Гаплогруппа I-M253
| Гаплогруппа I1 (M253) | |
|---|---|
| Возможное время происхождения | 3,170–4,600 [1] –5070 BP (сегодняшняя диверсификация) [2] [3] (ранее 11 000 BP [4] до 33 000 лет назад [5] ) 27 500 (диверсификация с помощью I2-FGC77992) [1] |
| Возможное место происхождения | Северная Европа [6] |
| Предок | Я* (М170) |
| Потомки | И1а (DF29/S438); I1b (S249/Z131); I1c (Y18119/Z17925) |
| Определение мутаций | M253, M307.2/P203.2, M450/S109, P30, P40, L64, L75, L80, L81, L118, L121/S62, L123, L124/S64, L125/S65, L157.1, L186, L187 |
Гаплогруппа I-M253 , также известная как I1 , представляет собой Y-хромосомы гаплогруппу . Генетическим маркером, идентифицирующим I-M253, являются SNP M253 , M307.2/P203.2, M450/S109, P30, P40, L64, L75, L80, L81, L118, L121/S62, L123, L124/S64, L125/S65, L157.1, L186 и L187. [7] Это основная ветвь гаплогруппы I-M170 (I*).
Считается, что гаплогруппа I1 присутствовала среди европейских охотников-собирателей верхнего палеолита как второстепенная линия, но из-за ее почти полного отсутствия в донеолитических образцах ДНК она не могла быть широко распространена. Образцы эпохи неолита I1 также очень редки, что позволяет предположить быстрое рассеивание, связанное с эффектом основателя в скандинавском бронзовом веке . Сегодня он достигает своего пика частоты в Швеции (52 процента мужчин в округе Вестра-Гёталанд ) и западной Финляндии (более 50 процентов в провинции Сатакунта ). [8] В среднем по стране I-M253 обнаруживается у 38–39% шведских мужчин. [9] [10] [8] 37% норвежских мужчин, [11] [12] [13] 34,8% датских мужчин, [14] [15] [16] 34,5% исландских мужчин, [17] [18] [19] и около 28% финских мужчин. [20] Брайан Сайкс в своей книге «Кровь островов » 2006 года дает членам – и условному патриарху-основателю I1 – имя « Водан ». [21]
Все известные ныне живущие члены происходят от общего предка, который в 6 раз моложе общего предка с I2. [1]
До реклассификации в 2008 г. [22] группа была известна как I1a , имя, которое с тех пор было присвоено первичной ветви, гаплогруппе I-DF29. Другими первичными ветвями I1 (M253) являются I1b (S249/Z131) и I1c (Y18119/Z17925).
Более 99% ныне живущих мужчин с I1 принадлежат к ветви DF29, которая, по оценкам, возникла в 2400 году до нашей эры. [23] [24] Все мужчины DF29 имеют общего предка, родившегося между 2500 и 2400 годами до нашей эры. [25] Самый старый найденный древний человек с I1-DF29 — Олл009, человек из Швеции раннего бронзового века . [26] [27]
Происхождение
[ редактировать ]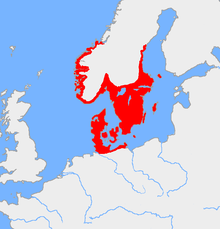
Хотя гаплогруппа I1, скорее всего, отошла от I* еще 27 000 лет назад в Граветте , до сих пор исследования ДНК смогли обнаружить ее только у трех палеолитических и мезолитических охотников-собирателей. По состоянию на ноябрь 2022 года только 6 древних образцов ДНК из человеческих останков, датируемых ранее скандинавского бронзового века, были отнесены к гаплогруппе I1:
- Охотник-собиратель из Азилиана в Испании в 11 466 году до н.э. классифицирован как имеющий ныне вымершую ветвь I-Z2699. [24] [28]
- Погребение SF11. Дата: 7500 лет назад. Первый — образец ДНК скандинавского охотника-собирателя с меткой SF11, найденный на Стора-Карлсё на Готланде . Было обнаружено, что SF11 несет 9 из 312 SNP, определяющих гаплогруппу I1. SF11 был классифицирован как I1-Z2699*. [29] [30] [31] [32] SF11 не был отнесен к конкретной археологической культуре, поскольку скелет был найден в пещере Стора Фёрвар на острове Стора Карлсё.
- Погребение BAB5. Дата: 7300-5900 лет назад. Второй представляет собой индивидуальный образец из Балатонсемеш-Багодомба с обозначением BAB5, из неолитической Венгрии . [33] Было обнаружено, что BAB5 несет 1 из 312 SNP, определяющих гаплогруппу I1. BAB5 также можно классифицировать как I1-Z2699*. [34] BAB5 имел генетическое родство с другими современными неолитическими земледельцами Центральной Европы .
- Захоронение RISE179. Дата: 4010–3776 гг. до н. э. – Кроме того, третий древний образец I1 принадлежит человеку, найденному в курганном захоронении, датируемом поздним неолитическим периодом кинжалов в Скандинавии, с обозначением RISE179. [35] Могила расположена недалеко от Аббекоса на южном побережье Сконе. RISE179 имел генетическое родство с популяциями культуры шнуровой керамики и культуры унетице . [35]
- Погребение oll009 Дата: 3930–3750 гг. до н.э. Четвертый древний образец I1, предшествовавший скандинавскому бронзовому веку (1700–500 гг. до н.э.), обозначен oll009 и был секвенирован в исследовании под названием «Геномное происхождение людей скандинавской культуры боевых топоров и их связь с более широкий горизонт шнуровой керамики». [36] Oll009 датируется скандинавским поздним неолитом и был найден в захоронении в Швеции недалеко от Олльсё на восточном побережье Сконе. Подобно RISE179, он имел высокий процент предков западных степных пастухов и имел генетическую близость к населению культуры боевых топоров и другим популяциям горизонта шнуровой керамики. [37] oll009 имеет Y11204, но, похоже, не имеет Y164553 или Y11205. [38]
Несмотря на высокую частоту гаплогруппы I1 у современных скандинавов, I1 полностью отсутствует среди ранних образцов ДНК земледельцев из неолитической Скандинавии. [39] [40] [31] (как и в случае с другими гаплогруппами по всей Европе). За исключением единственного образца ДНК (SF11), он также отсутствует у мезолитических охотников-собирателей в Скандинавии. I1 впервые начинает заметно чаще появляться в Скандинавии во время позднего неолита в связи с приходом в Скандинавию групп, несущих предков западностепных пастухов , но частота значительно не увеличивается до начала скандинавского бронзового века. [35] [41] [42]
Из-за очень небольшого количества древних образцов ДНК, отнесенных к I1 и датируемых ранее скандинавского бронзового века, в настоящее время неизвестно, присутствовал ли I1 в качестве редкой гаплогруппы среди скандинавских культур-собирателей, таких как костная посуда , прежде чем он был ассимилирован культура Боевого Топора , или если она была завезена в Скандинавию прибывшими группами, такими как Боевой Топор, которые, возможно, ассимилировали ее из таких культур, как культура Воронкообразных в Центральной Европе; или сама степь. Будущие исследования, скорее всего, смогут определить, какое из этих двух возможных источников имеет место.
