Генетическая история Африки

Генетическая история Африки обобщает генетический состав и популяционную историю африканских популяций в Африке , состоящую из общей генетической истории , включая региональные генетические истории Северной Африки , Западной Африки , Восточной Африки , Центральной Африки и Южной Африки , а также недавнее происхождение современного человека в Африке . Сахара . служила трансрегиональным проходом и местом проживания людей в Африке во время различных влажных периодов [1] [2] [3] и периоды на протяжении всей истории Африки . [4] [5]
Обзор
[ редактировать ]
Народы также Африки характеризуются региональной генетической субструктурой и гетерогенностью, зависящей от соответствующей этнолингвистической идентичности и отчасти объяснимой «многорегиональной эволюцией» современных человеческих линий в различных многочисленных регионах африканского континента, а более поздние события примеси, включая обратную миграцию из Евразии, как высокодифференцированных западных , так и восточноевразийских компонентов. [6]
Генетическое происхождение африканцев в значительной степени разделено по географическому признаку и языковой семье , при этом популяции, принадлежащие к одним и тем же этнолингвистическим группам, демонстрируют высокую генетическую однородность и согласованность. Поток генов, соответствующий событиям как краткосрочной, так и дальней миграции, за которыми следуют обширные события примеси и узкие места , повлиял на региональный генетический состав и демографическую структуру африканцев. Историческая экспансия банту оказала долгосрочное влияние на современный демографический состав Африки, что привело к большей генетической и языковой гомогенизации. [7] [8] Генетические, археологические и лингвистические исследования добавили дополнительное понимание этого движения: «Наши результаты раскрывают генетический континуум популяций, говорящих на Нигере и Конго, по всему континенту и расширяют наше нынешнее понимание маршрутов, сроков и масштабов миграции банту». [9]
В целом различные африканские популяции демонстрируют генетическое разнообразие и субструктуру, но могут быть объединены в отдельные, но частично перекрывающиеся группы: [10] [11] [12] [8] [13] [14]
- Койсан или « южноафриканские охотники-собиратели» из Южной Африки, представленные койсанскими народами ; они связаны с глубочайшим расхождением (около 270 000 лет назад) генетического разнообразия человека, образуя отдельный кластер. Впоследствии они разделились на Северную и Южную подгруппу, c. 30 000 лет назад. [а]
- «Центральноафриканские охотники-собиратели» или « охотники-собиратели тропических лесов » ( пигмеи ) Центральной Африки, представленные биака и мбути ; связано с другим глубоким расхождением (ок. 220 000 лет назад). Впоследствии они разделились на Восточную и Западную подгруппу, c. 20 000 лет назад. [б]
- « Предки евразийцев » представляют собой древнее население современных евразийцев незадолго до экспансии за пределы Африки; Предполагается, что они отделились от других африканских популяций, скорее всего, где-то в Северо-Восточной Африке , c. 70 000 лет назад.
- Предполагается, что различные на афроазиатском языке говорящие из Северной Африки и Африканского Рога отделились от других африканских групп c. 50 000 лет назад, но в настоящее время недостаточно данных, а поток генов от других групп затрудняет точную оценку даты расхождения. [15] Афроазиатскоязычное население демонстрирует различное количество западно-евразийских (в первую очередь натуфийских , но также неолитических, анатолийских и иранских ) примесей, а остальная часть в первую очередь связана с нилотским происхождением. Они также демонстрируют близость к палеолитическим североафриканским тафоральтов образцам иберомаврийской культуры. [с]
- « Восточноафриканские охотники-собиратели », представленные хадза , сандаве , носителями омотского языка и древним экземпляром мота; их филогенетическое родство с другими популяциями неясно, но они проявляют близость к современным популяциям Восточной и Западной Африки и содержат хэсаноподобный поток генов вдоль линии с северо-востока на юго-запад, а также более поздние (западные) евразийские примеси, но в меньших количествах, чем среди говорящих на афроазиатском языке. [д]
- « Древние восточноафриканцы » или « предки западных/восточных африканцев », связанные с общим предком современных нигер-конголезцев и нило-сахарцев, возникли около 28 000 лет назад, вероятно, в регионе долины Нила . Впоследствии они разошлись в c. 18 000 лет назад в предков западных и западно-центральноафриканских нигер-конголезцев и носителей банту, а также в восточноафриканских нило-сахарских/нилотских говорящих. Они представляют собой доминирующий и наиболее распространенный компонент предков современной Африки и связаны с относительным недавним ростом населения, связанным с сельским хозяйством и скотоводческим образом жизни. Генетические данные указывают на родство с более старыми группами охотников-собирателей в Восточной Африке, но их точное родство остается неясным. [10] [и] Есть свидетельства ограниченного потока генов (9-13%) от человеческой призрачной линии , называемой «западноафриканскими собирателями», с более глубоким или столь же глубоким временем дивергенции, чем у «койсанских охотников-собирателей», к современным западноафриканцам. [16] [17]
- , говорящие на австронезийском языке, Малагасийцы на Мадагаскаре получили значительную Восточной / Юго-Восточной Азии примесь , связанную с австронезийской экспансией , при этом остальная родословная в первую очередь связана с компонентами Западной-Центральной и Восточной Африки. Предполагаемая дата потока генов между этими источниками — ок. 2200 лет назад. [18]

Коренные африканцы
[ редактировать ]Термин « коренные африканцы » относится к населению с преимущественно коренным (неевразийским) происхождением, состоящему из носителей нигерско-конголезского языка , говорящих на нило-сахарском языке , расходящейся и разнообразной койсанской группы , а также нескольких неклассифицированных или изолированных этнолингвистических групп ( см. неклассифицированные языки Африки ). Происхождение афроазиатских языков остается спорным: некоторые предполагают ближневосточное происхождение, а другие поддерживают африканское происхождение. [19] с различной степенью евразийского и африканского компонентов. [15] Австронезийские языки зародились на юге Восточной Азии, а затем распространились за пределы Филиппин .
Нигерско -Конголезские языки , вероятно, возникли в районе, где на этих языках говорили до экспансии банту , или вблизи него (т. е. в Западной или Центральной Африке ). Его расширение могло быть связано с расширением сельского хозяйства в период африканского неолита после высыхания Сахары в ок. 3500 г. до н.э. Прото-Нигер-Конго , возможно, возник примерно за 10 000 лет до того, как он появился в « Зеленой Сахаре » Африки (примерно в Сахеле и южной Сахаре ), и его распространение можно соотнести с распространением лука и стрел мигрирующими охотниками-собирателями. , где позже развилось сельское хозяйство. [21] [22] [23]
Хотя существование нило-сахарской семьи остается спорным, регион между Чадом , Суданом и Центральноафриканской Республикой рассматривается как вероятный кандидат на ее родину до ее расселения около 10 000–8 000 лет до нашей эры. [24]
Предполагается, что южноафриканские охотники-собиратели (койсаны) представляют автохтонное население охотников-собирателей южной Африки до экспансии носителей языка банту из Западной / Центральной Африки и скотоводов Восточной Африки. Койсан демонстрирует наличие примеси, связанной с банту, в диапазоне от почти ~ 0% до ~ 87,1%. [25]
Мероприятие за пределами Африки
[ редактировать ]

« Недавнее африканское происхождение современных людей » предполагает «единое происхождение» Homo sapiens в Африке. Недавние генетические и археологические данные позволяют предположить, что подгруппы Homo sapiens возникли во многих регионах Африки, а не ограничивались одним субрегионом происхождения, причем последний общий предок всех современных людей распространился из одного региона, поглощая или заменяя различные глубокие линии происхождения. описываются как архаичные призраки). Предок H. sapiens собственно евразийцев, скорее всего, покинул Северо-Восточную Африку между 50 000 и 100 000 лет назад. [26] Модель «недавнего африканского происхождения» предполагает, что все современные неафриканские популяции произошли от одной или нескольких волн H. sapiens , покинувших Африку 70 000–60 000 лет назад. [27] [28] [29] [30]

По данным Дурвасулы и др. (2020), есть признаки того, что от 2% до 19% (≃6,6–7,0%) ДНК популяций Западной Африки могло происходить от неизвестного архаичного гоминина, который отделился от предка людей и неандертальцев между 360 и 1,02 млн лет назад. . Однако Дурвасула и др. (2020) также предполагает, что, по крайней мере, часть этой архаичной примеси также присутствует у евразийцев/неафриканцев и что событие или события примеси варьируются от 0 до 124 тыс. лет назад, что включает период до миграции из Африки. и до раскола Африки и Евразии (таким образом, частично затронувшего общих предков как африканцев, так и евразийцев/неафриканцев). [32] [33] [34] Чен и др. (2020) обнаружили, что африканцы имеют более высокое происхождение от неандертальцев, чем считалось ранее. 2504 африканских образца со всей Африки были проанализированы и проверены на неандертальское происхождение. Во всех африканских образцах были обнаружены доказательства незначительного неандертальского происхождения, но всегда на более низком уровне, чем у евразийцев. [35]
Генный поток между евразийскими и африканскими популяциями
[ редактировать ]

Значительная евразийская примесь встречается в Северной Африке , а также среди отдельных этнических групп Рога , Северного Судана , региона Сахеля , а также среди малагасийцев Мадагаскара Африканского . Различные исследования генома обнаружили доказательства множественных доисторических обратных миграций различных евразийских популяций и последующего смешения с местными группами. [37] Поток генов, связанный с Западной Евразией, прибыл в Северную Африку во время палеолита (от 30 000 до 15 000 лет назад), после чего последовали другие миграционные события донеолита и неолита. Генетические данные образцов Тафоральта «продемонстрировали, что Северная Африка получила значительный поток генов из Евразии еще до голоцена и развития методов ведения сельского хозяйства». Средневековые события потока генов, такие как арабская экспансия, также оставили следы в различных африканских популяциях. [26] [14] [38] Пикрелл и др. (2014) указали, что западно-евразийские предки в конечном итоге пришли через Северо-Восточную Африку (особенно Африканский Рог) в Юго-Восточную Африку и Южную Африку . [39]
Рамзи и др. (2018) также обнаружили доказательства значительной западно-евразийской примеси в различных частях Африки, как в результате древних, так и более поздних миграций, причем самый высокий уровень среди популяций из Северной Африки и некоторых групп Африканского Рога : [40]
В дополнение к внутреннему разнообразию внутри континента, обусловленному структурой населения и изоляцией, миграция евразийских популяций в Африку стала решающим фактором генетического разнообразия. Эти миграции включали приток разных евразийских популяций в разное время и в разные части Африки. Всесторонняя характеристика деталей этих миграций посредством генетических исследований существующих популяций могла бы помочь объяснить сильные генетические различия между некоторыми географически соседними популяциями.
