; Аль-Захери, Н.; Батталья, В.; Макчиони, Л.; Триантафиллидис, К.; Шен, П. (2004). «Происхождение, распространение и дифференциация гаплогрупп E и J Y-хромосомы: выводы о неолитизации Европы и более поздних миграционных событиях в Средиземноморье» . Американский журнал генетики человека . 74 (5): 1023–1034. дои : 10.1086/386295 . ПМК 1181965 . ПМИД 15069642 .
- ^ Фадлауи-Зид, К.; Хабер, М.; Мартинес-Крус, Б.; Заллуа, П.; Бенаммар Эльгаайед, А.; Комас, Д. (ноябрь 2013 г.). «Общегеномное и отцовское разнообразие раскрывает недавнее происхождение человеческих популяций в Северной Африке» . ПЛОС ОДИН . 8 (11): е80293. Бибкод : 2013PLoSO...880293F . дои : 10.1371/journal.pone.0080293 . ПМЦ 3842387 . ПМИД 24312208 .
- ^ Майлз, С.; Бузекри, Н.; Хаверфилд, Э.; Шеркауи, М.; Дюгужон, Ж.М.; Уорд, Р. (2005). «Генетические доказательства в поддержку общего происхождения молочных продуктов Евразии и Северной Африки». Генетика человека . 117 (1): 34–42. дои : 10.1007/s00439-005-1266-3 . ПМИД 15806398 . S2CID 23939065 .
- ^ Адамс, С.М.; Бош, Э.; Балареск, Польша; Баллеро, SPJ; Ли, AC; Арройо, Э.; Лопес-Парра, AM; Алер, М.; Грифо, MSG (2008). «Генетическое наследие религиозного разнообразия и нетерпимости: отцовские линии христиан, евреев и мусульман на Пиренейском полуострове» . Американский журнал генетики человека . 83 (6): 725–736. дои : 10.1016/j.ajhg.2008.11.007 . ПМК 2668061 . ПМИД 19061982 .
- ^ Заллуа, Пенсильвания; Платт, Делавэр; Эль Сибай, М.; Халифе, Дж.; Махул, Н.; Хабер, М.; Сюэ, Ю.; Изаабель, Х.; Бош, Э. (2008). «Выявление генетических следов исторической экспансии: финикийские следы в Средиземноморье» . Американский журнал генетики человека . 83 (5): 633–42. дои : 10.1016/j.ajhg.2008.10.012 . ПМК 2668035 . ПМИД 18976729 .
- ^ Бехар, DM; Юнусбаев Б.; Метспалу, М.; Метспалу, Э.; Россет, С.; Парик, младший; Роотси, С.; Чаубей, Г.; Кутуев, И. (2010). «Полногеномная структура еврейского народа». Природа . 466 (7303): 238–242. Бибкод : 2010Natur.466..238B . дои : 10.1038/nature09103 . ПМИД 20531471 . S2CID 4307824 .
- ^ Алонсо, С.; Флорес, К.; Кабрера, В.; Алонсо, А.; Мартин, П.; Альбарран, К.; Исагирре, Н.; Де Ла Руа, Китай; Гарсиа, О. (2005). «Место басков в европейском ландшафте разнообразия Y-хромосомы» . Европейский журнал генетики человека . 13 (12): 1293–1302. дои : 10.1038/sj.ejhg.5201482 . ПМИД 16094307 .
- ^ Jump up to: а б Кручиани, Ф.; Сантоламазца, П.; Шен, П.; Маколей, В.; Мораль, П.; Олкерс, А.; Модиано, Д.; Холмс, С.; Дестро-Бисол, Г. (2002). «Обратная миграция из Азии в страны Африки к югу от Сахары подтверждается анализом гаплотипов Y-хромосомы человека с высоким разрешением» . Американский журнал генетики человека . 70 (5): 1197–1214. дои : 10.1086/340257 . ПМЦ 447595 . ПМИД 11910562 .