Образцы SF11 и BAB5 отличаются от других образцов древней ДНК, отнесенных к I1, в том смысле, что они оба, похоже, представляют собой ныне вымершие ветви I1, которые еще не полностью развились в I-M253. Поэтому маловероятно, что они были предками современных носителей I1, которые все имели общего предка, жившего около 2600 г. до н.э.
Согласно исследованию, опубликованному в 2010 году, I-M253 возник между 3170 и 5000 лет назад в энеолитической Европе . [2] Новое исследование, проведенное в 2015 году, с использованием двух разных методов оценило происхождение от 3470 до 5070 лет назад или от 3180 до 3760 лет назад. [3]
В 2007 году было высказано предположение, что первоначально он расселился с территории нынешней Дании . [14] Однако профессор доктор Кеннет Нордтведт из Университета штата Монтана по поводу MRCA в 2009 году написал в личном сообщении: «Мы не знаем, где существовал этот человек, но большой бассейн нижней части Эльбы кажется сердцем I1».
Последние результаты (январь 2022 г.), опубликованные Y-Full , показывают, что I1 (I-M253) образовался 27 500 лет назад ( 95 CI : 29 800 лет назад - 25 200 лет назад) с TMRCA 4600 лет назад (95 CI: 5 200 лет назад - 4 000 лет назад). Поскольку самая последняя расчетная оценка TMRCA I1, как полагают, датируется примерно 2600 г. до н.э., это, вероятно, указывает на то, что предок всех ныне живущих людей I1 находится где-то в Северной Европе примерно в это время. Филогения I1 демонстрирует признаки быстрого звездообразного расширения. [43] [44] Это говорит о том, что I1 быстро превратился из редкого маркера в довольно распространенный. [3]
Структура
[ редактировать ]I-M253 ( M253, M307.2/P203.2, M450/S109, P30, P40, L64, L75, L80, L81, L118, L121/S62, L123, L124/S64, L125/S65, L157.1, L186 и L187 ) или I1 [7]
- И-DF29 (DF29/S438); I1a
- I-CTS6364 (CTS6364/Z2336); I1a1
- ФГК20030; Я1а1а~
- С4767; Я1а1а1~
- И-М227; я1а1а1а1а
- А394; I1a1a2~
- Y11221; I1a1a3~
- А5338; I1a1a4~
- С4767; Я1а1а1~
- CTS10028; I1a1b
- И-Л22 (Л22/С142); I1a1b1
- CTS11651/Z2338; I1a1b1a~
- И-П109; I1a1b1a1
- Я-Y3662; I1a1b1a1e~
- I-S14887; I1a1b1a1e2~
- Я-Y11203; I1a1b1a1e2d~
- Я-Y29630; I1a1b1a1e2d2~
- Я-Y11203; I1a1b1a1e2d~
- I-S14887; I1a1b1a1e2~
- Я-Y3662; I1a1b1a1e~
- CTS6017; I1a1b1a2
- И-Л205 (Л205.1/Л939.1/С239.1); I1a1b1a3
- CTS6868; I1a1b1a4
- И-З74; I1a1b1a4a
- CTS2208; I1a1b1a4a1~
- И-Л287; I1a1b1a4a1a
- И-Л258 (Л258/С335); I1a1b1a4a1a1
- И-Л287; I1a1b1a4a1a
- И-Л813; I1a1b1a4a2
- ФГК12562; I1a1b1a4a3~
- CTS2208; I1a1b1a4a1~
- И-З74; I1a1b1a4a
- И-П109; I1a1b1a1
- CTS11603/S4744; I1a1b1b~
- I-FT40464
- I-Y19934
- И-Л300 (Л300/С241); I1a1b1b1a1
- I-Y31032
- I-Y32014
- I-Y22918
- I-Y21972
- I-Y24013
- I-Y24015
- I-Y31032
- I-Y19933
- I-Y19932
- I-Y22015
- I-FT57000
- I-Y22015
- I-Y19932
- И-Л300 (Л300/С241); I1a1b1b1a1
- I-Y19934
- I-FT40464
- CTS11651/Z2338; I1a1b1a~
- ФГК10477/И13056; I1a1b2
- А8178, А8182, А8200, А8204; I1a1b3~
- Ф13534.2/И17263.2; I1a1b4~
- И-Л22 (Л22/С142); I1a1b1
- ФГК20030; Я1а1а~
- И-З58 (С244/З58); I1a2
- И-З59 (С246/З59); I1a2a
- И-З60 (С337/З60, С439/З61, З62); I1a2a1
- Я-Z140 (Z140, Z141)
- I-L338
- И-Ф2642 (Ф2642)
- Я-Z73
- I-L1302
- I-L573
- I-L803
- Я-Z140 (Z140, Z141)
- Я-Z382; I1a2a2
- И-З60 (С337/З60, С439/З61, З62); I1a2a1
- И-З138 (С296/З138, З139); I1a2b
- I-Z2541
- И-З59 (С246/З59); I1a2a
- И-З63 (С243/З63); I1a3
- I-BY151; I1a3a
- И-Л849.2; I1a3a1
- I-BY351; I1a3a2
- I-CTS10345
- I-Y10994
- I-Y7075
- I-CTS10345
- I-S2078
- I-S2077
- Я-Y2245 (Y2245/PR683)
- I-L1237
- Я-FGC9550
- I-S10360
- I-S15301
- I-Y7234
- I-L1237
- Я-Y2245 (Y2245/PR683)
- I-S2077
- I-BY62 (BY62); I1a3a3
- I-BY151; I1a3a
- I-CTS6364 (CTS6364/Z2336); I1a1
- И-З131 (З131/С249); I1b
- I-CTS6397 ; I1b1
- I-Z17943 (Y18119/Z17925, S2304/Z17937); I1c
Историческое расширение
[ редактировать ]
Гаплогруппа I1, а также субклады R1b, такие как R1b-U106, и субклады R1a, такие как R1a-Z284, тесно связаны с германскими народами и связаны с протогерманскими носителями нордического бронзового века . [45] [46] Текущие исследования ДНК показывают, что I1 практически не существовал в большей части Европы, за исключением Скандинавии и северной Германии, до периода миграции . Экспансия I1 напрямую связана с экспансией германских племен. Примерно с 900 г. до н.э. германские племена начали переселяться из южной Скандинавии и северной Германии в близлежащие земли между Эльбой и Одером. Между 600 и 300 гг. до н.э. другая волна германцев мигрировала через Балтийское море и поселилась вдоль Вислы . Переселение германцев в эту область привело к формированию культуры Вильбарк , связанной с готами . [47]
I1-Z63 был обнаружен в могильнике Ковалевко в Польше, датируемом римским железным веком . В 2017 году польские исследователи смогли успешно определить гаплогруппы YDNA 16 человек, похороненных на этом месте. Из этих 16 особей 8 принадлежали к I1. Что касается субкладов, то три принадлежали I-Z63, в частности субкладу I-L1237. [48] Археологический памятник Ковалевко был связан с культурой Вильбарк. Таким образом, субклад I-L1237 из I-Z63 можно рассматривать как генетический индикатор готского племени поздней античности. I1-Z63 также был найден в захоронении, связанном с останками готов и ломбардов в Колленьо, Италия. [49] [50] Кладбище датируется концом VI века, что позволяет предположить, что субклады I1-Z63 и нижележащие по течению связаны с миграциями готов в раннем средневековье.
