Гаплогруппа E-M35
| Гаплогруппа E-M35 | |
|---|---|
| Возможное время происхождения | 34 800 лет назад [1] |
| Возраст слияния | 23 900 лет назад [1] |
| Возможное место происхождения | Восточная Африка [2] |
| Предок | Э-М215 |
| Потомки | Э-В68 , Э-З827 |
| Определение мутаций | М35 |
E-M35 , также известная как E1b1b1-M35 , представляет собой гаплогруппу ДНК Y-хромосомы человека . E-M35 имеет две базальные ветви: E-V68 и E-Z827 . E-V68 и E-Z827 в основном распространены в Северной Африке и на Африканском Роге , реже встречаются на Ближнем Востоке , в Европе и Южной Африке .
Происхождение
[ редактировать ]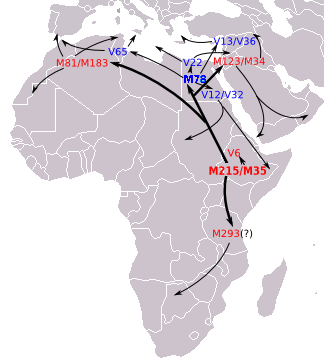
В июне 2015 г. Тромбетта и др. сообщил о ранее не оцененной большой разнице в возрасте между гаплогруппой E-M215 (38,6 тыс. лет назад; 95% ДИ 31,4–45,9 тыс лет назад) и ее подгаплогруппой E-M35 (25,0 тыс. лет назад; 95% ДИ 20,0–30,0 тыс лет назад) и оценил ее происхождение. находиться на Африканском Роге , где узел, разделяющий ветви E-V38 и E-M215, возник около 47 500 лет назад (95% ДИ: 41,3–56,8 тыс. лет назад). [7] E-M35 был датирован Батини в 2015 году периодом от 15 400 до 20 500 лет назад. [8]
Считается, что все основные ответвления E-M35 возникли в той же области, что и родительская клада: в Северной Африке , на Африканском Роге или в близлежащих районах Ближнего Востока . Предполагается, что некоторые ветви E-M35 покинули Африку тысячи лет назад, тогда как другие, возможно, прибыли с Ближнего Востока. Например, Андерхилл (2002) связывает распространение гаплогруппы с неолитической революцией , полагая, что структура и региональная структура субкладов E-M35 потенциально дают «реагенты, с помощью которых можно сделать выводы о конкретных эпизодах истории населения, связанных с неолитической сельскохозяйственной экспансией». . Батталья и др. (2008) также подсчитали, что E-M78 (названный в этой статье E1b1b1a1) существует в Европе уже более 10 000 лет. Соответственно, было показано, что человеческие останки, раскопанные в испанской погребальной пещере, датируемые примерно 7000 лет назад, принадлежат к этой гаплогруппе. [9] Еще два E-M78 были найдены в неолитических культурах Сопота и Лендьеля . [10]
Что касается E-M35 в Европе в рамках этой схемы, Underhill & Kivisild (2007) отметили, что E-M215, по-видимому, представляет собой миграцию позднего плейстоцена из Северной Африки в Европу через Синайский полуостров в Египте . [Примечание 1] Хотя это предложение остается неоспоримым, совсем недавно оно было предложено Тромбеттой и др. (2011) также имеются доказательства дополнительной миграции E-M215, переносящей мужчин напрямую из Северной Африки в юго-западную Европу морским путем (см. ниже).
Древняя ДНК
[ редактировать ]По данным Лазаридиса и др. (2016), натуфийские останки скелетов из древнего Леванта преимущественно содержали гаплогруппу Y-ДНК E1b1b. Из пяти натуфийских образцов, проанализированных на предмет отцовских линий, три принадлежали к субкладам E1b1b1b2(xE1b1b1b2a,E1b1b1b2b), E1b1(xE1b1a1,E1b1b1b1) и E1b1b1b2(xE1b1b1b2a,E1b1b1b2b) (60%). Гаплогруппа E1b1b также обнаруживалась с умеренной частотой среди окаменелостей последующей докерамической неолитической культуры B, при этом субклады E1b1b1 и E1b1b1b2 (xE1b1b1b2a, E1b1b1b2b) наблюдались в двух из семи образцов PPNB (~ 29%). Ученые предполагают, что ранние земледельцы Леванта, возможно, распространились на юг, в Восточную Африку, принося с собой западно-евразийские и базально-евразийские предковые компоненты, отдельные от тех, которые позже прибыли в Северную Африку.
Кроме того, гаплогруппа E1b1b1 была обнаружена в древней египетской мумии, раскопанной на археологическом памятнике Абусир-эль-Мелек в Среднем Египте, который датируется периодом между поздним Новым царством и римской эрой . [11] Окаменелости на иберомаврском стоянке Ифри-н'Амр или Мусса в Марокко , датированные примерно 5000 годом до нашей эры, также несли гаплотипы, относящиеся к субкладу E1b1b1b1a (E-M81). Эти древние люди несли автохтонный геномный компонент Магриба, который достигает максимума среди современных жителей Северной Африки , что указывает на то, что они были предками популяций в этом районе. [12] Гаплогруппа E1b1b также наблюдалась в древних окаменелостях гуанчей , раскопанных на Гран-Канарии и Тенерифе на Канарских островах , которые были датированы радиоуглеродом между 7 и 11 веками нашей эры. Особи-носители клады, которые были проанализированы на предмет отцовской ДНК, были захоронены на территории Тенерифе, при этом все эти экземпляры оказались принадлежащими к субкладу E1b1b1b1a1 или E-M183 (3/3; 100%). [13]
Лоосдрехт и др. (2018) проанализировали полногеномные данные семи древних иберомавров из Голубиной пещеры недалеко от Тафоральта на востоке Марокко. Окаменелости были напрямую датированы периодом от 15 100 до 13 900 лет назад. Ученые обнаружили, что пять экземпляров мужского пола с достаточной сохранностью ядерной ДНК принадлежали к субкладу E1b1b1a1 (M78), при этом один скелет нес родительскую линию E1b1b1a1b1 к E-V13, другой экземпляр мужского пола принадлежал к E1b1b (M215*). [14]
Распределение
[ редактировать ]E-M215 и E-M35 довольно распространены среди говорящих на афроазиатском языке . Лингвистическая группа и носители линии Е-М35 с высокой вероятностью возникли и рассеялись вместе из афроазиатской урхеймата . [15] Среди населения с афро-азиатскоязычным прошлым значительная часть палестинцев и евреев мужского пола принадлежит к E-M35. [16] Гаплогруппа E-M35, составляющая примерно 18% [2] до 20% [17] [18] 18% палестинцев и ашкенази и 8,6% [19] до 30% [2] сефардской Y-хромосомы, по-видимому , является одной из основных линий-основателей палестинского и еврейского населения. [20] [Примечание 2]
В следующую таблицу включены только выборочные популяции, в которых более 1% мужчин E-M215 со всеми известными субкладами по состоянию на июнь 2015 года. Она содержит кладу E-V1515, определенную Trombetta et al. 2015, и все субклады E1b1b, распространенные ниже Сахары (E-V42, E-M293, E-V92, E-V6), которые в прежней филогении были идентифицированы как базальные клады E-M35. [7]
| Население | Н | Область | Язык | Общий Э-М215 | E-V2009 | Э-М78* | E-V1477 | E-V1083* | Э-В13 | Э-В22 | Э-В12* | Э-В32 | Э-В259 | Э-В65 | Э-В257* | Э-М81 | Э-М123* | Э-М34 | E-V1515* | E-V1486* | E-V2881* | E-V1792 | Э-В92 | Э-М293* | E-V3065 | Э-В42 | E-V1785* | Э-В6 | Э-В16 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Северная Африка | |||||||||||||||||||||||||||||
| Марокканские арабы | 55 | Марокко | АА/семитский | 15.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 7.3 | 0.0 | 0.0 | 0.0 | 32.7 | 0.0 | 30.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Асни Берберы | 54 | Марокко | АА/берберский | 85.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.7 | 0.0 | 0.0 | 0.0 | 0.0 | 1.9 | 79.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Бухрия Берберы | 67 | Марокко | АА/берберский | 79.1 | 0.0 | 0.0 | 0.0 | 0.0 | 1.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 77.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Берберы Среднего Атласа | 69 | Марокко | АА/берберский | 81.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 10.1 | 0.0 | 71.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Марракешские берберы | 27 | Марокко | АА/берберский | 92.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.7 | 3.7 | 0.0 | 0.0 | 0.0 | 3.7 | 77.8 | 0.0 | 3.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Сусс Берберы | 34 | Марокко | АА/берберский | 79.4 | 2.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 76.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Уарзазат Берберы | 31 | Марокко | АА/берберский | 54.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 54.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Мозабитские берберы | 67 | Алжир | АА/берберский | 89.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 1.5 | 0.0 | 86.6 | 0.0 | 1.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Тунисские евреи | 10 | Тунис | АА/семитский | 20.0 | 0.0 | 0.0 | 10.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 10.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Ливийские арабы | 10 | Ливия | АА/семитский | 20.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 20.0 | 0.0 | 30.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Ливийские евреи | 23 | Ливия | АА/семитский | 26.1 | 0.0 | 0.0 | 0.0 | 0.0 | 4.3 | 0.0 | 0.0 | 0.0 | 0.0 | 4.3 | 0.0 | 0.0 | 0.0 | 17.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Северные египтяне | 49 | Египет | АА/семитский | 20.9 | 0.0 | 0.0 | 0.0 | 0.0 | 2.0 | 16.3 | 4.1 | 2.0 | 0.0 | 0.0 | 0.0 | 4.1 | 4.1 | 10.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Египетские берберы из Сивы | 93 | Египет | АА/семитский | 18.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.2 | 0.0 | 0.0 | 4.3 | 2.2 | 1.1 | 0.0 | 2.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 6.5 | 0.0 |
| Египтяне из Бахарии | 41 | Египет | АА/семитский | 56.1 | 0.0 | 0.0 | 0.0 | 0.0 | 2.4 | 22.0 | 14.6 | 0.0 | 0.0 | 2.4 | 7.3 | 4.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.4 | 0.0 |
| Египтяне из оазиса Гурна | 34 | Египет | АА/семитский | 17.6 | 0.0 | 5.9 | 0.0 | 0.0 | 0.0 | 0.0 | 8.8 | 2.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Южные египтяне | 47 | Египет | АА/семитский | 78.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 74.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.1 | 2.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Западная/Центральная Африка | |||||||||||||||||||||||||||||
| Манденка | 16 | Сенегал | Северная Каролина/Манде | 6.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 6.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Туареги | 22 | Нигер | АА/берберский | 13.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4.5 | 0.0 | 0.0 | 0.0 | 0.0 | 9.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Назад | 29 | Камерун (Север) | АА/Чадик | 3.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Путеводители | 9 | Камерун (Север) | АА/Чадик | 11.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 11.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Мандара | 82 | Камерун (Север) | АА/Чадик | 2.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 1.2 | 0.0 | 0.0 | 0.0 | 1.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Шува арабы | 5 | Камерун (Север) | АА/семитский | 20.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 20.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Фульбе из Камеруна | 76 | Камерун (Север) | Северная Каролина/Атлантика | 1.3 | 1.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| ОСТАНАВЛИВАТЬСЯ | 21 | Камерун (Север) | Северная Каролина/Адамава | 4.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Восточная Африка | |||||||||||||||||||||||||||||
| Тигре | 5 | Эритрея | АА/семитский | 100.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 60.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 20.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 20.0 | 0.0 |
| Нара | 15 | Эритрея | НС/Суданский | 60.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 6.7 | 0.0 | 13.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 13.3 | 6.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 13.3 | 0.0 | 6.7 | 0.0 |
| Кунаама | 20 | Эритрея | Н.С./Кунама | 65.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 5.0 | 0.0 | 20.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 5.0 | 0.0 | 0.0 | 5.0 | 0.0 | 0.0 | 0.0 | 0.0 | 15.0 | 10.0 | 5.0 | 0.0 |
| Сахо | 94 | Эритрея | АА/кушитский | 98.9 | 0.0 | 0.0 | 0.0 | 1.1 | 0.0 | 88.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 1.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 8.5 | 0.0 |
| Тигры | 32 | Эритрея/Эфиопия | АА/семитский | 71.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.1 | 3.1 | 21.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.1 | 3.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 31.3 | 6.3 | 0.0 |
| издалека | 25 | Джибути | АА/кушитский | 60.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 56.0 | 0.0 |
| Сомали | 40 | Джибути | АА/кушитский | 25.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 25.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Эфиопские евреи | 22 | Эфиопия | АА/кушитский | 31.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 9.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 13.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 9.1 | 0.0 | 0.0 | 0.0 |
| Амхара | 82 | Эфиопия | АА/семитский | 45.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.4 | 0.0 | 11.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 13.4 | 0.0 | 0.0 | 2.4 | 0.0 | 1.2 | 0.0 | 0.0 | 1.2 | 0.0 | 8.5 | 4.9 |
| Оромо | 62 | Эфиопия | АА/кушитский | 53.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 22.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4.8 | 0.0 | 0.0 | 17.7 | 0.0 | 0.0 | 1.6 | 0.0 | 3.2 | 0.0 | 1.6 | 1.6 |
| Волайта | 12 | Эфиопия | АА/Головокружение | 58.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 8.3 | 0.0 | 8.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 8.3 | 0.0 | 0.0 | 8.3 | 0.0 | 0.0 | 8.3 | 0.0 | 0.0 | 0.0 | 16.7 | 0.0 |
| Сомали | 12 | Эфиопия | АА/кушитский | 50.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 8.3 | 0.0 | 25.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 16.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Гураге | 7 | Эфиопия | АА/семитский | 42.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 28.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 14.3 | 0.0 |
