Гаплогруппа E-V68
| Гаплогруппа E-V68 | |
|---|---|
| Возможное время происхождения | в. 24000 лет назад [ 1 ] |
| Возраст слияния | в. 19 900 лет назад [ 1 ] |
| Возможное место происхождения | Египет / Ливия [ 2 ] или южный Египет/северный Судан [ 3 ] |
| Предок | Э-М35 [ 4 ] |
| Потомки | Э-М78 , [ 4 ] E-V1039 |
| Определение мутаций | В68, Л539, ПФ2203 [ 4 ] |
Гаплогруппа E-V68 , также известная как E1b1b1a , является основной гаплогруппой ДНК Y-хромосомы человека, обнаруженной в Северной Африке , на Африканском Роге , в Западной Азии и Европе . Это субклад более крупной и старой гаплогруппы, известной как E1b1b или E-M215 (также примерно эквивалентной E-M35). Линия E1b1b1a идентифицируется по наличию однонуклеотидного полиморфизма (SNP) мутации на Y-хромосоме , известной как V68. Это предмет обсуждения и изучения в генетике , а также в генетической генеалогии , археологии и исторической лингвистике .
В E-V68 доминирует более известный субклад E-M78 . В различных публикациях и E-V68, и E-M78 упоминались под другими именами, особенно в филогенетической номенклатуре, такой как «E3b1a», которая призвана показать их место в генеалогическом древе всех мужчин. Эти различные названия меняются по мере новых открытий и обсуждаются ниже.
Происхождение
[ редактировать ]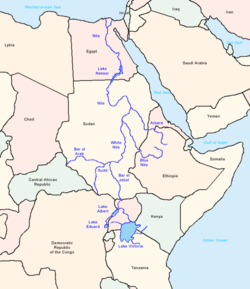
Считается, что E-M78, как и его родительская клада E-V68, имеет африканское происхождение. На основании STR данных генетической дисперсии Cruciani et al. (2007) предполагает, что этот субклад возник в «Северо-Восточной Африке», что в исследовании относится конкретно к региону Египта и Ливии . [ 5 ]
До Cruciani et al. (2007) , Семино и др. (2004) предложили место происхождения E-M78 южнее, в Восточной Африке . Это произошло из-за высокой частоты и разнообразия линий E-M78 в регионе Эфиопии. Однако Кручиани и др. (2007) смогли изучить больше данных и пришли к выводу, что в линиях E-M78 на Африканском Роге доминируют относительно недавние ветви (см. E-V32 ниже). Они пришли к выводу, что регион Египта был вероятным местом происхождения E-M78 на основании «периферийного географического распределения наиболее производных субгаплогрупп по отношению к северо-востоку Африки, а также результатов количественного анализа UEP и разнообразия микросателлитов».
Кручиани и др. (2007) также отмечают это как свидетельство существования «коридора двунаправленной миграции» между Северо-Восточной Африкой (Египет и Ливия по их данным) с одной стороны и Восточной Африкой с другой. Поскольку Кручиани и др. (2007) также предположили, что E-M35, родительская клада E-M78, возникла в Восточной Африке во время палеолита и впоследствии распространилась в регион Египта. Таким образом, E-M78 в Восточной Африке является результатом обратной миграции. Авторы полагают, что было «по крайней мере 2 эпизода между 23,9–17,3 тыс. лет назад и 18,0–5,9 тыс. лет назад».
Еще одну вероятную миграцию на юг из Египта отметили Хасан и др. (2008) на основе исследования Судана. В частности, E-V12 и E-V22 «могли быть завезены в Судан из Северной Африки после прогрессирующего опустынивания Сахары около 6000–8000 лет назад».
К северу от Египта и Ливии E-M78 мигрировал на Ближний Восток, но дополнительно Trombetta et al. (2011) предположили, что более ранняя популяция, носительница E-V68, могла мигрировать по морю непосредственно из Африки в юго-западную Европу, поскольку они наблюдали случаи E-V68* (без мутации M78) только на Сардинии , а не в образцах с Ближнего Востока. . Что касается E-M78, то, как и других форм E-V68, есть свидетельства существования множества путей экспансии за пределы африканской родины.
С другой стороны, хотя, очевидно, имели место прямые миграции из Северной Африки в Иберию и Южную Италию (людей, являющихся носителями E-V68*, E-V12, E-V22 и E-V65), большинство линий E-M78 обнаружили в Европе принадлежат к субкладу E-V13, который, по-видимому, проник в Европу в какое-то неопределенное время с Ближнего Востока , откуда он, по-видимому, и возник, через Балканы .
Придя к тем же выводам, что и команда Кручиани и Тромбетта, Батталья и др. (2008) , написанные до открытия E-V68, описывают Египет как «центр распространения различных географически локализованных субкладов, связанных с M78», и, основываясь на археологических данных, они предполагают, что точка происхождения E-M78 (в отличие от более поздних расселений из Египта), возможно, находился в рефугиуме , который «существовал на границе современного Судана и Египта, недалеко от озера Нубия , до начала влажная фаза около 8500 г. до н.э. Движущиеся на север пояса осадков в этот период могли также стимулировать быструю миграцию мезолитических собирателей на север в Африку, Левант и, в конечном итоге, дальше в Малую Азию и Европу, где каждый из них в конечном итоге дифференцировался на свои региональные особенности. ветки».
Разделение E-V68 на субклады, такие как E-V12, E-V13 и т. д., в значительной степени было работой итальянской команды, в которую входили Фульвио Кручиани, Бениамино Тромбетта, Розарио Скоццари и другие. Они начали на основе исследований STR в 2004 году, а затем в 2006 году объявили об открытии мутаций однонуклеотидного полиморфизма (SNP), которые могли бы с большей ясностью определить большинство основных ветвей, что затем обсуждалось дальше в 2007 году. [ 2 ] [ 6 ] [ 7 ] Эти статьи легли в основу обновленной филогении, найденной Карафетом и др. (2008) и ISOGG , что, в свою очередь, является основой филогении, приведенной ниже.
Кейта (2008) изучил опубликованный набор данных по Y-хромосоме афро-азиатских популяций и обнаружил, что ключевая линия E-M35 / E-M78 , субкладовая мутация гаплогруппы E, была общей для популяций в регионе, где первоначально жили египетские носители. и современные кушитские носители с Рога. Эти линии присутствуют у египтян, берберов, кушитов с Африканского Рога и семитов на Ближнем Востоке. Он отметил, что варианты также встречаются в Эгейском море и на Балканах, но происхождение субклада M35 было в Восточной Африке , и его клады доминировали в основной части афро-азиатскоязычного населения, которое включало кушитские , египетские и берберские группы, в Напротив, говорящие на семитском языке показали снижение частоты при движении с запада на восток в левантийско-сирийском регионе. Кейта выявил высокие частоты M35 (>50%) среди омотических популяций, но заявил, что это произошло на основе небольшой опубликованной выборки из 12 человек. Кейта также написал, что мутация PN2 была общей для линий M35 и M2, и эта определенная клада произошла с Востока. Африка. Он пришел к выводу, что «генетические данные дают профили популяций, которые четко указывают на мужчин африканского происхождения, а не азиатского или европейского происхождения», но признал, что биоразнообразие не указывает на какой-либо конкретный набор цветов кожи или черт лица, поскольку популяции подвергались воздействию микроэволюционное давление. [ 8 ] [ 9 ]
Лоосдрехт и др. (2018) проанализировали полногеномные данные семи древних иберомавров из Голубиной пещеры недалеко от Тафоральта на востоке Марокко . Окаменелости были непосредственно датированы периодом от 15 100 до 13 900 лет назад. Ученые обнаружили, что все экземпляры мужского пола с достаточной сохранностью ядерной ДНК принадлежали к субкладу E1b1b1a1 (M78), причем один скелет несет родительскую линию E1b1b1a1b1 к E-V13. [ 10 ] Мартиниано и др. (2022) позже отнесли все образцы тафоральта к гаплогруппе E-M78 и ни одного — к E-L618, предшественнику EV13. [ 11 ]
Возраст
[ редактировать ]Батталья и др. (2008) подсчитали, что E-M78 (названный в этой статье E1b1b1a1) существует в Европе уже более 10 000 лет. А совсем недавно Лакан и др. (2011) обнаружили, что человеческие останки, раскопанные в испанской погребальной пещере, датируемые примерно 7000 лет назад, находились в ветви E-V13 E-M78.
В июне 2015 года мутация M78 и последующее начало генеалогических древ E-M78 и E-V68 были датированы Trombetta et al. примерно 20 300–14 800 лет назад. [ 12 ]
Генеалогическое древо
[ редактировать ]Это филогенетическое дерево субкладов гаплогрупп основано на дереве ISOGG 2019, а также на дереве FamilyTreeDNA™.
| В68 |
Э-В68* (E1b1b1a*) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| М78 |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Распределение
[ редактировать ]сообщили о трех людях, которые находятся в E-V68, но не в E-M78, на Сардинии. На данный момент Тромбетта и др. (2011) , объявляя об открытии V68.
E-M78 широко распространен в Северной Африке , Африканском Роге , Западной Азии (до Южной Азии ) и Европе . [ 2 ] [ 7 ]
Самая базальная и редкая парагруппа E-M78* была обнаружена с самыми высокими частотами у египтян из оазиса Гурна (5,88%), с более низкими частотами также наблюдалась у марокканских арабов , сардинцев и балканцев . [ 2 ] [ 3 ] [ 13 ]
Самые высокие частоты всех определенных субкладов E-M78 в основном встречаются среди афроазиатскоязычного населения на большой территории, простирающейся от предполагаемого места происхождения гаплогруппы в Верхнем Египте до Судана и Африканского Рога . [ 6 ]
За пределами этой основной области распространения (Северная Африка и Африканский Рог) E-V68 также наблюдается в других частях континента с более низкими частотами из-за более недавнего расселения. Таким образом, сегодня он встречается в районах Великих африканских озер и Южной Африки благодаря первым афро-азиатскоязычным поселенцам из региона Рога. [ 12 ] и на запад до Гвинеи-Бисау , где его присутствие предположительно связано с транссахарским перемещением людей из Северной Африки. [ 14 ]
В распространении E-V68 в Европе преобладает его субклад E-V13, за исключением Иберии. E-V13 имеет пик частоты, сосредоточенный в некоторых частях Балкан ( примерно 20% в южных районах; почти 50% приходится на некоторые определенные места и группы населения). [ 15 ] [ 16 ] ) и Италия. Сегодня он имеет более низкие частоты в западных, центральных и северо-восточных районах, хотя E-V13 был найден в неолитическом захоронении в Каталонии. Более подробно это обсуждается ниже.