В 2015 году в ходе исследования ДНК были проверены гаплогруппы Y-ДНК 12 образцов, датированных 300–400 годами нашей эры, из Саксонии-Анхальт в Германии. 8 из них принадлежали к гаплогруппе I1. Эти данные ДНК согласуются с историческими миграциями германских племен из Скандинавии в Центральную Европу. [51]
Кроме того, I1-Z63 был обнаружен на позднеантичном участке Crypta Balbi в Риме, на этот раз вместе с нижестоящим субкладом I-Y7234. [52] Материальные находки, связанные с лангобардами, были раскопаны в крипте Бальби.
Вилла Пла-де-л'Орта недалеко от Жироны в Испании расположена в непосредственной близости от некрополя с рядом гробниц, связанных с вестготами . Погребальный инвентарь и типология гробниц указывают на вестготское происхождение людей. В ходе исследования 2019 года у небольшого количества людей, похороненных на этом месте, были взяты образцы ДНК для анализа. Один из образцов принадлежал к гаплогруппе I1. [53] Этот вывод согласуется с общим предковым происхождением вестготов и остготов .
Англо -саксонское поселение в Британии представило I1 на Британских островах. [54]
В эпоху викингов I-M253 претерпел еще одно расширение. Маргарян и др. В 2020 году было проанализировано 442 человека из мира викингов из различных археологических памятников Европы. I-M253 была наиболее распространенной Y-гаплогруппой, обнаруженной в исследовании. Норвежские и датские викинги привезли больше I1 в Великобританию и Ирландию, а шведские викинги завезли его в Россию и Украину и привезли еще больше в Финляндию и Эстонию. [55]
Географическое распространение
[ редактировать ]Наибольшая плотность I-M253 встречается в Северной Европе и других странах, которые испытали обширную миграцию из Северной Европы либо в период миграции , в эпоху викингов , либо в наше время. Его можно найти во всех местах, захваченных скандинавами.
В современную эпоху значительные популяции I-M253 также укоренились в иммигрантских странах и бывших европейских колониях, таких как США , Австралия , Новая Зеландия и Канада .
| Население | Размер выборки | Я (всего) | И1 (И-М253) | И1а1а (И-М227) | Источник |
|---|---|---|---|---|---|
| Албанцы ( Тирана ) | 55 | 21.82%=(12/55) | 3.6%=(2/55) | 0.0 | Батталья и др. 2008 год |
| Албанцы ( Северная Македония ) | 64 | 17.2%=(11/64) | 4.7%=(3/64) | 0.0 | Батталья и др. 2008 год |
| Албанцы ( Тирана ) Албанцы ( Северная Македония ) | 55+64=119 | 19.33%=(23/119) | 4.2%=(5/119) | 0.0 | Батталья и др. 2008 год |
| Косовские албанцы ( Приштина ) | 114 | 7.96%=(9/114) | 5.31%=(6/114) | 0.0 | Перичич и др. 2005 г. |
| Албанцы ( Тирана ) Албанцы ( Северная Македония ) Косовские албанцы ( Приштина ) | 55+64+114=233 | 13.73%=(32/233) | 4.72%=(11/233) | 0.0 | Перичич и др. 2005 г. Батталья и др. 2008 год |
| Австрия | 43 | 9.3 | 2.3 | 0.0 | Андерхилл и др. 2007 год |
| Беларусь: Витбск | 100 | 15 | 1.0 | 0.0 | Андерхилл и др. 2007 год |
| Беларусь: Брест | 97 | 20.6 | 1.0 | 0.0 | Андерхилл и др. 2007 год |
| Босния | 100 | 42 | 2.0 | 0.0 | Шведский и др. 2004 г. |
| Болгария | 808 | 26.6 | 4.3 | 0.0 | Карачанак и др. 2013 год |
| Чешская Республика | 47 | 31.9 | 8.5 | 0.0 | Андерхилл и др. 2007 год |
| Чешская Республика | 53 | 17.0 | 1.9 | 0.0 | Шведский и др. 2004 г. |
| Дания | 122 | 39.3% (48/122) | 34.8% (40/122) | 0.0 | Андерхилл и др. 2007 год |
| Англия | 104 | 19.2 | 15.4 | 0.0 | Андерхилл и др. 2007 год |
| Эстония | 210 | 18.6 | 14.8 | 0.5 | Шведский и др. 2004 г. |
| Эстония | 118 | 11.9 | Лаппалайнен и др. 2008 год | ||
| Финляндия (национальный) | 28.0 | Лаппалайнен и др. 2006 г. | |||
| Финляндия: Запад | 230 | 40% (92/230) | Лаппалайнен и др. 2008 год | ||
| Финляндия: Восток | 306 | 19% (58/306) | Лаппалайнен и др. 2008 год | ||
| Финляндия: регион Сатакунта | 50+ | Лаппалайнен и др. 20089 г. | |||
| Франция | 58 | 17.2 | 8.6 | 1.7 | Андерхилл и др. 2007 год |
| Франция | 12 | 16.7 | 16.7 | 0.0 | Канн и др. 2002 г. |
| Франция (Нижняя Нормандия) | 42 | 21.4 | 11.9 | 0.0 | Шведский и др. 2004 г. |
| Германия | 125 | 24 | 15.2 | 0.0 | Андерхилл и др. 2007 год |
| Греция | 171 | 15.8 | 2.3 | 0.0 | Андерхилл и др. 2007 год |
| Венгрия | 113 | 25.7 | 13.3 | 0.0 | Шведский и др. 2004 г. |
| Ирландия | 100 | 11 | 6.0 | 0.0 | Андерхилл и др. 2007 год |
| Волжские татары | 53 | 13.2 | 11.3 | 0.0 | Трофимова 2015г. |
| Латвия | 113 | 3.5 | Лаппалайнен и др. 2008 год | ||
| Литва | 164 | 4.9 | Лаппалайнен и др. 2008 год | ||
| Нидерланды | 93 | 20.4 | 14 | 0.0 | Андерхилл и др. 2007 год |
| Норвегия | 1766 | 37% (653/1766) | Стенерсен и др. 2006 г. | ||
| Россия (национальный) | 16 | 25 | 12.5 | 0.0 | Канн и др. 2002 г. |
| Россия: Псков | 130 | 16.9 | 5.4 | 0.0 | Андерхилл и др. 2007 год |
| Россия: Кострома | 53 | 26.4 | 11.3 | 0.0 | Андерхилл и др. 2007 год |
| Россия: Смоленск | 103 | 12.6 | 1.9 | 0.0 | Андерхилл и др. 2007 год |
| Россия: Воронеж | 96 | 19.8 | 3.1 | 0.0 | Андерхилл и др. 2007 год |
| Россия: Архангельск | 145 | 15.8 | 7.6 | 0.0 | Андерхилл и др. 2007 год |
| Россия: Казаки | 89 | 24.7 | 4.5 | 0.0 | Андерхилл и др. 2007 год |
| Россия: Карелы | 140 | 10 | 8.6 | 0.0 | Андерхилл и др. 2007 год |
| Россия: Карелы | 132 | 15.2 | Лаппалайнен и др. 2008 год | ||
| Россия: Вепса | 39 | 5.1 | 2.6 | 0.0 | Андерхилл и др. 2007 год |
| Словакия | 70 | 14.3 | 4.3 | 0.0 | Шведский и др. 2004 г. |
| Словения | 95 | 26.3 | 7.4 | 0.0 | Андерхилл и др. 2007 год |
| Швеция (национальный) | 160 | 35.6% (57/160) | Лаппалайнен и др. 2008 год | ||
| Швеция (национальный) | 38.0 | Лаппалайнен и др. 2009 год | |||
| Швеция: Вестра Гёталанд | 52 | Лаппалайнен и др. 2009 год | |||