Эта характерная евразийская примесь, по-видимому, произошла по крайней мере в течение трех периодов времени: древняя примесь в центральной части Западной Африки (например, йоруба из Нигерии) произошла между ~ 7,5 и 10,5 тыс. лет назад, более старая примесь в Восточной Африке (например, Эфиопия) произошла между ~ 2,4 тыс. лет назад. и 3,2 тыс. лет назад и более поздние примеси от ~0,15 до 1,5 тыс. лет назад в некоторых восточноафриканских (например, кенийских) популяциях.
Последующие исследования, основанные на распаде LD и совместном использовании гаплотипов в обширной группе африканских и евразийских популяций, подтвердили наличие евразийских признаков у жителей западной, восточной и южной Африки. На западе, помимо нигеро-конголезских носителей из Гамбии и Мали, моси из Буркина-Фасо показали старейшее событие евразийской примеси ~ 7 тыс. лет назад. На востоке эти анализы выявили евразийскую примесь в Кении за последние 4000 лет. [40]
Не существует однозначного согласия относительно того, когда и где существовала первоначальная родина афроазиатской языковой семьи . Некоторые предполагают, что они были распространены людьми преимущественно западно-евразийского происхождения во время неолитической революции в Северную Африку и на Африканский Рог, выходцами с Ближнего Востока , особенно из Леванта . [41] Другие утверждают, что первые носители протоафразийского языка проживали в Северо-Восточной Африке, потому что этот регион включает в себя большую часть разнообразия афроазиатской языковой семьи и имеет очень разнообразные группы в непосредственной географической близости, что иногда считается характерным признаком лингвистико-географической близости. источник. [42] Подмножество протоафразийского населения мигрировало в Левант во время позднего палеолита , сливаясь с местными западноевразийцами и приводя к образованию населения, которое позже дало начало натуфийской культуре , связанной с ранним развитием сельского хозяйства и ранними афроазиатскими языками. или конкретно препротосемитского происхождения . [43] [44] [ нужна страница ] [45] [46] [47] [48] Кроме того, Y-гаплогруппы сублиния E-M215 (также известная как «E1b1b) и ее производная E-M35 довольно распространены среди говорящих на афроазиатском языке, а юго-западная Эфиопия является вероятным источником этих гаплогрупп. [49] Согласно этой африканской модели, языковая группа и носители этой линии возникли и рассеялись вместе из Северо-Восточной Африки в эпоху мезолита , вероятно, уже развив модели существования скотоводства и интенсивного использования и сбора растений. [50] [51] [52] [53]
Гипотеза ближневосточных земледельцев не объясняет одомашнивания растений, эндемичных для Африканского Рога, таких как тефф , энсете и нигерское семя , а также не объясняет отсутствия доказательств существования интрузивных сельскохозяйственных популяций или выращивания пшеницы . ячмень или сорго в этом регионе до 3000 г. до н.э. [54] По мнению историка и лингвиста Кристофера Эрета , форма интенсивного сбора растений, практикуемая протоафразийским населением Северо-Восточной Африки, возможно, была предшественником других методов ведения сельского хозяйства, которые позже независимо развились в странах Плодородного полумесяца и Африканского Рога. [47] [55] [56]

Африканский Рог
[ редактировать ]Хотя многие исследования, проведенные на популяциях Африканского Рога, оценивают событие западно-евразийской примеси около 3000 лет назад, [57] [40] [39] [58] Ходжсон и др. (2014) обнаружили отчетливый западно-евразийский наследственный компонент среди изученных афроазиатскоязычных групп на Африканском Роге (и в меньшей степени в Северной Африке и Западной Азии ), наиболее распространенный среди этнических сомалийцев . Этот наследственный компонент, получивший название «эфио-сомалийский», наиболее тесно связан с «магребским» компонентом (с пиком у тунисцев ) и, как полагают, отделился от других неафриканских предков около 23 000 лет назад и мигрировал обратно в Африку до развития сельского хозяйства. (12–23 тыс. лет назад) с Ближнего Востока. Эта популяция должна была пересечь Синайский полуостров, а затем разделиться на две части: одна ветвь продолжила бы путь на запад через Северную Африку, а другая направилась на юг, к Африканскому Рогу. Авторы предполагают, что «эфиосомалийский» компонент мог быть существенным предковым компонентом протоафразиоязычного населения. Более поздняя миграция из Аравии в ТСЖ, начавшаяся около 3 тыс. лет назад, могла бы объяснить происхождение эфиосемитских языков в это время. [45] Анализ мтДНК, проведенный Gandini et al. (2016) предоставили дополнительные доказательства в поддержку доземледельческой обратной миграции из Западной Евразии на Африканский Рог с предполагаемой датой прибытия на Африканский Рог в раннем голоцене , возможно, в результате обсидиана сетей обмена . через Красное море. [59] Ходжсон и др. также подтвердили существование родового компонента Африканского Рога - «эфиопского» или «омотского» (Пагани и др.), - который наиболее распространен среди носителей омотской ветви афроазиатского языка на юго-западе Эфиопии. [45] [57] Эта линия связана с окаменелостью (Мота) возрастом 4500 лет, найденной в пещере на юго-западе Эфиопии, которая имеет высокую генетическую близость к современным эфиопским группам , особенно к эндогамной кузнецов касте народа Омотик Ари . Как и Мота, кузнецы Ари не обнаруживают доказательств смешения с жителями Западной Евразии, демонстрируя определенную степень преемственности населения в этом регионе на протяжении как минимум 4500 лет. В сравнительном анализе генома Моты, относящегося к современным популяциям, Gallego et al. (2016) пришли к выводу, что отличие омотического языка от других афроазиатских языков могло быть результатом относительной изоляции его носителей от внешних групп. [60]
В анализе 68 эфиопских этнических групп Лопес и др. (2021) выявили, что несколько групп, принадлежащих к трем классификациям АА: кушитской, омотской и семитской, в среднем демонстрируют высокое генетическое сходство друг с другом. Более того, говорящие на нило-сахарском языке на юго-западе имели более позднее происхождение с банту и нилотами, в отличие от афроазиатских говорящих на северо-востоке, которые имели более позднее происхождение с египтянами и другими выходцами из Западной Евразии. Данные также подтверждают широко распространенное в последнее время смешение различных этнических групп. [61]
Мадагаскар
[ редактировать ]
Конкретное восточноазиатское происхождение встречается среди малагасийцев Мадагаскара со средней частотой. Присутствие этого восточноазиатского происхождения в основном связано с экспансией австронезийских народов из Юго-Восточной Азии . [62] [63] [64] [65] Было установлено, что народы Борнео напоминают восточноазиатских путешественников, прибывших на Мадагаскар. Восточноазиатское происхождение среди малагасийцев оценивалось в среднем в 33%, но достигало ~75% среди некоторых групп горцев и групп высших каст. [66] [67] [65]
Северная Африка
[ редактировать ]Добон и др. (2015) выявили аутосомный наследственный компонент, который обычно встречается среди современного афроазиатскоязычного населения (а также нубийцев ) в Северо-Восточной Африке. Пик этого коптского компонента приходится на коптов в Судане , который отличается отсутствием арабского влияния, но имеет общее происхождение с населением Северной Африки и Ближнего Востока. Он появляется рядом с компонентом, который определяет нило-сахарские носители юго-западного Судана и Южного Судана . [68] Арауна и др. (2017), анализируя существующие генетические данные, полученные от популяций Северной Африки , таких как берберы , описали их как мозаику североафриканских (тафоральт), ближневосточных, европейских ( ранние европейские фермеры ) и африканских предков, родственных южнее Сахары . [69]
Чен и др. (2020) проанализировали 2504 африканских образца со всей Африки и обнаружили архаическое неандертальское происхождение среди всех протестированных африканских образцов с низкой частотой. Они также определили родственный европейцам (западно-евразийский) сегмент предков, который, по-видимому, во многом соответствует обнаруженным компонентам предков неандертальцев. По оценкам, примесь, связанная с европейцами, среди африканцев составляет от ~ 0% до ~ 30%, с пиком среди жителей Северной Африки. [70] По данным Чена и др. (2020): «Эти данные согласуются с гипотезой о том, что обратная миграция способствовала сигналу о неандертальском происхождении у африканцев. Более того, данные показывают, что эта обратная миграция произошла после разделения европейцев и аборигенов Восточной Азии от популяции, связанной к европейскому происхождению». [70]