| Сомали | 5 | Сомали | АА/кушитский | 100.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 80.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 20.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Туркана | 6 | Кения | НС/Суданский | 50.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 33.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 16.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Борана | 9 | Кения | АА/кушитский | 77.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 66.7 | 0.0 | 0.0 | 11.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Сомали | 6 | Кения | АА/кушитский | 100.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 66.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 16.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 16.7 | 0.0 |
| Нилотик Западная Кения | 11 | Кения | НС/Суданский | 45.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 9.1 | 9.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 18.2 | 0.0 | 0.0 | 0.0 | 9.1 | 0.0 |
| Лухья | 51 | Кения | Северная Каролина/банту | 9.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 5.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Другие банту | 17 | Кения | Северная Каролина/банту | 11.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 11.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Кикую | 9 | Кения | Северная Каролина/банту | 11.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 11.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Масаи | 45 | Кения | НС/Суданский | 37.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 6.7 | 0.0 | 6.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 24.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| тутси | 9 | Бурунди | Северная Каролина/банту | 22.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 22.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Южная Африка | |||||||||||||||||||||||||||||
| !Если | 64 | Ангола | КС | 10.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 9.4 | 1.6 | 0.0 | 0.0 | 0.0 | 0.0 |
| Хве | 26 | Намибия | КС | 30.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 30.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Помощь | 8 | ЮАР | Северная Каролина/банту | 12.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 12.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Европа | |||||||||||||||||||||||||||||
| Северный португальский | 50 | Португалия | ИЕ | 10.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4.0 | 0.0 | 0.0 | 2.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Южный португальский | 49 | Португалия | ИЕ | 16.3 | 0.0 | 0.0 | 0.0 | 0.0 | 4.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 12.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Пасиегос из Кантабрии | 56 | Испания | ИЕ | 42.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 1.8 | 41.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Астурийцы | 90 | Испания | ИЕ | 12.2 | 0.0 | 0.0 | 0.0 | 0.0 | 5.6 | 4.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Южные испанцы | 62 | Испания | ИЕ | 6.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.2 | 0.0 | 0.0 | 0.0 | 0.0 | 1.6 | 1.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Испанские баски | 55 | Испания | Баскский | 3.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Французский | 85 | Франция | ИЕ | 8.2 | 0.0 | 0.0 | 0.0 | 0.0 | 3.5 | 0.0 | 1.2 | 0.0 | 0.0 | 0.0 | 0.0 | 3.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Французские баски | 16 | Франция | Баскский | 6.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 6.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| корсиканцы | 140 | Франция | ИЕ | 6.4 | 0.0 | 0.0 | 0.0 | 0.0 | 4.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.7 | 0.0 | 0.0 | 1.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| датский | 35 | Дания | ИЕ | 2.9 | 0.0 | 0.0 | 0.0 | 0.0 | 2.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| немцы | 77 | Германия | ИЕ | 3.9 | 0.0 | 0.0 | 0.0 | 0.0 | 3.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Северные итальянцы | 80 | Италия | ИЕ | 11.3 | 0.0 | 0.0 | 0.0 | 0.0 | 6.3 | 2.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 1.3 | 0.0 | 1.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Центральные итальянцы | 356 | Италия | ИЕ | 12.9 | 0.0 | 0.0 | 0.0 | 0.0 | 5.3 | 2.0 | 0.3 | 0.0 | 0.0 | 0.3 | 0.3 | 0.8 | 0.0 | 3.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Южные итальянцы | 141 | Италия | ИЕ | 15.6 | 0.7 | 0.0 | 0.0 | 0.0 | 8.5 | 1.4 | 0.7 | 0.0 | 0.0 | 0.0 | 0.0 | 1.4 | 0.0 | 2.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| сицилийцы | 153 | Италия | ИЕ | 20.3 | 0.0 | 0.0 | 0.0 | 0.0 | 7.2 | 4.6 | 0.7 | 0.0 | 0.0 | 0.7 | 0.0 | 0.7 | 0.0 | 6.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| сардинцы | 374 | Италия | ИЕ | 8.3 | 0.8 | 0.0 | 0.0 | 0.3 | 1.1 | 0.8 | 0.3 | 0.0 | 0.0 | 1.1 | 0.3 | 0.3 | 0.0 | 3.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Польский | 40 | Польша | ИЕ | 2.5 | 0.0 | 0.0 | 0.0 | 0.0 | 2.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Словенцы | 104 | Словения | ИЕ | 2.9 | 0.0 | 0.0 | 0.0 | 0.0 | 2.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Эстонцы | 74 | Эстония | В | 5.4 | 0.0 | 0.0 | 0.0 | 0.0 | 4.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| венгры | 106 | Венгрия | В | 10.4 | 0.0 | 0.0 | 0.0 | 0.0 | 9.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| румыны | 30 | Румыния | ИЕ | 26.7 | 0.0 | 0.0 | 0.0 | 0.0 | 26.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| македонцы | 99 | Македония | ИЕ | 18.2 | 0.0 | 0.0 | 0.0 | 0.0 | 18.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Континентальные греки | 32 | Греция | ИЕ | 28.1 | 0.0 | 0.0 | 0.0 | 0.0 | 25.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| болгары | 112 | Болгария | ИЕ | 22.3 | 0.0 | 0.0 | 0.0 | 0.0 | 21.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Сефардские болгары | 20 | Болгария | ИЕ | 5.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 5.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| албанцы | 21 | Албания | ИЕ | 33.3 | 0.0 | 0.0 | 0.0 | 0.0 | 33.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Ближний Восток | |||||||||||||||||||||||||||||
| Сефардский турецкий | 19 | Турция | А | 10.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 5.3 | 0.0 | 5.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Стамбул Турецкий | 35 | Турция | А | 17.1 | 0.0 | 0.0 | 0.0 | 0.0 | 2.9 | 5.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 5.7 | 0.0 | 2.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Юго-западный турецкий | 40 | Турция | А | 7.5 | 0.0 | 0.0 | 0.0 | 0.0 | 2.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.5 | 0.0 | 2.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Северо-восточный турецкий | 41 | Турция | А | 2.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Центральная Анатолия | 61 | Турция | А | 9.8 | 0.0 | 0.0 | 0.0 | 0.0 | 4.9 | 0.0 | 1.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Юго-восточный турецкий | 24 | Турция | А | 8.3 | 0.0 | 0.0 | 0.0 | 0.0 | 4.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Эрзурум Турецкий | 25 | Турция | А | 12.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 8.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Турецкие киприоты | 46 | Турция | А | 23.9 | 0.0 | 0.0 | 0.0 | 0.0 | 10.9 | 2.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 8.7 | 0.0 | 2.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Бедуины | 28 | Израиль | АА/семитский | 14.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.6 | 0.0 | 7.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Друзы арабы | 28 | Израиль | АА/семитский | 14.3 | 0.0 | 0.0 | 0.0 | 0.0 | 10.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| палестинцы | 29 | Израиль | АА/семитский | 13.8 | 0.0 | 0.0 | 0.0 | 0.0 | 3.4 | 6.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Эмираты | 41 | Объединенные Арабские Эмираты | АА/семитский | 7.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Оманцы | 13 | Мой собственный | АА/семитский | 15.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 7.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 7.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| йеменцы | 94 | Йемен | АА/семитский | 14.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.1 | 3.2 | 0.0 | 0.0 | 0.0 | 1.1 | 0.0 | 7.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 1.1 |
Исключительные случаи мужчин, у которых M215-положительный, но M35-отрицательный («E-M215*»), были обнаружены на данный момент в двух амхарах Эфиопии и одном йемене . [3] [21] По крайней мере, некоторые из этих людей, а возможно, и все, с начала 2011 года известны как принадлежащие к редкой родственной кладе E-M35, известной как E-V16 или E-M281. [22] Об открытии M281 объявили Семино и др. (2002) , которые нашли его у двух эфиопских оромо . Тромбетта и др. (2011) обнаружили еще 5 особей из Эфиопии и SNP, эквивалентный M281, V16. Именно в статье 2011 года была обнаружена позиция генеалогического древа (M215+/M35-), как описано выше.Производная E-M215, E-M35, определяется SNP M35. 1 туркменский индивидуум из Джаузджана с мутацией, определяющей субклад, обозначен как E-M35*. [23] По состоянию на июнь 2015 года существует все более сложная древовидная структура, которая делит большинство мужчин в E-M35 на две ветви: E-V68 и E-Z827.
Наиболее часто описываемыми субкладами являются E-M78, часть E-V68, и E-M81 , являющийся ветвью E-Z827. Эти два субклада представляют наибольшую часть современной популяции E-M215. E-M78 встречается на большей части ареала, где встречается E-M215, за исключением Южной Африки. E-M81 встречается в основном в Северной Африке. E-M123 менее распространен, но широко разбросан, со значительными популяциями в определенных частях Африканского Рога, Леванта, Аравии, Иберии и Анатолии. Новая клада (E-V1515) была определена Тромбеттой и др. 2015, возникший около 12 тыс. лет назад (95% ДИ 8,6–16,4) в Восточной Африке, где он в основном распространен в настоящее время. Эта клада включает субклады E-V42, E-M293, E-V92 и E-V6, которые в предыдущей филогении были идентифицированы как базальные клады E-M35. [7]
Внутри E-M35 существуют поразительные параллели между двумя гаплогруппами, E-V68 и E-V257. Оба содержат линию, часто наблюдаемую в Африке (E-M78 и E-M81 соответственно), и группу недифференцированных хромосом, которые в основном встречаются в южной Европе. Эту географическую закономерность можно объяснить расширением авианосцев E-M35, возможно, с Ближнего Востока, как предлагают другие авторы, и разделения на две ветви, разделенные географическим барьером Средиземного моря. Однако отсутствие E-V68* и E-V257* на Ближнем Востоке делает морское распространение между Северной Африкой и Южной Европой более правдоподобной гипотезой.

TMRCA основных узлов в E-M35
[ редактировать ]| TMRCA (Кья) | Тромбетта 2015 | YПолный |
| Э-М215 | 39 | 35,4 |
| *E-M35 | 25 | 23,9 |
| **E-V68 | 20 | 20 |
| ***E-M78 | 15 | 13 |
| **E-Z827 | ? | 23,6 |
| ***E-V257/L19 | ? | 13,9 |
| ****Э-М81 | ? | 2,7 |
| ***E-Z830 | 20 | 19 |
| ****Э-М34 | ? | 15 |
| ****E-V1515 | 19 | ? |
Субклады
[ редактировать ]Э-В68 (E1b1b1a)
[ редактировать ]В E-V68 доминирует более известный субклад E-M78. сообщили о трех особях «E-V68*», которые находятся в E-V68, но не в E-M78 на Сардинии , Тромбетта и др. . (2011) , объявляя об открытии V68. Авторы отметили, что, поскольку E-V68* не был обнаружен в образцах с Ближнего Востока, это, по-видимому, является свидетельством морской миграции из Африки в юго-западную Европу. Е-М78 — широко встречающийся субклад, широко распространённый в Северной Африке , Африканском Роге , Западной Азии , (Ближний Восток и Ближний Восток) «вплоть до Южной Азии», [4] и вся Европа . [24] В европейском распространении пик частоты сосредоточен в некоторых частях Балкан (до почти 50% в некоторых регионах). [2] [25] и Сицилии, а снижение частоты наблюдается в направлении Западной, Центральной и Северо-Восточной Европы.
На основании STR данных генетической дисперсии Cruciani et al. (2007) предполагает, что E-M78 зародился в районе Египта и Ливии . [Примечание 3] около 18 600 лет назад (17 300 – 20 000 лет назад). [Примечание 4] Батталья и др. (2008) описывают Египет как «центр распространения различных географически локализованных субкладов, связанных с M78» и, основываясь на археологических данных, предполагают, что точка происхождения E-M78 (в отличие от более позднего распространения из Египта) может находились в рефугиуме , который «существовал на границе современного Судана и Египта, недалеко от озера Нубия , до наступления влажной фазы около 8500 г. до н.э. Движущиеся на север пояса осадков в этот период также могли стимулировать быструю миграцию мезолитических и, в конечном счете , собирателей на север в Африке, Леванте далее в Малую Азию и Европу, где каждый из них в конечном итоге дифференцировался на свои регионально отличительные ветви». На юге Хасан и др. (2008) также объясняют доказательства того, что некоторые субклады E-M78, особенно E-V12 и E-V22, «могли быть завезены в Судан из Северной Африки после прогрессирующего опустынивания Сахары около 6 000–8 000 лет назад». Аналогичным образом, Cruciani et al. (2007) предполагают, что E-M78 из Эфиопии, Сомали и прилегающих территорий обратно мигрировал в этот регион со стороны Египта после приобретения мутации E-M78.