| Швейцария | 144 | 7.6 | 5.6 | 0.0 | Шведский и др. 2004 г. |
| Турция | 523 | 5.4 | 1.1 | 0.0 | Андерхилл и др. 2007 год |
| Украина: Львов | 101 | 23.8 | 4.9 | 0.0 | Андерхилл и др. 2007 год |
| Украина: Ivanovo-Frankiv | 56 | 21.4 | 1.8 | 0.0 | Андерхилл и др. 2007 год |
| Украина: Хмельниц | 176 | 26.2 | 6.1 | 0.0 | Андерхилл и др. 2007 год |
| Ukraine: Cherkasy | 114 | 28.1 | 4.3 | 0.0 | Андерхилл и др. 2007 год |
| Ukraine: Bilhorod | 56 | 26.8 | 5.3 | 0.0 | Андерхилл и др. 2007 год |
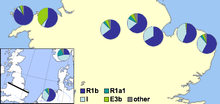
В 2002 году Майкл Э. Уил и его коллеги опубликовали статью, показывающую генетические доказательства популяционных различий между английским и валлийским населением, включая заметно более высокий уровень гаплогруппы I1 Y-ДНК в Англии, чем в Уэльсе. Они увидели в этом убедительное свидетельство массового вторжения англосаксов в восточную Великобританию из северной Германии и Дании в период миграции . [56] Авторы предположили, что популяции с большой долей гаплогруппы I1 произошли из северной Германии или южной Скандинавии, особенно из Дании, и что их предки мигрировали через Северное море вместе с англосаксонскими миграциями и датскими викингами . Основное утверждение исследователей заключалось в том,
что необходимо событие англосаксонской иммиграции, затронувшее 50–100% мужского генофонда Центральной Англии в то время. Однако мы отмечаем, что наши данные не позволяют нам отличить событие, которое просто пополнило генофонд мужчин коренных народов Центральной Англии, от события, когда мужчины из числа коренных народов были вытеснены в другие места, или от события, когда численность мужчин из числа коренных народов сократилась... Это исследование показывает что граница Уэльса была скорее генетическим барьером для потока генов англосаксонской Y-хромосомы, чем Северное море ... Эти результаты показывают, что политическая граница может быть более важной, чем геофизическая, в генетическом структурировании популяции.

В 2003 году Кристиан Капелли и его коллеги опубликовали статью, в которой поддержали, но изменили выводы Уила и его коллег. [57] В этой статье, в которой были отобраны образцы из Великобритании и Ирландии по сетке, была обнаружена меньшая разница между выборками из Уэльса и Англии с постепенным снижением частоты гаплогруппы I1, перемещающимся на запад на юге Великобритании. Результаты показали авторам, что норвежские викинги-завоеватели оказали сильное влияние на северную часть Британских островов, но что как английские, так и материковые шотландские образцы имеют немецко-датское влияние.
Выдающиеся члены I-M253
[ редактировать ]Путем прямого тестирования или тестирования их потомков и генеалогических данных было показано, что следующие известные люди являются I-M253:
- Александр Гамильтон , отец-основатель США . [58]
- Варяж Михаила Шимон , считающийся основателем русского дворянского рода Воронцовых (в том числе князя Семёновича Воронцова и Иллариона Ивановича Воронцова-Дашкова ), принадлежал к гаплогруппе I1-Y15024. [59] [60] [61] [62]
- Рюриковичский Владимира князь Святополк Окаянный (сын Великого ) принадлежал к I1-S2077. [63] [64] [65]
- Биргер Ярл , герцог Швеции Восточного Геатишского дома Бьяльбо , основатель Стокгольма ; его останки были эксгумированы и проверены в 2002 году и оказались I-M253. [66] Дом Бьяльбо также предоставил трех королей Норвегии и одного короля Дании в 14 веке.
- Британский музыкант Гордон Самнер, известный как Стинг [67]
- Уильям Брэдфорд , губернатор колонии Плимут [68]
- Уильям Брюстер , один из первых колонистов, эмигрировавших в Америку на корабле «Мэйфлауэр». [68]
- Генерал Конфедерации Роберт Э. Ли . Другими видными членами семьи Ли из Вирджинии и Мэриленда были Ричард Ли I и Ричард Генри Ли . [69]
- Роберт Брюс , король Шотландии, принадлежал к I1-Y17395. [70]
- Дом Гримальди принадлежит к скандинавскому субкладу I1, расположенному ниже по течению от I1-Y3549. [71]
- Президент США Эндрю Джексон . [72] [73]
- Русский писатель Лев Толстой . [74] [75]
- Исландский историк и поэт Снорри Стурлусон. [74]
- Шведский учёный и теолог Эмануэль Сведенборг [76] [77]
- Ренсбургский провидец , бурский патриотический деятель и мистик. [78] [79]
- Бьёрн Валроос , финский бизнесмен и миллионер. [74]
- Финский математик Рольф Неванлинна . [80] [81] [82]
- Американский изобретатель Сэмюэл Морзе . [83] [84] [85]
- Шведские футболисты Себастьян Ларссон и его отец Сванте Ларссон принадлежат к I1-Y24470. [86] [87] [88] [89]
- Шведский ютубер Феликс Чельберг (PewDiePie) принадлежит I1-L22. [90]
- Шведский актер Бьорн Андресен принадлежит к гаплогруппе I1-L22. [91] [92] [93] [94] Его предок Йохан Андресен жил по обе стороны шведско-норвежской границы. [95] [96]
- Американский актер Крис Пайн принадлежит к гаплогруппе I1-A13819. [97] [98]
- Шведский саамский хоккеист Бёрье Салминг . [99]
- Американский колонист Эдмунд Райс .
- Немецкий композитор Людвиг ван Бетховен . [100]
Маркеры
[ редактировать ]
Ниже приведены технические характеристики известных мутаций SNP и STR гаплогруппы I-M253.
Название: М253 [101]
- Тип: СНП
- Источник: М (Питер Андерхилл из Стэнфордского университета )
- Позиция: ChrY:13532101..13532101 (+ нить)
- Позиция (базовая пара): 283
- Общий размер (пар оснований): 400
- Длина: 1
- ИЗОГГ ХГ: I1
- Праймер F (вперед 5’→ 3’): GCAACAATGAGGGTTTTTTTG
- Праймер R (обратный 5'→ 3'): CAGCTCCACCTCTATGCAGTTT.
- YCC HG: I1
- Изменение аллелей нуклеотидов (мутация): C на T
Название: М307 [102]
- Тип: СНП
- Источник: М (Питер Андерхилл)
- Позиция: ChrY:21160339..21160339 (+ нить)
- Длина: 1
- ИЗОГГ ХГ: I1
- Праймер F: TTATTGGCATTTCAGGAAGTG
- Праймер R: GGGTGAGGCAGGAAAATAGC.