- ^ Jump up to: а б Бекада, Асмахан; Фрегель, Роза; Кабрера, Висенте М.; Ларруга, Хосе М.; Пестано, Хосе; Бенхамамуш, Сорая; Гонсалес, Ана М. (19 февраля 2013 г.). «Внедрение алжирской митохондриальной ДНК и профилей Y-хромосомы в ландшафт Северной Африки» . ПЛОС ОДИН . 8 (2): e56775. Бибкод : 2013PLoSO...856775B . дои : 10.1371/journal.pone.0056775 . ISSN 1932-6203 . ПМЦ 3576335 . ПМИД 23431392 .
- ^ Jump up to: а б Фрегель и др. 2009, Демографическая история мужского генофонда Канарских островов: замена местных линий европейскими.
- ^ Онофри и др. 2008, Распределение маркеров Y-хромосомы в Северной Африке: анализ SNP и STR высокого разрешения в популяциях Туниса и Марокко.
- ^ Jump up to: а б с д и Бош и др. 2001. Анализ вариаций Y-хромосомы человека с высоким разрешением показывает резкий разрыв и ограниченный поток генов между Северо-Западной Африкой и Пиренейским полуостровом.
- ^ Кручиани и др. 2004, Филогеографический анализ Y-хромосом гаплогруппы E3b (E-M215) выявляет множественные миграционные события внутри и из Африки.
- ^ Jump up to: а б Кручиани, Ф.; Ла Фратта, Р.; Сантоламазца, П.; Селитто, Д.; Пасконе, Р.; Мораль, П.; Уотсон, Э.; Гуида, В.; Коломб, Э.Б. (2004). «Филогеографический анализ гаплогруппы E3b (E-M215) Y-хромосом выявляет множественные миграционные события внутри Африки и из нее» . Американский журнал генетики человека . 74 (5): 1014–1022. дои : 10.1086/386294 . ПМК 1181964 . ПМИД 15042509 .
- ^ Jump up to: а б «Берберы: лингвистическое и генетическое разнообразие» (PDF) . ddl.ish-lyon.cnrs.fr. Архивировано из оригинала (PDF) 25 марта 2012 года . Проверено 22 марта 2015 г.
- ^ Jump up to: а б с Ахмед, Региг; Нурдин, Харич; Абдельхамид, Баракат; Хасан, Руба (2014). «Филогеография гаплогруппы E1b1b1b-M81 и анализ ее субкладов в Марокко» . Препринты по биологии человека в открытом доступе . 86 (2).
- ^ Y Гаплогруппы мира, онлайн-издание. Архивировано 11 декабря 2013 г. в Wayback Machine.
- ^ Абдалла Ларуи, История Магриба (Париж, 1970; Принстон, 1977) на 17, 60 (о свасианцах, ссылаясь на более раннюю работу Гселля).
- ^ Кэмпс, Габриэль (1996), Берберы , Эдисуд, стр. 11–14.65
- ^ Ахилли, А.; Ренго, К.; Батталья, В.; Пала, М.; Оливьери, А.; Форнарино, С.; Магри, К.; Скоццари, Р.; Бабудри, Н. (2005). «Саамы и берберы — неожиданная связь митохондриальной ДНК» . Американский журнал генетики человека . 76 (5): 883–886. дои : 10.1086/430073 . ПМК 1199377 . ПМИД 15791543 .
- ^ Ж. Десанж, «Протоберберы» 236–245, стр. 237, в «Всеобщей истории Африки», т. II «Древние цивилизации Африки» (ЮНЕСКО, 1990).
- ^ Марио Кертис Джордани, История Африки. До открытий (Петрополис, Бразилия: Editora Vozes 1985) на 42–43, 77–78. Джордани ссылается на Буске, «Берберы» (Париж, 1961).