Недавно E-M78 был датирован Тромбеттой и др. 2015. Между 20 300 и 14 800 лет назад. [7]
Субклады E-M78
[ редактировать ]Здесь перечислены основные субклады M78 по состоянию на июнь 2015 года. В субкладе E-M78 Trombetta et al. В 2015 году большая часть бывших хромосом E-M78* была распределена по трем новым отдельным ветвям: E-V1083*, E-V1477 и E-V259. Первая является сестрой парагруппы клад E-V22 и E-V13. Мутация V1477 определяет новую базальную ветвь, которая наблюдалась только в одном образце из Северной Африки. Наконец, сестринская клада E-V12, определенная V264, включает E-V65 и V259, новую линию, распространенную в Центральной Африке. [7] [24]
- E-M78 (E1b1b1a1) Северная Африка, Африканский Рог, Западная Азия, Европа (ранее «E1b1b1a»).
- E-M78* Встречается в Марокко, на юге Португалии, на юге Испании и в Иране (провинции Тегеран и Семнан).
- E-V1477 Встречается у тунисских евреев.
- E-V1083
- E-V1083* Встречается только в Эритрее (1,1%) и Сардинии (0,3%).
- E-V13 Это наиболее распространенный субклад E-M215, встречающийся в Европе. Особенно это распространено на Балканах.
- Э-В22 . Сосредоточены в Северо-Восточной Африке и на Ближнем Востоке. Пики среди Сахо.
- E-V1129
- Э-В12 . Встречается в Египте, Судане, Чаде и других местах. Имеет важный субклад
- E-V12* Наиболее распространенная линия среди южных египтян (74,5%).
- Э-В32 . Очень распространен среди сомалийцев, тигров и оромо.
- Э-В264
- E-V259 найден в Северном Камеруне.
- E-V65 Связан с Северной Африкой, но встречается также на Сицилии, а также в континентальной Италии.
- Э-В12 . Встречается в Египте, Судане, Чаде и других местах. Имеет важный субклад
- E-M521 Не упоминается Тромбеттой и др., 2015 г. Обнаружен у двух особей в Греции Battaglia et al. (2008) и у одного человека из восточно-альпийского региона Италии Coia et al. (2013)
E-Z827 (E1b1b1b)
[ редактировать ]В генетике человека E-Z827 — это название основной гаплогруппы ДНК Y-хромосомы человека, широко встречающейся в Северной Африке, особенно в странах Магриба , и в меньшей степени на Африканском Роге , на Ближнем Востоке и в Европе .
E-V257/E-L19 (E1b1b1b1)
[ редактировать ]Особи E-V257* в их выборках имели E-V257, но не E-M81. Борана из Кении, марракешский бербер, корсиканец, сардинец, южный испанец и кантабрийец. Как упоминалось выше, Тромбетта и др. (2011) предполагают, что отсутствие E-V257* на Ближнем Востоке (Yfull нашел молодой в иранском Азербайджане [26] и другой молодой в Армении [27] ) делает морское движение из северной Африки в южную Европу наиболее правдоподобной гипотезой, объясняющей его распространение. Yfull перечисляет 24 человека, все из которых принадлежат к одной ветви, которая на 30% моложе, чем их общий предок с M81. [28]
Э-М81
[ редактировать ]E-M81 — наиболее распространенный субклад гаплогруппы E-L19/V257. Он сконцентрирован в Магрибе , и в нем доминирует субклад E-M183. Считается, что E-M183 возникла в Северо-Западной Африке и ее предполагаемый возраст составляет 4700 лет назад. [29]
Эта гаплогруппа достигает средней частоты 61% в Магрибе и 51% в Северной Африке , частота снижается примерно с 80% до 100% в берберском населении. [30] [31] включая сахарцев, примерно до 29% к востоку от этого ареала в Египте. [32] [33] [34] [35] Из-за своего молодого возраста и распространенности среди этих групп, а также среди других, таких как мозабитцы , средний атлас , кабильцы и другие берберские группы, его иногда называют генетическим «берберским маркером». Перейра и др. (2010) сообщают о высоких уровнях туарегов в двух популяциях Сахары – 77,8% в районе Гором-Горома в Буркина-Фасо и 81,8% в Госи в Мали . В окрестностях Танута в Нигере частота встречаемости была гораздо ниже - 11,1% . E-M81 также довольно распространен среди арабоязычных групп Магриба . Обычно он встречается с частотой около 45% в прибрежных городах Алжира и Туниса ( Джиджель , Оран , Тизи-Узу , Алжир , Тунис , Сус ). [32] [36]
В этом ключевом районе от Египта до океана Атлантического Соле-Мората и др. (2017) сообщают о закономерности M183-SM001 уменьшения изменчивости микросателлитных гаплотипов (что подразумевает больший возраст линии в прежних областях) у племени регибат в Оране , и они обнаружили M183 * (не SM001) в Иберии, Ливии и Марокко. Арреди и др. (2004) , однако, показали уменьшение вариаций микросателлитов с востока на запад, сопровождающееся существенным увеличением частоты. На восточной окраине этого основного ареала Куджанова и др. (2009) обнаружили M81 у 28,6% (10 из 35 мужчин) в Эль-Хайезе в Ливийской пустыне в Египте.
Арреди и др. (2004) считают, что характер распределения и дисперсии соответствует гипотезе « демического распространения » с Востока. Автохтонного присутствия E-M81 на Ближнем Востоке нет (есть в Ливане). [37] ), что указывает на то, что M81, скорее всего, произошел от своей родительской клады M35 либо в Северной Африке, либо, возможно, на юге, вплоть до Африканского Рога. [38]
В Европе E-M81 широко распространен, но редок, на Пиренейском полуострове показывает среднюю частоту 4% (45/1140), на Пиренейском полуострове с частотами, достигающими 3,5% в Галисии , 4% в Западной Андалусии и Северо-Западной Кастилии . Однако в это исследование включены 153 человека с островов Майорка, Менорка и Ибица, а также 24 человека из Гаскони, которые не находятся на Пиренейском полуострове. Без этих 177 особей средний показатель по Пиренейскому полуострову составляет 4,1% (40/963), [39] он встречается на уровне, сравнимом с E-M78, со средней частотой около 5%. Его частота выше в западной половине полуострова: частота достигает 8% в Эстремадуре и южной Португалии, от 4% до 5% в Галисии , 5% в западной Андалусии и 4% на северо-западе Кастилии и от 9% до 17% в Кантабрии . [19] [40] [41] [42] [43] Самые высокие частоты этой клады, обнаруженные на данный момент в Европе, наблюдались в Валлес-Пазиегос из Кантабрии и варьируются от 5,5% (8/45). [43] до 41% (23/56). [3] Средняя частота 8,28% (54/652) также зарегистрирована на испанских Канарских островах, а частота более 10% на трех крупнейших островах: Тенерифе (10,68%), Гран-Канария (11,54%) и Фуэртевентура (13,33%). [44]
E-M81 также встречается во Франции , [3] 2,70% (15/555) в целом с частотой, превышающей 5% в Оверни (5/89) и Иль-де-Франс (5/91), [45] [46] от 0,7% до 5,8% на Сардинии , [47] [48] примерно 2,12% в целом на Сицилии (но до 7,14% на площади Пьяцца Армерина ), [49] и гораздо ниже частота около Лучеры (1,7%), в континентальной Италии , [42] возможно, из-за древних миграций во времена исламской , римской и карфагенской империй. В исследовании 2014 года, проведенном Стефанией Сарно и соавт. при 326 образцах из Козенцы , Реджо-ди-Калабрии , Лечче и пяти сицилийских провинций E-M81 показывает среднюю частоту 1,53%, но типичный магрибинский основной гаплотип 13-14-30-24-9-11-13 был обнаружен только в двое из пяти особей E-M81. Эти результаты, наряду с незначительным вкладом североафриканских популяций, выявленным с помощью анализа примесных графиков, предполагают лишь незначительное влияние транссредиземноморских потоков генов на текущий генетический пул SSI. [49] [50]
В результате своего распространения в Старом Свете этот субклад встречается по всей Бразилии. [51] 5,4% в Бразилии (Рио-де-Жанейро), [Примечание 5] а среди латиноамериканцев из Калифорнии и Гавайев - 2,4%. [52]
В меньших количествах мужчин E-M81 можно найти в районах, соприкасающихся с Северной Африкой , как вокруг Сахары , в таких местах, как Судан , и вокруг Средиземноморья, в таких местах, как Ливан , Турция , и среди евреев-сефардов .
Есть два признанных субклада E-M81, хотя один из них гораздо важнее другого. [ который? ]
Субклад E-M81 был обнаружен в древних окаменелостях гуанчей (бимбапе), раскопанных в Пунта-Азуле, Эль-Йерро , Канарские острова , которые датируются 10 веком (~ 44%). [53]
Э-М107
[ редактировать ]Андерхилл (2000) нашел один экземпляр E-M107 в Мали .
Э-М183
[ редактировать ]E-M183 чрезвычайно доминирует (более 99% [54] ) внутри E-M81. Карафет и др. (2008) впервые описали его как субклад E-M81.Известные субклады E-M183 включают:
- E-M165 Андерхилл и др. (2000) нашли один пример на Ближнем Востоке.
- E-L351 Обнаружен у двух родственных участников проекта E-M35 Phylogeny Project .
E-Z830 (E1b1b1b2)
[ редактировать ]Это недавно открытый субклад, который еще не включен в большинство деревьев гаплогрупп. E-Z830 включает подтвержденные субклады E-M123, E-V1515 (E-M293, E-V42, E-V6, E-V92), и E-Z830*, и является родственной кладой E-L19. В настоящее время, [55] проект филогении E-M35] выделяет четыре отдельных кластера носителей Z830*, два из которых имеют исключительно еврейское происхождение. Остальные два значительно меньше и включают разрозненных особей в Германии, Испании, Латинской Америке, Египте и Эфиопии. [56] [57] [58] [59]
Э-М123
[ редактировать ]E-M123 в основном известен своим основным субкладом E-M34, который доминирует в этом кладе. [Примечание 6]
E-V1515
[ редактировать ]Новая клада (E-V1515) была определена Тромбеттой и др. 2015, возникший около 12 тыс. лет назад (95% ДИ 8,6–16,4) в Восточной Африке, где он в основном распространен в настоящее время. Эта клада включает все гаплогруппы к югу от Сахары (E-V42, E-M293, E-V92, E-V6), о которых сообщалось как базальные клады E-M35 в предыдущей филогении. [7]
Э-М293
[ редактировать ]E-M293 является субкладом E-V1515. Впервые он был идентифицирован ISOGG как вторая клада внутри E-Z830. Он был обнаружен до E-Z830, о чем было объявлено Henn et al. (2008) , которые связали это с распространением скотоводства из Восточной Африки в Южную Африку . На данный момент высокие уровни были обнаружены у определенных этнических групп в Танзании и Южной Африке. Самыми высокими были народы Датуга (43%), Кхве (Ксоэ) (31%), Бурунге (28%) и Сандаве (24%). Хенн (2008) в своем исследовании также обнаружил двух кенийских мужчин, говорящих на языке банту, с мутацией M293. [5]
Другие субклады E-M215 редки в Южной Африке. Авторы заявляют: «Без информации о M293 у масаи, хема и других популяций в Кении, Судане и Эфиопии мы не можем с большей уверенностью определить точный географический источник M293. Однако имеющиеся данные указывают на современную Танзанию как на ранний и важный географический локус эволюции M293». Они также говорят, что «M293 встречается только в Африке к югу от Сахары, что указывает на отдельную филогенетическую историю для образцов M35.1 * (бывший) дальше на север».
E-P72 появляется в «Карафете» (2008). Тромбетта и др. (2011) объявили, что это субклад E-M293.
Э-В42
[ редактировать ]Тромбетта и др. (2011) объявили об обнаружении E-V42 у двух человек из Бета-Израиль . Было высказано предположение, что оно может быть ограничено регионом вокруг Эфиопии. Однако дальнейшее тестирование, проведенное коммерческими компаниями по тестированию ДНК, подтвердило множество положительных результатов для этого субклада в Саудовской Аравии, Кувейте и у одного человека в Португалии, имеющего корни из Аравии. [60]
Э-В6
[ редактировать ]Субклад E-V6 E-V1515 определяется V6. Кручиани и др. (2004) выявили значительное присутствие этих линий в Эфиопии, а также в соседних сомалийцах . Среди выборок эфиопов и сомалийцев самые высокие показатели были 14,7% среди амхара и 16,7% среди волайта .
Южнее Тишкофф и др. (2007) выявили одного мужчину с V6+ в выборке из 35 жителей Датуги из Танзании. А дальше на север Dugoujon et al. (2009) выявили еще 6 мужчин из выборки из 93 человек из оазиса Сива , который является берберским населением.
Э-В92
[ редактировать ]Тромбетта и др. (2011) объявили об открытии E-V92 в двух Амхарах. Как и E-V6 и E-V42, он, возможно, существует только на территории Эфиопии.