| Область | Население | н | Э-М78 | Э-М78* | Э-В12* | Э-В13 | Э-В22 | Э-В32 | Э-В65 | Изучать |
|---|---|---|---|---|---|---|---|---|---|---|
| Европа | албанцы | 55 | 25.46% = (14/55) | 1.82% = (1/55) | 23.64% = (13/55) | [ 17 ] | ||||
| Европа | Македонские албанцы | 64 | 35.94% = (23/64) | 1.56% = (1/64) | 34.38% = (22/64) | [ 17 ] | ||||
| Европа | албанцы и Македонские албанцы |
55+ 64= 119 |
31.09% = (37/119) | 1.68% = (2/119) | 29.41% = (35/119) | [ 17 ] | ||||
| Европа | Косовские албанцы | 114 | 45.61% = (52/114) | 1.75% = (2/114) | 43.86% = (50/114) | Перичич и др. (2005) | ||||
| Европа | албанцы | 96 | 32.29% = (31/96) | 32.29% = (31/96) | Кручиани и др. (2007) | |||||
| Европа | Косовские албанцы и македонские албанцы и албанцы |
119+ 114+ 96= 329 |
36.47% = (120/329) | 1.22% = (4/329) | 35.26% = (116/329) | [ 17 ] Перичич и др. (2005) Кручиани и др. (2007) | ||||
| Европа | Македонские арумыны | 57 | 29.82 | 29.82 | Перичич и др. (2005) | |||||
| Европа | сербы | 113 | 20.35 | 1.77 | 18.58 | Перичич и др. (2005) | ||||
| Европа | хорваты | 108 | 5.60 | 5.60 | Перичич и др. (2005) | |||||
| Европа | Крит | 193 | 6.7% = 13/193 | 6.7% = 13/193 | Кинг и др. (2008) | |||||
| Европа | Греки из Неа Никомедеи | 57 | 15.8% = 9/57 | 1.8% = 1/57 | 14.0% = 8/57 | Кинг и др. (2008) | ||||
| Европа | Греки из Сескло/Димини | 57 | 38.6% = 22/57 | 3.5% = 2/57 | 35.1% = 20/57 | Кинг и др. (2008) | ||||
| Европа | Греки из Лерны/Франхти | 57 | 35.1% = 20/57 | 35.1% = 20/57 | Кинг и др. (2008) | |||||
| Европа | Греки с Крита и Греки из Неа Никомедеи Греки из Сескло/Димини из Лерны/Франчти |
193+ 57+ 57+ 57= 364 |
17.58% = 64/364 | 0.82% = 3/364 | 16.76% = 61/364 | Кинг и др. (2008) | ||||
| Европа | Континентальные греки | 147 | 19.05% = 28/147 | 17.69% = 26/147 | 0.68% = 1/147 | 0.68% = 1/147 | Кручиани и др. (2007) | |||
| Европа | Греки с Крита | 215 | 6.51% = 14/215 | 0.93% = 2/215 | 5.58% = 12/215 | Кручиани и др. (2007) | ||||
| Европа | Греки с островов Эгейского моря | 71 | 16.9% = 12/71 | 15.49% = 11/71 | 1.41% = 1/71 | Кручиани и др. (2007) | ||||
| Европа | Континентальные греки Греки с Крита Греки с островов Эгейского моря |
147+ 215+ 71= 433 |
12.47% = 54/433 | 0.46% = 2/433 | 11.32% = 49/433 | 0.46% = 2/433 | 0.23% = 1/433 | Кручиани и др. (2007) | ||
| Европа | Греки с Крита и Греки из Неа Никомедеи Греки из Сескло/Димини из Лерны/Франчти Континентальные греки Греки с Крита Греки с островов Эгейского моря |
364+ 433= 797 |
14.81% = 118/797 | 0.38% = 3/797 | 0.25% = 2/797 | 13.8% = 110/797 | 0.25% = 2/797 | 0.13% = 1/797 | Кинг и др. (2008) Кручиани и др. (2007) | |
| Европа | сицилийцы | 236 | 11.43 | 1.27 | 5.93 | 3.81 | 0.42 | Ди Гаэтано и др. (2009) | ||
| Европа | Уэльва Андалусийцы | 167 | 6.59 | 1.20 | 4.19 | 0.60 | 0.60 | Амвросий и др. (2010) | ||
| Европа | македонцы | 99 | 18.18 | 17.17 | 1.01 | Кручиани и др. (2007) | ||||
| Европа | болгары | 204 | 16.67 | 0.49 | 16.18 | Кручиани и др. (2007) | ||||
| Европа | сицилийцы | 153 | 13.07 | 0.65 | 7.19 | 4.58 | 0.65 | Кручиани и др. (2007) | ||
| Европа | Северные итальянцы | 94 | 7.45 | 5.32 | 2.13 | Кручиани и др. (2007) | ||||
| Европа | Центральные итальянцы | 356 | 7.87 | 0.28 | 5.34 | 1.97 | 0.28 | Кручиани и др. (2007) | ||
| Европа | Южные итальянцы | 141 | 10.64 | 0.71 | 8.51 | 1.42 | Кручиани и др. (2007) | |||
| Европа | сардинцы | 374 | 3.48 | 0.27 | 0.27 | 1.07 | 0.8 | 1.07 | Кручиани и др. (2007) | |
| Европа | Северный португальский | 50 | 4 | 4 | Кручиани и др. (2007) | |||||
| Европа | Южный португальский | 49 | 4.08 | 4.08 | Кручиани и др. (2007) | |||||
| Европа | Пасиегос из Кантабрии | 56 | Кручиани и др. (2007) | |||||||
| Европа | Астурийцы | 90 | 10 | 5.56 | 4.44 | Кручиани и др. (2007) | ||||
| Европа | Южные испанцы | 62 | 3.23 | 3.23 | Кручиани и др. (2007) | |||||
| Европа | Испанские баски | 55 | Кручиани и др. (2007) | |||||||
| Европа | Французские баски | 16 | 6.25 | 6.25 | Кручиани и др. (2007) | |||||
| Европа | Французский | 225 | 4.44 | 0.44 | 4 | Кручиани и др. (2007) | ||||
| Европа | Английский | 28 | Кручиани и др. (2007) | |||||||
| Европа | датский | 35 | 2.86 | 2.86 | Кручиани и др. (2007) | |||||
| Европа | немцы | 77 | 3.9 | 3.9 | Кручиани и др. (2007) | |||||
| Европа | Польский | 40 | 2.5 | 2.5 | Кручиани и др. (2007) | |||||
| Европа | чехи | 268 | 4.85 | 4.85 | Кручиани и др. (2007) | |||||
| Европа | словаки | 24 | 8.33 | 8.33 | Кручиани и др. (2007) | |||||
| Европа | Словенцы | 104 | 2.88 | 2.88 | Кручиани и др. (2007) | |||||
| Европа | Эстонцы | 74 | 4.05 | 4.05 | Кручиани и др. (2007) | |||||
| Европа | белорусы | 40 | Кручиани и др. (2007) | |||||||
| Европа | Северные русские | 82 | 3.66 | 3.66 | Кручиани и др. (2007) | |||||
| Европа | Южные русские | 92 | 2.17 | 2.17 | Кручиани и др. (2007) | |||||
| Европа | украинцы | 11 | 9.09 | 9.09 | Кручиани и др. (2007) | |||||
| Европа | молдаване | 77 | 7.79 | 7.79 | Кручиани и др. (2007) | |||||
| Европа | венгры | 106 | 9.43 | 9.43 | Кручиани и др. (2007) | |||||
| Европа | румыны | 265 | 7.55 | 7.17 | 0.38 | Кручиани и др. (2007) | ||||
| Северо-Западная Африка | Марокканские арабы | 55 | 40 | 3.64 | 7.27 | 29.09 | Кручиани и др. (2007) | |||
| Северо-Западная Африка | Асни Берберы | 54 | 3.7 | 3.7 | Кручиани и др. (2007) | |||||
| Северо-Западная Африка | Бухрия Берберы | 67 | 1.49 | 1.49 | Кручиани и др. (2007) | |||||
| Северо-Западная Африка | Берберы Среднего Атласа | 69 | 10.14 | 10.14 | Кручиани и др. (2007) | |||||
| Северо-Западная Африка | Марракешские берберы | 29 | 6.9 | 3.45 | 3.45 | Кручиани и др. (2007) | ||||
| Северо-Западная Африка | Марокканские евреи | 50 | 12 | 2 | 2 | 8 | Кручиани и др. (2007) | |||
| Северо-Западная Африка | Мозабитские берберы | 20 | Кручиани и др. (2007) | |||||||
| Северо-Восточная Африка | Ливийские евреи | 25 | 8 | 4 | 4 | Кручиани и др. (2007) | ||||
| Северо-Восточная Африка | Ливийские арабы | 10 | 20 | 20 | Кручиани и др. (2007) | |||||
| Северо-Восточная Африка | Северные египтяне (Дельта) | 72 | 23.61 | 5.56 | 1.39 | 13.89 | 2.78 | Кручиани и др. (2007) | ||
| Северо-Восточная Африка | Египетские берберы | 93 | 6.45 | 2.15 | 4.3 | Кручиани и др. (2007) | ||||
| Северо-Восточная Африка | Египтяне из Бахари | 41 | 41.46 | 14.63 | 2.44 | 21.95 | 2.44 | Кручиани и др. (2007) | ||
| Северо-Восточная Африка | Египтяне из оазиса Гурна | 34 | 17.65 | 5.88 | 8.82 | 2.94 | Кручиани и др. (2007) | |||
| Северо-Восточная Африка | Египтяне | 70 | 79 | 79 | Труба (2015) | |||||
| Северо-Восточная Африка | Южные египтяне | 79 | 50.63 | 44.3 | 1.27 | 3.8 | 1.27 | Кручиани и др. (2007) | ||
| Восточная Африка | Шитье | 26 | 15.38 | 3.85 | 11.54 | Хасан и др. (2008) | ||||
| Восточная Африка | Шиллук | 15 | 13.33 | 13.33 | Хасан и др. (2008) | |||||
| Восточная Африка | Нуэр | 12 | 16.67 | 16.67 | Хасан и др. (2008) | |||||
| Восточная Африка | Деревня | 26 | 15.38 | 3.85 | 11.54 | Хасан и др. (2008) | ||||
| Восточная Африка | пасмурно | 28 | 25 | 3.57 | 3.57 | 7.14 | 10.71 | Хасан и др. (2008) | ||
| Восточная Африка | Можно сказать | 32 | 71.88 | 3.13 | 15.63 | 53.13 | Хасан и др. (2008) | |||
| Восточная Африка | Шерсть | 32 | 59.38 | 18.75 | 40.63 | Хасан и др. (2008) | ||||
| Восточная Африка | нубийцы | 39 | 15.38 | 12.82 | 2.56 | Хасан и др. (2008) | ||||
| Восточная Африка | Фулани из Судана | 26 | 34.62 | 30.77 | 3.85 | Хасан и др. (2008) | ||||
| Восточная Африка | Хауса из Судана | 32 | 3.13 | 3.13 | Хасан и др. (2008) | |||||
| Восточная Африка | Египетские копты из Судана | 33 | 15.15 | 15.15 | Хасан и др. (2008) | |||||
| Восточная Африка | Бежа | 42 | 35.71 | 4.76 | 30.95 | Хасан и др. (2008) | ||||
| Восточная Африка | галлов | 50 | 18.00 | 6.00 | 6.00 | 6.00 | Хасан и др. (2008) | |||
| Восточная Африка | Работа | 28 | 14.29 | 3.57 | 10.71 | Хасан и др. (2008) | ||||
| Восточная Африка | Аракиен | 24 | 16.67 | 8.33 | 4.17 | 4.17 | Хасан и др. (2008) | |||
| Восточная Африка | Амхара | 34 | 8.82 | 8.82 | Кручиани и др. (2007) | |||||
| Восточная Африка | Эфиопские евреи | 22 | 9.09 | 9.09 | Кручиани и др. (2007) | |||||
| Восточная Африка | Смешанные эфиопы | 12 | 33.33 | 25 | 8.33 | Кручиани и др. (2007) | ||||
| Восточная Африка | Борана/Оромо (Кения/Эфиопия) | 32 | 40.63 | 40.63 | Кручиани и др. (2007) | |||||
| Восточная Африка | Волайта | 12 | 16.67 | 8.33 | 8.33 | Кручиани и др. (2007) | ||||
| Восточная Африка | Сахо из Эритреи | 94 | 88.3 | 88.3 | Труба (2015) | |||||
| Восточная Африка | Сомали из Эфиопии | 12 | 33.3 | 8.3 | 25 | Труба (2015) | ||||
| Восточная Африка | Сомали из Сомали | 5 | 80 | 80 | Труба (2015) | |||||
| Восточная Африка | Сомали из Кении | 6 | 80 | 80 | Труба (2015) | |||||
| Восточная Африка | Нилотик из Кении | 18 | 11.11 | 11.11 | Кручиани и др. (2007) | |||||
| Восточная Африка | Банту из Кении | 28 | 3.57 | 3.57 | Кручиани и др. (2007) | |||||
| Восточная Африка | Западная Африка | 123 | 0.81 | 0.81 | Кручиани и др. (2007) | |||||
| Восточная Африка | Центральная Африка | 150 | 0.67 | 0.67 | Кручиани и др. (2007) | |||||
| Восточная Африка | Южная Африка | 105 | Кручиани и др. (2007) | |||||||
| Западная Азия | Стамбул Турецкий | 35 | 8.57 | 2.86 | 5.71 | Кручиани и др. (2007) | ||||
| Западная Азия | Юго-западный турецкий | 40 | 2.5 | 2.5 | Кручиани и др. (2007) | |||||
| Западная Азия | Северо-восточный турецкий | 41 | Кручиани и др. (2007) | |||||||
| Западная Азия | Юго-восточный турецкий | 24 | 4.17 | 4.17 | Кручиани и др. (2007) | |||||
| Западная Азия | Эрзурум Турецкий | 25 | 4 | 4 | Кручиани и др. (2007) | |||||
| Западная Азия | Центральная Анатолия | 61 | 6.56 | 1.64 | 4.92 | Кручиани и др. (2007) | ||||
| Западная Азия | Турецкие киприоты | 46 | 13.04 | 10.87 | 2.17 | Кручиани и др. (2007) | ||||
| Западная Азия | Самаритянские левиты | 2 | 100 | 100 | [ 18 ] | |||||
| Западная Азия | Сефардский турецкий | 19 | Кручиани и др. (2007) | |||||||
| Западная Азия | палестинцы | 29 | 10.34 | 3.45 | 6.9 | Кручиани и др. (2007) | ||||
| Западная Азия | Друзы арабы | 28 | 10.71 | 10.71 | Кручиани и др. (2007) | |||||
| Западная Азия | Бедуин | 28 | 3.57 | 3.57 | Кручиани и др. (2007) | |||||
| Западная Азия | Сирийцы | 100 | 2 | 2 | Кручиани и др. (2007) | |||||
| Западная Азия | Курды из Ирака | 20 | Кручиани и др. (2007) | |||||||
| Западная Азия | Арабы из Объединенных Арабских Эмиратов | 40 | 2.5 | 2.5 | Кручиани и др. (2007) | |||||
| Западная Азия | Оманит | 106 | 0.94 | 0.94 | Кручиани и др. (2007) | |||||
| Западная Азия | Адыгей | 18 | Кручиани и др. (2007) | |||||||
| Западная Азия | азербайджанский | 97 | 2.06 | 2.06 | Кручиани и др. (2007) |
Субклады M78
[ редактировать ]
Здесь перечислены основные субклады M78 по состоянию на июнь 2015 года. В субкладе E-M78 Trombetta et al. В 2015 году большая часть бывших хромосом E-M78* была распределена по трем новым отдельным ветвям: E-V1083*, E-V1477 и E-V259. Первая является сестрой парагруппы клад E-V22 и E-V13. Мутация V1477 определяет новую базальную ветвь, наблюдаемую только в одном образце из Северной Африки. Наконец, сестринская клада E-V12, определенная V264, включает E-V65 и новую центральноафриканскую линию, определенную V259. [ 12 ] Редкая субгаплогруппа M78 E1b1b1a1-PF2186 встречается с наибольшей частотой среди населения Тубу, населяющего Чад (21%). [ 20 ]
- E-M78 (E1b1b1a1) Северная Африка, Африканский Рог, Западная Азия, Европа (ранее E1b1b1a).
- Э-М78*
- E-V1477 Встречается у тунисских евреев.
- E-V1083
- PF2186 Встречается среди тубу в районе озера Чад.
- E-V1083* Встречается только в Эритрее (1,1%) и Сардинии (0,3%).
- Э-В13 (E1b1b1a1b)
- Э-В22 (E1b1b1a1s)
- E-V1129
- Э-В12 (E1b1b1a1a)
- Э-В12*
- Э-В32 (E1b1b1a1a2)
- Э-В264
- E-V259 Встречается у чадских (афро-азиатских) говорящих из Северного Камеруна.
- Э-В65 (E1b1b1a1d)
- Э-В12 (E1b1b1a1a)
Э-В12
[ редактировать ]
Этот субклад E-M78, по-видимому, первым отделился от остальных (он возник около 13,7–15,2 тыс. лет назад). [ 21 ] ). По данным Кручиани и др. (2007) , сублиния E-V12, вероятно, возникла в Северной Африке .