- ^ Jump up to: а б Малярчук Б.А.; Деренко, М.; Перкова, М.; Гжибовский, Т.; Ванечек, Т.; Лазур, Дж. (2008). «Реконструкция филогении африканских линий митохондриальной ДНК у славян» . Европейский журнал генетики человека . 16 (9): 1091–1096. дои : 10.1038/ejhg.2008.70 . ПМИД 18398433 .
- ^ Черни, Л.; Фернандес, В.Н.; Перейра, Дж.Б.; Коста, доктор медицины; Гойос, А.; Фриги, С.; Якуби-Луэслати, Б.; Амор, МБ; Слама, А. (2009). «Максимальное расширение после последнего ледникового периода от Иберии до Северной Африки, выявленное путем точной характеристики гаплогруппы H мтДНК в Тунисе». Американский журнал физической антропологии . 139 (2): 253–260. дои : 10.1002/ajpa.20979 . ПМИД 19090581 .
- ^ Брейс, CL (2006). «Сомнительный вклад неолита и бронзового века в европейскую черепно-лицевую форму» . Труды Национальной академии наук . 103 (1): 242–247. Бибкод : 2006PNAS..103..242B . дои : 10.1073/pnas.0509801102 . ПМК 1325007 . ПМИД 16371462 .
- ^ Кавалли-Сфорца, Л.; Меноцци, П.; Пьяцца, А. (1993). «Демическая экспансия и эволюция человека». Наука . 259 (5095): 639–646. Бибкод : 1993Sci...259..639C . дои : 10.1126/science.8430313 . ПМИД 8430313 . S2CID 27056263 .
- ^ Бар-Йосеф О (1987) Плейстоценовые связи между Африкой и Юго-Западной Азией: археологическая перспектива. Африканский археологический обзор ; Глава 5, стр. 29–38.
- ^ Андерхилл, Пенсильвания; Кивисилд, Т. (2007). «Использование структуры популяции Y-хромосомы и митохондриальной ДНК для отслеживания миграций человека». Ежегодный обзор генетики . 41 : 539–564. дои : 10.1146/annurev.genet.41.110306.130407 . ПМИД 18076332 . S2CID 24904955 .
- ^ Делсон, Эрик; Таттерсолл, Ян; Куверинг, Джон Ван; Брукс, Элисон С. (23 ноября 2004 г.). Энциклопедия эволюции и предыстории человека: второе издание . Рутледж. п. 97. ИСБН 9781135582272 .
- ^ Кефи Р., Бузаид Э., Стеванович А., Беро-Коломб Э. «МИТОХОНДРИАЛЬНАЯ ДНК И ФИЛОГЕНЕТИЧЕСКИЙ АНАЛИЗ ДОИСТОРИЧЕСКИХ ПОПУЛЯЦИЙ СЕВЕРНОЙ АФРИКИ» (PDF) . ИСАБС. Архивировано из оригинала (PDF) 11 марта 2016 года . Проверено 17 января 2016 г.
- ^ Бернар Сешер; Роза Фрегель; Хосе М. Ларруга; Висенте М. Кабрера; Филипп Эндикотт; Хосе Дж. Пестано; Ана М. Гонсалес (2014). «История потока гена гаплогруппы U6 североафриканской митохондриальной ДНК на африканский, евразийский и американский континенты» . Эволюционная биология BMC . 14 :109. дои : 10.1186/1471-2148-14-109 . ПМК 4062890 . ПМИД 24885141 .
- ^ Лоосдрехт, Марике ван де; Бузуггар, Абдельджалил; Хамфри, Луиза; Пост, Козимо; Бартон, Ник; Аксиму-Петри, Айинуэр; Никель, Биргит; Нагель, Сара; Талби, Эль Хасан (15 марта 2018 г.). «Плейстоценовые геномы Северной Африки связывают человеческие популяции Ближнего Востока и Африки к югу от Сахары» . Наука . 360 (6388): 548–552. Бибкод : 2018Sci...360..548V . дои : 10.1126/science.aar8380 . ISSN 0036-8075 . ПМИД 29545507 .