Существует незначительный поток генов из Северной Африки в некоторых частях Южной Европы , это подтверждается наличием специфичной для Африки митохондриальной гаплогруппы среди одного из четырех образцов возрастом 4000 лет. [71] Многочисленные исследования также обнаружили доказательства потока генов африканского происхождения в Евразию, особенно в Европу и на Ближний Восток. Анализ 40 различных популяций Западной Евразии выявил африканскую примесь с частотой от 0% до ~ 15%. [72] [73] [74] [75]
Западная Африка
[ редактировать ]Холлфельдер и др. (2021) пришли к выводу, что западноафриканские народы йоруба , которые ранее использовались в качестве «несмешанной эталонной популяции» для коренных африканцев, имеют незначительные уровни неандертальского происхождения, что может быть в значительной степени связано с обратной миграцией исходной популяции, «подобной европейскому предку». . [6]
Полногеномное исследование сообщества фулани из Буркина-Фасо выявило два основных события примеси в этой группе, датируемые примерно 1800 и 300 лет назад. Первое событие смешения произошло между западноафриканскими предками фула и предками североафриканских кочевых групп. Второе событие примеси, относительно недавнее, предполагает источник из Юго-Западной Европы или предполагает дополнительный поток генов между группами фулани и Северной Африки, которые несут пропорции примеси от европейцев. [76] В сахельских популяциях, таких как тубу, также наблюдалась примесь евразийцев. [77]
Южная Африка
[ редактировать ]Низкий уровень западно-евразийского происхождения (европейского или ближневосточного) обнаруживается у говорящих на языке хое-квади хоесан. Оно могло быть приобретено косвенно, путем смешения с мигрирующими скотоводами из Восточной Африки. Эта гипотеза о потоке генов из восточной Африки в южную подтверждается и другими генетическими и археологическими данными, документирующими распространение скотоводства с Востока в Южную Африку. [78]
Региональный геномный обзор
[ редактировать ]Северная Африка
[ редактировать ]ДНК архаичного человека
[ редактировать ]В то время как денисовское и неандертальское происхождение у неафриканцев за пределами Африки более достоверно, архаическое человеческое происхождение у африканцев менее достоверно, и его слишком рано устанавливать с уверенностью. [79]
Древняя ДНК
[ редактировать ]Дэниел Шрайнер (2018), используя современное население в качестве ориентира, показал, что натуфийцы несли 61,2% арабов, 21,2% североафриканцев, 10,9% западноазиатов и небольшую часть восточноафриканского происхождения (6,8%), что связано с современным Группы, говорящие на омотическом языке, обитают на юге Эфиопии. [49]
Египет
[ редактировать ]Хнум-аа , Хнум-Нахт , Нахт-Анх и JK2911 несли материнскую гаплогруппу M1a1 . [80] [57]
Джехутинахт (10А) нес материнскую гаплогруппу U5b2b5 . [81] JK2888 нес материнскую гаплогруппу U6a2 . [57]
Туя , Тийе , мать Тутанхамона, и Тутанхамон несли материнскую К. гаплогруппу [80]
JK2134 нес материнскую гаплогруппу J1d. [57] и JK2887 нес материнскую гаплогруппу J2a1a1 . [57]
Аменхотеп III , Эхнатон и Тутанхамон несли отцовскую гаплогруппу R1b . [80]
Рамсес III и «Неизвестный человек E», возможно, Пентавер , несли отцовскую гаплогруппу E1b1a . [80] [82] [83]
JK2134 и JK2911 несли отцовскую J. гаплогруппу [57]
Такабути был носителем материнской гаплогруппы H4a1. [84] и YM:KMM A 63 несли материнскую гаплогруппу HV . [85]
OM:KMM A 64 нес материнскую гаплогруппу T2c1a . [85]
JK2888 нес отцовскую гаплогруппу E1b1b1a1b2 . [57]
Ливия
[ редактировать ]В каменном убежище Такаркори в Ливии две естественным образом мумифицированные женщины, относящиеся к среднему пастырскому периоду (7000 лет назад), несли базальную материнскую гаплогруппу N. [86]
Марокко
[ редактировать ]Ван де Лоорсдрехт и др. что из семи образцов тафоральтов Марокко кал , датированных радиоуглеродом 100 и 13 900 . 2018) обнаружили , между 15 ( Было обнаружено, что шесть из шести мужчин являются носителями отцовской гаплогруппы E1b1b . Было обнаружено, что они имеют 63,5% предков, связанных с натуфийцами , и 36,5% предков, связанных с выходцами из Африки к югу от Сахары . Компонент стран к югу от Сахары наиболее сильно представлен современными западноафриканскими группами, такими как йоруба и менде . Образцы также содержат дополнительную близость к внешним группам Южной, Центральной и Восточной Африки, которую нельзя объяснить какими-либо известными древними или современными популяциями. [87] При проецировании на график анализа главных компонентов африканских и западно-евразийских популяций особи тафоральта образуют отдельный кластер, занимающий промежуточное положение между современными североафриканцами (например, берберами , мозабитами , сахарцами ) и восточноафриканцами (например, афарами , оромосами , сомалийцами) . ). [87] Чон (2020), сравнивая тафоральтов иберомаврийской культуры с современными популяциями, обнаружил, что генетический компонент тафоральтов в Африке к югу от Сахары может быть лучше всего представлен современными западноафриканцами (например, йоруба). [88]
Y-хромосомная ДНК
[ редактировать ]Митохондриальная ДНК
[ редактировать ]В период голоцена , включая климатический оптимум голоцена в 8000 лет назад, африканцы, несущие гаплогруппу L2, распространились в Западной Африке, а африканцы, несущие гаплогруппу L3, распространились в Восточной Африке. [89] Будучи крупнейшей миграцией со времен миграции из Африки , миграция из стран Африки к югу от Сахары в Северную Африку произошла среди жителей Западной Африки, Центральной Африки и Восточной Африки, что привело к миграции в Европу и Азию ; следовательно, митохондриальная ДНК стран Африки к югу от Сахары была завезена в Европу и Азию. [89] В ранний период голоцена 50 % митохондриальной ДНК стран Африки к югу от Сахары было завезено в Северную Африку выходцами из Западной Африки , а остальные 50% были завезены выходцами из Восточной Африки . [89] В современный период большее количество жителей Западной Африки привезли митохондриальную ДНК из Африки к югу от Сахары в Северную Африку, чем жители Восточной Африки. [89]
Митохондриальные гаплогруппы L3, M и N встречаются среди суданских народов (например, беджа , нилоты , нубийцы , нубийцы ), у которых нет известных взаимодействий (например, история миграции/примеси) с европейцами или азиатами; не в контексте миграции после выхода из Африки. митохондриальная макрогаплогруппа L3/M/N и ее последующее развитие в отдельные митохондриальные гаплогруппы (например, гаплогруппа L3 , гаплогруппа M , гаплогруппа N ) могли возникнуть в Восточной Африке, а время, которое значительно предшествует миграционному событию из Африки, произошедшему 50 000 лет назад. [90]
Аутосомная ДНК
[ редактировать ]Медицинская ДНК
[ редактировать ]Геномы африканцев, обычно подвергающихся адаптации, представляют собой регуляторную ДНК , и многие случаи адаптации, обнаруженные среди африканцев, связаны с диетой , физиологией и эволюционным давлением со стороны патогенов. [91]
Стойкость лактазы
[ редактировать ]эпохи неолита Земледельцы , которые, возможно, проживали в Северо-Восточной Африке и на Ближнем Востоке , возможно, были источником вариантов персистенции лактазы , включая –13910*T, и могли быть впоследствии вытеснены более поздними миграциями народов. [92] Западноафриканские фулани к югу от Сахары . , североафриканские туареги и европейские земледельцы , являющиеся потомками этих неолитических земледельцев, разделяют вариант персистенции лактазы –13910*T [92] Хотя вариант Фулани -13910*T является общим для пастухов фулани и туарегов, по сравнению с вариантом туарегов он претерпел более длительный период дифференциации гаплотипов. [92] Вариант персистенции лактазы Фулани -13910*T, возможно, распространился вместе со скотоводством между 9686 и 7534 годами назад, возможно, около 8500 лет назад; Подтверждая эти временные рамки для фулани, по крайней мере, на 7500 лет назад, есть свидетельства того, что пастухи занимались доением в Центральной Сахаре . [92]
Западная Африка
[ редактировать ]ДНК архаичного человека
[ редактировать ]Архаические черты, обнаруженные в человеческих окаменелостях Западной Африки (например, окаменелости Ихо Элеру , датируемые 13 000 лет назад) и Центральной Африки (например, окаменелости Ишанго , которые датируются между 25 000 и 20 000 лет назад), возможно, развились в результате смешения архаичных людей и современных людей или может быть свидетельством существования позднесохраняющихся ранних современных людей . [79] В то время как денисовское и неандертальское происхождение у неафриканцев за пределами Африки более достоверно, архаическое человеческое происхождение у африканцев менее достоверно, и его слишком рано устанавливать с уверенностью. [79]
Древняя ДНК
[ редактировать ]По состоянию на 2017 год древняя ДНК человека не была обнаружена в регионе Западной Африки . [93] По состоянию на 2020 год древняя ДНК человека в регионе Западной Африки не обнаружена. [88]