Филогенетика
[ редактировать ]Филогенетическая история
[ редактировать ]До 2002 года в научной литературе существовало как минимум семь систем наименования филогенетического дерева Y-хромосомы. Это привело к значительной путанице. В 2002 году основные исследовательские группы объединились и сформировали Консорциум Y-хромосомы (YCC). Они опубликовали совместный документ, в котором было создано единое новое дерево, которое все согласились использовать. Позже группа гражданских ученых, интересующихся популяционной генетикой и генетической генеалогией, сформировала рабочую группу для создания любительского дерева, стремясь быть прежде всего своевременным. В таблице ниже собраны все эти работы, относящиеся к знаковому дереву YCC 2002 года. Это позволяет исследователю, просматривающему старую опубликованную литературу, быстро перемещаться между номенклатурами.
| YCC 2002/2008 (сокращенно) | (а) | (б) | (с) | (г) | (е) | (г) | (или) | YCC 2002 (от руки) | YCC 2005 (от руки) | YCC 2008 (от руки) | YCC 2010r (от руки) | ИСОГГ 2006 г. | ИСОГГ 2007 | ИСОГГ 2008 | ИСОГГ 2009 | ИСОГГ 2010 | ИЗОГГ 2011 | ИЗОГГ 2012 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| E-P29 | 21 | III | 3А | 13 | Еу3 | Н2 | Б | И* | И | И | И | И | И | И | И | И | И | И |
| Э-М33 | 21 | III | 3А | 13 | Еу3 | Н2 | Б | Е1* | Е1 | Е1а | Е1а | Е1 | Е1 | Е1а | Е1а | Е1а | Е1а | Е1а |
| Э-М44 | 21 | III | 3А | 13 | Еу3 | Н2 | Б | Е1а | Е1а | Е1а1 | Е1а1 | Е1а | Е1а | Е1а1 | Е1а1 | Е1а1 | Е1а1 | Е1а1 |
| Э-М75 | 21 | III | 3А | 13 | Еу3 | Н2 | Б | Е2а | Е2 | Е2 | Е2 | Е2 | Е2 | Е2 | Е2 | Е2 | Е2 | Е2 |
| Э-М54 | 21 | III | 3А | 13 | Еу3 | Н2 | Б | Е2б | Е2б | Е2б | E2b1 | - | - | - | - | - | - | - |
| Е-П2 | 25 | III | 4 | 14 | Еу3 | Н2 | Б | Е3* | Е3 | Е1б | E1b1 | Е3 | Е3 | E1b1 | E1b1 | E1b1 | E1b1 | E1b1 |
| Э-М2 | 8 | III | 5 | 15 | ЕС2 | Н2 | Б | Е3а* | Е3а | E1b1 | E1b1a | Е3а | Е3а | E1b1a | E1b1a | E1b1a | E1b1a1 | E1b1a1 |
| Э-М58 | 8 | III | 5 | 15 | ЕС2 | Н2 | Б | Е3а1 | Е3а1 | E1b1a1 | E1b1a1 | Е3а1 | Е3а1 | E1b1a1 | E1b1a1 | E1b1a1 | E1b1a1a1a | E1b1a1a1a |
| Э-М116.2 | 8 | III | 5 | 15 | ЕС2 | Н2 | Б | E3a2 | E3a2 | E1b1a2 | E1b1a2 | E3a2 | E3a2 | E1b1a2 | E1b1a2 | E1ba12 | удаленный | удаленный |
| Э-М149 | 8 | III | 5 | 15 | ЕС2 | Н2 | Б | Е3а3 | Е3а3 | E1b1a3 | E1b1a3 | Е3а3 | Е3а3 | E1b1a3 | E1b1a3 | E1b1a3 | E1b1a1a1c | E1b1a1a1c |
| Э-М154 | 8 | III | 5 | 15 | ЕС2 | Н2 | Б | Е3а4 | Е3а4 | E1b1a4 | E1b1a4 | Е3а4 | Е3а4 | E1b1a4 | E1b1a4 | E1b1a4 | E1b1a1a1g1c | E1b1a1a1g1c |
| Э-М155 | 8 | III | 5 | 15 | ЕС2 | Н2 | Б | Е3а5 | Е3а5 | E1b1a5 | E1b1a5 | Е3а5 | Е3а5 | E1b1a5 | E1b1a5 | E1b1a5 | E1b1a1a1d | E1b1a1a1d |
| Э-М10 | 8 | III | 5 | 15 | ЕС2 | Н2 | Б | Е3а6 | Е3а6 | E1b1a6 | E1b1a6 | Е3а6 | Е3а6 | E1b1a6 | E1b1a6 | E1b1a6 | E1b1a1a1e | E1b1a1a1e |
| Э-М35 | 25 | III | 4 | 14 | Еу4 | Н2 | Б | E3b* | E3b | E1b1b1 | E1b1b1 | E3b1 | E3b1 | E1b1b1 | E1b1b1 | E1b1b1 | удаленный | удаленный |
| Э-М78 | 25 | III | 4 | 14 | Еу4 | Н2 | Б | E3b1* | E3b1 | E1b1b1a | E1b1b1a1 | E3b1a | E3b1a | E1b1b1a | E1b1b1a | E1b1b1a | E1b1b1a1 | E1b1b1a1 |
| Э-М148 | 25 | III | 4 | 14 | Еу4 | Н2 | Б | E3b1a | E3b1a | E1b1b1a3a | E1b1b1a1c1 | E3b1a3a | E3b1a3a | E1b1b1a3a | E1b1b1a3a | E1b1b1a3a | E1b1b1a1c1 | E1b1b1a1c1 |
| Э-М81 | 25 | III | 4 | 14 | Еу4 | Н2 | Б | E3b2* | E3b2 | E1b1b1b | E1b1b1b1 | E3b1b | E3b1b | E1b1b1b | E1b1b1b | E1b1b1b | E1b1b1b1 | E1b1b1b1a |
| Э-М107 | 25 | III | 4 | 14 | Еу4 | Н2 | Б | E3b2a | E3b2a | E1b1b1b1 | E1b1b1b1a | E3b1b1 | E3b1b1 | E1b1b1b1 | E1b1b1b1 | E1b1b1b1 | E1b1b1b1a | E1b1b1b1a1 |
| Э-М165 | 25 | III | 4 | 14 | Еу4 | Н2 | Б | E3b2b | E3b2b | E1b1b1b2 | E1b1b1b1b1 | E3b1b2 | E3b1b2 | E1b1b1b2a | E1b1b1b2a | E1b1b1b2a | E1b1b1b2a | E1b1b1b1a2a |
| Э-М123 | 25 | III | 4 | 14 | Еу4 | Н2 | Б | E3b3* | E3b3 | E1b1b1c | E1b1b1c | E3b1c | E3b1c | E1b1b1c | E1b1b1c | E1b1b1c | E1b1b1c | E1b1b1b2a |
| Э-М34 | 25 | III | 4 | 14 | Еу4 | Н2 | Б | E3b3a* | E3b3a | E1b1b1c1 | E1b1b1c1 | E3b1c1 | E3b1c1 | E1b1b1c1 | E1b1b1c1 | E1b1b1c1 | E1b1b1c1 | E1b1b1b2a1 |
| Э-М136 | 25 | III | 4 | 14 | Еу4 | Н2 | Б | E3ba1 | E3b3a1 | E1b1b1c1a | E1b1b1c1a1 | E3b1c1a | E3b1c1a | E1b1b1c1a1 | E1b1b1c1a1 | E1b1b1c1a1 | E1b1b1c1a1 | E1b1b1b2a1a1 |
Научные публикации
[ редактировать ]Следующие исследовательские группы в соответствии с их публикациями были представлены в создании Дерева YCC.
Филогенетические деревья
[ редактировать ]Кладограмма с основными субкладами:
| E1b1b ( M215 ) |
| ||||||
Следующее филогенетическое дерево основано на дереве YCC 2008 и последующих опубликованных исследованиях, обобщенных ISOGG. Он включает все известные субклады по состоянию на июнь 2015 г. (Тромбетта и др., 2015). [61] [62] [63]
- Э-М215 (E1b1b)
- Э-М215* . Редкий или несуществующий.
- Э-М35 (E1b1b1)
- Э-В68 (E1b1b1a)
- E-V2009 . Встречается у особей на Сардинии и в Марокко.
- Э-М78 (Е1б1б1а1). Северная Африка, Африканский Рог, Западная Азия, Сицилия. (Ранее «E1b1b1a».)
- Э-М78*
- E-V1477 . Встречается у тунисских евреев.
- E-V1083 .
- E-V1083* . Встречается только в Эритрее (1,1%) и Сардинии (0,3%).
- Э-В13
- Э-В22
- E-V1129
- Э-В12
- Э-В12*
- Э-В32
- Э-В264
- Э-В259 . Найден в Северном Камеруне.
- Э-В65
- E-CTS194
- Э-В12
- E-Z827 (E1b1b1b) [64]
- Э-В257/Л19 (Л19, В257) – Е1b1b1b1 [64]
- E-PF2431
- Э-М81 (М81)
- E-PF2546
- E-PF2546*
- E-CTS12227
- Э-МЗ11
- Э-МЗ12
- Э-МЗ11
- E-A929
- E-Z5009
- E-Z5009*
- E-Z5010
- E-Z5013
- E-Z5013*
- E-A1152
- E-A2227
- Е-А428
- Э-МЗ16
- E-PF6794
- E-PF6794*
- E-PF6789
- Э-МЗ21
- Э-МЗ23
- Э-МЗ80
- Е-А930
- E-Z2198/E-MZ46
- Е-А601
- E-L351
- E-Z5009
- E-PF2546
- E-Z830 (Z830) – E1b1b1b2 [64]
- Э-М123 (М123)
- Э-М34 (М34)
- Э-М84 (М84)
- Э-М136 (М136)
- Э-М290 (М290)
- Э-В23 (В23)
- Э-Л791 (Л791,Л792)
- Э-М84 (М84)
- Э-М34 (М34)
- Э-В1515 . E-V1515 и его субклады распространены в основном в Восточной Африке.
- E-V1515*
- E-V1486
- E-V1486*
- E-V2881
- E-V2881*
- E-V1792
- Э-В92
- Э-М293 (М293)
- Э-М293*
- Э-П72 (П72)
- E-V3065*
- Э-В1700
- Э-В42 (В42)
- E-V1785
- E-V1785*
- Э-V6 (V6)
- Э-М123 (М123)
- Э-В257/Л19 (Л19, В257) – Е1b1b1b1 [64]
- E-V16/E-M281 (E1b1b2). Редкий. Встречается у особей в Эфиопии, Йемене и Саудовской Аравии.
- Э-В68 (E1b1b1a)
См. также
[ редактировать ]Генетика
[ редактировать ]- Африканская примесь в Европе
- генетическая генеалогия
- Гаплогруппа D
- Гаплогруппа DE
- Гаплогруппа
- Гаплотип
- Гаплогруппа ДНК Y-хромосомы человека
- Молекулярная филогенетика
- Парагруппа
- Субклад
- Гаплогруппы Y-хромосомы в популяциях мира
- Гаплогруппы Y-ДНК по этническим группам
- Гаплогруппы Y-ДНК в населении стран Африки к югу от Сахары
Y-ДНК и субклады
[ редактировать ]- Гаплогруппа E-L485
- Гаплогруппа E-M123
- Гаплогруппа E-M180
- Гаплогруппа E-M215
- Гаплогруппа E-M33
- Гаплогруппа E-M521
- Гаплогруппа E-M75
- Гаплогруппа E-M96
- Гаплогруппа E-P147
- Гаплогруппа E-P177
- Гаплогруппа E-P2
- Гаплогруппа E-V12
- Гаплогруппа E-V13
- Гаплогруппа E-V22
- Гаплогруппа E-M2
- Гаплогруппа E-V65
- Гаплогруппа E-V68
- Гаплогруппа E-Z820
- Гаплогруппа E-Z827
Основополагающее дерево Y-ДНК
[ редактировать ]Примечания
[ редактировать ]- ^ «Данные Y-хромосомы показывают сигнал об отдельной миграции позднего плейстоцена из Африки в Европу через Синай, о чем свидетельствует распределение линий гаплогруппы E3b, что не проявляется в распределении гаплогрупп мтДНК». Андерхилл и Кивисилд (2007 : 547)
- ^ «Парагруппа E-M35 * и гаплогруппа J-12f2a * соответствуют критериям основных линий-основателей AJ, поскольку они широко распространены как в популяциях AJ, так и в популяциях Ближнего Востока, и встречаются с гораздо меньшей частотой в европейских нееврейских популяциях». Бехар и др. (2004)
- ^ Кручиани и др. (2007) используют термин «Северо-Восточная Африка» для обозначения Египта и Ливии, как показано в Таблице 1 исследования. До Cruciani et al. (2007) , Семино и др. (2004) Восточная Африка как возможное место происхождения E-M78, на основе испытаний в Эфиопии. Это произошло из-за высокой частоты и разнообразия линий E-M78 в регионе Эфиопии. Однако Кручиани и др. (2007) смогли изучить больше данных, включая популяции из Северной Африки, которые не были представлены в исследовании Semino et al. (2004) провели исследование и обнаружили доказательства того, что линии E-M78, составляющие значительную часть некоторых популяций в этом регионе, были относительно молодыми ветвями (см. E-V32 ниже). Поэтому они пришли к выводу, что «Северо-Восточная Африка» была вероятным местом происхождения E-M78, основываясь на «периферийном географическом распределении наиболее производных субгаплогрупп по отношению к северо-востоку Африки, а также на результатах количественного анализа UEP и разнообразия микросателлитов». . Так, по данным Cruciani et al. (2007) E-M35, родительская клада E-M78, возникла в Восточной Африке, впоследствии распространилась в Северо-Восточную Африку, а затем произошла «обратная миграция» хромосом E-M215, которые приобрели мутацию E-M78. Кручиани и др. (2007) поэтому отмечают это как свидетельство наличия «коридора двунаправленной миграции» между Северо-Восточной Африкой (Египет и Ливия по их данным) с одной стороны и Восточной Африкой с другой. Авторы полагают, что было «по крайней мере 2 эпизода между 23,9–17,3 тыс. лет назад и 18,0–5,9 тыс. лет назад».