- ^ Фадлауи-Зид, К.; Плаза, С.; Калафель, Ф.; Бен Амор, М.; Комас, Д.; Беннамар, А.; Гаайед, Э. (2004). «Гетерогенность митохондриальной ДНК у тунисских берберов». Анналы генетики человека . 68 (3): 222–233. дои : 10.1046/j.1529-8817.2004.00096.x . ПМИД 15180702 . S2CID 6407058 .
- ^ Кудрэ, К.; Оливьери, А.; Ахилли, А.; Пала, М.; Мелауи, М.; Шеркауи, М.; Эль-Ченнави, Ф.; Коссманн, М.; Торрони, А.; Дюгужон, ЖМ (2009). «Сложный и разнообразный митохондриальный генофонд берберских популяций». Анналы генетики человека . 73 (2): 196–214. дои : 10.1111/j.1469-1809.2008.00493.x . ПМИД 19053990 . S2CID 21826485 .
- ^ Черни, Л.; Фернандес, В.Н.; Перейра, Дж.Б.; Коста, доктор медицины; Гойос, А.; Фриги, С.; Якуби-Луэслати, Б.; Амор, МБ; Слама, А. (2009). «Максимальное расширение после последнего ледникового периода от Иберии до Северной Африки, выявленное путем точной характеристики гаплогруппы H мтДНК в Тунисе». Американский журнал физической антропологии . 139 (2): 253–260. дои : 10.1002/ajpa.20979 . ПМИД 19090581 .
- ^ Харич, Н.; Коста, доктор медицины; Фернандес, В.Н.; Кандил, М.; Перейра, Дж.Б.; Сильва, Нью-Мексико; Перейра, Л.С. (2010). «Транссахарская работорговля – подсказки из интерполяционного анализа и характеристики линий митохондриальной ДНК с высоким разрешением» . Эволюционная биология BMC . 10 :138. дои : 10.1186/1471-2148-10-138 . ПМЦ 2875235 . ПМИД 20459715 .
- ^ Фриги, С.; Черни, Л.; Фадлауи-Зид, К.; Бенаммар-Эльгаайед, А. (август 2010 г.). «Древняя локальная эволюция африканских гаплогрупп мтДНК в тунисских берберских популяциях » человека Биология 82 (4): 367–384. дои : 10.3378/027.082.0402 . ПМИД 21082907 . S2CID 27594333 .
- ^ Эстебан, Э.; Гонсалес-Перес, Э.; Харич, Н.; Лопес-Аломар, А.; Виа, М.; Луна, Ф.; Мораль, П. (2004). «Генетические связи среди берберов и южных испанцев на основе микросателлитов CD4/гаплотипов Alu». Анналы биологии человека . 31 (2): 202–212. дои : 10.1080/03014460310001652275 . ПМИД 15204363 . S2CID 24805101 .
- ^ Харич и др. 2010 г.
Цитируемые работы
[ редактировать ]- Батталья, Винченца; Форнарино, Симона; Аль-Захери, Надя; Оливьери, Анна; Лопата, Мэри; Майрес, Натали М; Кинг, Рой Дж; Корни, Сирия; и др. (2008), «Y-хромосомные свидетельства культурного распространения сельского хозяйства в юго-восточной Европе», European Journal of Human Genetics , 17 (6): 820–830, : 10.1038 /ejhg.2008.249 , PMC 2947100 , PMID doi
- Босх, Елена; Калафель, Франческ; Комас, Дэвид; Офнер, Питер Дж.; Андерхилл, Питер А.; Бертранпети, Жауме (2001), «Анализ вариаций Y-хромосомы человека с высоким разрешением показывает резкий разрыв и ограниченный поток генов между северо-западной Африкой и Пиренейским полуостровом», Am J Hum Genet , 68 (4): 1019–1029 , doi : 10.1086/319521 , PMC 1275654 , PMID 11254456
- Капелли С., Онофри В., Бризигелли Ф., Боски И., Скарниччи Ф., Масулло М. и др. (июнь 2009 г.). «Мавры и сарацины в Европе: оценка средневекового мужского наследия Северной Африки в южной Европе» . Европейский журнал генетики человека . 17 (6): 848–52. дои : 10.1038/ejhg.2008.258 . ПМЦ 2947089 . ПМИД 19156170 .