Y-хромосомная ДНК
[ редактировать ]Восемь мужчин из Гвинеи-Бисау , два мужчины из Нигера , один мужчина из Мали и один мужчина из Кабо-Верде несли гаплогруппу A1a . [94]
Поскольку гаплогруппа D0, базальная ветвь гаплогруппы DE, была обнаружена у трех нигерийских мужчин, возможно, гаплогруппа DE , а также ее подлинии D0 и E произошли из Африки . [95]
По состоянию на 19 000 лет назад африканцы, несущие гаплогруппу E1b1a-V38 , вероятно, пересекали Сахару с востока на запад . [96] E1b1a1-M2, вероятно, возник в Западной или Центральной Африке . [97]
Митохондриальная ДНК
[ редактировать ]Около 18 000 лет назад население менде , наряду с народами Гамбии , увеличилось в численности. [98]
За 15 000 лет до нашей эры носители языка Нигер-Конго возможно, мигрировали из Сахельского региона Западной Африки вдоль реки Сенегал и завезли L2a1 в Северную Африку , в результате чего его унаследовали современные мавританские народы и берберы Туниса , . [99]
Между 11 000 и 10 000 BP населения йоруба и эсана выросла. численность [98]
Еще 11 000 лет назад жители Западной Африки к югу от Сахары, несущие макрогаплогруппу L (например, L1b1a11 , L1b1a6a, L1b1a8, L1b1a9a1, L2a1k , L3d1b1a ), возможно, мигрировали через Северную Африку и в Европу , в основном в южную Европу (например, Иберию) . ). [100]
Аутосомная ДНК
[ редактировать ]В ранний период голоцена , в 9000 году до нашей эры, родственные койсанам народы смешались с предками народа игбо , возможно, в западной Сахаре . [101] [102]
Между 2000 и 1500 годами назад носители нило-сахарского языка, возможно, мигрировали через Сахель , из Восточной Африки в Западную Африку , и смешались с нигерско-конгоязычным народом бером . [103] В 710 году н. э. родственные западноафриканскому населению (например, нигер-конго -говорящие люди бером , говорящие на банту ) и восточноафриканскому населению ( нило-сахароязычные эфиопы , нило-сахароязычные чадцы ) смешались друг с другом в северная Нигерия и северный Камерун . [104]
Фан и др. (2019) обнаружили, что народ фулани проявляет генетическую близость к изолированным афроазиатскоязычным группам в Восточной Африке , особенно к омотоязычным народам, таким как народ ари . Хотя фулани имеют почти исключительное коренное африканское происхождение (определяемое западно- и восточноафриканским происхождением), они также демонстрируют следы западно-евразийской примеси, поддерживая прародину где-то в Северной или Восточной Африке, и экспансию на запад во время неолита, возможно, вызвано приходом и экспансией западно-евразийских групп. [105] Фан и др. (2023) обнаружили, что фулани , которые на 50% связаны с амхарой и на 50% связаны с тикари , а также занимают такие регионы, как Западная Африка , Центральная Африка и Судан , в качестве кочевых пастухов , возможно, изначально были афроазиатскими носителями языка, которые впоследствии подверглись языковой замене и стали говорить на нигерско-конголезском языке . [106]
Медицинская ДНК
[ редактировать ]Геномы африканцев, обычно подвергающихся адаптации, представляют собой регуляторную ДНК , и многие случаи адаптации, обнаруженные среди африканцев, связаны с диетой , физиологией и эволюционным давлением со стороны патогенов. [91] По всей Африке к югу от Сахары генетическая адаптация (например, мутация rs334, группа крови Даффи , повышенный уровень дефицита G6PD , серповидно-клеточная анемия ) к малярии была обнаружена среди африканцев к югу от Сахары , которая, возможно, первоначально развилась около 7300 лет назад. [91] У африканцев к югу от Сахары более 90% генотипа Duffy-null . [107]
Педикулюс
[ редактировать ]В медный век и раннюю исламскую эпоху древнего Израиля возможно западноафриканцы, , мигрировали в древний Израиль и завезли головную вошь из Западной Африки . [108]
Серповидная клетка
[ редактировать ]В Зеленой Сахаре мутация серповидноклеточной анемии возникла в Сахаре. [96] или в северо-западном лесном регионе западной части Центральной Африки (например, Камерун) [96] [109] по крайней мере 7300 лет назад, [96] [109] хотя, возможно, еще 22 000 лет назад. [110] [109] Наследственный серповидно-клеточный гаплотип современных гаплотипов (например, гаплотипов Камеруна / Центральноафриканской Республики и Бенина / Сенегала ), возможно, впервые возник у предков современных жителей Западной Африки, несущих гаплогруппы E1b1a1-L485 и E1b1a1-U175 или их предковую гаплогруппу E1b1a1-M4732. . [96] Жители Западной Африки (например, йоруба и эсан из Нигерии), несущие серповидноклеточный гаплотип Бенина, возможно, мигрировали через северо-восточный регион Африки в западный регион Аравии . [96] Жители Западной Африки (например, Менде из Сьерра-Леоне), несущие сенегальский серповидноклеточный гаплотип, [111] [96] возможно, мигрировали в Мавританию (современный уровень встречаемости 77%) и Сенегал (100%); они также могли мигрировать через Сахару в Северную Африку и из Северной Африки в Южную Европу , Турцию и регион вблизи северного Ирака и южной Турции. [111] Некоторые из них, возможно, мигрировали и завезли серповидноклеточные гаплотипы Сенегала и Бенина в Басру , Ирак, где оба встречаются одинаково. [111] Жители Западной Африки, несущие серповидноклеточный гаплотип Бенина, возможно, мигрировали в северный регион Ирака (69,5%), Иорданию (80%), Ливан (73%), Оман (52,1%) и Египет (80,8%). [111]
Шистосомы
[ редактировать ]По данным Стевердинга (2020), хотя и неопределенным: вблизи Великих африканских озер шистосомы (например, S. mansoni , S. haematobium ) претерпели эволюцию. [112] Впоследствии произошло расширение вдоль Нила. [112] Из Египта присутствие шистосом могло распространиться через мигрирующих людей йоруба в Западную Африку. [112] После этого шистосомы , возможно, распространились через мигрирующих народов банту в остальную часть Африки к югу от Сахары (например, в Южную Африку , Центральную Африку ). [112]
Талассемия
[ редактировать ]Путями, пройденными караванами или путешествиями в период Альмоваридов , население (например, к югу от Сахары жители Западной Африки ) могло внедрить мутацию -29 (A → G) β-талассемии (обнаруженную в заметных количествах среди афроамериканцев ). в североафриканский регион Марокко . [113]
ДНК домашних животных
[ редактировать ]Хотя миграция Нигера и Конго могла происходить из Западной Африки в Кордофан, возможно, из Кордофана , Судан , нигер-конголезские носители в сопровождении неодомашненных цесарок в шлемах , возможно, проникли в Западную Африку, одомашнили цесарок в шлемах к 3000 г. до н. э. и через банту. расширение , перешедшее в другие части Африки к югу от Сахары (например, Центральную Африку , Восточную Африку , Южную Африку ). [114]
Центральная Африка
[ редактировать ]ДНК архаичного человека
[ редактировать ]Архаические черты, обнаруженные в человеческих окаменелостях Западной Африки (например, окаменелости Ихо Элеру , датируемые 13 000 лет назад) и Центральной Африки (например, окаменелости Ишанго , которые датируются между 25 000 и 20 000 лет назад), возможно, развились в результате смешения архаичных людей и современных людей или может быть свидетельством существования позднесохраняющихся ранних современных людей . [79] В то время как денисовское и неандертальское происхождение у неафриканцев за пределами Африки более достоверно, архаическое человеческое происхождение у африканцев менее достоверно, и его слишком рано устанавливать с уверенностью. [79]
Древняя ДНК
[ редактировать ]В 4000 году до нашей эры могла существовать популяция, которая перешла из Африки (например, Западной Африки или Западно- Центральной Африки ) через Гибралтарский пролив на Пиренейский полуостров , где смешалась африканцы и иберийцы (например, северной Португалии , южной Испании ). [115]
Камерун
[ редактировать ]Было обнаружено , что западноафриканские охотники-собиратели в регионе западной части Центральной Африки (например, Шум-Лака , Камерун ), особенно между 8000 и 3000 лет назад, были связаны с современными центральноафриканскими охотниками-собирателями (например, Бака , Бакола , Биака) . , Бедзан ). [116]
Демократическая Республика Конго
[ редактировать ]В Киндоки, в Демократической Республике Конго , было три человека, датированные протоисторическим периодом (230 г. до н. э., 150 г. до н. э., 230 г. до н. э.); один нес гаплогруппы E1b1a1a1d1a2 (E-CTS99, E-CTS99) и L1c3a1b , другой нес гаплогруппу E (E-M96, E-PF1620) , а последний нес гаплогруппы R1b1 (R-P25 1, R-M415) и L0a1b1a1 . [117] [118]
Y-хромосомная ДНК
[ редактировать ]Считается, что гаплогруппа R1b-V88 возникла в Европе и мигрировала в Африку вместе с фермерами или пастухами в период неолита, ок. 5500 г. до н.э. [119] [120] [121] [122] R1b-V88 часто встречается среди чадскоязычных народов, таких как хауса , [77] а также в Канембу , [123] Некоторый , [124] и Тубу [77] населения.