Недифференцированные линии E-V12*
[ редактировать ]Недифференцированные линии E-V12* (не E-V32 или E-M224, поэтому названные «E-V12*») имеют пик частоты среди южных египтян (до 74,5%). [ 22 ] Субклады также широко разбросаны в небольших количествах как в Северной Африке, так и в Европе, но практически не распространены в Западной Азии, за исключением Турции. [ 2 ] Эти линии E-V12* ранее были включены (наряду со многими линиями E-V22*) [ Примечание 1 ] ) в оригинальном «дельта-кластере» Кручиани и др. (2004 г.), который он определил с помощью Y-STR профилей . С открытием определяющего SNP Cruciani et al. (2007) сообщили, что самые высокие концентрации V12* были обнаружены в Египте, особенно в Южном Египте. Хасан и др. (2008) сообщают о значительном присутствии E-V12* в соседнем Судане, включая 5/33 коптов и 5/39 нубийцев . E-V12* составлял примерно 20% суданской E-M78. Они предполагают, что субклады E-V12 и E-V22 E-M78 могли быть завезены в Судан из места своего происхождения в Северной Африке после прогрессирующего опустынивания Сахары около 6000–8000 лет назад. Внезапное изменение климата могло заставить несколько неолитических культур/людей мигрировать на север, в Средиземноморье, и на юг, в Сахель и долину Нила. [ 23 ] E-V12* Парагруппа также наблюдается в Европе (например, среди французских басков ) и Восточной Анатолии (например, у Эрзурума турок ). [ 2 ]
Небазальная субгаплогруппа E1b1b-V12/E3b1a1 чаще всего обнаруживается в различных афроазиатскоязычных популяциях Восточной Африки, включая Гарре (74,1%), Габру (58,6%), Вату (55,6%) и Борану (50,0%). ), Санье (41,7%), Бежа (33,3%) и Рендиль (29,0%). [ 24 ]
Субклады E-V12
[ редактировать ]Э-М224
[ редактировать ]E-M224 был обнаружен в Израиле среди йеменского населения (5%) и, по-видимому, представляет собой второстепенный субклад.
О его открытии было объявлено в работе Underhill et al. (2001) и Шен и др. (2004) обнаружили одного из 20 йеменских израильтян , которых они протестировали. Кручиани и др. (2006) назвали M224 «редким и довольно неинформативным» и не нашли экземпляров.
Э-В32
[ редактировать ]
Кручиани и др. (2007) предполагают, что этот субклад E-V12 зародился в Северной Африке , а затем впоследствии распространился дальше на юг, в район Африканского Рога , где он и сейчас распространен. [ Примечание 2 ] До открытия V32 Cruciani et al. (2004) назвали те же линии, что и «гамма-кластер», который, по оценкам, возник около 8500 лет назад. Они заявили, что «наибольшая частота встречается у трех кушитоязычных групп: борана из Кении (71,4%), оромо из Эфиопии (32,0%) и сомалийцев (52,2%). За пределами Восточной Африки он был обнаружен только у двух испытуемых из Египта (3,6%) и у одного араба из Марокко». Санчес и др. (2005) обнаружили, что это чрезвычайно заметно у сомалийских мужчин, и заявили, что «мужское население Сомали представляет собой ветвь популяции Африканского Рога, тесно связанную с оромо в Эфиопии и Северной Кении (Боранас)» и что их линии гамма-кластера «вероятно были завезен в население Сомали 4000–5000 лет назад». Совсем недавно Тиллмар и др. (2009) типировали 147 мужчин из Сомали по 12 локусам Y-STR и заметили, что 77% (113/147) имели типичные гаплотипы E-V32. В настоящее время это самая высокая частота E-V32, обнаруженная в любой выборке популяции. Сходным образом, Хасан и др. (2008) в своем исследовании отметили, что это наиболее распространенный из субкладов E-M78, обнаруженных в Судане , особенно среди беджа , масалит и фур . Бежа, как сомалийцы и оромо, говорят на афро-азиатском языке и живут вдоль «коридора» от Африканского Рога до Египта. Хасан и др. (2008) интерпретируют это как усиление «сильной корреляции между языковым и генетическим разнообразием» и признаков родства между беджа и народами Африканского Рога, такими как амхара и оромо. С другой стороны, масалиты и фур живут в Дарфуре и говорят на нило-сахарском языке. В своем исследовании авторы отметили, что «масалиты обладают безусловно самой высокой частотой гаплогруппы E-M78 и E-V32», что, по их мнению, предполагает «либо недавнее узкое место в популяции, либо близость к происхождению гаплогруппа». Однако совсем недавно Тиллмар и др. (2009) типировали 147 мужчин из Сомали по 12 локусам Y-STR и заметили, что 77% (113/147) имели типичные гаплотипы E-V32. Это самая высокая частота E-V32, обнаруженная в любой выборке популяции.
Данные STR Cruciani et al. (2007) относительно E-V12 можно резюмировать следующим образом.
| Гаплотип | описание | YCAIIa | YCAIIb | ДИС413а | ДИС413б | ДИС19 | ДИС391 | ДИС393 | ДИС439 | ДИС460 | ДИС461 | А10 |
| Э-В12* | капитал | 19 | 22 | 22 | 22 | 13 | 10 | 13 | 11 | 11 | 9 | 13 |
| мин | 18 | 21 | 20 | 21 | 11 | 10 | 12 | 11 | 8 | 8 | 11 | |
| Макс | 19 | 22 | 22 | 23 | 15 | 12 | 14 | 13 | 12 | 10 | 14 | |
| число | 40 | 40 | 40 | 40 | 40 | 40 | 40 | 40 | 40 | 40 | 40 | |
| Э-В32 | капитал | 19 | 21 | 22 | 23 | 11 | 10 | 13 | 12 | 10 | 10 | 13 |
| мин | 19 | 19 | 20 | 21 | 11 | 9 | 12 | 11 | 9 | 10 | 11 | |
| Макс | 20 | 22 | 22 | 24 | 11 | 11 | 13 | 13 | 12 | 11 | 14 | |
| число | 35 | 35 | 35 | 35 | 35 | 35 | 35 | 35 | 35 | 35 | 35 | |
| Все E-V12 | капитал | 19 | 22 | 22 | 23 | 11 | 10 | 13 | 11 | 11 | 10 | 13 |
| мин | 18 | 19 | 20 | 21 | 11 | 9 | 12 | 11 | 8 | 8 | 11 | |
| Макс | 20 | 22 | 22 | 24 | 15 | 12 | 14 | 13 | 12 | 11 | 14 | |
| число | 75 | 75 | 75 | 75 | 75 | 75 | 75 | 75 | 75 | 75 | 75 | |
Э-В13
[ редактировать ]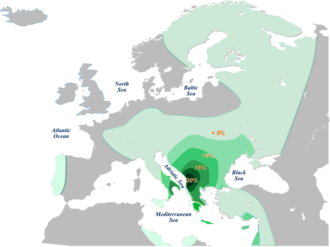
Клада E-V13 эквивалентна «альфа-кластеру» E-M78, о котором сообщалось Cruciani et al. (2004) и впервые был определен SNP V13 Cruciani et al. (2006) . Для этой клады известен другой SNP, V36, о котором сообщает Cruciani et al. (2007) . Все известные положительные тесты на V13 также положительны и на V36. Таким образом, E-V13 в настоящее время считается «филогенетически эквивалентным» E-V36.
Гаплогруппа E-V13 — единственная линия, которая достигает самых высоких частот за пределами Африки. Фактически, он представляет около 85% европейских хромосом E-M78 с клинальным паттерном распределения частот от южной части Балканского полуострова (19,6%) до Западной Европы (2,5%). Та же гаплогруппа с более низкими частотами присутствует в Анатолии (3,8%), на Ближнем Востоке (2,0%) и на Кавказе (1,8%). В Африке гаплогруппа E-V13 встречается редко и наблюдается только в северной Африке с низкой частотой (0,9%).
В Европе E-V13 особенно распространен на Балканах и в некоторых частях Италии. В различных исследованиях особенно высокая частота наблюдалась у косовских албанцев (45,6%), [ 25 ] Македонские албанцы (34,4%), [ 17 ] албанцы (32,29%), [ 26 ] и в некоторых частях Греции (около 35%). [ 27 ] [ 28 ] В целом, высокие частоты также были обнаружены в других районах Греции, а также среди болгар , румын , македонцев и сербов . [ 6 ] [ 16 ] [ 29 ] [ 30 ]
В Италии частота, как правило, выше в Южной Италии . [ 2 ] с особенно высокими результатами, иногда наблюдаемыми в определенных областях; например, в Санта-Нинфе и площади Пьяцца-Армерина на Сицилии . [ 31 ] Высокие частоты, по-видимому, существуют также в некоторых северных районах. [ Примечание 3 ] например вокруг Венеции , [ Примечание 4 ] Генуя [ 32 ] и Римини , [ 33 ] а также на острове Корсика [ 34 ] и регион Прованс на юге Франции, [ 35 ] а также встречается в разрозненных и небольших количествах у ливийских евреев и Египта, но это, скорее всего, результат миграции из Европы или Ближнего Востока. [ 2 ]
E-V13 и древние миграции
[ редактировать ]Очевидное перемещение линий E-M78 с Ближнего Востока в Европу и их последующая быстрая экспансия делают субклад E-V13 особенно интересным предметом для размышлений о древних миграциях людей.
Был сделан вывод, что хромосомы E-M78 начали распространяться в другие регионы именно в северо-восточной Африке, а не в восточной Африке. [ 36 ] Наиболее правдоподобный сценарий заключается в том, что E-V13 зародился в Западной Азии. [ 26 ] Гипотеза состоит в том, что носители E-M78, лишенные мутации V13, покинули Африку и что слияние произошло позже на Ближнем Востоке/Анатолии. [ 26 ] Данные показывают, что западноазиатские носители V13 распространились в Европе не ранее 5300 лет назад. [ 26 ] TMRCA . европейского V13 приходится на 4700–4000 лет назад [ 26 ] Филогенетический анализ предполагает, что европейский v13 распространился по Европе с Балкан в результате «быстрой демографической экспансии». [ 26 ]
До этого мутация SNP V13, по-видимому, впервые возникла в Западной Азии около 10 тысяч лет назад, и, хотя она там не широко распространена, она, например, встречается в больших количествах (> 10% мужского населения) у киприотов-турок и . арабских друзов . [ 2 ] Друзы считаются генетически изолированным сообществом и поэтому представляют особый интерес. [ 37 ] Сигнатура ДНК STR некоторых мужчин E-V13 среди них изначально была классифицирована в дельта-кластере Cruciani et al. (2004) . Это означает, что друзский E-V13 кластеризовался вместе с большинством E-V12 и E-V22, а не с европейским E-V13, который в основном находился в альфа-кластере.
| гаплотип | описание | YCAIIa | YCAIIb | ДИС413а | ДИС413б | ДИС19 | ДИС391 | ДИС393 | ДИС439 | ДИС460 | ДИС461 | А10 |
| Все E-V13 | капитал | 19 | 21 | 23 | 24 | 13 | 10 | 13 | 12 | 9 | 10 | 13 |
| Друзы V13 | 1 | 19 | 21 | 23 | 23 | 13 | 10 | 13 | 13 | 11 | 9 | 12 |
| Друзы V13 | 2 | 19 | 21 | 23 | 23 | 13 | 10 | 13 | 13 | 11 | 9 | 13 |
| Все E-V22 | капитал | 19 | 22 | 22 | 23 | 14 | 10 | 13 | 12 | 11 | 10 | 12 |
| Все E-V12* | капитал | 19 | 22 | 22 | 22 | 13 | 10 | 13 | 11 | 11 | 9 | 13 |
Ранняя миграция с Ближнего Востока в Европу
[ редактировать ]Часто считается, что распространение и разнообразие V13 представляют собой внедрение ранних технологий земледелия во время неолитической экспансии в Европу через Балканы. [ 15 ] Гаплогруппа J2b (J-M12) также часто обсуждалась в связи с V13 как гаплогруппа с, казалось бы, очень похожим распределением и предысторией. [ 3 ] [ 6 ] [ 15 ] (Нет единого мнения относительно обстоятельств и сроков его развития.)
Кручиани и др. (2007) говорит, что для этой географической области запланировано как минимум четыре крупных демографических события:
- «Расширение после последнего ледникового максимума (около 20 тысяч лет назад)»
- « Младший дриас – реэкспансия голоцена (около 12 тыс. лет назад)»
- «Рост населения, связанный с внедрением методов ведения сельского хозяйства (около 8 тыс. лет назад)»
- «Развитие технологии бронзы (около 5 тыс. лет назад)»
Последние два, кажется, находятся в пределах возможного периода времени для V13, учитывая возраст его STR, предположительно возникший на Ближнем Востоке. В пользу связи с сельским хозяйством было показано, что человеческие останки, раскопанные в испанской погребальной пещере, датируемые примерно 7000 лет назад, принадлежат к этой гаплогруппе. [ 38 ]
Однако возможен и более ранний выход в Европу. Батталья и др. (2008) , например, предполагают, что линия E-M78*, являющаяся предком всех современных людей E-V13, быстро вышла из родины Южного Египта в более влажные условия раннего голоцена ; прибыли на Балканы только с технологиями мезолита и лишь впоследствии интегрировались с неолитическими культурами, которые прибыли на Балканы позже.
E-V13 в любом случае часто описывается в популяционной генетике как один из компонентов европейского генетического состава , что указывает на относительно недавнюю связь популяций с Ближнего Востока с проникновением в Европу и, предположительно, связанную с внедрением новых технологий. [ 39 ] [ 40 ] [ 41 ] По существу, также иногда отмечают, что это относительно недавнее генетическое перемещение из Африки в Евразию и описывается как «сигнал для отдельной позднеплейстоценовой миграции из Африки в Европу через Синай ... проявляется в распределении гаплогрупп мтДНК ». [ 42 ]
После его первоначального проникновения в Европу произошло его распространение с Балкан на остальную Европу. Для этого движения также существует широкий спектр возможностей. Батталья и др. (2008) предполагают, что субклад E-V13 из E-M78 возник in situ в Европе, и предполагают, что первое крупное распространение E-V13 с Балкан, возможно, произошло в направлении Адриатического моря с неолитической тисненой керамики культурой . часто называемый Impressa или Cardial . Упомянутая выше находка архаичного E-V13 в Испании подтверждает это предположение.