- ^ Кручиани и др. (2007) используют два метода расчета для оценки возраста E-M78, которые дают очень разные результаты. Для основной 18600-летней давности метод ASD используется , а для второго « метода ρ », используемого в качестве проверки, дает 13,7 тыс. лет назад со стандартным отклонением 2,3 тыс. лет назад, но разница между двумя методами велика только для оценка возраста E-M78, а не его субкладов. Авторы заявляют, что большая разница «обусловлена соответствующим отклонением от звездообразной структуры из-за повторяющихся эффектов основателя ».
- ^ (6 из 112), «Присутствие хромосом североафриканского происхождения (E3b1b-M81; Cruciani et al., 2004) также можно объяснить португальским, но особенно итальянским притоком, поскольку эта гаплогруппа достигает частоты 4,6% в Португалии и 4,8% в Италии , что очень похоже на частоту, обнаруженную в Рио-де-Жанейро (4,4%) среди европейских вкладчиков». Сильва и др. (2006)
- ^ Например, по состоянию на 11 ноября 2008 г. проект филогении E-M35. [ постоянная мертвая ссылка ] имел записи о четырех тестах E-M123* по сравнению с 93 результатами тестов E-M34.
Ссылки
[ редактировать ]- ^ Перейти обратно: а б «Э-М35 YДерево» .
- ^ Перейти обратно: а б с д Семино и др. (2004)
- ^ Перейти обратно: а б с д Кручиани и др. (2004)
- ^ Перейти обратно: а б Кручиани и др. (2007)
- ^ Перейти обратно: а б Хенн и др. (2008)
- ^ Хасан и др. (2008)
- ^ Перейти обратно: а б с д и ж Тромбетта, Бениамино; и др. (июль 2015 г.). «Филогеографическое уточнение и крупномасштабное генотипирование гаплогруппы E Y-хромосомы человека дают новое представление о расселении ранних скотоводов на африканском континенте» . Геномная биология и эволюция . 7 (7): 1940–1950. дои : 10.1093/gbe/evv118 . ISSN 1759-6653 . OCLC 5854174538 . ПМЦ 4524485 . ПМИД 26108492 . S2CID 16352575 .
- ^ Крупномасштабное недавнее расширение европейских отцовских линий, продемонстрированное повторным секвенированием населения , Кьяра Батини и др., Nature.com , 2015.
- ^ Лакан и др. (2011)
- ^ «Молекулярно-генетическое исследование истории неолитического населения Западного Карпатского бассейна» (PDF) . Архивировано из оригинала (PDF) 21 июля 2015 г.
- ^ Шунеманн, Верена Дж.; и др. (2017). «Геномы древнеегипетских мумий предполагают увеличение африканского происхождения к югу от Сахары в постримские периоды» . Природные коммуникации . 8 : 15694. Бибкод : 2017NatCo...815694S . дои : 10.1038/ncomms15694 . ПМЦ 5459999 . ПМИД 28556824 .
- ^ Фрегель; и др. (2018). «Древние геномы из Северной Африки свидетельствуют о доисторических миграциях в Магриб как из Леванта, так и из Европы». bioRxiv 10.1101/191569 .
- ^ Родригес-Варела; и др. (2017). «Геномный анализ человеческих останков с Канарских островов доевропейского завоевания показывает близкое родство с современными жителями Северной Африки» . Современная биология . 27 (1–7): 3396–3402.e5. Бибкод : 2017CBio...27E3396R . дои : 10.1016/j.cub.2017.09.059 . hdl : 2164/13526 . ПМИД 29107554 .
- ^ Ван Де Лоосдрехт, «Божьи коровки»; Бузугар, Абдельджалил; Хамфри, Луиза; Пост, Козимо; Бартон, Ник; Ачиму-Петри, Айнуэр; Никель, Биргит; Нагель, Сара; Талби, Эль-Хасан; Эль-Хаджрауи, Мохаммед Абдельджалил; Амзази, Саид; Юблин, Жан-Жак; Паабо, Сванте; Шиффельс, Стефан; Мейер, Матиас; Хаак, Вольфганг; Чон, Чунгвон; Краузе, Йоханнес (4 мая 2018 г.). «Плейстоценовые геномы Северной Африки связывают человеческие популяции Ближнего Востока и Африки к югу от Сахары» . Наука . 360 (6388): 548–552. Бибкод : 2018Sci...360..548V . дои : 10.1126/science.aar8380 . ПМИД 29545507 . S2CID 206666517 .
- ^ Эрет, Кейта и Ньюман (2004) ; Кейта и Бойс (2005) ; Кейта (2008 )
- ^ Бехар и др. (2003)
- ^ Бехар и др. (2004)
- ^ Шен и др. (2004)
- ^ Перейти обратно: а б Адамс и др. (2008)
- ^ Небель и др. (2001)
- ^ Каденас и др. (2007)
- ^ Тромбетта и др. (2011)
- ^ (Рисунок S.7) Дж. Д. Кристофаро и др., 2013, «Афганский Гиндукуш: где сходятся потоки генов на субконтиненте Евразии», http://www.plosone.org/article/info%3Adoi%2F10.1371% 2Fjournal.pone.0076748
- ^ Перейти обратно: а б Кручиани и др. (2006)
- ^ Перичич и др. (2005)
- ^ «E-Y133414 YДерево» .
- ^ «E-FGC18960 YДерево» .
- ^ "E-L19 YTree" .
- ^ «E-M81 YTree» .
- ^ Кручиани и др. (2004) .
- ^ Фадхлауи-Зид, Карима; Мартинес-Крус, Бегона; Ходжет-эль-Хиль, Хусейн; Мендисабаль, Изабель; Бенаммар-Эльгаайед, Амель; Комас, Дэвид (октябрь 2011 г.). «Генетическая структура тунисских этнических групп, выявленная по отцовской линии». Американский журнал физической антропологии . 146 (2): 271–280. дои : 10.1002/ajpa.21581 . ISSN 1096-8644 . ПМИД 21915847 .
- ^ Перейти обратно: а б Арреди и др. (2004)
- ^ Альварес и др. (2009)
- ^ Бош и др. (2001)
- ^ Куянова, Мартина; Перейра, Луиза; ФЕРНАНДЕС, Вероника; Перейра, Джоана Б.; Черны, Виктор (октябрь 2009 г.). «Генетический вклад ближневосточного неолита в небольшой оазис Западной египетской пустыни». Американский журнал физической антропологии . 140 (2): 336–346. дои : 10.1002/ajpa.21078 . ISSN 1096-8644 . ПМИД 19425100 .
- ^ Робин и др. (2008)
- ^ «E-A5604 YTree» .
- ^ Кейта (2008)
- ^ см. таблицу. Архивировано 30 июля 2013 г. на Wayback Machine .
- ^ Флорес и др. (2005)
- ^ Красота и др. (2006)
- ^ Перейти обратно: а б Капелли и др. (2009)
- ^ Перейти обратно: а б Мака-Мейер и др. (2003)
- ^ Фрегель и др. (2009) , см. таблицу
- ^ Рамос-Луиза и др. (2009)
- ^ Анализировались только мужчины с французской фамилией, чтобы попытаться исключить более поздних иммигрантов.
- ^ Грюни, Виола; Равеан, Алессандро; Коломбо, Джулия; Ники, Кармен; Кробу, Франческа; Онгаро, Линда; Батталья, Винченца; Санна, Дарья; Аль-Захери, Надя; Фиорани, Орнелла; Лиза, Антонелла (16 октября 2019 г.). «Анализ Y-хромосомы и фамилий для реконструкции прошлых структур населения: население Сардинии как тестовый пример» . Международный журнал молекулярных наук . 20 (22): 5763. doi : 10.3390/ijms20225763 . ISSN 1422-0067 . ПМК 6888588 . ПМИД 31744094 .
- ^ Франкалаччи и др. (2013), Низкопроходное секвенирование ДНК 1200 сардинцев реконструирует европейскую филогению Y-хромосомы
- ^ Перейти обратно: а б Ди Гаэтано и др. (2009)
- ^ Сарно, С; Боаттини, А; Карта, М; Ферри, Дж; Алу, М; Яо, ДЮ; Чиани, Дж; Петтенер, Д; Луизелли, Д. (2014). «Древний средиземноморский плавильный котел: исследование однородительской генетической структуры и истории населения Сицилии и Южной Италии» . ПЛОС ОДИН . 9 (4): е96074. Бибкод : 2014PLoSO...996074S . дои : 10.1371/journal.pone.0096074 . ПМЦ 4005757 . ПМИД 24788788 .
 Эта статья содержит цитаты из этого источника, который доступен по лицензии Creative Commons Attribution 4.0 International (CC BY 4.0) .
Эта статья содержит цитаты из этого источника, который доступен по лицензии Creative Commons Attribution 4.0 International (CC BY 4.0) . - ^ (8 из 132), Mendizabal et al. (2008)
- ^ (7 из 295), Paracchini et al. (2003)
- ^ Ордоньес, AC; Фрегель, Р.; Трухильо-Медерос, А.; Эрвелла, М.; де-ла-Руа, К.; Арне-де-ла-Роса, М. (2017). «Генетические исследования доиспанского населения, захороненного в пещере Пунта-Асуль (Эль-Йерро, Канарские острова)». Журнал археологической науки . 78 : 20–28. Бибкод : 2017JArSc..78...20O . дои : 10.1016/j.jas.2016.11.004 .
- ^ «E-M81 YTree» .
- ^ [1] [ постоянная мертвая ссылка ]
- ^ «Данные проекта E-M35» . Архивировано из оригинала 24 сентября 2015 г. Проверено 1 апреля 2012 г.
- ^ «Данные проекта E-M35» . Архивировано из оригинала 24 сентября 2015 г. Проверено 1 апреля 2012 г.
- ^ «Данные проекта E-M35» . Архивировано из оригинала 24 сентября 2015 г. Проверено 1 апреля 2012 г.
- ^ «Данные проекта E-M35» . Архивировано из оригинала 24 сентября 2015 г. Проверено 1 апреля 2012 г.
- ^ «Данные проекта E-M35» . Архивировано из оригинала 24 сентября 2015 г. Проверено 14 января 2013 г.
- ^ ИЗОГГ (2011)
- ^ Карафет и др. (2008)
- ^ Консорциум Y-хромосомы "YCC" (2002)
- ^ Перейти обратно: а б с ИЗОГГ 2015
Библиография
[ редактировать ]- Адамс, Сьюзен М; Босх, Елена; Балареск, Патрисия Л.; Баллеро, Стефан Ж.; Ли, Эндрю С.; Арройо, Эдуардо; Лопес-Парра, Ана М.; Алер, Мерседес; и др. (2008), «Генетическое наследие религиозного разнообразия и нетерпимости: отцовские линии христиан, евреев и мусульман на Пиренейском полуострове», Американский журнал генетики человека , 83 (6): 725–36, doi : 10.1016/j .ajhg.2008.11.007 , PMC 2668061 , PMID 19061982
- Альварес; Святые, Кристина; Монтьель, Рафаэль; Каейро, Бласкес; Крылья, Абделлатиф; Дюгужон, Жан-Мишель; Алуджа, Мария Пилар (2009), «Вариации Y-хромосомы в Южной Иберии: взгляд на вклад Северной Африки», Американский журнал человеческой биологии , 21 ): 407–409, doi : 10.1002/ajhb.20888 , PMID ( 3 , S2CID 7041905
- Арреди, Б; Полони, Э; Параккини, С ; Зержал, Т; Фаталлах, Д; Макрелуф, М; Паскали, В; Новеллетто, А; Тайлерсмит, К. (2004), «Преимущественно неолитическое происхождение вариаций Y-хромосомной ДНК в Северной Африке», Американский журнал генетики человека , 75 (2): 338–345, doi : 10.1086/423147 , PMC 1216069 , PMID 15202071
- Бадро, Даниэль А.; Дуэи, Бушра; Хабер, Марк; Юханна, Соня С.; Саллум, Анжелика; Гассибе-Саббах, Мишелла; Джонсруд, Брайан; Хазен, Жорж; Матису-Смит, Элизабет; Сория-Эрнанц, Дэвид Ф.; Уэллс, Р. Спенсер; Тайлер-Смит, Крис; Платт, Дэниел Э.; Заллуа, Пьер А. (2013), «Генетика Y-хромосомы и мтДНК выявляет значительные различия в сходстве современного населения Ближнего Востока с населением Европы и Африки», PLOS ONE , 8 (1: e54616): e54616, Bibcode : 2013PLoSO.. .854616B , doi : 10.1371/journal.pone.0054616 , PMC 3559847 , PMID 23382925
- Батталья, Винченца; Форнарино, Симона; Аль-Захери, Надя; Оливьери, Анна; Лопата, Мэри; Майрес, Натали М; Кинг, Рой Дж; Корни, Сирия; и др. (2008), «Y-хромосомные свидетельства культурного распространения сельского хозяйства в юго-восточной Европе», European Journal of Human Genetics , 17 (6): 820–830, doi : 10.1038/ejhg.2008.249 , PMC 2947100 , PMID 19107149
- Бехар, Дорон М.; Томас, Марк Г.; Скорецкий, Карл; Хаммер, Майкл Ф.; Булыгина Екатерина; Розенгартен, Дрор; Джонс, Эбигейл Л.; Держись, Карен; и др. (октябрь 2003 г.), «Множественное происхождение ашкеназских левитов: свидетельства Y-хромосомы как ближневосточного, так и европейского происхождения», Am. Дж. Хум. Жене. , том. 73, нет. 4, стр. 768–779, doi : 10.1086/378506 , PMC 1180600 , PMID 13680527 . Также на http://www.ucl.ac.uk/tcga/tcgapdf/Behar-AJHG-03.pdf и https://web.archive.org/web/20090304100321/http://www.familytreedna.com/ pdf/400971.pdf
- Бехар; Гарриган; Каплан; Мобашер; Розенгартен (ноябрь 2004 г.), «Контрастные закономерности вариаций Y-хромосомы у евреев-ашкенази и нееврейского европейского населения» (PDF) , Hum. Жене. , том. 114, нет. 4, стр. 354–365, doi : 10.1007/s00439-003-1073-7 , PMID 14740294 , S2CID 10310338 , заархивировано из оригинала (PDF) 10 ноября 2011 г. , получено 11 сентября 2009 г.