Митохондриальная ДНК
[ редактировать ]Через 150 000 лет назад африканцы (например, центральноафриканцы , восточноафриканцы ), несущие гаплогруппу L1 , разошлись. [89] Между 75 000 и 60 000 лет назад африканцы, несущие гаплогруппу L3, появились в Восточной Африке и в конечном итоге мигрировали и стали присутствовать у современных жителей Западной Африки , Центральной Африки и неафриканцев . [89] В период голоцена, включая климатический оптимум голоцена в 8000 лет назад, африканцы, несущие гаплогруппу L2, распространились в Западной Африке, а африканцы, несущие гаплогруппу L3, распространились в Восточной Африке. [89] Будучи крупнейшей миграцией со времен миграции из Африки , миграция из стран Африки к югу от Сахары в Северную Африку произошла среди жителей Западной Африки, Центральной Африки и Восточной Африки, что привело к миграции в Европу и Азию ; следовательно, митохондриальная ДНК стран Африки к югу от Сахары была завезена в Европу и Азию. [89]
Митохондриальная гаплогруппа L1c прочно связана с пигмеями, особенно с Бамбенга . группами [125] Распространенность L1c оценивалась по-разному: 100% в Ба-Коле , 97% в Аке (Ба-Бензеле) и 77% в Биаке . [126] 100% бедзанов (тикар) и 100% народа бака Габона , 97 % и Камеруна соответственно, [127] 97% в «Они придут» (97%) и 82% в «Они придут» [125] Митохондриальные гаплогруппы L2a и L0a преобладают среди бамбути . [125] [128]
Аутосомная ДНК
[ редактировать ]Генетически африканские пигмеи имеют некоторые ключевые различия между ними и народами банту . [129] [130]
Медицинская ДНК
[ редактировать ]Имеющиеся данные свидетельствуют о том, что по сравнению с другими популяциями Африки к югу от Сахары , популяции африканских пигмеев демонстрируют необычно низкие уровни экспрессии генов, кодирующих человеческий гормон роста и его рецептор, что связано с низкими в сыворотке уровнями инсулиноподобного фактора роста-1 крови и низким ростом. . [131]
Геномы африканцев, обычно подвергающихся адаптации, представляют собой регуляторную ДНК , и многие случаи адаптации, обнаруженные среди африканцев, связаны с диетой , физиологией и эволюционным давлением со стороны патогенов. [91] По всей Африке к югу от Сахары генетическая адаптация (например, мутация rs334, группа крови Даффи , повышенный уровень дефицита G6PD , серповидно-клеточная анемия ) к малярии была обнаружена среди африканцев к югу от Сахары , которая, возможно, первоначально развилась около 7300 лет назад. [91] У африканцев к югу от Сахары более 90% генотипа Duffy-null . [107] В тропических лесах Центральной Африки генетическая адаптация к факторам, не связанным с ростом (например, иммунные особенности , репродукция , функция щитовидной железы ) и низкорослостью (например, EHB1 и PRDM5 – синтез костей ; OBSCN и COX10 – мышечное развитие ; HESX1 и ASB14) . – производство/секреция гормона роста гипофиза ) был обнаружен среди охотников-собирателей тропических лесов . [91]
Восточная Африка
[ редактировать ]От региона Кении и Танзании до Южной Африки восточные африканцы, говорящие на языке банту, составляют генетическую линию с севера на юг ; кроме того, от восточной Африки до южной Африки свидетельства генетической однородности указывают на последовательный эффект основателя и события примеси , произошедшие между африканцами, говорящими на банту, и другими африканскими популяциями к тому времени, когда миграция банту распространилась на Южную Африку. [91]
ДНК архаичного человека
[ редактировать ]В то время как денисовское и неандертальское происхождение у неафриканцев за пределами Африки более достоверно, архаическое человеческое происхождение у африканцев менее достоверно, и его слишком рано устанавливать с уверенностью. [79]
Древняя ДНК
[ редактировать ]Эфиопия
[ редактировать ]В Моте , в Эфиопии , человек, живший, по оценкам, в 5-м тысячелетии до нашей эры, нес гаплогруппы E1b1 и L3x2a . [132] [133] Индивид Мота генетически связан с группами, проживающими вблизи региона Мота, и, в частности, в значительной степени генетически связан с народом Ари , особенно с кастой кузнецов этой группы. [134] [135]
Кения
[ редактировать ]В Джавуойо Рокшелтер, в округе Кисуму , Кения , собиратель позднего каменного века нес гаплогруппы E1b1b1a1b2/E-V22 и L4b2a2c . [136] [137]
В Ол Калу, в округе Ньяндаруа , Кения , скотовод эпохи пасторального неолита носил гаплогруппы E1b1b1b2b2a1/E-M293 и L3d1d . [136] [137]
В Кокурматакоре, в округе Марсабит , Кения , скотовод пасторального железного века носил гаплогруппы E1b1b1/E-M35 и L3a2a . [136] [137]
В Уайт-Рок-Пойнт, в округе Хома-Бей , Кения , жили два собирателя позднего каменного века ; один нес гаплогруппы BT (xCT) , вероятно, B и L2a4 , а другой, вероятно, нес гаплогруппу L0a2 . [136] [137]
В каменном убежище Ньяринди в Кении обитали два человека, датированные позднем каменным веком (3500 лет назад); один нес гаплогруппу L4b2a , а другой нес гаплогруппу E (E-M96, E-P162) . [117] [118]
На холме Лукения в Кении обитали два человека, датированные пасторальным неолитом (3500 лет назад); один нес гаплогруппы E1b1b1b2b (E-M293, E-CTS10880) и L4b2a2b , а другой нес гаплогруппу L0f1 . [117] [118]
В Хайракс-Хилле в Кении человек, относящийся к пасторальному неолиту (2300 лет назад), нес гаплогруппы E1b1b1b2b (E-M293, E-M293) и L5a1b . [117] [118]
В пещере Моло в Кении обитали два человека, датированные пасторальным неолитом (1500 лет назад); в то время как у одного были неопределенные гаплогруппы, у другого были гаплогруппы E1b1b1b2b (E-M293, E-M293) и L3h1a2a1 . [117] [118]
В Какапеле, в Кении , было обнаружено три особи: одна датируется поздней каменным веком (3900 лет назад), а две датируются поздним железным веком (300 лет назад, 900 лет назад); один нес гаплогруппы CT (CT-M168, CT-M5695) и L3i1 , другой нес гаплогруппу L2a1f , а последний нес гаплогруппу L2a5 . [117] [118]
В Панга-я-Саиди , в Кении , человек, возраст которого, по оценкам, на сегодняшний день составляет от 496 до 322 лет назад, нес гаплогруппы E1b1b1b2 и L4b2a2 . [138]
В Килифи , Мтвапа , в Кении , человек, живший между 1250 и 1650 годами нашей эры, нес гаплогруппу L3b1a1a . [139]
Танзания
[ редактировать ]В каменном убежище Мламбаласи в Танзании человек, датируемый периодом от 20 345 до 17 025 лет назад, нес неопределенные гаплогруппы. [140]
В пещере Гишимангеда, в округе Карату , Танзания , жили одиннадцать скотоводов пасторального неолита ; один нес гаплогруппы E1b1b1a1b2/E-V22 и HV1b1 , другой нес гаплогруппу L0a , другой нес гаплогруппу L3x1 , другой нес гаплогруппу L4b2a2b , другой нес гаплогруппы E1b1b1b2b2a1/E-M293 и L3i2 , другой нес гаплогруппу L3h1a2a1 , третий нес гаплогруппы E1b1b 1b2b2/E-V1486 , вероятно, E-M293 и L0f2a1 , а другой нес гаплогруппы E1b1b1b2b2/E-V1486 , вероятно, E-M293 и T2+150 ; хотя большинство гаплогрупп среди трех скотоводов остались неопределенными, один из них был определен как несущий гаплогруппу BT , вероятно B. , [136] [137]
В Килве, на побережье Танзании , человек, живший между 1300 и 1600 годами нашей эры, нес гаплогруппы J2a2a1a1a2a~ и L2a1h . [139]
В Линди , в Танзании , человек, датируемый между 1511 и 1664 годами нашей эры, нес гаплогруппы E1b1a1a1a2a1a3a1d~ и L0a1a2 . [139]
В пещере Макангале на Пемба острове в Танзании человек, живший, по оценкам, между 1421 и 1307 годами назад, нес гаплогруппу L0a . [138]
В Сонго Мнара , в Танзании , человек, датируемый между 1294 и 1392 годами нашей эры, нес гаплогруппы R1a и L3e3a . [139]
Уганда
[ редактировать ]В Мунсе , в Уганде , человек, живший в эпоху позднего железного века (500 лет до н. э.), нес гаплогруппу L3b1a1 . [117] [118]
Y-хромосомная ДНК
[ редактировать ]По состоянию на 19 000 лет назад африканцы, несущие гаплогруппу E1b1a-V38 , вероятно, пересекали Сахару с востока на запад . [96]
До восточные периода работорговли африканцы, несущие гаплогруппу E1b1a-M2 , распространились в Аравию , что привело к различным уровням наследования по всей Аравии (например, 2,8% Катар , 3,2% Йемен , 5,5% Объединенные Арабские Эмираты , 7,4% Оман ). [141]
Митохондриальная ДНК