Напротив, Cruciani et al. (2007) предполагают, что перемещение с Балкан могло произойти совсем недавно, чем 5300 лет назад. Авторы предполагают, что по большей части современные E-V13 происходят от популяции, которая оставалась на Балканах вплоть до балканского бронзового века . Они считают, что «распространение гаплогрупп E-V13 и J-M12, по-видимому, в основном происходило по речным путям, соединяющим южные Балканы с северо-центральной Европой». Перичич и др. (2005) предлагают реки Вардар-Морава-Дунай в качестве возможного пути распространения неолита в Центральную Европу. Бёрд (2007) предполагает еще более недавнее расселение с Балкан, примерно во времена Римской империи.
По мнению Лакана и др. (2011) , Неолитические скелеты (возрастом около 7000 лет), которые были раскопаны в пещере Авелланер в Каталонии , на северо-востоке Испании, включали экземпляр мужского пола, несущий гаплогруппу E1b1b. Эта окаменелость принадлежала к субкладу E1b1b1a1b (V13) и обладала идентичными гаплотипами, обнаруженными у современных европейцев (пять албанцев, два француза из Прованса, два корсиканца, два боснийца, один итальянец, один сицилийский и один грек). Присутствие этой гаплогруппы в неолитической Испании позволяет предположить, что она связана с неолитической сельскохозяйственной упаковкой . Древний фермер также носил кладу мтДНК U5 , раннюю европейскую материнскую гаплогруппу. Его аутосомные маркеры STR также были наиболее типичны для Европы. Кроме того, образец был гомозиготным C/C по SNP персистентности лактазы LP-13910-C/T , что указывает на непереносимость лактозы .
Лоосдрехт и др. (2018) нашли один скелет в Голубиной пещере недалеко от Тафоральта на востоке Марокко , который нес гаплогруппу E1b1b1a1b1, предшественник EV13. Скелет был прямо датирован от 15 100 до 13 900 лет назад. [ 10 ] Мартиниано и др. (2022) позже отнесли все образцы тафоральта к гаплогруппе E-M78 и ни одного — к E-L618, предшественнику EV13. [ 11 ] Фернандес и др. (2016) и Липсон и др. (2017) обнаружили гаплогруппу E-L618 у двух особей из Венгрии и Хорватии, отнесенных к культуре Лендьель . [ 43 ] [ 44 ]
Греческие солдаты в Пакистане
[ редактировать ]И E-M78, и J-M12 также использовались в исследованиях, направленных на поиск доказательств сохраняющегося греческого присутствия в Афганистане и Пакистане , начиная со времен Александра Великого.
Обширный анализ разнообразия Y среди греков и трех пакистанских популяций – бурушо , калашей и патанов – которые заявляют о своем происхождении от греческих солдат, позволил нам сравнить линии Y внутри этих популяций и переоценить их предполагаемое греческое происхождение. Это исследование в целом, похоже, исключает большой вклад Греции в какое-либо население Пакистана, что подтверждает предыдущие наблюдения. Тем не менее, это дает убедительные доказательства в поддержку греческого происхождения небольшой части патанов, о чем свидетельствует сеть клады E и низкие попарные генетические расстояния между этими двумя популяциями.
Однако в этом исследовании проверялся только M78, а не V13, типичный тип M78 с Балкан. Однако более поздний и детальный анализ E-V13 в этом регионе показал, что эта гипотеза неверна и что обнаруженные там варианты не являются типами, типичными для Балкан. [ 45 ] Вместо этого «происхождение Афганистана коррелирует с жителями Ближнего Востока и иранцами, но не с населением Балкан». [ 46 ]
Древняя Британия
[ редактировать ]Значительные частоты E-V13 также наблюдались в городах Уэльса , вокруг Честера (древняя Дева-Виктрикс ) в Англии и Шотландии . В частности, в старом торговом городе Абергеле на северном побережье Уэльса 7 из 18 протестированных местных жителей принадлежали к этой линии (приблизительно 40%), как сообщается в Weale et al. (2002) .
Некоторые ученые (например, Берд (2007) приписывают присутствие E-V13 в Великобритании , особенно в районах с высокой частотой, римским поселениям в период с 1 по 4 века нашей эры. Римская армия включала мужчин балканского происхождения, в том числе фракийцев , Иллирийцы и даки , в частности, Стивен Берд предлагает связь с современным регионом, охватывающим Косово, южную Сербию, северную Македонию и крайнюю северо-западную Болгарию – регионом, соответствующим римской провинции Верхняя Мезия , которая была определена Перичичем и др. 2005) как имеющий самую высокую частоту этого субклада в мире. [ Примечание 5 ]
Также примечательно, что E-V13, по-видимому, отсутствует в современной центральной Англии, особенно в Уэст-Мидлендсе и Южном Мидлендсе . [ Примечание 6 ] Берд (2007) отмечает, что коллективный генетический профиль английского Мидлендса аналогичен профилю голландской провинции Фрисландия , которая не была колонизирована Римом, но, как и Англия, подвергалась англосаксонскому заселению . Так называемая «дыра E3b» в Центральной Англии, по мнению Стивена Берда, может отражать замену населения – романо- британцев англосаксами . [ Примечание 7 ] Томас и др. (2006) поднимает вопрос о возможности существования по типу « апартеида » социальных структур с доминированием элиты в англосаксонской Англии. Бёрд (2007) соглашается: «Дыра E3b предполагает, что либо (а) массовое перемещение... романо-британского населения в результате вторжения, либо (б) существенная генетическая замена романо-британской Y-ДНК через модель доминирования элиты («апартеида»)... Независимо от механизма, регион Центральной Англии... с его отсутствием гаплотипов E3b является территорией, имеющей наиболее «поразительное сходство в распределении Y-хромосом» с Фрисландией. "
Субклады E-V13
[ редактировать ]Хотя у большинства особей E-V13 не обнаруживаются какие-либо известные нижестоящие мутации SNP, и поэтому они классифицируются как E-V13*, существует несколько признанных субкладов, каждый из которых может быть очень небольшим. Это один из двух случаев, когда Карафет и др. (2008) отметили, что на момент написания этой статьи не было уверенности в том, что эти две клады действительно были отдельными («положения этих мутаций не были решены из-за отсутствия образца ДНК, содержащего производное состояние в V27») .
- Э-В27. Определено V27. Кручиани и др. (2007) обнаружили один случай на Сицилии .
- Е-Р65. Определяется P65.
- E-L17. Определяется L17.
- E-L143. Определено L143.
- Э-М35.2. Определено M35.2.
- E-L241. Определяется L241.
- Э-Л250. Определяется L250, L251 и L252.
Э-В22
[ редактировать ]
В эту кладу входит большинство тех, что классифицированы в «дельта-кластере» Cruciani et al. (2004) . Кручиани и др. (2006) позже отметили, что «хромосомы E-V22 и E-V12* перемешаны и нечетко дифференцированы по микросателлитным гаплотипам».
Самая высокая частота E-V22 до сих пор наблюдалась среди самаритянских левитов с частотой 100%. [ 18 ]
Другие частоты, о которых сообщили Cruciani et al. (2007) включают марокканских арабов (7,27%, 55 человек), палестинцев (6,9% из 29 человек). Каденас и др. (2007) обнаружили 7% присутствия в ОАЭ . [ 47 ]
Субклады E-V22
[ редактировать ]Есть два признанных субклада, которые, по-видимому, являются отдельными, хотя Karafet et al. (2008) отметили, что на момент написания этой статьи «положения этих мутаций не были определены из-за отсутствия образца ДНК, содержащего производное состояние [...] V19».
- E-M148 Определяется M148. Андерхилл и др. (2000) нашли 1 экземпляр на Индийском субконтиненте . Кручиани и др. (2006) называет M148 «редким и довольно неинформативным».
- E-V19 Определен V19. Кручиани и др. (2007) нашли 2 экземпляра на Сардинии .
Э-В65
[ редактировать ]Этот субклад, эквивалентный ранее классифицированному «бета-кластеру», в больших количествах встречается в регионах Магриба на крайнем севере Африки. Кручиани и др. (2007) сообщают об уровне около 20% среди ливийских арабов и около 30% среди марокканских арабов . Похоже, что это менее распространено среди берберов , но все же присутствует на уровне> 10%. Авторы предполагают североафриканское происхождение этой линии. В Европе лишь несколько особей были обнаружены в Италии и Греции. Результаты статьи можно резюмировать следующим образом...
| Э-В65 | YCAIIa | YCAIIb | ДИС413а | ДИС413б | ДИС19 | ДИС391 | ДИС393 | ДИС439 | ДИС460 | ДИС461 | А10 |
| капитал | 19 | 21 | 21 | 23 | 13 | 10 | 13 | 10 | 10 | 11 | 13 |
| мин | 19 | 20 | 20 | 22 | 11 | 10 | 13 | 10 | 9 | 9 | 12 |
| Макс | 21 | 21 | 22 | 23 | 14 | 11 | 14 | 11 | 11 | 12 | 13 |
| число | 38 | 38 | 38 | 38 | 38 | 38 | 38 | 38 | 38 | 38 | 38 |
Капелли и др. (2009) изучали бета-кластер в Европе. Они обнаружили небольшие количества в Южной Италии, а также следы в Кантабрии, Португалии и Галисии, при этом в Кантабрии был самый высокий уровень в Европе по их исследованию - 3,1% (5 из 161 человека). Рядом с Кантабрией Родригес и др. (2021) обнаружили высокие частоты E-V65 среди баскских автохтонных жителей провинции Алава (17,3%), провинции Бискайя (10,9%) и провинции Гипускоа (3,3%).
Э-М521
[ редактировать ]Об открытии этого субклада было объявлено в работе Battaglia et al. (2008) Они обнаружили, что у 2 из 92 греков есть эта мутация.
Филогенетика
[ редактировать ]Филогенетическая история
[ редактировать ]До 2002 года в научной литературе существовало как минимум семь систем наименования филогенетического дерева Y-хромосомы. Это привело к значительной путанице. В 2002 году основные исследовательские группы объединились и сформировали Консорциум Y-хромосомы (YCC). Они опубликовали совместный документ, в котором было создано единое новое дерево, которое все согласились использовать. Позже группа гражданских ученых, интересующихся популяционной генетикой и генетической генеалогией, сформировала рабочую группу для создания любительского дерева, стремясь быть прежде всего своевременным. В таблице ниже собраны все эти работы, относящиеся к знаковому дереву YCC 2002 года. Это позволяет исследователю, просматривающему старую опубликованную литературу, быстро перемещаться между номенклатурами.
| ЮКК 2002/2008 (Сокращенно) |
(а) | (б) | (с) | (г) | (е) | (г) | (или) | ЮКК 2002 | ЮКК 2005 г. | ЮКК 2008 | ЮКК 2010р | ИЗОГГ | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (от руки) | 2006 | 2007 | 2008 | 2009 | 2010 | 2011 | 2012 | |||||||||||
| E-P29 | 21 | III | 3А | 13 | Еу3 | Н2 | Б | И* | И | И | И | И | И | И | И | И | И | И |
| Э-М33 | 21 | III | 3А | 13 | Еу3 | Н2 | Б | Е1* | Е1 | Е1а | Е1а | Е1 | Е1 | Е1а | Е1а | Е1а | Е1а | Е1а |
| Э-М44 | 21 | III | 3А | 13 | Еу3 | Н2 | Б | Е1а | Е1а | Е1а1 | Е1а1 | Е1а | Е1а | Е1а1 | Е1а1 | Е1а1 | Е1а1 | Е1а1 |
| Э-М75 | 21 | III | 3А | 13 | Еу3 | Н2 | Б | Е2а | Е2 | Е2 | Е2 | Е2 | Е2 | Е2 | Е2 | Е2 | Е2 | Е2 |
| Э-М54 | 21 | III | 3А | 13 | Еу3 | Н2 | Б | Е2б | Е2б | Е2б | E2b1 | - | - | - | - | - | - | - |
| Е-П2 | 25 | III | 4 | 14 | Еу3 | Н2 | Б | Е3* | Е3 | Е1б | E1b1 | Е3 | Е3 | E1b1 | E1b1 | E1b1 | E1b1 | E1b1 |
| Э-М2 | 8 | III | 5 | 15 | ЕС2 | Н2 | Б | Е3а* | Е3а | E1b1 | E1b1a | Е3а | Е3а | E1b1a | E1b1a | E1b1a | E1b1a1 | E1b1a1 |
| Э-М58 | 8 | III | 5 | 15 | ЕС2 | Н2 | Б | Е3а1 | Е3а1 | E1b1a1 | E1b1a1 | Е3а1 | Е3а1 | E1b1a1 | E1b1a1 | E1b1a1 | E1b1a1a1a | E1b1a1a1a |
| Э-М116.2 | 8 | III | 5 | 15 | ЕС2 | Н2 | Б | E3a2 | E3a2 | E1b1a2 | E1b1a2 | E3a2 | E3a2 | E1b1a2 | E1b1a2 | E1ba12 | удаленный | удаленный |
| Э-М149 | 8 | III | 5 | 15 | ЕС2 | Н2 | Б | Е3а3 | Е3а3 | E1b1a3 | E1b1a3 | Е3а3 | Е3а3 | E1b1a3 | E1b1a3 | E1b1a3 | E1b1a1a1c | E1b1a1a1c |
| Э-М154 | 8 | III | 5 | 15 | ЕС2 | Н2 | Б | Е3а4 | Е3а4 | E1b1a4 | E1b1a4 | Е3а4 | Е3а4 | E1b1a4 | E1b1a4 | E1b1a4 | E1b1a1a1g1c | E1b1a1a1g1c |
| Э-М155 | 8 | III | 5 | 15 | ЕС2 | Н2 | Б | Е3а5 | Е3а5 | E1b1a5 | E1b1a5 | Е3а5 | Е3а5 | E1b1a5 | E1b1a5 | E1b1a5 | E1b1a1a1d | E1b1a1a1d |
| Э-М10 | 8 | III | 5 | 15 | ЕС2 | Н2 | Б | Е3а6 | Е3а6 | E1b1a6 | E1b1a6 | Е3а6 | Е3а6 | E1b1a6 | E1b1a6 | E1b1a6 | E1b1a1a1e | E1b1a1a1e |
| Э-М35 | 25 | III | 4 | 14 | Еу4 | Н2 | Б | E3b* | E3b | E1b1b1 | E1b1b1 | E3b1 | E3b1 | E1b1b1 | E1b1b1 | E1b1b1 | удаленный | удаленный |
| Э-М78 | 25 | III | 4 | 14 | Еу4 | Н2 | Б | E3b1* | E3b1 | E1b1b1a | E1b1b1a1 | E3b1a | E3b1a | E1b1b1a | E1b1b1a | E1b1b1a | E1b1b1a1 | E1b1b1a1 |
| Э-М148 | 25 | III | 4 | 14 | Еу4 | Н2 | Б | E3b1a | E3b1a | E1b1b1a3a | E1b1b1a1c1 | E3b1a3a | E3b1a3a | E1b1b1a3a | E1b1b1a3a | E1b1b1a3a | E1b1b1a1c1 | E1b1b1a1c1 |
| Э-М81 | 25 | III | 4 | 14 | Еу4 | Н2 | Б | E3b2* | E3b2 | E1b1b1b | E1b1b1b1 | E3b1b | E3b1b | E1b1b1b | E1b1b1b | E1b1b1b | E1b1b1b1 | E1b1b1b1a |
| Э-М107 | 25 | III | 4 | 14 | Еу4 | Н2 | Б | E3b2a | E3b2a | E1b1b1b1 | E1b1b1b1a | E3b1b1 | E3b1b1 | E1b1b1b1 | E1b1b1b1 | E1b1b1b1 | E1b1b1b1a | E1b1b1b1a1 |
| Э-М165 | 25 | III | 4 | 14 | Еу4 | Н2 | Б | E3b2b | E3b2b | E1b1b1b2 | E1b1b1b1b1 | E3b1b2 | E3b1b2 | E1b1b1b2a | E1b1b1b2a | E1b1b1b2a | E1b1b1b2a | E1b1b1b1a2a |
| Э-М123 | 25 | III | 4 | 14 | Еу4 | Н2 | Б | E3b3* | E3b3 | E1b1b1c | E1b1b1c | E3b1c | E3b1c | E1b1b1c | E1b1b1c | E1b1b1c | E1b1b1c | E1b1b1b2a |
| Э-М34 | 25 | III | 4 | 14 | Еу4 | Н2 | Б | E3b3a* | E3b3a | E1b1b1c1 | E1b1b1c1 | E3b1c1 | E3b1c1 | E1b1b1c1 | E1b1b1c1 | E1b1b1c1 | E1b1b1c1 | E1b1b1b2a1 |
| Э-М136 | 25 | III | 4 | 14 | Еу4 | Н2 | Б | E3ba1 | E3b3a1 | E1b1b1c1a | E1b1b1c1a1 | E3b1c1a | E3b1c1a | E1b1b1c1a1 | E1b1b1c1a1 | E1b1b1c1a1 | E1b1b1c1a1 | E1b1b1b2a1a1 |
Научные публикации
[ редактировать ]Следующие исследовательские группы в соответствии с их публикациями были представлены при создании дерева YCC.