- Кручиани, Ф.; Ла Фратта, Р.; Тромбетта, Б.; Сантоламазца, П.; Селитто, Д.; Коломб, Э.Б.; Дюгужон, Ж.-М.; Кривелларо, Ф.; и др. (2007), «Отслеживание прошлых человеческих перемещений мужчин в Северной / Восточной Африке и Западной Евразии: новые подсказки по гаплогруппам Y-хромосомы E-M78 и J-M12» , Molecular Biology and Evolution , 24 (6): 1300–1311, doi : 10.1093/molbev/msm049 , PMID 17351267 , заархивировано из оригинала 10 октября 2017 г. Также см. Дополнительные данные .
- Хасан, Хишам Ю.; Андерхилл, Питер А.; Кавалли-Сфорца, Лука Л.; Ибрагим, Мунтасер Э. (2008), «Вариации Y-хромосомы среди суданцев: ограниченный поток генов, соответствие языку, географии и истории» (PDF) , Американский журнал физической антропологии , 137 (3): 316–23, doi : 10.1002/ajpa.20876 , PMID 18618658 , заархивировано из оригинала (PDF) 4 марта 2009 г.
- Хенн, Б.М.; Жиньу, К.; Лин, Алиса А; Офнер, Питер Дж.; Шен, П.; Скоццари, Р.; Кручиани, Ф.; Тишкофф, С.А.; Маунтин, JL; Андерхилл, Пенсильвания (2008), «Y-хромосомные свидетельства миграции скотоводов через Танзанию в южную Африку», PNAS , 105 (31): 10693–8, Bibcode : 2008PNAS..10510693H , doi : 10.1073/pnas.0801184105 , PMC 2504844 , ПМИД 18678889 . См. комментарии в блоге Dienekes , комментарии в блоге Spitoon и публичный выпуск .
- ИЗОГГ (2008). «Дерево гаплогрупп Y-ДНК 2008» .
- Карафет Т.М., Мендес Ф.Л., Мейлерман М.Б., Андерхилл П.А., Зегура С.Л., Хаммер М.Ф. (2008). «Новые бинарные полиморфизмы изменяют форму и увеличивают разрешение дерева гаплогрупп Y-хромосомы человека» . Геномные исследования . 18 (5): 830–38. дои : 10.1101/гр.7172008 . ПМК 2336805 . ПМИД 18385274 .
- Кейта, СОИ; Бойс, AJ (Энтони Дж.) (2005), «Генетика, Египет и история: интерпретация географических закономерностей вариаций Y-хромосомы» , History in Africa , 32 : 221–246, doi : 10.1353/hia.2005.0013 , S2CID 163020672
- Луис, Дж; Роуолд, Д; Регейро, М; Каейро, Б; Чинниоглу, К; Роузман, К; Андерхилл, П; Каваллисфорса, Л; Эррера, Р. (2004), «Левант против Африканского Рога: доказательства двунаправленных коридоров миграции людей» (PDF) , Американский журнал генетики человека , 74 (3): 532–544, doi : 10.1086/382286 , PMC 1182266 , PMID 14973781 , заархивировано из оригинала (PDF) 16 февраля 2012 г. (Также см. Ошибки )
- Андерхилл, Питер А.; Шен, Пейдун; Лин, Алиса А.; Джин, Ли; и др. (ноябрь 2000 г.). «Вариации последовательности Y-хромосомы и история человеческих популяций». Природная генетика . 26 (3): 358–361. дои : 10.1038/81685 . ПМИД 11062480 . S2CID 12893406 .
| Подтемы | |
|---|---|
| Генетическая история по регионам | |
| Популяционная генетика по группе |
|