[ редактировать ]Через 150 000 лет назад африканцы (например, центральноафриканцы , восточноафриканцы ), несущие гаплогруппу L1 , разошлись. [89] Через 130 000 лет назад африканцы, несущие гаплогруппу L5, разошлись в Восточной Африке . [89] Между 130 000 и 75 000 лет назад поведенческая современность возникла среди жителей Южной Африки долгосрочные взаимодействия между регионами Южной Африки и Восточной Африки . и установились [89] Между 75 000 и 60 000 лет назад африканцы, несущие гаплогруппу L3, появились в Восточной Африке и в конечном итоге мигрировали и стали присутствовать у современных жителей Западной Африки , Центральной Африки и неафриканцев . [89] В период голоцена, включая климатический оптимум голоцена в 8000 лет назад, африканцы, несущие гаплогруппу L2, распространились в Западной Африке, а африканцы, несущие гаплогруппу L3, распространились в Восточной Африке. [89] Будучи крупнейшей миграцией со времен миграции из Африки , миграция из стран Африки к югу от Сахары в Северную Африку произошла среди жителей Западной Африки, Центральной Африки и Восточной Африки, что привело к миграции в Европу и Азию ; следовательно, митохондриальная ДНК стран Африки к югу от Сахары была завезена в Европу и Азию. [89] В ранний период голоцена 50 % митохондриальной ДНК стран Африки к югу от Сахары было завезено в Северную Африку выходцами из Западной Африки , а остальные 50% были завезены выходцами из Восточной Африки . [89] В современный период большее количество жителей Западной Африки привезли митохондриальную ДНК из Африки к югу от Сахары в Северную Африку, чем жители Восточной Африки. [89] Между 15 000 и 7 000 лет назад 86% митохондриальной ДНК стран Африки к югу от Сахары было завезено в Юго-Западную Азию выходцами из Восточной Африки , в основном из региона Аравии , которые составляют 50% митохондриальной ДНК стран Африки к югу от Сахары в современной Юго -Западной Азии . [89] В современный период 68% митохондриальной ДНК стран Африки к югу от Сахары было введено выходцами из Восточной Африки, а 22% - жителями Западной Африки, что составляет 50% митохондриальной ДНК стран Африки к югу от Сахары в современной Юго-Западной Азии. [89]
Аутосомная ДНК
[ редактировать ]Во всех районах Мадагаскара среднее происхождение малагасийцев составило 4% западноевразийцев , 37% австронезийцев и 59% банту . [66]
Медицинская ДНК
[ редактировать ]Геномы африканцев, обычно подвергающихся адаптации, представляют собой регуляторную ДНК , и многие случаи адаптации, обнаруженные среди африканцев, связаны с диетой , физиологией и эволюционным давлением со стороны патогенов. [91] По всей Африке к югу от Сахары генетическая адаптация (например, мутация rs334, группа крови Даффи , повышенный уровень дефицита G6PD , серповидно-клеточная анемия ) к малярии была обнаружена среди африканцев к югу от Сахары , которая, возможно, первоначально развилась около 7300 лет назад. [91] У африканцев к югу от Сахары более 90% генотипа Duffy-null . [107] В высокогорной Эфиопии реакцией на генетическая адаптация (например, rs10803083, SNP, связанный со скоростью и функцией гемоглобина ; BHLHE41, ген, связанный с циркадным ритмом и гипоксию ; EGNL1, ген, тесно связанный с кислородным гомеостазом у млекопитающих) к гипоксии у народа амхара было обнаружено низкое атмосферное давление , которое, возможно, возникло за последние 5000 лет. [91] В Танзании генетическая адаптация (например, большее количество генов амилазы , чем у африканских популяций, потребляющих продукты с низким содержанием крахмала) была обнаружена у народа хадза из-за пищевого рациона, который особенно включает потребление клубней . [91]
Южная Африка
[ редактировать ]От региона Кении и Танзании до Южной Африки восточные африканцы, говорящие на языке банту, составляют генетическую линию с севера на юг ; кроме того, от восточной Африки до южной Африки свидетельства генетической однородности указывают на последовательный эффект основателя и события примеси , произошедшие между африканцами, говорящими на банту, и другими африканскими популяциями к тому времени, когда миграция банту распространилась на Южную Африку. [91]
ДНК архаичного человека
[ редактировать ]В то время как денисовское и неандертальское происхождение у неафриканцев за пределами Африки более достоверно, архаическое человеческое происхождение у африканцев менее достоверно, и его слишком рано устанавливать с уверенностью. [79]
Древняя ДНК
[ редактировать ]Три позднего каменного века охотника-собирателя несли древнюю ДНК, похожую на ДНК охотников-собирателей, говорящих на койсанском языке . [142] До миграции банту в этот регион, о чем свидетельствует древняя ДНК из Ботсваны , из Восточной Африки пастухи мигрировали в Южную Африку. [142] Из четырех века банту земледельцев- западноафриканского происхождения железного двое более ранних земледельцев несли древнюю ДНК, похожую на ДНК народов тсонга и венда , а два более поздних земледельца несли древнюю ДНК, похожую на ДНК людей нгуни ; это указывает на то, что в общей миграции банту происходили различные перемещения народов, что привело к усилению взаимодействия и смешения между народами , говорящими на банту , и народами, говорящими на койсанском языке. [142]
Ботсвана
[ редактировать ]В Нкоме, в Ботсване , человек, живший в раннем железном веке (900 лет до н. э.), нес гаплогруппу L2a1f . [117] [118]
В Таукоме, в Ботсване , человек, датируемый ранним железным веком (1100 лет назад), нес гаплогруппы E1b1a1 (E-M2, E-Z1123) и L0d3b1 . [117] [118]
В Ксаро, в Ботсване , обитали два человека, датированные ранним железным веком (1400 г. до н. э.); один нес гаплогруппы E1b1a1a1c1a и L3e1a2 , а другой нес гаплогруппы E1b1b1b2b (E-M293, E-CTS10880) и L0k1a2 . [117] [118]
Малави
[ редактировать ]В каменном убежище Фингира в Малави человек, датируемый периодом между 6179 и 2341 годами до нашей эры, нес гаплогруппы B2 и L0d1 . [140]
В Ченчерере, в Малави , человек, возраст которого, по оценкам, составляет от 5400 до 4800 лет назад, нес гаплогруппу L0k2 . [138]
В каменном убежище Хора 1 в Малави человек, датируемый между 16 897 и 15 827 годами назад, нес гаплогруппы B2b и L5b . [140]
ЮАР
[ редактировать ]В Дунсайде , в Южной Африке , человек, живший на сегодняшний день между 2296 и 1910 годами до нашей эры, был носителем гаплогруппы L0d2 . [143] [144]
В Баллито заливе , Южная Африка , человек, живший, по оценкам, между 1986 и 1831 годами, нес гаплогруппы A1b1b2 и L0d2c1 . [143] [144]
В каменном убежище Калемба в Замбии человек, датируемый между 5285 и 4975 годами до нашей эры, нес гаплогруппу L0d1b2b . [140]
Y-хромосомная ДНК
[ редактировать ]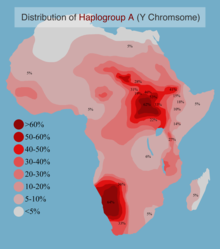

Различные исследования Y-хромосомы показывают, что народ сан несет в себе некоторые из наиболее расходящихся (старейших) гаплогрупп Y-хромосомы человека . Эти гаплогруппы представляют собой особые подгруппы гаплогрупп A и B , двух самых ранних ветвей дерева Y-хромосомы человека . [145] [146] [147]
Митохондриальная ДНК
[ редактировать ]За 200 000 лет назад африканцы (например, койсаны из Южной Африки), несущие гаплогруппу L0, отделились от других африканцев, несущих гаплогруппу L1'6 , которые, как правило, проживали к северу от Южной Африки . [89] Между 130 000 и 75 000 лет назад среди жителей Южной Африки возникла поведенческая современность долгосрочные взаимодействия между регионами Южной Африки и Восточной Африки . и установились [89]
Исследования митохондриальной ДНК также свидетельствуют о том, что сан несут высокие частоты самых ранних ветвей гаплогрупп в дереве митохондриальной ДНК человека. Эта ДНК наследуется только от матери. Самая дивергентная (самая старая) митохондриальная гаплогруппа, L0d , была идентифицирована с самой высокой частотой в южноафриканских группах сан. [145] [148] [149] [150]
Аутосомная ДНК
[ редактировать ]Хенн и др. (2011) обнаружили, что хомани сан, а также народы сандаве и хадза в Танзании были наиболее генетически разнообразными из всех изученных живых людей. Такая высокая степень генетического разнообразия намекает на происхождение анатомически современного человека . [151] [152]
Медицинская ДНК
[ редактировать ]Среди древней ДНК трех охотников-собирателей, имеющих генетическое сходство с народом сан , и четырех земледельцев железного века , их SNP показали, что они несли варианты устойчивости к сонной болезни и Plasmodium vivax . [153] В частности, двое из четырех земледельцев железного века имели варианты устойчивости к сонной болезни, а трое из четырех земледельцев железного века имели отрицательные по Даффи варианты устойчивости к малярии . [153] В отличие от земледельцев железного века, из числа охотников-собирателей, родственных сану, шестилетний мальчик мог умереть от шистосомоза . [153] В Ботсване мужчина, датируемый 1400 годом до нашей эры, возможно, также был носителем отрицательного варианта Даффи по устойчивости к малярии. [153]