См. также
[ редактировать ]Генетика
[ редактировать ]- Африканская примесь в Европе
- Генетическая генеалогия
- Гаплогруппа D (Y-ДНК)
- Гаплогруппа DE (Y-ДНК)
- Гаплогруппа
- Гаплотип
- Гаплогруппа ДНК Y-хромосомы человека
- Молекулярная филогенетика
- Парагруппа
- Субклад
- Гаплогруппы Y-хромосомы в популяциях мира
- Гаплогруппы Y-ДНК по этническим группам
- Гаплогруппы Y-ДНК в населении стран Африки к югу от Сахары
Y-ДНК и субклады
[ редактировать ]- Гаплогруппа E-L485 (Y-ДНК)
- Гаплогруппа E-M180 (Y-ДНК)
- Гаплогруппа E-M33 (Y-ДНК)
- Гаплогруппа E-M96 (Y-ДНК)
- Гаплогруппа E-P147 (Y-ДНК)
- Гаплогруппа E-P177 (Y-ДНК)
- Гаплогруппа E-P2 (Y-ДНК)
- Гаплогруппа E-V12 (Y-ДНК)
- Гаплогруппа E-V13 (Y-ДНК)
- Гаплогруппа E-V22 (Y-ДНК)
- Гаплогруппа E-V65 (Y-ДНК)
- Гаплогруппа E-V38 (Y-ДНК)
- Гаплогруппа E-M215 (Y-ДНК)
- Гаплогруппа E-M123 (Y-ДНК)
- Гаплогруппа E-M75 (Y-ДНК)
- Гаплогруппа E-V68 (Y-ДНК)
- Гаплогруппа E-Z820 (Y-ДНК)
- Гаплогруппа E-Z827 (Y-ДНК)
- Гаплогруппа E-M521 (Y-ДНК)
Основополагающее дерево Y-ДНК
[ редактировать ]Примечания
[ редактировать ]- ^ Кручиани и др. (2004) : «Хромосомы E-V22 и E-V12* перемешаны и нечетко дифференцированы по микросателлитным гаплотипам». В Cruciani et al. (2007) те же авторы показывают, что ветвь E-V13, обнаруженная среди арабов-друзов, также находится в дельта-кластере. (Сравните таблицы данных Cruciani et al. (2007) и Cruciani et al. (2004) ).
- ^ Кручиани и др. (2007) : Рис. 2/С
- ^ Не все генетические исследования проверяют одни и те же маркеры.
- ^ Скоццари и др. 2001 . См. кладу 25.1. Тот же набор данных позже был использован Cruciani et al. (2004) и Кручиани и др. (2007) .
- ^ Сомнения по поводу этой линии рассуждений были выражены , потому что: (а) новые данные, появившиеся в King et al. (2008) указывает на то, что высокие концентрации E-V13 также наблюдались в Греции и (б) данные Pericic et al. (2005) показывают, что область с самой высокой частотой не имеет самого высокого разнообразия, а это означает, что V13 прибыл туда позже, чем в Грецию.
- ^ Берд использует три источника: Weale et al. (2002) , Капелли и др. (2003) и Сайкс (2006) . Ни Капелли, ни Уил не имеют данных из района в английском Мидлендсе, где Берд предполагает отсутствие E1b1b [ редактор E-M243 ]. В 2006 году Бёрд упомянул, что у Сайкса было 193 центральноанглийских гаплотипа.
- ^ Однако на картах распространения E3b, опубликованных в собственной статье Берда, в районе Норфолка показан высокий процент E3b. Норфолк является частью эпицентра предполагаемого вторжения англичан .
- ^ Перейти обратно: а б «E-L539 YДерево» .
- ^ Перейти обратно: а б с д и ж г час я дж Кручиани и др. (2007)
- ^ Перейти обратно: а б с Батталья и др. (2008)
- ^ Перейти обратно: а б с ISOGG, авторские права принадлежат 2016 г. «ISOGG 2017 Гаплогруппа E Y-ДНК» . isogg.org . Проверено 7 января 2019 г.
{{cite web}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Кручиани и др. (2007) Таблица 1
- ^ Перейти обратно: а б с д Кручиани и др. (2004)
- ^ Перейти обратно: а б Кручиани и др. (2006)
- ^ Кейта, СОЙ (2008). «География, избранные афро-азиатские семьи и вариации линии Y-хромосомы: исследование лингвистики и филогеографии». В поисках языка в доисторические времена: очерки в четырех областях антропологии . Амстердам: Паб John Benjamins. стр. 3–17. ISBN 978-9027232526 .
- ^ Кейта, Шомарка Омар (3 декабря 2008 г.). География, избранные афро-азиатские семьи и вариации линии Y-хромосомы: исследование лингвистики и филогеографии . Издательство Джона Бенджамина. ISBN 978-90-272-3252-6 .
- ^ Перейти обратно: а б Лоосдрехт; и др. (2018). «Плейстоценовые геномы Северной Африки связывают человеческие популяции Ближнего Востока и Африки к югу от Сахары» . Наука . 360 (6388): 548–552. Бибкод : 2018Sci...360..548V . дои : 10.1126/science.aar8380 . ПМИД 29545507 .
- ^ Перейти обратно: а б Мартиниано, Руи; Де Санктис, Бьянка; Халласт, Таблетки; Дурбин, Ричард (февраль 2022 г.). «Помещение древних последовательностей ДНК в эталонные филогении» . Молекулярная биология и эволюция . 39 (2). дои : 10.1093/molbev/msac017 . ПМЦ 8857924 . ПМИД 35084493 .
- ^ Перейти обратно: а б с Труба (2015)
- ^ Амвросий и др. (2010)
- ^ Роуз и др. (2007)
- ^ Перейти обратно: а б с Семино и др. (2004)
- ^ Перейти обратно: а б Перичич и др. (2005)
- ^ Перейти обратно: а б с д и Батталья и др. 2008 год .
- ^ Перейти обратно: а б Шен, Пейдун; Лави, Таль; Кивисилд, Тоомас; Чоу, Вивиан; Сенгун, Дениз; Гефель, Дов; Шпирер, Иссак; Вульф, Эйлон; Гилель, Йосси; Фельдман, Маркус В.; Офнер, Питер Дж. (сентябрь 2004 г.). «Реконструкция отцовских и матрилиний самаритян и других израильских популяций на основе вариаций последовательностей Y-хромосомы и митохондриальной ДНК» . Человеческая мутация . 24 (3): 248–260. дои : 10.1002/humu.20077 . ISSN 1098-1004 . ПМИД 15300852 . S2CID 1571356 .
- ^ Перейти обратно: а б с Д'Атанасио Э., Тромбетта Б., Бонито М., Финоккио А., Ди Вито Г., Сегиззи М. и др. (2018). «Заселение последней Зеленой Сахары, выявленное путем широкомасштабного повторения последовательности транссахарских отцов» . Геном Биол . 19 (1): 20. дои : 10.1186/s13059-018-1393-5 . ПМК 5809971 . ПМИД 29433568 .
- ^ Хабер, Марк; и др. (2016). «Генетическое разнообразие Чада раскрывает африканскую историю, отмеченную многочисленными голоценовыми евразийскими миграциями» . Американский журнал генетики человека . 99 (6): 1316–1324. дои : 10.1016/j.ajhg.2016.10.012 . ПМК 5142112 . ПМИД 27889059 . - Частоты гаплогрупп Y-хромосомы в Таблице S.4.
- ^ См. рисунок 1.
- ^ Бениамино Тромбетта (2015). «Филогеографическое уточнение и крупномасштабное генотипирование гаплогруппы E Y-хромосомы человека дают новое представление о расселении ранних скотоводов на африканском континенте» . Геномная биология и эволюция . 7 (7): 1940–1950. дои : 10.1093/gbe/evv118 . ПМЦ 4524485 . ПМИД 26108492 .
- ^ Хасан и др. (2008)
- ^ Хирбо, Джибрил Бору. «Комплексная генетическая история человеческих популяций Восточной Африки» (PDF) . Университет Мэриленда, Колледж-Парк . Проверено 13 июля 2017 г.
- ^ Перичич и др. 2005 .
- ^ Перейти обратно: а б с д и ж Кручиани и др. 2007 , «Гаплогруппа E-V13: миграции и демографическая экспансия в Западной Евразии».
- ^ Кинг и др. 2008 год .
- ^ Семино и др. (2004) . предполагают, что уровни E-M78 на Пелопоннесе могут превышать 40%. Они нашли там 17 из 36 (47%), но оправдали выводы из этой небольшой выборки, сославшись также на Di Giacomo et al. (2003)
- ^ Россер и др. 2000 г.
- ^ Кинг и др. (2008)
- ^ Ди Гаэтано и др. (2009)
- ^ Ди Джакомо и др. (2003)
- ^ Пелотти и др. 2007 год
- ^ Франкалаччи и др. 2003 г.
- ^ Кинг, Рой Дж.; и др. (2011). «Приход греков в Прованс и Корсику: модели Y-хромосомы архаической греческой колонизации западного Средиземноморья» . Эволюционная биология BMC . 11 (1): 69. Бибкод : 2011BMCEE..11...69K . дои : 10.1186/1471-2148-11-69 . ПМК 3068964 . ПМИД 21401952 .
- ^ Кручиани и др. 2007 , «Определение происхождения гаплогруппы E-M78».
- ^ Слаш и др. (2008)
- ^ Лакан и др. (2011)
- ^ Семино (2000)
- ^ Кинг и Андерхилл (2002)
- ^ Андерхилл (2002)
- ^ Андерхилл и Каменный мост (2007)
- ^ Липсон 2017 .
- ^ Фернандес 2016 .
- ^ Лакау и др. (2012)
- ^ Хабер и др. (2012)
- ^ Каденас, Алисия М.; Животовский Лев А.; Кавалли-Сфорца, Лука Л.; Андерхилл, Питер А.; Эррера, Рене Дж. (март 2008 г.). «Разнообразие Y-хромосом характеризует Оманский залив» . Европейский журнал генетики человека . 16 (3): 374–386. дои : 10.1038/sj.ejhg.5201934 . ISSN 1476-5438 . ПМИД 17928816 . S2CID 32386262 .
Источники таблиц преобразования
[ редактировать ]- Капелли, Кристиан; Уилсон, Джеймс Ф.; Ричардс, Мартин; Штумпф, Майкл П.Х.; и др. (февраль 2001 г.). «Отцовское наследие преимущественно коренных народов австронезийскоязычных народов островной Юго-Восточной Азии и Океании» . Американский журнал генетики человека . 68 (2): 432–443. дои : 10.1086/318205 . ПМЦ 1235276 . ПМИД 11170891 .
- Хаммер, Майкл Ф.; Карафет Татьяна М.; Редд, Алан Дж.; Джарджанази, Хамди; и др. (1 июля 2001 г.). «Иерархические закономерности глобального разнообразия Y-хромосом человека» . Молекулярная биология и эволюция . 18 (7): 1189–1203. doi : 10.1093/oxfordjournals.molbev.a003906 . ПМИД 11420360 .
- Джоблинг, Марк А.; Тайлер-Смит, Крис (2000), «Новое использование новых гаплотипов», Trends in Genetics , 16 (8): 356–62, doi : 10.1016/S0168-9525(00)02057-6 , PMID 10904265
- Каладжиева, Люба; Калафель, Франческ; Джоблинг, Марк А; Ангеличева, Дора; и др. (февраль 2001 г.). «Закономерности меж- и внутригруппового генетического разнообразия у Vlax Roma, выявленные с помощью Y-хромосомы и линий митохондриальной ДНК» . Европейский журнал генетики человека . 9 (2): 97–104. дои : 10.1038/sj.ejhg.5200597 . ПМИД 11313742 . S2CID 21432405 .