- КРАСОТА, Сандра; Гусмао, Леонор; ЛОПЕС, Александра; АЛВЕС, Синтия; ГОМЕС, Ива; Джузели, Мария; Калафель, Франческ; Карраседо, Анхель; Аморим, Антонио (2006), «Микрофилогеографическая и демографическая история португальских мужских линий», Annals of Human Genetics , 70 (2): 181–194, doi : 10.1111/j.1529-8817.2005.00221.x , PMID 16626329 , S2CID 4652154
- Бёрд, Стивен (2007), «Гаплогруппа E3b1a2 как возможный индикатор поселения в Римской Британии солдат балканского происхождения» , Журнал генетической генеалогии , 3 (2), заархивировано из оригинала 22 апреля 2016 г. , получено в 2023 г. 04-20
- Бортолини; Томас, Марк Г.; Чихи, Лурдес; Игл, Джон А.; Кастро-Оф-Вар, Динора; Сальзано, Фрэнсис М.; Руис-Линарес, Андрес (2004), «Типология Рибейро, геномы и испанский колониализм с точки зрения Гран-Канарии и Колумбии» (PDF) , Генетика и молекулярная биология , 27 (1): 1–8, doi : 10.1590/S1415 - 47572004000100001
- Босх, Елена; Калафель, Франческ; Комас, Дэвид; Офнер, Питер Дж.; Андерхилл, Питер А.; Бертранпети, Жауме (2001), «Анализ вариаций Y-хромосомы человека с высоким разрешением показывает резкий разрыв и ограниченный поток генов между северо-западной Африкой и Пиренейским полуостровом», Am J Hum Genet , 68 (4): 1019–1029 , doi : 10.1086/319521 , PMC 1275654 , PMID 11254456
- Бош, Э.; Калафель, Ф.; Гонсалес-Нейра, А.; Флейз, К.; Матеу, Э.; Шейль, Х.-Г.; Хакенбек, В.; Ефремовская, Л.; и др. (2006), «Отцовские и материнские линии на Балканах демонстрируют однородный ландшафт, несмотря на языковые барьеры, за исключением изолированных аромунов» , Annals of Human Genetics , 70 (4): 459–487, doi : 10.1111/j.1469-1809.2005. .00251.x , PMID 16759179 , S2CID 23156886 , заархивировано из оригинала 10 декабря 2012 г.
- Каденас; Животовский Лев А; Кавалли-Сфорца, Лука Л; Андерхилл, Питер А; Эррера, Рене Дж. (2007), «Разнообразие Y-хромосомы характеризует Оманский залив», Европейский журнал генетики человека , 16 (3): 1–13, doi : 10.1038/sj.ejhg.5201934 , PMID 17928816
- Капелли, Кристиан; Рыжая, Никола; Абернети, Джулия К.; Гратрикс, Фиона; Уилсон, Джеймс Ф.; Моен, Торольф; Хервиг, Тор; Ричардс, Мартин; и др. (2003), «Перепись хромосом AY на Британских островах», Current Biology , 13 (11): 979–84, Бибкод : 2003CBio…13..979C , doi : 10.1016/S0960-9822(03)00373-7 , hdl : 20.500.11820/8acb01f3-a7c1-45f5-89de-b796266d651e , PMID 12781138 также в [2]
- Каратти; Джино, С.; Торре, К.; Робино, К. (2009), «Подтипирование Y-хромосомной гаплогруппы E-M78 (E1b1b1a) с помощью анализа SNP и его судебно-медицинское применение», International Journal of Legal Medicine , 123 (4): 357–360, doi : 10.1007/s00414 -009-0350-y , PMID 19430804 , S2CID 5657112
- Волосы, Кристиан; Онофри, Валерио; Бризигелли, Франческа; Боски, Илария; Скарниччи, Франческа; Масулло, Мара; Ферри, Джанмарко; Тофанелли, Серджио; и др. (2009), «Мавры и сарацины в Европе: оценка средневекового североафриканского мужского наследия в южной Европе», European Journal of Human Genetics , 17 (6): 848–852, doi : 10.1038/ejhg.2008.258 , PMC 2947089 , PMID 19156170
- Чинниоглу, Ченгиз; Кинг, Рой; Кивисилд, Тоомас; Калфоглу, Эрси; Атасой, Севиль; Каваллери, Джанпьеро Л.; Лилли, Анита С.; Роузман, Чарльз К.; и др. (2004), «Раскопки слоев гаплотипов Y-хромосомы в Анатолии», Hum Genet , 114 (2): 127–48, doi : 10.1007/s00439-003-1031-4 , PMID 14586639 , S2CID 10763736
- Конту, Даниэла; Морелли, Даниэла; Сантони, Федерико; Фостер, Джейми В.; Франкалаччи, Паоло; Кукка, Франческо (2008), «Свидетельства на основе Y-хромосомы донеолитического происхождения генетически однородного, но разнообразного населения Сардинии: выводы для сканирования ассоциаций», PLOS ONE , 3 (1): e1430, Bibcode : 2008PLoSO...3.1430 C , doi : 10.1371/journal.pone.0001430 , PMC 2174525 , PMID 18183308
- Кручиани, Фульвио; Сантоламазца, Пьеро; Шен, Пейдун; Маколей, Винсент; Мораль, Педро; Олкерс, Антонель; Модиано, Дэвид; Холмс, Сьюзан (2002), «Обратная миграция из Азии в страны Африки к югу от Сахары поддерживается анализом гаплотипов Y-хромосомы человека с высоким разрешением», American Journal of Human Genetics , 70 (5): 1197–1214, doi : 10.1086/340257 , ПМК 447595 , ПМИД 11910562
- Кручиани; Ла Фратта; Сантоламазца; Селлитто (май 2004 г.), «Филогеографический анализ Y-хромосом гаплогруппы E3b (E-M215) выявляет множественные миграционные события внутри и из Африки» (PDF) , American Journal of Human Genetics , 74 (5): 1014–1022, doi : 10.1086/386294 , PMC 1181964 , PMID 15042509 , заархивировано из оригинала (PDF) 26 июня 2008 г. , получено 17 мая 2008 г.
- Кручиани; Ла Фратта; Торрони; Андерхилл; Скоццари (2006), «Молекулярное рассечение гаплогруппы E-M78 (E3b1a) Y-хромосомы: апостериорная оценка подхода, основанного на микросателлитных сетях, с помощью шести новых биаллельных маркеров», Human Mutation , 27 (8): 831–2, doi : 10.1002/humu.9445 , PMID 16835895 , S2CID 26886757
- Кручиани, Ф.; Ла Фратта, Р.; Тромбетта, Б.; Сантоламазца, П.; Селитто, Д.; Коломб, Э.Б.; Дюгужон, Ж.-М.; Кривелларо, Ф.; и др. (2007), «Отслеживание прошлых перемещений мужчин-мужчин в Северной/Восточной Африке и Западной Евразии: новые подсказки по гаплогруппам Y-хромосомы E-M78 и J-M12» , Molecular Biology and Evolution , 24 (6): 1300–1311, doi : 10.1093/molbev/msm049 , PMID 17351267 , заархивировано из оригинала 10 октября 2017 г. Также см. Дополнительные данные .
- Ди Гаэтано; Черутти, Франческа; Кробу, Карло; Робино (2009), «Дифференциальная миграция из Греции и Северной Африки на Сицилию подтверждается генетическими данными Y-хромосомы», European Journal of Human Genetics , 17 (1): 91–99, doi : 10.1038/ejhg.2008.120 , PMC 2985948 , PMID 18685561
- Дюгужон; Кудрэ; Торрони; Кручиани; Скоццари; Мораль; Луали; Коссманн (2009), д'Эррико; Хомберт (ред.), «Бербер и берберы: генетическое и языковое разнообразие» (PDF) , Становясь красноречивыми, достижения в возникновении языка, человеческого познания и современных культур : 123–146, doi : 10.1075/z.152.05ch4 , ISBN 978-90-272-3269-4 , заархивировано из оригинала (PDF) 19 октября 2016 г. , получено 20 апреля 2023 г.
- Эрет, К.; Кейта, СО; Ньюман, П. (2004), «Происхождение афроазиатского языка», Science , 306 (5702): 1680, doi : 10.1126/science.306.5702.1680c , PMID 15576591 , S2CID 8057990
- Эль-Сибай, Мирват; Платт, Дэниел Э.; Хабер, Марк; Сюэ, Яли; Юханна, Соня С.; Уэллс, Р. Спенсер; Изаабель, Хасан; Санюра, Мэй Ф.; и др. (2009), «Географическая структура Y-хромосомного генетического ландшафта Леванта: контраст прибрежных и внутренних территорий», Annals of Human Genetics , 73 (6): 568–581, doi : 10.1111/j.1469-1809.2009.00538 .x , PMC 3312577 , PMID 19686289
- Фирасат; Халик, Шагуфта; Мохьюддин, Аиша; Папайоанну, Мирто; Тайлер-Смит, Крис; Андерхилл, Питер А; Аюб, Касим (2006), «Y-хромосомные доказательства ограниченного вклада греков в патанскую популяцию Пакистана», European Journal of Human Genetics , 15 (1): 121–126, doi : 10.1038/sj.ejhg.5201726 , ПМК 2588664 , ПМИД 17047675
- Флорес, Карлос; Мака-Мейер, Николь; Гонсалес, Ана М; Офнер, Питер Дж; Шен, Пейдун; Перес, Хосе А; Рохас, Антонио; Ларруга, Хосе М; Андерхилл, Питер А. (2004), «Сокращенная генетическая структура Пиренейского полуострова, выявленная с помощью анализа Y-хромосомы: последствия для демографии населения», Европейский журнал генетики человека , 12 (10): 855–863, doi : 10.1038/sj. ejhg.5201225 , PMID 15280900 , S2CID 16765118
- Флорес; Мака-Мейер, Николь; Ларруга, Хосе М.; Кабрера, Висенте М.; Карадшех, Наиф; Гонсалес, Ана М. (2005), «Изоляты в коридоре миграций: анализ вариаций Y-хромосомы с высоким разрешением в Иордании», J Hum Genet , 50 (9): 435–441, doi : 10.1007/s10038- 005-0274-4 , ПМИД 16142507
- Франкалаччи, П.; Морелли, Л.; Андерхилл, Пенсильвания; Лилли, А.С.; Пассарино, Г.; Усели, А.; Мадедду, Р.; Паоли, Г.; и др. (2003), «Население трех средиземноморских островов (Корсика, Сардиния и Сицилия), основанное на биаллельной изменчивости Y-хромосомы», Американский журнал физической антропологии , 121 (3): 270–279, doi : 10.1002/ajpa.10265 , ПМИД 12772214
- Фрегель, Роза; ГОМЕС, Вероника; ГУСМИО, Леонор; Гонсалес, Ана М; Кабрера, Висенте М; АМОРИМ, Антониу; Ларруга, Хосе М. (2009), «Демографическая история мужского генофонда Канарских островов: замена местных линий Европой», BMC Evolutionary Biology , 9 : 181, Bibcode : 2009BMCEE...9..181F , doi : 10.1186 /1471-2148-9-181 , PMC 2728732 , PMID 19650893
- Жерар; Берриш, С; Ауизерат, А; Дитерлен, Ф; Люкотт, Дж. (2006), «Североафриканское берберское и арабское влияние в западном Средиземноморье, выявленное гаплотипами ДНК Y-хромосомы» , Human Biology , 78 (3): 307–316, doi : 10.1353/hub.2006.0045 , PMID 17216803 , S2CID 13347549
- Гонсалвеш, Р; Фрейтас, А; Белый, М; РОЗА, А; Фернандес, AT; Животовский, Л.А.; Андерхилл, Пенсильвания; Кивисилд, Т; Брем, А. (2005), «Линии Y-хромосомы из Португалии, Мадейры и Азорских островов фиксируют элементы сефардского и берберского происхождения» , Annals of Human Genetics , 69 (Pt 4): 443–454, doi : 10.1111/j.1529- 8817.2005.00161.x , hdl : 10400.13/3018 , PMID 15996172 , S2CID 3229760 [ мертвая ссылка ]
- Хаммер (2003), «Структура человеческой популяции и ее влияние на выборку вариаций последовательности Y-хромосомы», Genetics , 164 (4): 1495–1509, doi : 10.1093/genetics/164.4.1495 , PMC 1462677 , PMID 12930755
- Хасан, Хишам Ю.; Андерхилл, Питер А.; Кавалли-Сфорца, Лука Л.; Ибрагим, Мунтасер Э. (2008), «Вариации Y-хромосомы среди суданцев: ограниченный поток генов, соответствие языку, географии и истории» (PDF) , Американский журнал физической антропологии , 137 (3): 316–23, doi : 10.1002/ajpa.20876 , PMID 18618658 , заархивировано из оригинала (PDF) 4 марта 2009 г.