Геномы африканцев, обычно подвергающихся адаптации, представляют собой регуляторную ДНК , и многие случаи адаптации, обнаруженные среди африканцев, связаны с диетой , физиологией и эволюционным давлением со стороны патогенов. [91] По всей Африке к югу от Сахары генетическая адаптация (например, мутация rs334, группа крови Даффи , повышенный уровень дефицита G6PD , серповидно-клеточная анемия ) к малярии была обнаружена среди африканцев к югу от Сахары , которая, возможно, первоначально развилась около 7300 лет назад. [91] У африканцев к югу от Сахары более 90% генотипа Duffy-null . [107] В регионе пустыни Калахари различные возможные генетические адаптации (например, адипонектин , индекс массы тела , обмен веществ были обнаружены в Африке среди народа хомани ) . [91] У африканцев к югу от Сахары более 90% генотипа Duffy-null . [107] В Южной Африке генетическая адаптация (например, rs28647531 на хромосоме 4q22 ) и сильная восприимчивость к туберкулезу была обнаружена среди цветных . [91]
Недавнее африканское происхождение современных людей
[ редактировать ]Между 500 000 и 300 000 лет назад анатомически современные люди могли появиться в Африке. [154] Поскольку африканцы (например, Y-хромосомный Адам , Митохондриальная Ева ) мигрировали из мест своего происхождения в Африке в другие места Африки, и поскольку время расхождения восточноафриканских , центральноафриканских и западноафриканских линий схоже со временем Из-за расхождения южноафриканской линии недостаточно доказательств для определения конкретного региона происхождения людей в Африке . [79] За 100 000 лет назад анатомически современные люди мигрировали из Африки в Евразию . [155] Впоследствии, спустя десятки тысяч лет, предки всех современных евразийцев мигрировали из Африки в Евразию и в конечном итоге смешались с денисовцами и неандертальцами . [155]
Археологические и ископаемые свидетельства подтверждают африканское происхождение homo sapiens и современность поведения . [156] модели, отражающие панафриканское происхождение (несколько мест происхождения в Африке) и эволюцию современного человека . Были разработаны [156] Поскольку идея «современности» становится все более проблематизированной, исследования «начали распутывать, что подразумевается под «современным» генетическим происхождением, морфологией скелета и поведением, признавая, что они вряд ли образуют единый пакет». [156]
По сравнению с неафриканским геномом, африканский геном имеет примерно на 25% большее количество полиморфизмов . [91] или в 3-5 раз больше, [107] а генетические варианты , которые редки за пределами Африки, в большом количестве встречаются внутри Африки. [91] Установлено, что большая часть генетического разнообразия, обнаруженного среди неафриканцев, в целом представляет собой подмножество генетического разнообразия, обнаруженного среди африканцев. [91] Геномы африканцев, обычно подвергающихся адаптации, представляют собой регуляторную ДНК , и многие случаи адаптации, обнаруженные среди африканцев, связаны с диетой , физиологией и эволюционным давлением со стороны патогенов. [91] По всей Африке к югу от Сахары генетическая адаптация (например, мутация rs334, группа крови Даффи , повышенный уровень дефицита G6PD , серповидно-клеточная анемия ) к малярии была обнаружена среди африканцев к югу от Сахары , которая, возможно, первоначально развилась около 7300 лет назад. [91] По всей Африке различные генетические адаптации (например, аполипопротеин L1 (APOL1): устойчивость гаплотипов G1 и G2 к трипаносомозу и повышенному риску заболеваний почек ; гены человеческого лейкоцитарного антигена (HLA); главный комплекс гистосовместимости (MHC)) к ВИЧ-1 , оспе Среди африканцев трипаносомоз (африканская сонная болезнь) туберкулез обнаружены и . [91] Биомедицинские тесты на конкретные генетические варианты (например, rs1799853 в гене CYP2C9 ), которые были одобрены Управлением по контролю за продуктами и лекарствами США и предназначены для подтверждения правильного назначения варфарина , оказываются все более нерелевантными для африканцев, поскольку эти варианты редко встречается в Африке. [91] поскольку коэффициент частоты учитывается при рассмотрении и принятии решения вариантов о патогенности и обобщаемых полигенных показателях . Современные клинические классификации патогенности генетических вариантов оказываются неадекватными из-за отсутствия генетического разнообразия в биомедицинских исследованиях, [91] Fan et al (2023) недавно обнаружили около 5,3 миллиона уникальных генетических вариантов в 180 африканских популяциях охотников-собирателей, а среди существующих классификаций вариантов, которые , вероятно, являются «патогенными», около 29% (44/154) из этих «патогенных» классифицируются. Было обнаружено, что такие варианты часто встречаются среди африканских охотников-собирателей. [91]
См. также
[ редактировать ]Примечания
[ редактировать ]- ^ Линия, ведущая к Кхое-Сан, является базовой для всех других человеческих линий с предполагаемым временем расхождения 300–200 тысяч лет назад (например, Джу|'Хоан с самым низким уровнем недавней примеси разошлась ~ 270 ± 12 тысяч лет назад).
- ^ Впоследствии мбути (RHG) отошли на ~ 220 ± 10 тысяч лет назад от всех других человеческих линий, образовав вторую базальную линию (Schlebusch et al. 2020) (рис. 1).
- ^ Во-первых, современное происхождение североафриканцев характеризуется автохтонным магрибским компонентом, связанным с палеолитической обратной миграцией в Африку из Евразии. ... Этот результат позволяет предположить, что иберомаврские популяции в Северной Африке были связаны с палеолитическими людьми в Леванте, но также и о том, что миграции африканского происхождения из стран к югу от Сахары достигли Магриба во время плейстоцена. ... Этот результат согласуется с нашим предыдущим выводом о том, что кушитское происхождение образовалось в результате смешения нило-сахарских и арабских предков39. ... Хотя эти результаты показывают, что население, связанное с левант-неолитом, внесло решающий вклад в происхождение современных восточноафриканцев (Lazaridis et al., 2016), современные кушитские носители, такие как сомалийцы, не могут быть просто приспособлены как имеющий происхождение из Танзании_Люксманда_3100BP. Наиболее подходящая модель для сомалийцев включает происхождение Танзании_Luxmanda_3100BP, происхождение, связанное с динка, и 16% ± 3% происхождение, связанное с иранским неолитом (p = 0,015). Это говорит о том, что предки, относящиеся к иранскому неолиту, появились в Восточной Африке после более раннего потока генов, связанного с неолитическими популяциями Леванта, сценарий, который становится более правдоподобным благодаря генетическим свидетельствам примеси предков, связанных с иранским неолитом, по всему Леванту ко времени бронзового века (Lazaridis et al., 2016), а в Древнем Египте – железного века (Schuenemann et al., 2017).
- ^ Это может либо указывать на глубокую структуру населения, в которой EAHG и южные группы охотников-собирателей прослеживают некоторые из своих предков до базовой центральноафриканской линии RHG (Lipson et al. 2020, 2022), либо на поток генов между южноафриканскими и центральноафриканскими собирателями, как указано. благодаря четкому паттерну совместного использования аллелей между !Xun/Ju|'Hoan и Mbuti (Scheinfeldt et al. 2019; Bergström et al. 2020; Schlebusch et al. 2020). ... В настоящее время недостаточно данных, чтобы оценить время (даже более древнее) восточноафриканско-омотской дивергенции.
- ^ Для пары предков из Западной и Западной и Центральной Африки точечная оценка времени расхождения составила 6900 лет назад. ... Западноафриканское происхождение является преобладающим среди населения территории вокруг Сенегала и Гамбии, тогда как западно-центральноафриканское происхождение преобладает среди населения территории вокруг Нигерии. ... При сравнении двух манденка и одного гамбийца с двумя эсанами и одним йоруба время разделения оценивалось в <4600 лет назад, что, как ожидается, будет заниженной оценкой по сравнению со временем, основанным на FST, из-за присутствия 0–11,1. % западно-центральноафриканского происхождения у жителей Западной Африки и 26,7–35,0% западноафриканского происхождения у жителей Западной Центральной Африки. ... В свою очередь, восточноафриканское происхождение, характерное для современных нилотов, и общий предок западных и западно-центральноафриканских предков произошли от общего предка 18 000 лет назад на основе разложения FST или <13 800 лет назад на основе msmc. анализ двух динка по сравнению либо с одним гамбийцем и одним манденка, либо с двумя эсанами. Последнее время относительно недооценено из-за присутствия 22,6–26,1% западного или западно-центральноафриканского происхождения у восточноафриканцев. Этот общий предок, вероятно, существовал в долине Нила. ... В настоящее время недостаточно данных, чтобы оценить время (даже более древнее) восточноафриканско-омотской дивергенции.