- Карафет, Татьяна; Сюй, Липин; Ду, Руофу; Ван, Уильям; и др. (сентябрь 2001 г.). «История отцовского населения Восточной Азии: источники, закономерности и микроэволюционные процессы» . Американский журнал генетики человека . 69 (3): 615–628. дои : 10.1086/323299 . ПМЦ 1235490 . ПМИД 11481588 .
- Семино, О.; Пассарино, Дж; Оефнер, П.Дж.; Лин, А.А.; и др. (2000), «Генетическое наследие палеолитического Homo sapiens sapiens у современных европейцев: хромосомная перспектива AY», Science , 290 (5494): 1155–9, Бибкод : 2000Sci...290.1155S , doi : 10.1126/science.290.5494. 1155 , ПМИД 11073453
- Су, Бинг; Сяо, Цзюньхуа; Андерхилл, Питер; Дека, Ранджан; и др. (декабрь 1999 г.). «Y-хромосома свидетельствует о миграции современных людей на север в Восточную Азию во время последнего ледникового периода» . Американский журнал генетики человека . 65 (6): 1718–1724. дои : 10.1086/302680 . ПМЦ 1288383 . ПМИД 10577926 .
- Андерхилл, Питер А.; Шен, Пейдун; Лин, Алиса А.; Джин, Ли; и др. (ноябрь 2000 г.). «Вариации последовательности Y-хромосомы и история человеческих популяций». Природная генетика . 26 (3): 358–361. дои : 10.1038/81685 . ПМИД 11062480 . S2CID 12893406 .
Ссылки
[ редактировать ]- Амвросий, Б; Дюгужон, Ж.М.; Эрнандес, К; Из Фонтана, Д; Гонсалес-Мартин, А; Форт-Лима, Калифорния; Новеллетто, А; Родригес, JN; Кальдерон, Р; и др. (2010), «Андалузское население Уэльвы демонстрирует высокую диверсификацию отцовских линий Y-ДНК от гаплогруппы E: Идентификация человеческих мужских движений в средиземноморском пространстве», Annals of Human Biology , 37 (1): 86–107, doi : . http://dx.doi.org/10.3109/03014460903229155 , PMID19939195 , S2CID S2CID1667431
- Адамс, Сьюзен М; Босх, Елена; Балареск, Патрисия Л.; Баллеро, Стефан Ж.; Ли, Эндрю С.; Арройо, Эдуардо; Лопес-Парра, Ана М.; Алер, Мерседес; и др. (2008), «Генетическое наследие религиозного разнообразия и нетерпимости: отцовские линии христиан, евреев и мусульман на Пиренейском полуострове», Американский журнал генетики человека , 83 (6): 725–36, doi : 10.1016/j .ajhg.2008.11.007 , PMC 2668061 , PMID 19061982
- Альварес; Святые, Кристина; Монтьель, Рафаэль; Каейро, Бласкес; Крылья, Абделлатиф; Дюгужон, Жан-Мишель; Алуха, Мария Пилар (2009), «Вариации Y-хромосомы в Южной Иберии: взгляд на вклад Северной Африки», Американский журнал человеческой биологии , 21 ): 407–409, doi : 10.1002/ajhb.20888 , PMID ( 3 , S2CID 7041905
- Арреди, Б; Полони, Э; Параккини, С; Зержал, Т; Фаталлах, Д; Макрелуф, М; Паскали, В; Новеллетто, А; Тайлерсмит, К. (2004), «Преимущественно неолитическое происхождение вариаций Y-хромосомной ДНК в Северной Африке», Американский журнал генетики человека , 75 (2): 338–345, doi : 10.1086/423147 , PMC 1216069 , PMID 15202071
- Батталья, Винченца; Форнарино, Симона; Аль-Захери, Надя; Оливьери, Анна; Лопата, Мэри; Майрес, Натали М; Кинг, Рой Дж; Корни, Сирия; и др. (2008), «Y-хромосомные свидетельства культурного распространения сельского хозяйства в юго-восточной Европе», European Journal of Human Genetics , 17 (6): 820–830, doi : 10.1038/ejhg.2008.249 , PMC 2947100 , PMID 19107149
- Бехар, Дорон М.; Томас, Марк Г.; Скорецкий, Карл; Хаммер, Майкл Ф.; Булыгина Екатерина; Розенгартен, Дрор; Джонс, Эбигейл Л.; Держись, Карен; и др. (октябрь 2003 г.), «Множественное происхождение ашкеназских левитов: свидетельства Y-хромосомы как ближневосточного, так и европейского происхождения», Am. Дж. Хум. Жене. , том. 73, нет. 4, стр. 768–779, doi : 10.1086/378506 , PMC 1180600 , PMID 13680527 . Также на http://www.ucl.ac.uk/tcga/tcgapdf/Behar-AJHG-03.pdf и https://web.archive.org/web/20090304100321/http://www.familytreedna.com/ pdf/400971.pdf
- Бехар; Гарриган; Каплан; Мобашер; Розенгартен (ноябрь 2004 г.), «Контрастные закономерности вариаций Y-хромосомы у евреев-ашкенази и нееврейского европейского населения» (PDF) , Hum. Жене. , том. 114, нет. 4, стр. 354–365, doi : 10.1007/s00439-003-1073-7 , PMID 14740294 , S2CID 10310338 , заархивировано из оригинала (PDF) 10 ноября 2011 г. , получено 14 января 2012 г.
- КРАСОТА, Сандра; Гусмао, Леонор; ЛОПЕС, Александра; АЛВЕС, Синтия; ГОМЕС, Ива; Джузели, Мария; Калафель, Франческ; Карраседо, Анхель; Аморим, Антонио (2006), «Микрофилогеографическая и демографическая история португальских мужских линий» , Annals of Human Genetics , 70 (2): 181–194, doi : 10.1111/j.1529-8817.2005.00221.x , PMID 16626329 , S2CID 4652154 [ мертвая ссылка ]
- Бёрд, Стивен (2007), «Гаплогруппа E3b1a2 как возможный индикатор поселения в Римской Британии солдат балканского происхождения» , Журнал генетической генеалогии , 3 (2), заархивировано из оригинала 22 апреля 2016 г. , получено в 2009 г. 07-15
- Бортолини; Томас, Марк Г.; Чихи, Лурдес; Игл, Джон А.; Кастро-Оф-Вар, Динора; Сальзано, Фрэнсис М.; Руис-Линарес, Андрес (2004), «Типология Рибейро, геномы и испанский колониализм с точки зрения Гран-Канарии и Колумбии» (PDF) , Генетика и молекулярная биология , 27 (1): 1–8, doi : 10.1590/S1415 - 47572004000100001
- Босх, Елена; Калафель, Франческ; Комас, Дэвид; Офнер, Питер Дж.; Андерхилл, Питер А.; Бертранпети, Жауме (2001), «Анализ вариаций Y-хромосомы человека с высоким разрешением показывает резкий разрыв и ограниченный поток генов между северо-западной Африкой и Пиренейским полуостровом», Am J Hum Genet , 68 (4): 1019–1029 , doi : 10.1086/319521 , PMC 1275654 , PMID 11254456
- Бош, Э.; Калафель, Ф.; Гонсалес-Нейра, А.; Флейз, К.; Матеу, Э.; Шейль, Х.-Г.; Хакенбек, В.; Ефремовская, Л.; и др. (2006), «Отцовские и материнские линии на Балканах демонстрируют однородный ландшафт, несмотря на языковые барьеры, за исключением изолированных аромунов» , Annals of Human Genetics , 70 (4): 459–487, doi : 10.1111/j.1469-1809.2005. .00251.x , PMID 16759179 , S2CID 23156886 , заархивировано из оригинала 10 декабря 2012 г.
- Каденас; Животовский Лев А; Кавалли-Сфорца, Лука Л; Андерхилл, Питер А; Эррера, Рене Дж. (2007), «Разнообразие Y-хромосомы характеризует Оманский залив», Европейский журнал генетики человека , 16 (3): 1–13, doi : 10.1038/sj.ejhg.5201934 , PMID 17928816
- Капелли, Кристиан; Рыжая, Никола; Абернети, Джулия К.; Гратрикс, Фиона; Уилсон, Джеймс Ф.; Моен, Торольф; Хервиг, Тор; Ричардс, Мартин; и др. (2003), «Перепись хромосом AY на Британских островах» (PDF) , Current Biology , 13 (11): 979–84, doi : 10.1016/S0960-9822(03)00373-7 , PMID 12781138 , S2CID 526263 , также на сайте [1]
- Каратти; Джино, С.; Торре, К.; Робино, К. (2009), «Подтипирование Y-хромосомной гаплогруппы E-M78 (E1b1b1a) с помощью анализа SNP и его судебно-медицинское применение», International Journal of Legal Medicine , 123 (4): 357–360, doi : 10.1007/s00414 -009-0350-y , PMID 19430804 , S2CID 5657112
- Волосы, Кристиан; Онофри, Валерио; Бризигелли, Франческа; Боски, Илария; Скарниччи, Франческа; Масулло, Мара; Ферри, Джанмарко; Тофанелли, Серджио; и др. (2009), «Мавры и сарацины в Европе: оценка средневекового североафриканского мужского наследия в южной Европе», European Journal of Human Genetics , 17 (6): 848–852, doi : 10.1038/ejhg.2008.258 , PMC 2947089 , PMID 19156170
- Чинниоглу, Ченгиз; Кинг, Рой; Кивисилд, Тоомас; Калфоглу, Эрси; Атасой, Севиль; Каваллери, Джанпьеро Л.; Лилли, Анита С.; Роузман, Чарльз К.; и др. (2004), «Раскопки слоев гаплотипов Y-хромосомы в Анатолии», Hum Genet , 114 (2): 127–48, doi : 10.1007/s00439-003-1031-4 , PMID 14586639 , S2CID 10763736
- Конту, Даниэла; Морелли, Даниэла; Сантони, Федерико; Фостер, Джейми В.; Франкалаччи, Паоло; Кукка, Франческо (2008), «Свидетельства на основе Y-хромосомы донеолитического происхождения генетически однородного, но разнообразного населения Сардинии: выводы для сканирования ассоциаций», PLOS ONE , 3 (1): e1430, Bibcode : 2008PLoSO...3.1430 C , doi : 10.1371/journal.pone.0001430 , PMC 2174525 , PMID 18183308
- Кручиани, Фульвио; Сантоламазца, Пьеро; Шен, Пейдун; Маколей, Винсент; Мораль, Педро; Олкерс, Антонель; Модиано, Дэвид; Холмс, Сьюзан (2002), «Обратная миграция из Азии в страны Африки к югу от Сахары поддерживается анализом гаплотипов Y-хромосомы человека с высоким разрешением», American Journal of Human Genetics , 70 (5): 1197–1214, doi : 10.1086/340257 , ПМК 447595 , ПМИД 11910562
- Кручиани; Ла Фратта; Сантоламазца; Селлитто (май 2004 г.), «Филогеографический анализ гаплогруппы E3b (E-M215) Y-хромосом выявляет множественные миграционные события внутри и из Африки» (PDF) , American Journal of Human Genetics , 74 (5): 1014–1022, doi : 10.1086/386294 , ПМЦ 1181964 , PMID 15042509 , заархивировано из оригинала (PDF) 26 июня 2008 г. , получено 15 июля 2009 г.
- Кручиани; Ла Фратта; Торрони; Андерхилл; Скоццари (2006), «Молекулярное рассечение гаплогруппы E-M78 (E3b1a) Y-хромосомы: апостериорная оценка подхода, основанного на микросателлитных сетях, с помощью шести новых биаллельных маркеров», Human Mutation , 27 (8): 831–2, doi : 10.1002/humu.9445 , PMID 16835895 , S2CID 26886757
- Кручиани, Ф.; Ла Фратта, Р.; Тромбетта, Б.; Сантоламазца, П.; Селитто, Д.; Коломб, Э.Б.; Дюгужон, Ж.-М.; Кривелларо, Ф.; и др. (2007), «Отслеживание прошлых перемещений мужчин-мужчин в Северной/Восточной Африке и Западной Евразии: новые подсказки по гаплогруппам Y-хромосомы E-M78 и J-M12» , Molecular Biology and Evolution , 24 (6): 1300–1311, doi : 10.1093/molbev/msm049 , PMID 17351267 , заархивировано из оригинала 10 октября 2017 г. Также см. Дополнительные данные .
- Ди Гаэтано; Черутти, Франческа; Кробу, Карло; Робино (2009), «Дифференциальная миграция из Греции и Северной Африки на Сицилию подтверждается генетическими данными Y-хромосомы», European Journal of Human Genetics , 17 (1): 91–99, doi : 10.1038/ejhg.2008.120 , PMC 2985948 , PMID 18685561
- Ди Джакомо, Ф.; Лука, Ф.; Ананью, Н.; Чаварелла, Г.; Корбо, Р.М.; Креста, М.; Куччи, Ф.; Ди Стази, Л.; Августиан, В.; Гипараки, М.; Лутрадис, А.; Мамми, К.; Михалодимитракис, EN; Папола, Ф.; Педичини, Г.; Плата, Э.; Терренато, Л.; Тофанелли, С.; Маласпина, П.; Новеллетто, А. (сентябрь 2003 г.). «Клинические закономерности разнообразия Y-хромосомы человека в континентальной Италии и Греции определяются эффектами дрейфа и основателя». Молекулярная филогенетика и эволюция . 28 (3): 387–395. дои : 10.1016/S1055-7903(03)00016-2 . ПМИД 12927125 .