- Хенн, Б.М.; Жиньу, К.; Лин, Алиса А; Офнер, Питер Дж.; Шен, П.; Скоццари, Р.; Кручиани, Ф.; Тишкофф, С.А.; Маунтин, JL; Андерхилл, Пенсильвания (2008), «Y-хромосомные свидетельства миграции скотоводов через Танзанию в южную Африку», PNAS , 105 (31): 10693–8, Bibcode : 2008PNAS..10510693H , doi : 10.1073/pnas.0801184105 , PMC 2504844 , ПМИД 18678889 . См. комментарии в блоге Dienekes , комментарии в блоге Spitoon и публичный выпуск .
- Гаплогруппа E Y-ДНК и ее субклады – 2011 г. , Международное общество генетической генеалогии (ISOGG), 2011 г. [ нужны разъяснения ]
- Джоблинг, Массачусетс; Тайлер-Смит, К. (2000), «Новые варианты использования новых гаплотипов Y-хромосомы человека, болезни и отбор», Trends Genet. , 16 (8): 356–362, doi : 10.1016/S0168-9525(00)02057-6 , PMID 10904265
- Карафет, ТМ; Мендес, Флорида; Мейлерман, МБ; Андерхилл, Пенсильвания; Зегура, СЛ; Хаммер, М.Ф. (май 2008 г.), «Новые бинарные полиморфизмы изменяют форму и увеличивают разрешение дерева гаплогрупп Y-хромосомы человека» , Genome Research , 18 (5): 830–8, doi : 10.1101/gr.7172008 , PMC 2336805 , PMID 18385274 . Опубликовано в Интернете 2 апреля 2008 г. См. также дополнительные материалы.
- Кейта, Шомарка (2008), «География, избранные афро-азиатские семьи и вариации линии Y-хромосомы» , «В поисках языка в предыстории: очерки в четырех областях антропологии: в честь Гарольда Крейна Флеминга» , Джон Бенджаминс, ISBN 978-90-272-3252-6
- Кейта, СОИ; Бойс, AJ (Энтони Дж.) (2005), «Генетика, Египет и история: интерпретация географических закономерностей вариаций Y-хромосомы» , History in Africa , 32 : 221–246, doi : 10.1353/hia.2005.0013 , S2CID 163020672
- Кинг, Р.Дж.; Озджан, СС; Картер, Т.; Калфоглу, Э.; Атасой, С.; Триантафиллидис, К.; Коваци, А.; Лин, А.А.; и др. (2008), «Дифференциальное анатолийское влияние Y-хромосомы на греческий и критский неолит» (PDF) , Annals of Human Genetics , 72 (2): 205–214, doi : 10.1111/j.1469-1809.2007.00414.x , PMID 18269686 , S2CID 22406638 , заархивировано из оригинала (PDF) 5 марта 2009 г.
- Король; Андерхилл (2002), «Соответствующее распределение расписной керамики и керамических фигурок эпохи неолита с линиями Y-хромосомы», Antiquity , 76 (293): 707–14, doi : 10.1017/S0003598X00091158 , S2CID 160359661
- Куянова; Перейра; Фернандес; Перейра; Черни (2009), «Ближневосточный неолитический генетический вклад в небольшой оазис западной египетской пустыни», Американский журнал физической антропологии , 140 (2): 336–346, doi : 10.1002/ajpa.21078 , PMID 19425100
- Лакан, Мари; Кейзер, Кристина; Рико, Франсуа-Ксавье; Брукато, Николас; Таррус, Хосеп; Босх, Ангел; Гилейн, Жан; Крубези, Эрик; Людес, Бертран (2011), «Древняя ДНК предполагает ведущую роль мужчин в распространении неолита», PNAS , 108 (45): 18255–9, Bibcode : 2011PNAS..10818255L , doi : 10.1073/pnas.1113061108 , PMC 3215063 , ПМИД 22042855
- Ланкастер, Эндрю (2009), «Гаплогруппы Y, археологические культуры и языковые семьи: обзор междисциплинарных сравнений на примере E-M35» (PDF) , Журнал генетической генеалогии , 5 (1), заархивировано из оригинала ( PDF) от 06 мая 2016 г. , получено 20 апреля 2023 г.
- Луис, Дж; Роуолд, Д; Регейро, М; Каейро, Б; Чинниоглу, К; Роузман, К; Андерхилл, П; Каваллисфорса, Л; Эррера, Р. (2004), «Левант против Африканского Рога: доказательства двунаправленных коридоров миграции людей» (PDF) , Американский журнал генетики человека , 74 (3): 532–544, doi : 10.1086/382286 , PMC 1182266 , PMID 14973781 , заархивировано из оригинала (PDF) 16 февраля 2012 г. (Также см. Ошибки )
- Мака-Мейер Н., Санчес-Веласко П., Флауэрс С., Ларруга Дж.М., Гонсалес А.М., Отерино А., Лейва-Кобиан Ф. и др. (2003), «Характеристика Y-хромосомы и митохондриальной ДНК Pasiegos, человеческого изолята из Кантабрии (Испания)», Genetics , 67 (Pt 4): CiteSeerX 329–339 , of Human Annals j.1469-1809.2003.00045.x , PMID 12914567 , S2CID 40355653 .
- Мартинес, Лэйзель; Андерхилл, Питер А; Животовский Лев А; Гайден, Тензин; Мошонас, Николас К; Чоу, Шерил-Эмилиан Т; Конти, Саймон; Мамолини, Элизабетта; Кавалли-Сфорца, Л. Лука; Эррера, Рене (1 апреля 2007 г.), «Палеолитическое наследие Y-гаплогруппы преобладает на Критском высокогорном плато», European Journal of Human Genetics , 15 (4): 485–493, doi : 10.1038/sj.ejhg.5201769 , ISSN 1018-4813 , PMID 17264870
- Мендисабаль, Изабель; Сандовал, Карла; Берниэль-Ли, Джемма; Калафель, Франческ; Салас, Антонио; Мартинес-Фуэнтес, Антонио; Комас, Дэвид (2008), «Генетическое происхождение, примесь и асимметрия в материнских и отцовских линиях человека на Кубе», BMC Evol. Биол. , 8 : 213, Bibcode : 2008BMCEE...8..213M , doi : 10.1186/1471-2148-8-213 , PMC 2492877 , PMID 18644108
- Небель, Альмут; Филон, Д; Бринкманн, Б; Маджамдер, П; Фаерман, М; Оппенгейм, А. (2001), «Пул Y-хромосомы евреев как часть генетического ландшафта Ближнего Востока», Американский журнал генетики человека , 69 (5): 1095–1112, doi : 10.1086/324070 , PMC 1274378 , ПМИД 11573163
- Онофри, Валерио; Алессандрини, Федерика; Турки, Кьяра; Песарези, Мауро; Бушеми, Лоредана; Тальябраччи, Адриано (2006), «Разработка мультиплексных ПЦР для эволюционных и судебно-медицинских применений 37 SNP Y-хромосомы человека» (PDF) , Forensic Science International , 157 (1): 23–35, doi : 10.1016/j.forsciint.2005.03 .014 , PMID 15896936 [ постоянная мертвая ссылка ]
- Параккини; Пирс, CL; Колонель, Л.Н.; Альтшулер, Д; Хендерсон, Бельгия; Тайлер-Смит, К. (2003), «Влияние хромосомы AY на риск рака простаты: многоэтническое когортное исследование», J Med Genet , 40 (11): 815–819, doi : 10.1136/jmg.40.11.815 , PMC 1735314 , ПМИД 14627670
- Пелотти; Чеккарди, С; Лугареси, Ф; Трейн, Р; Фалькони, М; Бини, К; Уиллувейт, С; Рёвер, Л. (2007), «Микрогеографическая генетическая изменчивость Y-хромосомы в выборке населения района Равенны в регионе Эмилия-Романья (север Италии)», Forensic Science International: Genetics Supplement Series , 1 (1): 242–243 , doi : 10.1016/j.fsigss.2007.10.025
- Перейра, Луиза; Черный, Виктор; Сересо, Мария; Сильва, Нуно М; Гаек, Мартин; Вашикова, Альжбета; Куянова, Мартина; Брдичка, Радим; Салас, Антонио (2010), «Связывание генофондов стран к югу от Сахары и Западной Евразии: материнское и отцовское наследие кочевников-туарегов из африканского Сахеля» (PDF) , European Journal of Human Genetics , 18 (8): 915–923 , doi : 10.1038/ejhg.2010.21 , PMC 2987384 , PMID 20234393 , заархивировано из оригинала (PDF) 28 мая 2013 г.
- Перичич, М.; Лаук, LB; Кларич, И.М.; Рутси, С; Яничиевич, Б; Рудан, я; Терзич, Р; Чолак, я; и др. (2005), «Филогенетический анализ с высоким разрешением в Юго-Восточной Европе прослеживает основные эпизоды потока отцовских генов среди славянских популяций» , Mol. Биол. Эвол. , том. 22, нет. 10, стр. 1964–75, doi : 10.1093/molbev/msi185 , PMID 15944443 , заархивировано из оригинала 30 июня 2012 года .
- Понтикос Д. "Филогеографическое уточнение гаплогруппы Е" http://dienekes.blogspot.ru/2015/07/phylogeographic-refinement-of.html
- Рамос-Луиза, Э.; Бланко-Вереа, А.; Брион, М.; Ван Хаффель, В.; Карраседо, А.; Санчес-Дис, П. (2009), «Филогеография французских мужских линий (неопубликованные данные 23-го Международного конгресса ISFG)», Forensic Science International , 2 : 439–441, doi : 10.1016/j.fsigss.2009.09.026 , S2CID 85134429 [ мертвая ссылка ]
- Регейру, М.; Каденас, AM; Гайден, Т.; Андерхилл, Пенсильвания; Эррера, Р.Дж. (2006), «Иран: трехконтинентальная связь для миграции, вызванной Y-хромосомой», Hum Hered , 61 (3): 132–143, doi : 10.1159/000093774 , PMID 16770078 , S2CID 7017701
- Робино, К.; Кробу, Ф.; Гаэтано, К.; Бекада, А.; Бенхамамуш, С.; Черутти, Н.; Пьяцца, А.; Интурри, С.; Торре, К. (2008), «Анализ гаплогрупп SNP Y-хромосомы и гаплотипов STR в выборке населения Алжира», Journal International Journal of Legal Medicine , 122 (3): 251–5, doi : 10.1007/s00414-007- 0203-5 , ПМИД 17909833 , С2КИД 11556974
- Роза, Александра; Орнелас, Каролина; Джоблинг, Марк А; Брем, Антониу; Виллемс, Ричард (2007), «Y-хромосомное разнообразие населения Гвинеи-Бисау: многонациональная перспектива», BMC Evolutionary Biology , 7 : 124, Bibcode : 2007BMCEE...7..124R , doi : 10.1186/1471- 2148-7-124 , ПМЦ 1976131 , ПМИД 17662131
- Россер, З; Зержал, Т; Херлс, М; Адохаан, М; Алавантик, Д; Аморим, А; Амос, В; Арментерос, М; и др. (2000), «Разнообразие Y-хромосомы в Европе является клиническим и зависит в первую очередь от географии, а не от языка» , Американский журнал генетики человека , 67 (6): 1526–1543, doi : 10.1086/316890 , PMC 1287948 , PMID 11078479 , заархивировано из оригинала 6 мая 2008 г.
- Санчес, Хуан Дж; Халленберг, Шарлотта; Бёрстинг, Клаус; Эрнандес, Алексис; Горлин, Р.Дж. (2005), «Высокая частота линий Y-хромосомы, характеризующихся E3b1, DYS19-11, DYS392-12 у сомалийских мужчин», European Journal of Human Genetics , 13 (7): 856–866, doi : 10.1038/sj. .ejhg.5201390 , PMID 15756297 . Опубликовано в Интернете 9 марта 2005 г.
- Скоццари, Розарий; Кручиани, Ф; Панграцио, А; Сантоламазца, П; Видишь, Г; Мораль, П; латынь, В; Вареси, Л; и др. (2001), «Вариации Y-хромосомы человека в районе Западного Средиземноморья: последствия для заселения региона» ( PDF) , Human Immunology 62 ( 9) CiteSeerX 871–884 , : , /S0198-8859(01)00286- 5 , PMID 11543889 , заархивировано из оригинала (PDF) 17 декабря 2012 г. , получено 2 апреля 2023 г.
- Семино, О.; Пассарино, Дж; Оефнер, П.Дж.; Лин, А.А.; Арбузова С; Бекман, Ле; Де Бенедиктис, Дж; Франкалаччи, П; и др. (2000), «Генетическое наследие палеолитического Homo sapiens sapiens у современных европейцев: хромосомная перспектива AY» (PDF) , Science , vol. 290, нет. 5494, стр. 1155–59, Bibcode : 2000Sci...290.1155S , doi : 10.1126/science.290.5494.1155 , PMID 11073453 , заархивировано из оригинала (PDF) 25 ноября 2003 г.