- YCC HG: I1
- Изменение аллелей нуклеотидов (мутация): G на A
Имя: P30 [103]
- Тип: СНП
- Источник: PS ( Майкл Хаммер из Университета Аризоны и Джеймс Ф. Уилсон из Эдинбургского университета)
- Позиция: ChrY:13006761..13006761 (+ нить)
- Длина: 1
- ИЗОГГ ХГ: I1
- Праймер F: GGTGGGCTTGTTTTGAAAAAGA
- Праймер R: AGCCAAATACCAGTCGTCAC
- YCC HG: I1
- Изменение аллелей нуклеотидов (мутация): G на A
- Регион: РСДП
Имя: P40 [104]
- Тип: СНП
- Источник: PS (Майкл Хаммер и Джеймс Ф. Уилсон)
- Позиция: ChrY:12994402..12994402 (+ нить)
- Длина: 1
- ИЗОГГ ХГ: I1
- Праймер F: GGAGAAAAGGTGAGAAACC
- Праймер R: GGACAAGGGGCAGATT.
- YCC HG: I1
- Изменение аллелей нуклеотидов (мутация): C на T
- Регион: РСДП
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с «I1 YTree» . Yfull.com. 06.04.2022 . Проверено 19 апреля 2022 г.
- ^ Jump up to: а б Педро Соарес, Алессандро Ачилли, Орнелла Семино, Уильям Дэвис, Винсент Маколей, Ханс-Юрген Бандельт, Антонио Торрони и Мартин Б. Ричардс, Археогенетика Европы, Современная биология , том. 20 (23 февраля 2010 г.), С174–Р183. yДНК Гаплогруппа I: Субклад I1 , Генеалогическое древо ДНК,
- ^ Jump up to: а б с Батини С., Халласт П., Задик Д., Дельсер П.М., Бенаццо А., Гиротто С. и др. (май 2015 г.). «Крупномасштабное недавнее расширение европейских отцов, продемонстрированное изменением последовательности населения» . Природные коммуникации . 6 : 7152. Бибкод : 2015NatCo...6.7152B . дои : 10.1038/ncomms8152 . ПМЦ 4441248 . ПМИД 25988751 .
- ^ Рутси С., Магри С., Кивисилд Т., Бенуцци Г., Хелп Х., Бермишева М. и др. (июль 2004 г.). «Филогеография гаплогруппы I Y-хромосомы выявляет отдельные области доисторического потока генов в Европе» (PDF) . Американский журнал генетики человека . 75 (1): 128–37. дои : 10.1086/422196 . ПМК 1181996 . ПМИД 15162323 . Архивировано из оригинала (PDF) 24 июня 2009 г. Проверено 20 марта 2008 г.
- ^ Андерхилл П.А., Майрес Н.М., Рутси С., Чоу К.Т., Лин А.А., Отиллар Р.П. и др. (2007). «Новые филогенетические взаимоотношения гаплогруппы I Y-хромосомы: переоценка ее филогеографии и предыстории». В Мелларс П., Бойл К., Бар-Йосеф О., Стринг С. (ред.). Переосмысление человеческой революции . Кембридж, Великобритания: Монографии Института Макдональда. стр. 33–42. ISBN 978-1-902937-46-5 .
- ^ Лаппалайнен, Т.; Лайтинен, В.; Салмела, Э.; Андерсен, П.; Хуопонен, К.; Савонтаус, М.-Л.; Лаэрмо, П. (2008). «Миграционные волны в регион Балтийского моря» . Анналы генетики человека . 72 (3): 337–348. дои : 10.1111/j.1469-1809.2007.00429.x . ПМИД 18294359 . S2CID 32079904 .
- ^ Jump up to: а б ISOGG, Гаплогруппа I Y-ДНК и ее субклады – 2017 г. (31 января 2017 г.).
- ^ Jump up to: а б Лаппалайнен Т., Лайтинен В., Салмела Э., Андерсен П., Хуопонен К., Савонтаус М.Л., Лаэрмо П. (май 2008 г.). «Миграционные волны в регион Балтийского моря» . Анналы генетики человека . 72 (Часть 3): 337–348. дои : 10.1111/j.1469-1809.2007.00429.x . ПМИД 18294359 . S2CID 32079904 .
- ^ Лаппалайнен Т., Ханнелиус У., Салмела Е., фон Дёбельн У., Линдгрен К.М., Хуопонен К. и др. (январь 2009 г.). «Структура населения в современной Швеции - анализ Y-хромосомной и митохондриальной ДНК». Анналы генетики человека . 73 (1): 61–73. дои : 10.1111/j.1469-1809.2008.00487.x . ПМИД 19040656 . S2CID 205598345 .
- ^ «FamilyTreeDNA – ПРОЕКТ ДНК Швеции – Sverigeprojektet» .
- ^ Дюпюи Б.М., Стенерсен М., Лу Т.Т., Олайсен Б. (декабрь 2006 г.). «Географическая гетерогенность Y-хромосомных линий в Норвегии». Международная судебно-медицинская экспертиза . 164 (1): 10–19. doi : 10.1016/j.forsciint.2005.11.009 . ПМИД 16337760 .
- ^ «FamilyTreeDNA – Норвежский проект ДНК – Norgesprosjektet» . Familytreedna.com . Проверено 26 ноября 2020 г.
- ^ «Y-ДНК Гаплогруппер» . ДНК Норвегии Norgesprosjektet . Проверено 27 декабря 2020 г.
- ^ Jump up to: а б Питер А. Андерхилл и др., Новые филогенетические отношения для гаплогруппы I Y-хромосомы: переоценка ее филогеографии и предыстории, в «Переосмыслении человеческой революции» (2007), стр. 33–42. П. Мелларс, К. Бойл, О. Бар-Йосеф, К. Стрингер (ред.) Институт археологических исследований Макдональда, Кембридж, Великобритания.
- ^ Санчес Джей-Джей (2004). «Гаплогруппы SNP Y-хромосомы у датчан, гренландцев и сомалийцев» (PDF) . Серия международных конгрессов . 1261 : 347–49. doi : 10.1016/S0531-5131(03)01635-2 – через isfg.org.
- ^ «FamilyTreeDNA – Проект ДНК Дании» . Familytreedna.com . Проверено 10 декабря 2020 г.
- ^ Хельгасон А., Сигурет Ардоттир С., Николсон Дж., Сайкс Б., Хилл Э.В., Брэдли Д.Г. и др. (сентябрь 2000 г.). «Оценка скандинавского и гэльского происхождения мужчин-поселенцев Исландии» . Американский журнал генетики человека . 67 (3): 697–717. дои : 10.1086/303046 . ПМЦ 1287529 . ПМИД 10931763 .
- ^ «Региональный проект Y-ДНК Больших Северных стран» . семьятридна . апрель 2021 г.
- ^ Эбенесерсдоттир С.С., Сандовал-Веласко М., Гуннарсдоттир Э.Д., Джагадисан А., Гудмундсдоттир В.Б., Тордардоттир Э.Л. и др. (июнь 2018 г.). «Древние геномы из Исландии раскрывают формирование человеческой популяции» . Наука . 360 (6392): 1028–1032. Бибкод : 2018Sci...360.1028E . дои : 10.1126/science.aar2625 . hdl : 10852/71890 . ПМИД 29853688 .