- Эрет, К.; Кейта, СО; Ньюман, П. (2004), «Происхождение афроазиатского языка», Science , 306 (5702): 1680, doi : 10.1126/science.306.5702.1680c , PMID 15576591 , S2CID 8057990
- Эль-Сибай, Мирват; Платт, Дэниел Э.; Хабер, Марк; Сюэ, Яли; Юханна, Соня С.; Уэллс, Р. Спенсер; Изаабель, Хасан; Санюра, Мэй Ф.; и др. (2009), «Географическая структура Y-хромосомного генетического ландшафта Леванта: контраст прибрежных и внутренних территорий», Annals of Human Genetics , 73 (6): 568–581, doi : 10.1111/j.1469-1809.2009.00538 .x , PMC 3312577 , PMID 19686289
- Фернандес, Даниэль (2016). «Предварительные результаты временных рядов древней ДНК доисторического человека из прибрежных и внутренних районов Хорватии» . Исследовательские ворота . ИУАЭС . дои : 10.13140/RG.2.1.2555.9920 .
- Фирасат; Халик, Шагуфта; Мохьюддин, Аиша; Папайоанну, Мирто; Тайлер-Смит, Крис; Андерхилл, Питер А; Аюб, Касим (2006), «Y-хромосомные доказательства ограниченного вклада греков в патанскую популяцию Пакистана», European Journal of Human Genetics , 15 (1): 121–126, doi : 10.1038/sj.ejhg.5201726 , ПМК 2588664 , ПМИД 17047675
- Флорес, Карлос; Мака-Мейер, Николь; Гонсалес, Ана М; Офнер, Питер Дж; Шен, Пейдун; Перес, Хосе А; Рохас, Антонио; Ларруга, Хосе М; Андерхилл, Питер А. (2004), «Сокращенная генетическая структура Пиренейского полуострова, выявленная с помощью анализа Y-хромосомы: последствия для демографии населения», European Journal of Human Genetics , 12 (10): 855–863, doi : 10.1038/sj. ejhg.5201225 , PMID 15280900 , S2CID 16765118
- Флорес; Мака-Мейер, Николь; Ларруга, Хосе М.; Кабрера, Висенте М.; Карадшех, Наиф; Гонсалес, Ана М. (2005), «Изоляты в коридоре миграций: анализ вариаций Y-хромосомы с высоким разрешением в Иордании», J Hum Genet , 50 (9): 435–441, doi : 10.1007/s10038- 005-0274-4 , ПМИД 16142507
- Франкалаччи, П.; Морелли, Л.; Андерхилл, Пенсильвания; Лилли, А.С.; Пассарино, Г.; Усели, А.; Мадедду, Р.; Паоли, Г.; и др. (2003), «Население трех средиземноморских островов (Корсика, Сардиния и Сицилия), определяемое биаллельной изменчивостью Y-хромосомы», Американский журнал физической антропологии , 121 (3): 270–279, doi : 10.1002/ajpa.10265 , ПМИД 12772214
- Фрегель, Роза; ГОМЕС, Вероника; ГУСМИО, Леонор; Гонсалес, Ана М; Кабрера, Висенте М; АМОРИМ, Антониу; Ларруга, Хосе М. (2009), «Демографическая история мужского генофонда Канарских островов: замена местных линий Европой», BMC Evolutionary Biology , 9 (1): 181, Bibcode : 2009BMCEE...9..181F , doi : 10.1186/1471-2148-9-181 , PMC 2728732 , PMID 19650893
- Жерар; Берриш, С; Ауизерат, А; Дитерлен, Ф; Люкотт, Г. (2006), «Североафриканские берберы и арабские влияния в западном Средиземноморье, выявленные гаплотипами ДНК Y-хромосомы» , Human Biology , 78 (3): 307–316, doi : 10.1353/hub.2006.0045 , PMID 17216803 , S2CID 13347549
- Гонсалвеш, Р; Фрейтас, А; Белый, М; РОЗА, А; Фернандес, AT; Животовский, Л.А.; Андерхилл, Пенсильвания; Кивисилд, Т; Брем, А. (2005), «Линии Y-хромосомы из Португалии, Мадейры и Азорских островов фиксируют элементы сефардского и берберского происхождения» , Annals of Human Genetics , 69 (Pt 4): 443–454, doi : 10.1111/j.1529- 8817.2005.00161.x , hdl : 10400.13/3018 , PMID 15996172 , S2CID 3229760 [ мертвая ссылка ]
- Хабер, Марк; Платт, Дэниел Э.; Ашрафиан Бонаб, Мазиар; Юханна, Соня С.; Сория-Эрнанц, Дэвид Ф.; Мартинес-Крус, Бегонья; Дуэи, Бушра; Гассибе-Саббах, Мишелла; Рафатпана, Хошанг; Ганбари, Мохсен; Кит, Джон; Балановский Олег; Уэллс, Р. Спенсер; Комас, Дэвид; Тайлер-Смит, Крис; Заллуа, Пьер А. (2012), «Этнические группы Афганистана разделяют Y-хромосомное наследие, структурированное историческими событиями», PLOS ONE , 7 (3): e34288, Bibcode : 2012PLoSO...734288H , doi : 10.1371/journal.pone .0034288 , PMC3314501 , PMID PMID22470552
- Хаммер (2003), «Структура человеческой популяции и ее влияние на выборку вариаций последовательности Y-хромосомы», Genetics , 164 (4): 1495–1509, doi : 10.1093/genetics/164.4.1495 , PMC 1462677 , PMID 12930755
- Хасан, Хишам Ю.; Андерхилл, Питер А.; Кавалли-Сфорца, Лука Л.; Ибрагим, Мунтасер Э. (2008), «Вариации Y-хромосомы среди суданцев: ограниченный поток генов, соответствие языку, географии и истории» (PDF) , Американский журнал физической антропологии , 137 (3): 316–23, doi : 10.1002/ajpa.20876 , PMID 18618658 , заархивировано из оригинала (PDF) 4 марта 2009 г.
- Хенн, Б.М.; Жиньу, К.; Лин, Алиса А; Офнер, Питер Дж.; Шен, П.; Скоццари, Р.; Кручиани, Ф.; Тишкофф, С.А.; Маунтин, JL; Андерхилл, Пенсильвания (2008), «Y-хромосомные свидетельства миграции скотоводов через Танзанию в южную Африку», PNAS , 105 (31): 10693–8, Bibcode : 2008PNAS..10510693H , doi : 10.1073/pnas.0801184105 , PMC 2504844 , ПМИД 18678889 . См. комментарии в блоге Dienekes , комментарии в блоге Spitoon и публичный выпуск .
- ISOGG (2013), Гаплогруппа E Y-ДНК и ее субклады - 2013 , Международное общество специалистов по генетической генеалогии «ISOGG»
- Джоблинг, Массачусетс; Тайлер-Смит, К. (2000), «Новые варианты использования новых гаплотипов Y-хромосомы человека, болезни и отбор», Trends Genet. , 16 (8): 356–362, doi : 10.1016/S0168-9525(00)02057-6 , PMID 10904265
- Карафет, ТМ; Мендес, Флорида; Мейлерман, МБ; Андерхилл, Пенсильвания; Зегура, СЛ; Хаммер, М.Ф. (май 2008 г.), «Новые бинарные полиморфизмы изменяют форму и увеличивают разрешение дерева гаплогрупп Y-хромосомы человека» , Genome Research , 18 (5): 830–8, doi : 10.1101/gr.7172008 , PMC 2336805 , PMID 18385274 . Опубликовано в Интернете 2 апреля 2008 г. См. также дополнительные материалы.
- Кейта, Шомарка (2008), «География, избранные афро-азиатские семьи и вариации линии Y-хромосомы» , В «Горячей погоне за языком в предыстории: очерки в четырех областях антропологии: в честь Гарольда Крейна Флеминга» , Джон Бенджаминс, ISBN 978-90-272-3252-6
- Кейта, СОИ; Бойс, AJ (Энтони Дж.) (2005), «Генетика, Египет и история: интерпретация географических закономерностей вариаций Y-хромосомы» , History in Africa , 32 : 221–246, doi : 10.1353/hia.2005.0013 , S2CID 163020672
- Кинг, Р.Дж.; Озджан, СС; Картер, Т.; Калфоглу, Э.; Атасой, С.; Триантафиллидис, К.; Коваци, А.; Лин, А.А.; и др. (2008), «Дифференциальное анатолийское влияние Y-хромосомы на греческий и критский неолит» (PDF) , Annals of Human Genetics , 72 (2): 205–214, doi : 10.1111/j.1469-1809.2007.00414.x , ПМИД 18269686 , S2CID 22406638 , заархивировано из оригинала (PDF) 5 марта 2009 г.
- Король; Андерхилл (2002), «Соответствующее распределение расписной керамики и керамических фигурок эпохи неолита с линиями Y-хромосомы», Antiquity , 76 (293): 707–14, doi : 10.1017/S0003598X00091158 , S2CID 160359661
- Куянова; Перейра; Фернандес; Перейра; Черни (2009), «Ближневосточный неолитический генетический вклад в небольшой оазис западной египетской пустыни», Американский журнал физической антропологии , 140 (2): 336–346, doi : 10.1002/ajpa.21078 , PMID 19425100
- Лакан, Мари; Кейзер, Кристина; Рико, Франсуа-Ксавье; Брукато, Николас; Таррус, Хосеп; Босх, Ангел; Гилейн, Жан; Крубези, Эрик; Людес, Бертран (2011), «Древняя ДНК предполагает ведущую роль мужчин в распространении неолита», PNAS , 108 (45): 18255–9, Bibcode : 2011PNAS..10818255L , doi : 10.1073/pnas.1113061108 , PMC 3215063 , ПМИД 22042855
- Лако, Харлетт; Гайден, Тензин; Регейро, Мария; Ченнакришнайя, Шилпа; Бухари, Аридж; Андерхилл, Питер; Гарсиа-Бертран, Ральф; Эррера, Рене (2012), «Афганистан с точки зрения Y-хромосомы», European Journal of Human Genetics , 20 (10): 1063–70, doi : 10.1038/ejhg.2012.59 , PMC 3449065 , PMID 22510847
- Ланкастер, Эндрю (2009), «Гаплогруппы Y, археологические культуры и языковые семьи: обзор междисциплинарных сравнений на примере E-M243» (PDF) , Журнал генетической генеалогии , 5 (1), заархивировано из оригинала ( PDF) от 06 мая 2016 г. , получено 15 июля 2009 г.
- Липсон, Марк (16 ноября 2017 г.). «Параллельные палеогеномные разрезы раскрывают сложную генетическую историю ранних европейских земледельцев» . Природа . 551 (7680). Исследования природы : 368–372. Бибкод : 2017Natur.551..368L . дои : 10.1038/nature24476 . ПМК 5973800 . ПМИД 29144465 .
- Луис, Дж; Роуолд, Д; Регейро, М; Каейро, Б; Чинниоглу, К; Роузман, К; Андерхилл, П; Каваллисфорса, Л; Эррера, Р. (2004), «Левант против Африканского Рога: доказательства двунаправленных коридоров миграции людей» (PDF) , Американский журнал генетики человека , 74 (3): 532–544, doi : 10.1086/382286 , PMC 1182266 , PMID 14973781 , заархивировано из оригинала (PDF) 16 февраля 2012 г. (Также см. Ошибки )
- Мака-Мейер, Н. ; Санчес-Веласко, П.; Цветы, К.; Ларруга, Дж. М.; Гонсалес, AM; Отерино, А; Лейва-Кобиан, Ф; и др. (2003), «Характеристика Y-хромосомы и митохондриальной ДНК Pasiegos, человеческого изолята из Кантабрии (Испания)», Genetics , 67 (Pt 4): CiteSeerX 329–339 , of Human Annals j.1469-1809.2003.00045.x , PMID 12914567 , S2CID 40355653 .
- Мартинес, Лэйзель; Андерхилл, Питер А; Животовский Лев А; Гайден, Тензин; Мошонас, Николас К; Чоу, Шерил-Эмилиан Т; Конти, Саймон; Мамолини, Элизабетта; Кавалли-Сфорца, Л. Лука; Эррера, Рене (1 апреля 2007 г.), «Палеолитическое наследие Y-гаплогруппы преобладает на Критском высокогорном плато», European Journal of Human Genetics , 15 (4): 485–493, doi : 10.1038/sj.ejhg.5201769 , ISSN 1018-4813 , PMID 17264870
- Мендисабаль, Изабель; Сандовал, Карла; Берниэль-Ли, Джемма; Калафель, Франческ; Салас, Антонио; Мартинес-Фуэнтес, Антонио; Комас, Дэвид (2008), «Генетическое происхождение, примесь и асимметрия в материнских и отцовских линиях человека на Кубе», BMC Evol. Биол. , 8 (1): 213, Bibcode : 2008BMCEE...8..213M , doi : 10.1186/1471-2148-8-213 , PMC 2492877 , PMID 18644108
- Небель; Филон, Д; Бринкманн, Б; Маджамдер, П; Фаерман, М; Оппенгейм, А. (2001), «Пул Y-хромосомы евреев как часть генетического ландшафта Ближнего Востока», Американский журнал генетики человека , 69 (5): 1095–1112, doi : 10.1086/324070 , PMC 1274378 , ПМИД 11573163
- Онофри, Валерио; Алессандрини, Федерика; Турки, Кьяра; Песарези, Мауро; Бушеми, Лоредана; Тальябраччи, Адриано (2006), «Разработка мультиплексных ПЦР для эволюционных и судебно-медицинских применений 37 SNP Y-хромосомы человека» (PDF) , Forensic Science International , 157 (1): 23–35, doi : 10.1016/j.forsciint.2005.03 .014 , PMID 15896936 [ постоянная мертвая ссылка ]
- Параккини; Пирс, CL; Колонель, Л.Н.; Альтшулер, Д; Хендерсон, Бельгия; Тайлер-Смит, К. (2003), «Влияние хромосомы AY на риск рака простаты: многоэтническое когортное исследование», J Med Genet , 40 (11): 815–819, doi : 10.1136/jmg.40.11.815 , PMC 1735314 , ПМИД 14627670
- Пелотти; Чеккарди, С; Лугареси, Ф; Трейн, Р; Фалькони, М; Бини, К; Уиллувейт, С; Рёвер, Л. (2007), «Микрогеографическая генетическая изменчивость Y-хромосомы в выборке населения района Равенны в регионе Эмилия-Романья (север Италии)», Forensic Science International: Genetics Supplement Series , 1 (1): 242–243 , doi : 10.1016/j.fsigss.2007.10.025
- Перейра, Луиза; Черный, Виктор; Сересо, Мария; Сильва, Нуно М; Гаек, Мартин; Вашикова, Альжбета; Куянова, Мартина; Брдичка, Радим; Салас, Антонио (2010), «Связывание генофондов стран к югу от Сахары и Западной Евразии: материнское и отцовское наследие кочевников-туарегов из африканского Сахеля» (PDF) , European Journal of Human Genetics , 18 (8): 915–923 , doi : 10.1038/ejhg.2010.21 , PMC 2987384 , PMID 20234393 , заархивировано из оригинала (PDF) 28 мая 2013 г.