- Семино; Сантакьяра-Бенерекетти, А. Сильвана; Фаласки, Франческо; Кавалли-Сфорца, Л. Лука; Андерхилл, Питер А. (2002), «Эфиопы и койсанцы разделяют самые глубокие клады филогении Y-хромосомы человека» (PDF) , Am J Hum Genet , vol. 70, нет. 1, стр. 265–268, doi : 10.1086/338306 , PMC 384897 , PMID 11719903 , заархивировано из оригинала (PDF) 15 марта 2006 г.
- Семино, Орнелла; Магри, Кьяра; Бенуцци, Джорджия; Лин, Алиса А.; Аль-Захери, Надя; Батталья, Винченца; Макчиони, Лилиана; Триантафиллидис, Костас; и др. (2004), «Происхождение, распространение и дифференциация гаплогрупп E и J Y-хромосомы: выводы о неолитизации Европы и более поздних миграционных событиях в Средиземноморье», American Journal of Human Genetics , vol. 74, нет. 5, стр. 1023–1034, номер документа : 10.1086/386295 , PMC 1181965 , PMID 15069642.
- Шен, Пейдун; Лави, Таль; Кивисилд, Тоомас; Чоу, Вивиан; Сенгун, Дениз; Гефель, Дов; Шпирер, Иссак; Вульф, Эйлон; и др. (2004), «Реконструкция отцовских и матрилиний самаритян и других израильских популяций на основе вариаций Y-хромосомы и митохондриальной ДНК» (PDF) , Human Mutation , vol. 24, нет. 3, стр. 248–60, doi : 10.1002/humu.20077 , PMID 15300852 , S2CID 1571356 , заархивировано из оригинала (PDF) 05 марта 2009 г. , получено 7 ноября 2008 г.
- Сильва; Карвалью, Элизеу; КОСТА, Гильерме; ТАВАРЕС, Лигия; АМОРИМ, Антониу; Гусмао, Леонор (2006), «Генетические вариации Y-хромосомы в популяции Рио-де-Жанейро» , Американский журнал человеческой биологии , 18 (6): 829–837, doi : 10.1002/ajhb.20567 , PMID 17039481 , S2CID 23778828 , в архиве. из оригинала от 18 октября 2012 г.
- Шлаш; Бехар, Дорон М.; Юдковский, Геннадий; Темплтон, Алан; Хадид, Ярин; Базис, Фуад; Хаммер, Майкл; Ицковиц, Шалев; Скорецки, Карл (2008), Геммелл, Нил Джон (редактор), «Друзы: популяционно-генетическое убежище Ближнего Востока», PLOS ONE , 3 (5): e2105, Bibcode : 2008PLoSO...3.2105S , doi : 10.1371/journal.pone.0002105 , PMC 2324201 , PMID 18461126
- Соле-Мората, Неус; Гарсиа-Фернандес, Карла; Урасин, Вадим; Бекада, Асмахан; Фадхлауи-Зид, Карима; Заллуа, Пьер; Комас, Дэвид; Калафель, Франческ (21 ноября 2017 г.), «Полные последовательности Y-хромосомы показывают чрезвычайно недавнее происхождение наиболее распространенной отцовской линии Северной Африки E-M183 (M81)», Scientific Reports , 7 (1): 15941, Bibcode : 2017NatSR. ..715941S , doi : 10.1038/s41598-017-16271-y , ISSN 2045-2322 , PMC 5698413 , PMID 29162904
- Сайкс, Брайан (2006), Кровь островов: исследование генетических корней нашей племенной истории , Bantam, ISBN 978-0-593-05652-3
- Тишкофф; Гондер; Хенн; Мортенсен; Найт (2007), «История говорящих по щелчку популяций Африки, выведенная на основе генетических вариаций мтДНК и Y-хромосомы» , Mol Biol Evol , 24 (10): 2180–2195, doi : 10.1093/molbev/msm155 , PMID 17656633 , заархивировано из оригинал от 10 октября 2011 г.
- Томас; Штумпф, магистр здравоохранения; Харк, Х. (2006), «Доказательства социальной структуры, подобной апартеиду, в ранней англосаксонской Англии» (PDF) , Proceedings of the Royal Society B , 273 (1601): 2651–2657, doi : 10.1098/rspb. 2006.3627 , PMC 1635457 , PMID 17002951 , заархивировано из оригинала (PDF) 5 марта 2009 г.
- Тромбетта, Бениамино; Кручиани, Фульвио; Селитто, Даниэле; Скоццари, Розария (2011), Маколей, Винсент (редактор), «Новая топология гаплогруппы E1b1 (E-P2) Y-хромосомы человека, выявленная с помощью недавно охарактеризованных бинарных полиморфизмов», PLOS ONE , 6 (1): e16073, Bibcode : 2011PLoSO...616073T , doi : 10.1371/journal.pone.0016073 , PMC 3017091 , PMID 21253605
- Тромбетта Б. «Филогеографическое уточнение и крупномасштабное генотипирование гаплогруппы E Y-хромосомы человека дают новое представление о расселении ранних скотоводов на африканском континенте» http://gbe.oxfordjournals.org/content/7/7/1940.long
- Андерхилл, Питер А.; Шен, Пейдун; Лин, Алиса А.; Джин, Ли; Пассарино, Джузеппе; Ян, Вэй Х.; Кауфман, Эрин; Бонне-Тамир, Батшева; и др. (2000), «Вариации последовательности Y-хромосомы и история человеческих популяций», Nat Genet , vol. 26, нет. 3, стр. 358–361, doi : 10.1038/81685 , PMID 11062480 , S2CID 12893406
- Андерхилл; Пассарино, Г.; Лин, А.А.; Шен, П.; Миразон Лар, М.; Фоли, РА; Оефнер, П.Дж.; Кавалли-Сфорца, LL (2001), «Филогеография бинарных гаплотипов Y-хромосомы и происхождение современных человеческих популяций», Ann Hum Genet , vol. 65, нет. Часть 1, стр. 43–62, doi : 10.1046/j.1469-1809.2001.6510043.x , PMID 11415522 , S2CID 9441236
- Андерхилл (2002), «Выводы об истории неолитического населения с использованием гаплотипов Y-хромосомы», в Беллвуде, Питер; Ренфрю, А. Колин (ред.), Исследование гипотезы расселения сельского хозяйства и языков , Кембридж: Институт археологических исследований Макдональда, ISBN. 978-1-902937-20-5
- Андерхилл, Питер А.; Кивисилд, Тоомас (2007), «Использование структуры популяции Y-хромосомы и митохондриальной ДНК для отслеживания миграций человека», Annu. Преподобный Жене. , 41 : 539–64, doi : 10.1146/annurev.genet.41.110306.130407 , PMID 18076332
- Уил, Мэн; Вайс, Д.А.; Ягер, РФ; Брэдман, Н.; Томас, М.Г. (2002), «Свидетельства Y-хромосомы о массовой миграции англосаксонцев» (PDF) , Mol. Биол. Эвол. , том. 19, нет. 7, стр. 1008–1021, doi : 10.1093/oxfordjournals.molbev.a004160 , PMID 12082121 , заархивировано из оригинала (PDF) 29 октября 2005 г.
- Уил; Шах, Т; Джонс, Алабама; Гринхал, Дж; Уилсон, Дж. Ф.; Нимадава, П; Зейтлин, Д; Коннелл, бакалавр; и др. (1 сентября 2003 г.), «Редкие линии Y-хромосомы с глубокими корнями у человека: уроки филогеографии» , Genetics , vol. 165, нет. 1, стр. 229–234, doi : 10.1093/genetics/165.1.229 , PMC 1462739 , PMID 14504230
- Древесина; Стовер; Эрет; Destro-Bisol (2005), «Контрастные закономерности вариаций Y-хромосомы и мтДНК в Африке: свидетельства демографических процессов, связанных с полом», European Journal of Human Genetics , 13 (7): 867–876, doi : 10.1038/sj.ejhg .5201408 , PMID 15856073
- Консорциум Y-хромосомы "YCC" (2002), "Номенклатурная система для дерева бинарных гаплогрупп Y-хромосомы человека", Genome Research , vol. 12, нет. 2, стр. 339–348, doi : 10.1101/gr.217602 , PMC 155271 , PMID 11827954
- Заллуа, Пьер А.; Сюэ, Яли; Халифе, Джейд; Махул, Надин; Дебиан, Лабиб; Платт, Дэниел Э.; Ройюру, Аджай К.; Эррера, Рене Дж.; и др. (2008), «Разнообразие Y-хромосомы в Ливане структурировано недавними историческими событиями», Американский журнал генетики человека , 82 (4): 873–882, doi : 10.1016/j.ajhg.2008.01.020 , PMC 2427286 , PMID 18374297
- Заллуа, Питер А.; Платт, Дэниел Э.; Сибай, Мирват; Халифе, Джейд; Махул, Надин; Хабер, Марк; Сюэ, Он был; Изабель, Хасан; и др. (2008), «Идентификация генетических следов исторической экспансии: финикийские следы в Средиземноморье», Американский журнал генетики человека , 83 (5): 633–642, doi : 10.1016/j.ajhg.2008.10.012 , PMC 2668035 . ПМИД 18976729
- Зержал (1999), Использование вариаций ДНК Y-хромосомы для изучения истории популяции; в Папиха С.С., Дека Р., Чакраборти Р. (ред.): Геномное разнообразие: применение в популяционной генетике человека , Kluwer Academic/Plenum Publishers, стр. 91–101.
- Чжао; Хан, Фейсал; Боркар, Минал; Эррера, Рене; Агравал, Суракша (2009), «Наличие трех разных отцовских линий среди жителей Северной Индии: исследование 560 Y-хромосом», Annals of Human Biology , 36 (1): 1–14, doi : 10.1080/03014460802558522 , PMC 2755252 , PMID 19058044
Источники таблиц преобразования
[ редактировать ]- Капелли, Кристиан; Уилсон, Джеймс Ф.; Ричардс, Мартин; Штумпф, Майкл П.Х.; и др. (февраль 2001 г.). «Отцовское наследие преимущественно коренных народов австронезийскоязычных народов островной Юго-Восточной Азии и Океании» . Американский журнал генетики человека . 68 (2): 432–443. дои : 10.1086/318205 . ПМЦ 1235276 . ПМИД 11170891 .
- Хаммер, Майкл Ф.; Карафет Татьяна М.; Редд, Алан Дж.; Джарджанази, Хамди; и др. (1 июля 2001 г.). «Иерархические закономерности глобального разнообразия Y-хромосом человека» . Молекулярная биология и эволюция . 18 (7): 1189–1203. doi : 10.1093/oxfordjournals.molbev.a003906 . ПМИД 11420360 .
- Каладжиева, Люба; Калафель, Франческ; Джоблинг, Марк А; Ангеличева, Дора; и др. (февраль 2001 г.). «Закономерности меж- и внутригруппового генетического разнообразия у влакс-рома, выявленные с помощью Y-хромосомы и линий митохондриальной ДНК» . Европейский журнал генетики человека . 9 (2): 97–104. дои : 10.1038/sj.ejhg.5200597 . ПМИД 11313742 . S2CID 21432405 .
- Карафет, Татьяна; Сюй, Липин; Ду, Руофу; Ван, Уильям; и др. (сентябрь 2001 г.). «История отцовского населения Восточной Азии: источники, закономерности и микроэволюционные процессы» . Американский журнал генетики человека . 69 (3): 615–628. дои : 10.1086/323299 . ПМЦ 1235490 . ПМИД 11481588 .
- Семино, О.; Пассарино, Дж; Оефнер, П.Дж.; Лин, А.А.; и др. (2000), «Генетическое наследие палеолитического Homo sapiens sapiens у современных европейцев: хромосомная перспектива AY», Science , 290 (5494): 1155–9, Бибкод : 2000Sci...290.1155S , doi : 10.1126/science.290.5494. 1155 , ПМИД 11073453
- Су, Бинг; Сяо, Цзюньхуа; Андерхилл, Питер; Дека, Ранджан; и др. (декабрь 1999 г.). «Y-хромосома свидетельствует о миграции современных людей на север в Восточную Азию во время последнего ледникового периода» . Американский журнал генетики человека . 65 (6): 1718–1724. дои : 10.1086/302680 . ПМЦ 1288383 . ПМИД 10577926 .
- Андерхилл, Питер А.; Шен, Пейдун; Лин, Алиса А.; Джин, Ли; и др. (ноябрь 2000 г.). «Вариации последовательности Y-хромосомы и история человеческих популяций». Природная генетика . 26 (3): 358–361. дои : 10.1038/81685 . ПМИД 11062480 . S2CID 12893406 .
Внешние ссылки
[ редактировать ]- E-M35 Phylogeny Project вики
- Проект филогении E-M35 (все лаборатории)
- Общественный форум проекта «Филогения E-M35»
- Еврейский проект E3b в FTDNA
- Карта распространения E-M35 в Европе
- Гаплогруппа E3B1 (E1b1b1) Группа Facebook (сообщество M35)
- E1b1b1b*-Проект
- E1b1.org - Международный проект Y-ДНК гаплогруппы E1b1 и ее субкладов. Архивировано 17 декабря 2014 г. в Wayback Machine.