- ^ Лаппалайнен Т., Койвумяки С., Салмела Э., Хуопонен К., Систонен П., Савонтаус М.Л., Лаэрмо П. (июль 2006 г.). «Региональные различия среди финнов: взгляд на Y-хромосому». Джин . 376 (2): 207–15. дои : 10.1016/j.gene.2006.03.004 . ПМИД 16644145 .
- ^ «Кровь островов: исследование генетических корней нашей племенной истории» . История Ирландии . 22 февраля 2013 г. Проверено 10 декабря 2020 г.
- ^ Карафет Т.М., Мендес Ф.Л., Мейлерман М.Б., Андерхилл П.А., Зегура С.Л., Хаммер М.Ф. (май 2008 г.). «Новые бинарные полиморфизмы изменяют форму и увеличивают разрешение дерева гаплогрупп Y-хромосомы человека» . Геномные исследования . 18 (5): 830–38. дои : 10.1101/гр.7172008 . ПМК 2336805 . ПМИД 18385274 .
- ^ Батини С., Халласт П., Задик Д., Дельсер П.М., Бенаццо А., Гиротто С. и др. (май 2015 г.). «Крупномасштабное недавнее расширение европейских отцов, продемонстрированное изменением последовательности населения» . Природные коммуникации . 6 (1): 7152. Бибкод : 2015NatCo...6.7152B . дои : 10.1038/ncomms8152 . ПМЦ 4441248 . ПМИД 25988751 .
- ^ Jump up to: а б «Добро пожаловать в FamilyTreeDNA Discover (бета)» . FamilyTreeDNA Discover (бета) . Проверено 25 декабря 2022 г.
- ^ «I-DF29 YTree» . yfull.com . Проверено 25 декабря 2022 г.
- ^ Мальмстрем Х., Гюнтер Т., Свенссон Э.М., Юрас А., Фрейзер М., Мунтерс А.Р. и др. (октябрь 2019 г.). «Геномное происхождение людей скандинавской культуры боевых топоров и их связь с более широким горизонтом шнуровой керамики» . Слушания. Биологические науки . 286 (1912): 20191528. doi : 10.1098/rspb.2019.1528 . ПМК 6790770 . ПМИД 31594508 .
- ^ «I-Y11204 YДерево» . yfull.com . Проверено 25 декабря 2022 г.
- ^ «Haplotree.info – Ancientdna.info. Карта на основе All Ancient DNA v. 2.07.26» .
- ^ Гюнтер Т. (2017). «Геномика мезолитической Скандинавии раскрывает пути колонизации и адаптацию к высоким широтам» (PDF) . Природа . 23 :6 – через Биорксив.
- ^ SF11 – Стора Фёрвар, Стора Карлсё I-Z2699*. «Пример Haplotree.info: SF11» . haplotree.info .
{{cite web}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Jump up to: а б Скоглунд П., Мальмстрем Х., Омрак А., Рагхаван М., Вальдиозера С., Гюнтер Т. и др. (май 2014 г.). «Геномное разнообразие и примеси различаются у скандинавских собирателей и фермеров каменного века» . Наука . 344 (6185): 747–50. Бибкод : 2014Sci...344..747S . дои : 10.1126/science.1253448 . ПМИД 24762536 . S2CID 206556994 .
- ^ "familytreedna.com дерево I-Z2699" . семьятридна . апрель 2021 г.
- ^ Сечени-Надь А., Брандт Г., Хаак В., Кирл В., Якуч Дж., Мёллер-Рикер С. и др. (апрель 2015 г.). «Отслеживание генетического происхождения первых фермеров Европы позволяет лучше понять их социальную организацию» . Слушания. Биологические науки . 282 (1805). дои : 10.1098/rspb.2015.0339 . ПМЦ 4389623 . ПМИД 25808890 .
- ^ «BAB5 I-Z2699*» . haplotree.info .
- ^ Jump up to: а б с Аллентофт М.Э., Сикора М., Шегрен К.Г., Расмуссен С., Расмуссен М., Стендеруп Дж. и др. (июнь 2015 г.). «Популяционная геномика Евразии бронзового века» . Природа . 522 (7555): 167–72. Бибкод : 2015Natur.522..167A . дои : 10.1038/nature14507 . ПМИД 26062507 . S2CID 4399103 .
- ^ «YFull | Геномное происхождение людей скандинавской культуры боевых топоров и их связь с более широким горизонтом шнуровой керамики» . yfull.com . Проверено 24 января 2021 г.
- ^ Мальмстрем Х (июль 2019 г.). «Геномное происхождение людей скандинавской культуры боевых топоров и их связь с более широким горизонтом шнуровой керамики» (PDF) . Уппсала Геномикс . 1 :3 – через jakobssonlab.iob.uu.se/.
- ^ «I-Y11204 YДерево» .
- ^ Санчес-Куинто Ф., Мальмстрем Х., Фрейзер М., Гирдланд-Флинк Л., Свенссон Э.М., Симойнс Л.Г. и др. (май 2019 г.). «Мегалитические гробницы в западной и северной неолитической Европе были связаны с родственным обществом» . Труды Национальной академии наук Соединенных Штатов Америки . 116 (19): 9469–74. Бибкод : 2019PNAS..116.9469S . дои : 10.1073/pnas.1818037116 . ПМК 6511028 . ПМИД 30988179 .
- ^ Мальмстрем Х., Линдерхольм А., Скоглунд П., Стора Дж., Сьёдин П., Гилберт М.Т. и др. (январь 2015 г.). «Древняя митохондриальная ДНК с северной окраины неолитической экспансии земледелия в Европе проливает свет на процесс расселения» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 370 (1660): 20130373. doi : 10.1098/rstb.2013.0373 . ПМЦ 4275881 . ПМИД 25487325 .
- ^ Карлссон А.О., Валлерстрем Т., Гётерстрем А., Холмлунд Г. (август 2006 г.). «Разнообразие Y-хромосомы в Швеции – долгосрочная перспектива» . Европейский журнал генетики человека . 14 (8): 963–70. дои : 10.1038/sj.ejhg.5201651 . ПМИД 16724001 . S2CID 23227271 .
- ^ Мальмстрем Х., Гюнтер Т., Свенссон Э.М., Юрас А., Фрейзер М., Мунтерс А.Р. и др. (октябрь 2019 г.). «Геномное происхождение людей скандинавской культуры боевых топоров и их связь с более широким горизонтом шнуровой керамики» . Слушания. Биологические науки . 286 (1912): 20191528. doi : 10.1098/rspb.2019.1528 . ПМК 6790770 . ПМИД 31594508 .
- ^ Позник Г.Д., Сюэ Ю., Мендес Ф.Л., Виллемс Т.Ф., Массайя А., Уилсон Сейрес М.А. и др. (июнь 2016 г.). «Периодические всплески демографии мужского пола, выявленные на основе 1244 последовательностей Y-хромосомы по всему миру» . Природная генетика . 48 (6): 593–99. дои : 10.1038/ng.3559 . hdl : 11858/00-001M-0000-002A-F024-C . ПМЦ 4884158 . ПМИД 27111036 .
- ^ Вудли М. (февраль 2017 г.). «Голоценовый отбор вариантов, связанных с когнитивными способностями: сравнение древних и современных геномов» (PDF) . biorxiv.org/ . дои : 10.1101/109678 . S2CID 196631764 . Проверено 27 января 2021 г.