- Перичич, М.; Лаук, LB; Кларич, И.М.; Рутси, С; Яничиевич, Б; Рудан, я; Терзич, Р; Чолак, я; и др. (2005), «Филогенетический анализ с высоким разрешением в Юго-Восточной Европе прослеживает основные эпизоды потока отцовских генов среди славянских популяций» , Mol. Биол. Эвол. , том. 22, нет. 10, стр. 1964–75, doi : 10.1093/molbev/msi185 , PMID 15944443 [ мертвая ссылка ] .
- Рамос-Луиза, Э.; Бланко-Вереа, А.; Брион, М.; Ван Хаффель, В.; Карраседо, А.; Санчес-Дис, П. (2009), «Филогеография французских мужских линий (неопубликованные данные 23-го Международного конгресса ISFG)», Forensic Science International , 2 : 439–441, doi : 10.1016/j.fsigss.2009.09.026 , S2CID 85134429 [ мертвая ссылка ]
- Регейру, М.; Каденас, AM; Гайден, Т.; Андерхилл, Пенсильвания; Эррера, Р.Дж. (2006), «Иран: трехконтинентальная связь для миграции, вызванной Y-хромосомой», Hum Hered , 61 (3): 132–143, doi : 10.1159/000093774 , PMID 16770078 , S2CID 7017701
- Робино, К.; Кробу, Ф.; Гаэтано, К.; Бекада, А.; Бенхамамуш, С.; Черутти, Н.; Пьяцца, А.; Интурри, С.; Торре, К. (2008), «Анализ гаплогрупп SNP Y-хромосомы и гаплотипов STR в выборке населения Алжира», Journal International Journal of Legal Medicine , 122 (3): 251–5, doi : 10.1007/s00414-007- 0203-5 , ПМИД 17909833 , С2КИД 11556974
- Родригес, Хавьер; Паленсия-Мадрид, Лейре; Мендоса, Вивиан С; Гарсиа-Бертран, Ральф; Панкорбо, Мириан М де; Эррера, Рене Дж. (2021), «Y-хромосома автохтонных баскских популяций и замена бронзового века», Scientific Reports , 11:5607 (1): 5607, Bibcode : 2021NatSR..11.5607L , doi : 10.1038/s41598-021 -84915-1 , ПМК 7970938 , ПМИД 33692401
- Роза, Александра; Орнелас, Каролина; Джоблинг, Марк А; Брем, Антониу; Виллемс, Ричард (2007), «Разнообразие Y-хромосом в населении Гвинеи-Бисау: многонациональная перспектива», BMC Evolutionary Biology , 7 (1): 124, Bibcode : 2007BMCEE...7..124R , doi : 10.1186 /1471-2148-7-124 , PMC 1976131 , PMID 17662131
- Россер, З; Зержал, Т; Херлс, М; Адохаан, М; Алавантик, Д; Аморим, А; Амос, В; Арментерос, М; и др. (2000), «Разнообразие Y-хромосомы в Европе является клиническим и зависит в первую очередь от географии, а не от языка» , Американский журнал генетики человека , 67 (6): 1526–1543, doi : 10.1086/316890 , PMC 1287948 , PMID 11078479 , заархивировано из оригинала 6 мая 2008 г.
- Санчес, Хуан Дж; Халленберг, Шарлотта; Бёрстинг, Клаус; Эрнандес, Алексис; Горлин, Р.Дж. (2005), «Высокая частота линий Y-хромосомы, характеризующихся E3b1, DYS19-11, DYS392-12 у сомалийских мужчин», European Journal of Human Genetics , 13 (7): 856–866, doi : 10.1038/sj. .ejhg.5201390 , PMID 15756297 . Опубликовано в Интернете 9 марта 2005 г.
- Скоццари, Розарий; Кручиани, Ф; Панграцио, А; Сантоламазца, П; Видишь, Г; Мораль, П; латынь, В; Вареси, Л; и др. (2001), «Вариации Y-хромосомы человека в районе Западного Средиземноморья: последствия для заселения региона» ( PDF) , Human Immunology 62 ( 9) CiteSeerX 871–884 , : , /S0198-8859(01)00286- 5 , PMID 11543889 , заархивировано из оригинала (PDF) 17 декабря 2012 г. , получено 1 июля 2009 г.
- Семино, О.; Пассарино, Дж; Оефнер, П.Дж.; Лин, А.А.; Арбузова С; Бекман, Ле; Де Бенедиктис, Дж; Франкалаччи, П; и др. (2000), «Генетическое наследие палеолитического Homo sapiens sapiens у современных европейцев: хромосомная перспектива AY» (PDF) , Science , vol. 290, нет. 5494, стр. 1155–59, Bibcode : 2000Sci...290.1155S , doi : 10.1126/science.290.5494.1155 , PMID 11073453 , заархивировано из оригинала (PDF) 25 ноября 2003 г.
- Семино; Сантакьяра-Бенерекетти, А. Сильвана; Фаласки, Франческо; Кавалли-Сфорца, Л. Лука; Андерхилл, Питер А. (2002), «Эфиопы и койсанцы разделяют самые глубокие клады филогении Y-хромосомы человека» (PDF) , Am J Hum Genet , vol. 70, нет. 1, стр. 265–268, doi : 10.1086/338306 , PMC 384897 , PMID 11719903 , заархивировано из оригинала (PDF) 15 марта 2006 г.
- Семино, Орнелла; Магри, Кьяра; Бенуцци, Джорджия; Лин, Алиса А.; Аль-Захери, Надя; Батталья, Винченца; Макчиони, Лилиана; Триантафиллидис, Костас; и др. (2004), «Происхождение, распространение и дифференциация гаплогрупп E и J Y-хромосомы: выводы о неолитизации Европы и более поздних миграционных событиях в Средиземноморье», American Journal of Human Genetics , vol. 74, нет. 5, стр. 1023–1034, номер документа : 10.1086/386295 , PMC 1181965 , PMID 15069642.
- Шен, Пейдун; Лави, Таль; Кивисилд, Тоомас; Чоу, Вивиан; Сенгун, Дениз; Гефель, Дов; Шпирер, Иссак; Вульф, Эйлон; и др. (2004), «Реконструкция отцовских и матрилиний самаритян и других израильских популяций на основе вариаций Y-хромосомы и митохондриальной ДНК» (PDF) , Human Mutation , vol. 24, нет. 3, стр. 248–60, doi : 10.1002/humu.20077 , PMID 15300852 , S2CID 1571356 , заархивировано из оригинала (PDF) 05 марта 2009 г. , получено 15 июля 2009 г.
- Сильва; Карвалью, Элизеу; КОСТА, Гильерме; ТАВАРЕС, Лигия; АМОРИМ, Антониу; Гусмао, Леонор (2006), «Генетические вариации Y-хромосомы в популяции Рио-де-Жанейро» , Американский журнал человеческой биологии , 18 (6): 829–837, doi : 10.1002/ajhb.20567 , PMID 17039481 , S2CID 23778828 , в архиве. из оригинала от 18 октября 2012 г.
- Шлаш; Бехар, Дорон М.; Юдковский, Геннадий; Темплтон, Алан; Хадид, Ярин; Базис, Фуад; Хаммер, Майкл; Ицковиц, Шалев; Скорецки, Карл (2008), Геммелл, Нил Джон (редактор), «Друзы: популяционно-генетическое убежище Ближнего Востока», PLOS ONE , 3 (5): e2105, Bibcode : 2008PLoSO...3.2105S , doi : 10.1371/journal.pone.0002105 , PMC 2324201 , PMID 18461126
- Сайкс, Брайан (2006), Кровь островов: исследование генетических корней нашей племенной истории , Bantam, ISBN 978-0-593-05652-3
- Томас; Штумпф, магистр здравоохранения; Харк, Х. (2006), «Свидетельства социальной структуры, подобной апартеиду, в ранней англосаксонской Англии» (PDF) , Proceedings of the Royal Society B , 273 (1601): 2651–2657, doi : 10.1098/rspb. 2006.3627 , PMC 1635457 , PMID 17002951 , заархивировано из оригинала (PDF) 5 марта 2009 г.
- Тиллмар, Андреас О.; Монтелиус, Керстин (декабрь 2009 г.). «Популяционные данные по 12 локусам Y-STR из сомалийского населения». Forensic Science International: Серия дополнений по генетике . 2 (1): 413–415. дои : 10.1016/j.fsigss.2009.08.078 .
- Тромбетта, Бениамино; Кручиани, Фульвио; Селитто, Даниэле; Скоццари, Розария (2011), Маколей, Винсент (ред.), «Новая топология гаплогруппы E1b1 (E-P2) Y-хромосомы человека, выявленная с помощью недавно охарактеризованных бинарных полиморфизмов», PLOS ONE , 6 (1): e16073, Bibcode : 2011PLoSO...616073T , doi : 10.1371/journal.pone.0016073 , PMC 3017091 , PMID 21253605
- Тромбетта (2015), «Филогеографическое уточнение и крупномасштабное генотипирование гаплогруппы E Y-хромосомы человека дают новое представление о расселении первых скотоводов на африканском континенте», Genome Biology and Evolution , 7 (7): 1940–1950, doi : 10.1093 /gbe/evv118 , PMC 4524485 , PMID 26108492
- Андерхилл, Питер А.; Шен, Пейдун; Лин, Алиса А.; Джин, Ли; Пассарино, Джузеппе; Ян, Вэй Х.; Кауфман, Эрин; Бонне-Тамир, Батшева; и др. (2000), «Вариации последовательности Y-хромосомы и история человеческих популяций», Nat Genet , vol. 26, нет. 3, стр. 358–361, doi : 10.1038/81685 , PMID 11062480 , S2CID 12893406
- Андерхилл; Пассарино, Г.; Лин, А.А.; Шен, П.; Миразон Лар, М.; Фоли, РА; Оефнер, П.Дж.; Кавалли-Сфорца, LL (2001), «Филогеография бинарных гаплотипов Y-хромосомы и происхождение современных человеческих популяций», Ann Hum Genet , vol. 65, нет. Часть 1, стр. 43–62, doi : 10.1046/j.1469-1809.2001.6510043.x , PMID 11415522 , S2CID 9441236
- Андерхилл (2002), «Выводы об истории неолитического населения с использованием гаплотипов Y-хромосомы», в Беллвуде, Питер; Ренфрю, А. Колин (ред.), Исследование гипотезы расселения сельского хозяйства и языков, Институт археологических исследований Макдональда , Кембридж: Институт археологических исследований Макдональда, ISBN 978-1-902937-20-5
- Андерхилл, Питер А.; Кивисилд, Тоомас (2007), «Использование структуры популяции Y-хромосомы и митохондриальной ДНК для отслеживания миграций человека», Annu. Преподобный Жене. , 41 : 539–64, doi : 10.1146/annurev.genet.41.110306.130407 , PMID 18076332
- Уил, Мэн; Вайс, Д.А.; Ягер, РФ; Брэдман, Н.; Томас, М.Г. (2002), «Свидетельства Y-хромосомы о массовой миграции англосаксонцев» (PDF) , Mol. Биол. Эвол. , том. 19, нет. 7, стр. 1008–1021, doi : 10.1093/oxfordjournals.molbev.a004160 , PMID 12082121 , заархивировано из оригинала (PDF) 29 октября 2005 г.
- Уил; Шах, Т; Джонс, Алабама; Гринхал, Дж; Уилсон, Дж. Ф.; Нимадава, П; Зейтлин, Д; Коннелл, бакалавр; и др. (1 сентября 2003 г.), «Редкие линии Y-хромосомы с глубокими корнями у человека: уроки филогеографии» , Genetics , vol. 165, нет. 1, стр. 229–234, doi : 10.1093/genetics/165.1.229 , PMC 1462739 , PMID 14504230
- Консорциум Y-хромосомы "YCC" (2002), "Номенклатурная система для дерева бинарных гаплогрупп Y-хромосомы человека", Genome Research , vol. 12, нет. 2, стр. 339–348, doi : 10.1101/гр.217602 , PMC 155271 , PMID 11827954
- Заллуа, Пьер А.; Сюэ, Яли; Халифе, Джейд; Махул, Надин; Дебиан, Лабиб; Платт, Дэниел Э.; Ройюру, Аджай К.; Эррера, Рене Дж.; и др. (2008), «Разнообразие Y-хромосомы в Ливане структурировано недавними историческими событиями», Американский журнал генетики человека , 82 (4): 873–882, doi : 10.1016/j.ajhg.2008.01.020 , PMC 2427286 , PMID 18374297
- Заллуа, Питер А.; Платт, Дэниел Э.; Сибай, Мирват; Халифе, Джейд; Махул, Надин; Хабер, Марк; Сюэ, Он был; Изабель, Хасан; и др. (2008), «Идентификация генетических следов исторической экспансии: финикийские следы в Средиземноморье», Американский журнал генетики человека , 83 (5): 633–642, doi : 10.1016/j.ajhg.2008.10.012 , PMC 2668035 . ПМИД 18976729
- Зержал (1999), Использование вариаций ДНК Y-хромосомы для изучения истории популяции; в Папиха С.С., Дека Р., Чакраборти Р. (ред.): Геномное разнообразие: применение в популяционной генетике человека , Kluwer Academic/Plenum Publishers, стр. 91–101.
- Чжао; Хан, Фейсал; Боркар, Минал; Эррера, Рене; Агравал, Суракша (2009), «Наличие трех разных отцовских линий среди жителей Северной Индии: исследование 560 Y-хромосом», Annals of Human Biology , 36 (1): 1–14, doi : 10.1080/03014460802558522 , PMC 2755252 , PMID 19058044