- ^ Шмидт К.Х. (1991). «Кельты и этногенез германского народа» . Historische Sprachforschung/Историческая лингвистика . 104 (1): 129–52. ISSN 0935-3518 . JSTOR 40849016 .
- ^ Бергербрант С. (май 2007 г.). «Идентичность бронзового века: костюмы, конфликты и контакты в Северной Европе 1600–1300 гг. До н.э.» (PDF) . Стокгольмские исследования по археологии . 43 : 7–201 – через diva-portal.org.
- ^ Теска М, Михаловски А (2014). «Связь Великопольши с регионом Балтийского моря в римском железе» . Археология Литвы . 14 : 63–77. дои : 10.15388/ArchLit.2013.0.2641 . S2CID 56295624 .
- ^ Зенчак М, Хандшу Л, Юрас А, Марцинковска-Свояк М, Филипс А, Пионтек Дж, Столарек И, Фиглерович М (2017). Присвоение гаплогруппы Y-хромосомы посредством секвенирования следующего поколения обогащенных библиотек древней ДНК . Антропологическая генетика. п. Номер презентации: АГ 16.
- ^ Аморим CE, Вай С., Пост С., Моди А., Конц И., Хакенбек С. и др. (сентябрь 2018 г.). «Понимание социальной организации и миграции варваров VI века с помощью палеогеномики» . Природные коммуникации . 9 (1): 3547. Бибкод : 2018NatCo...9.3547A . дои : 10.1038/s41467-018-06024-4 . ПМК 6134036 . ПМИД 30206220 .
- ^ Эстес Р. (16 октября 2020 г.). «Древняя ДНК лонгобардов из Паннонии и Италии – о чем нам говорит их ДНК? Вы родственники?» . DNAeXplained – Генетическая генеалогия . Проверено 11 декабря 2020 г.
- ^ Лабудд Д. (июль 2015 г.). «Гендерное распределение находок при раскопках периода Римской империи и миграции» . Исследовательские ворота . 1 :2 – через ResearchGate.net.
- ^ Антонио М.Л., Гао З., Мутс Х.М., Луччи М., Кандилио Ф., Сойер С. и др. (ноябрь 2019 г.). «Древний Рим: генетический перекресток Европы и Средиземноморья» . Наука . 366 (6466): 708–14. Бибкод : 2019Sci...366..708A . дои : 10.1126/science.aay6826 . ПМК 7093155 . ПМИД 31699931 .
- ^ Олальде И., Маллик С., Паттерсон Н., Роланд Н., Вильяльба-Моуко В., Сильва М. и др. (март 2019 г.). «Геномная история Пиренейского полуострова за последние 8000 лет» . Наука . 363 (6432): 1230–34. Бибкод : 2019Sci...363.1230O . дои : 10.1126/science.aav4040 . ПМК 6436108 . ПМИД 30872528 .
- ^ Мартиниано Р., Каффелл А., Холст М., Хантер-Манн К., Монтгомери Дж., Мюльднер Г. и др. (январь 2016 г.). «Геномные сигналы миграции и преемственности в Британии до англосаксов» . Природные коммуникации . 7 (1): 10326. Бибкод : 2016NatCo...710326M . дои : 10.1038/ncomms10326 . ПМЦ 4735653 . ПМИД 26783717 .
- ^ Маргарян А., Лоусон Д.Д., Сикора М., Расимо Ф., Расмуссен С., Мольтке И. и др. (сентябрь 2020 г.). «Популяционная геномика мира викингов». Природа . 585 (7825): 390–96. Бибкод : 2020Natur.585..390M . bioRxiv 10.1101/703405 . дои : 10.1038/s41586-020-2688-8 . hdl : 10852/83989 . ПМИД 32939067 . S2CID 201195157 .
- ^ Уил М.Э., Вайс Д.А., Джагер Р.Ф., Брэдман Н., Томас М.Г. (июль 2002 г.). «Y-хромосома свидетельствует о массовой миграции англосаксов» . Молекулярная биология и эволюция . 19 (7): 1008–21. doi : 10.1093/oxfordjournals.molbev.a004160 . ПМИД 12082121 .
- ^ Капелли С., Редхед Н., Абернети Дж.К., Гратрикс Ф., Уилсон Дж.Ф., Моен Т. и др. (май 2003 г.). «Перепись хромосом AY на Британских островах» . Современная биология . 13 (11): 979–84. дои : 10.1016/S0960-9822(03)00373-7 . hdl : 20.500.11820/8acb01f3-a7c1-45f5-89de-b796266d651e . ПМИД 12781138 . S2CID 526263 .
- ^ «ДНК отца-основателя» . isogg.org .
- ^ «FamilyTreeDNA – Генетическое тестирование на предмет происхождения, семейной истории и генеалогии» . Familytreedna.com . Проверено 10 декабря 2020 г.
- ^ «Фадерслинжен, ДНК» . sikaby.se . Проверено 10 декабря 2020 г.
- ^ «FamilyTreeDNA – RussiaDNA Project» . Familytreedna.com . Проверено 10 декабря 2020 г.
- ^ «Наш вождь викингов на востоке» . sikaby.se . Проверено 10 декабря 2020 г.
- ^ «Образец Homo sapiens – BioSample – NCBI» . ncbi.nlm.nih.gov . Проверено 10 декабря 2020 г.
- ^ Маргарян А., Лоусон Д.Д., Сикора М., Расимо Ф., Расмуссен С., Мольтке И. и др. (сентябрь 2020 г.). «Популяционная геномика мира викингов» . Природа . 585 (7825): 390–96. Бибкод : 2020Natur.585..390M . дои : 10.1038/s41586-020-2688-8 . hdl : 10852/83989 . ПМИД 32939067 . S2CID 221769227 .
- ^ Дучко В (2004). Русь викингов: Исследования присутствия скандинавов в Восточной Европе . Брилл. ISBN 978-90-04-13874-2 .
- ^ Мальмстрем Х., Вретемарк М., Тиллмар А., Дурлинг М.Б., Скоглунд П., Гилберт М.Т. и др. (январь 2012 г.). «В поисках основателя Стокгольма - исследование родства на основе Y-хромосомной, аутосомной и митохондриальной ДНК». Анналы анатомии – Anatomischer Anzeiger . Специальный выпуск: Древняя ДНК. 194 (1): 138–45. дои : 10.1016/j.aanat.2011.03.014 . ПМИД 21596538 .
- ^ Британское вторжение обретает свои корни
- ^ Jump up to: а б «Проект ДНК Мэйфлауэра» . mayflowerdna.org . Проверено 23 ноября 2020 г.
- ^ «FamilyTreeDNA - Проект исследования ДНК фамилии Ли (и Ли, Леа и т. Д.)» . Familytreedna.com . Проверено 10 декабря 2020 г.
- ^ «СемейноедеревоДНК – I1-S4795» . Familytreedna.com . Проверено 10 декабря 2020 г.
- ^ «FamilyTreeDNA - Генеалогический проект Гримальди в FtDNA» . Familytreedna.com . Проверено 11 декабря 2020 г.
- ^ «Проект ДНК Джексона» . СемейноеДревоДНК . 11 декабря 2020 г.
- ^ Хэй М (июль 2020 г.). «Происхождение и история гаплогруппы I1 (Y-ДНК)» . Исследовательские ворота . 1 :9.
- ^ Jump up to: а б с Масиамо Э. «Гаплогруппа I1 (Y-ДНК)» . Евпедия . Проверено 11 декабря 2020 г.
- ^ Петр Толстой. Моя родословная. Выпуск от 18.04.2010 (in Russian) , retrieved 2020-12-15
- ^ "I-BY229 YДерево" . yfull.com . Проверено 10 декабря 2020 г.
- ^ «Сведенборг» . Хойен (на шведском языке). Архивировано из оригинала 26 октября 2020 г. Проверено 10 декабря 2020 г.
- ^ «Клаас Янс ван Ренсбург, SV/PROG» . geni_family_tree . Проверено 03 января 2021 г.
- ^ «Янсе / Янсен ван Ренсбург I-M253, обсуждение генеалогии» . geni_family_tree . Проверено 03 января 2021 г.
- ^ «Рольф Х. Неванлинна» . geni_family_tree . Проверено 26 декабря 2020 г.
- ^ Оленус (30 марта 2018 г.). «I1: Рольф Неванлинна (урожденный Неовиус)» . Потомки гаплогруппы IJ-M429 . Проверено 26 декабря 2020 г.
- ^ «Арне Эдвард Неванлинна» . geni_family_tree . Проверено 26 декабря 2020 г.
- ^ «Тестирование ДНК Морса/Мосса» . morsesociety.org . Проверено 10 декабря 2020 г.
- ^ «FamilyTreeDNA – Проект ДНК Морса/Мосса» . Familytreedna.com . Проверено 10 декабря 2020 г.
- ^ «Генеалогическое древо Питера Морса» . geni.com . Проверено 10 декабря 2020 г.
- ^ «FamilyTreeDNA – Шведский проект ДНК – Sverigeprojektet» . Familytreedna.com . Проверено 14 февраля 2021 г.
- ^ «Муниципалитет Эскильстуны · EM GN398 – Семья Ларссон, Торшелла, около 1900 г.» . Муниципалитет Эскильстуна (на шведском языке) . Проверено 14 февраля 2021 г.
- ^ "I-Y24470 YTree" . yfull.com . Проверено 14 февраля 2021 г.
- ^ «Предки семьи Ларссон» . hosserudkullen.se . Проверено 14 февраля 2021 г.
- ^ Архивировано в Ghostarchive и Wayback Machine : «Я сделал тест ДНК… (Думаю, меня отменили)» – через YouTube .
- ^ «FamilyTreeDNA – Норвежский проект ДНК – Norgesprosjektet» . Familytreedna.com . Проверено 2 февраля 2021 г.
- ^ Товсет А (июнь 2018 г.). «Семейное исследование Андрисена, Фернскога и Хансена» . Kjellivar.tripod.com . Проверено 2 февраля 2021 г.
- ^ «Люди с именем Андресен | Släktingar.se» . мясники.se . Проверено 2 февраля 2021 г.
- ^ «Бьорн Андресен: Моя страсть к матери так и не получила ответа – встречается Катарина Хар» . Радио Швеции (на шведском языке) . Проверено 2 февраля 2021 г.
- ^ «Йохан Петер Андресен – Родословная» . предки.se . Проверено 2 февраля 2021 г.
- ^ «Генеалогическое древо Даниэля Андресена» . Гененет . Проверено 2 февраля 2021 г.
- ^ «FamilyTreeDNA - Проект ДНК генеалогии Пайн/Пайн» . Familytreedna.com . Проверено 10 декабря 2020 г.
- ^ «Джеймс Пайн-старший» . geni_family_tree . Проверено 10 декабря 2020 г.
- ^ Янлинд Ф (20 февраля 2021 г.). «Бьянка Салминг о своих отношениях с Бёрье: «Чувствую себя ужасно » [Бьянка Салминг о своих отношениях с Бёрье: «Чувствую себя ужасно»]. Goteborgs-Posten (на шведском языке).
- ^ Мерседес (26 марта 2023 г.). «Открытие ДНК Бетховена – выясните, являетесь ли вы родственниками» . Из кого ты сделан? . Проверено 7 октября 2023 г.
- ^ snpdev. «Отчет кластера эталонного SNP (refSNP): rs9341296» . nih.gov .
- ^ snpdev. «Отчет кластера эталонного SNP (refSNP): rs13447354» . nih.gov .
- ^ С30 [ постоянная мертвая ссылка ]
- ^ С40 [ постоянная мертвая ссылка ]
Дальнейшее чтение
[ редактировать ]- Аллентофт М.Э., Сикора М., Шегрен К.Г., Расмуссен С., Расмуссен М., Стендеруп Дж. и др. (июнь 2015 г.). «Популяционная геномика Евразии бронзового века» . Природа . 522 (7555): 167–72. Бибкод : 2015Natur.522..167A . дои : 10.1038/nature14507 . ПМИД 26062507 . S2CID 4399103 .
- Брюнель С., Беннетт Э.А., Карден Л., Гарро Д., Барран Эмам Х., Бейлиер А. и др. (июнь 2020 г.). «Древние геномы современной Франции раскрывают 7000-летнюю ее демографическую историю» . Труды Национальной академии наук Соединенных Штатов Америки . 117 (23): 12791–98. Бибкод : 2020PNAS..11712791B . дои : 10.1073/pnas.1918034117 . ПМЦ 7293694 . ПМИД 32457149 .
- Мальмстрем Х., Гилберт М.Т., Томас М.Г., Брандстрем М., Стора Дж., Молнар П. и др. (ноябрь 2009 г.). «Древняя ДНК показывает отсутствие преемственности между неолитическими охотниками-собирателями и современными скандинавами» . Современная биология . 19 (20): 1758–62. дои : 10.1016/j.cub.2009.09.017 . ПМИД 19781941 . S2CID 9487217 .
- Мальмстрём Х., Гюнтер Т., Свенссон Э.М., Юрас А., Фрейзер М., Мунтерс А.Р., Поспешны Л., Тырв М., Линдстрем Й., Гётерстрем А., Стора Й., Якобссон М. (октябрь 2019 г.). «Геномное происхождение людей скандинавской культуры боевых топоров и их связь с более широким горизонтом шнуровой керамики» . Слушания. Биологические науки . 286 (1912): 20191528. doi : 10.1098/rspb.2019.1528 . ПМК 6790770 . ПМИД 31594508 .
- Вильяльба-Моуко В., ван де Лоосдрехт М.С., Пост С., Мора Р., Мартинес-Морено Дж., Рохо-Гуэрра М. и др. (апрель 2019 г.). «Сохранение предков охотников-собирателей позднего плейстоцена на Пиренейском полуострове» . Современная биология . 29 (7). Cell Press : 1169–77.e7. дои : 10.1016/j.cub.2019.02.006 . hdl : 10261/208851 . ПМИД 30880015 . S2CID 76663708 .
Внешние ссылки
[ редактировать ]- Базы данных гаплогруппы I
- Проект гаплогруппы I1 в FTDNA
- Датский региональный проект ДНК Демес в FTDNA
- Проект гаплогруппы I-P109
- Проект ДНК Британских островов
- Общие базы данных Y-ДНК
Существует несколько баз данных общего доступа, в которых используется I-M253, в том числе:
- [1] Архивировано 4 января 2011 г. на Wayback Machine.
- YHRD: Справочная база данных гаплотипов STR Y-хромосомы
- I1 YДерево