Ссылки
[ редактировать ]- ^ Осборн А.Х. и др. (октябрь 2008 г.). «Влажный коридор через Сахару для миграции ранних современных людей из Африки 120 000 лет назад» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (43): 16444–16447. Бибкод : 2008PNAS..10516444O . дои : 10.1073/pnas.0804472105 . ПМЦ 2575439 . ПМИД 18936490 . S2CID 10418009 .
- ^ Дрейк Н., Бриз П. (2016). «Изменение климата и современное заселение Сахары человеком из MIS 6-2». Африка из МИС 6-2 . Палеобиология и палеоантропология позвоночных. Африка из МИС 6-2. стр. 103–122. дои : 10.1007/978-94-017-7520-5_6 . ISBN 978-94-017-7519-9 . S2CID 131383927 .
- ^ Эль-Шенави М.И. и др. (2018). «Спелеотемы свидетельствуют об озеленении Сахары и его последствиях для раннего расселения людей из стран Африки к югу от Сахары». Четвертичные научные обзоры . 188 : 67–76. Бибкод : 2018QSRv..188...67E . doi : 10.1016/j.quascirev.2018.03.016 . S2CID 134694090 .
- ^ Шееле Дж. (август 2016 г.). Перекресток регионов: Сахара . Оксфордские справочники в Интернете. дои : 10.1093/oxfordhb/9780199935369.013.18 . ISBN 978-0-19-993536-9 .
- ^ Виппель С (2020). «Сахара как мост, а не барьер: эссе и рецензия на книгу о последних трансрегиональных перспективах» . Новая политическая литература . 65 (3): 449–472. дои : 10.1007/s42520-020-00318-y . S2CID 224855920 .
- ^ Перейти обратно: а б Хольфельдер Н., Бретон Г., Сьёдин П., Якобссон М. (апрель 2021 г.). «Глубокая история населения Африки» . Молекулярная генетика человека . 30 (R1): R2–R10. дои : 10.1093/hmg/ddab005 . ПМЦ 8117439 . ПМИД 33438014 .
- ^ Фан С., Келли Д.Э., Белтрам М.Х., Хансен М.Е., Маллик С., Ранчиаро А. и др. (апрель 2019 г.). «Эволюционная история Африки выведена на основе данных последовательности всего генома 44 коренных африканских популяций» . Геномная биология . 20 (1): 82. дои : 10.1186/s13059-019-1679-2 . ПМК 6485071 . ПМИД 31023338 .
- ^ Перейти обратно: а б Бейкер Дж.Л., Ротими С.Н., Шрайнер Д. (8 мая 2017 г.). «Происхождение человека коррелирует с языком и показывает, что раса не является объективным геномным классификатором» . Научные отчеты . 7 (1): 1572. Бибкод : 2017НатСР...7.1572Б . дои : 10.1038/s41598-017-01837-7 . ISSN 2045-2322 . ПМЦ 5431528 . ПМИД 28484253 .
- ^ «Великая африканская миграция генов» . www.cosmosmagazine.com . 29 октября 2020 г. Проверено 11 января 2022 г.
- ^ Перейти обратно: а б Ананьо Чоудхури, Шон Арон, Дхрити Сенгупта, Скотт Хейзелхерст, Мишель Рамзи (1 августа 2018 г.). «Генетическое разнообразие Африки дает новое понимание истории эволюции и местных адаптаций» . Academic.oup.com . Проверено 21 октября 2022 г.
- ^ Пфенниг, Петерсен, Качамбва, Лашанс (29 марта 2023 г.). «Эволюционная генетика и примесь в африканских популяциях» . Геномная биология и эволюция . 15 (4). дои : 10.1093/gbe/evad054 . ПМЦ 10118306 . ПМИД 36987563 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Фан С., Келли Д.Э., Белтрам М.Х., Хансен М.Е., Маллик С., Ранчиаро А. и др. (26 апреля 2019 г.). «Эволюционная история Африки выведена на основе данных последовательности всего генома 44 коренных африканских популяций» . Геномная биология . 20 (1): 82. дои : 10.1186/s13059-019-1679-2 . ISSN 1474-760X . ПМК 6485071 . ПМИД 31023338 .
- ^ Скоглунд П., Томпсон Дж.К., Прендергаст М.Е., Миттник А., Сирак К., Хайдиньяк М. и др. (сентябрь 2017 г.). «Реконструкция структуры доисторического населения Африки» . Клетка . 171 (1): 59–71.e21. дои : 10.1016/j.cell.2017.08.049 . ISSN 0092-8674 . ПМЦ 5679310 . ПМИД 28938123 .
- ^ Перейти обратно: а б с Висенте М., Шлебуш К.М. (1 июня 2020 г.). «История населения Африки: взгляд на древнюю ДНК» . Текущее мнение в области генетики и развития . Генетика происхождения человека. 62 : 8–15. дои : 10.1016/j.где.2020.05.008 . ISSN 0959-437X . ПМИД 32563853 . S2CID 219974966 .
- ^ Перейти обратно: а б Бейкер Дж.Л., Ротими С.Н., Шрайнер Д. (8 мая 2017 г.). «Происхождение человека коррелирует с языком и показывает, что раса не является объективным геномным классификатором» . Научные отчеты . 7 (1): 1572. Бибкод : 2017НатСР...7.1572Б . дои : 10.1038/s41598-017-01837-7 . ISSN 2045-2322 . ПМЦ 5431528 . ПМИД 28484253 .
- ^ Скоглунд П., Томпсон Дж.К., Прендергаст М.Е., Миттник А., Сирак К., Хайдиньяк М. и др. (сентябрь 2017 г.). «Реконструкция структуры доисторического населения Африки» . Клетка . 171 (1): 59–71.e21. дои : 10.1016/j.cell.2017.08.049 . ISSN 0092-8674 . ПМЦ 5679310 . ПМИД 28938123 .
- ^ Липсон М., Рибо И., Маллик С., Роланд Н., Олальде И., Адамски Н. и др. (январь 2020 г.). «Древние западноафриканские собиратели в контексте истории населения Африки» . Природа . 577 (7792): 665–670. Бибкод : 2020Natur.577..665L . дои : 10.1038/s41586-020-1929-1 . ISSN 1476-4687 . ПМЦ 8386425 . ПМИД 31969706 .
Западноафриканская клада отличается примесью глубокого происхождения, которую можно смоделировать как сочетание современного человеческого и архаичного происхождения. Современный человеческий компонент расходится почти в той же точке, что и охотники-собиратели Центральной и Южной Африки, и предположительно связан с глубоким источником, способствующим происхождению Моты, тогда как архаический компонент расходится близко к расколу между неандертальцами и современными людьми (раздел 3 «Дополнительная информация»). ).
- ^ Кусума П., Брукато Н., Кокс М.П., Пьеррон Д., Разафиндразака Х., Аделаар А. и др. (18 мая 2016 г.). «Контраст лингвистического и генетического происхождения азиатских исходных популяций малагасийцев» . Научные отчеты . 6 : 26066. Бибкод : 2016NatSR...626066K . дои : 10.1038/srep26066 . ISSN 2045-2322 . ПМК 4870696 . ПМИД 27188237 .
- ^ Бленч Р. (2006). Археология, язык и африканское прошлое . АльтаМира Пресс. ISBN 978-0-7591-0465-5 .
- ^ Ван С., Зёлльнер С., Розенберг Н.А. (август 2012 г.). «Количественное сравнение сходства между генами и географией человеческих популяций во всем мире» . ПЛОС Генетика . 8 (8): e1002886. дои : 10.1371/journal.pgen.1002886 . ПМЦ 3426559 . ПМИД 22927824 .
- ^ Мэннинг К., Тимпсон А. (1 октября 2014 г.). «Демографический ответ на изменение климата в Сахаре в голоцене» . Четвертичные научные обзоры . 101 : 28–35. Бибкод : 2014QSRv..101...28M . doi : 10.1016/j.quascirev.2014.07.003 . ISSN 0277-3791 . S2CID 54923700 .
- ^ Бленч Р. (2006). Археология, язык и африканское прошлое . АльтаМира Пресс. ISBN 978-0-7591-0465-5 .
- ^ Бленч Р. (сентябрь 2016 г.). Можем ли мы посетить могилы первых говорящих на нигерско-конголезском языке? (PDF) . 2-й Международный конгресс: На пути к Прото-Нигер-Конго: сравнение и реконструкция. Париж.
- ^ Воссен Р., Dimensions GJ (13 марта 2020 г.). Оксфордский справочник африканских языков Издательство Оксфордского университета. ISBN 978-0-19-960989-5 .
- ^ Висенте М., Якобссон М., Эббесен П., Шлебуш К.М. (сентябрь 2019 г.). «Генетическая близость среди охотников-собирателей Южной Африки и влияние смешения популяций фермеров и пастухов» . Молекулярная биология и эволюция . 36 (9): 1849–1861. дои : 10.1093/molbev/msz089 . ПМЦ 6735883 . ПМИД 31288264 .
- ^ Перейти обратно: а б ван де Лоосдрехт М., Бузуггар А., Хамфри Л., Пост С., Бартон Н., Аксиму-Петри А. и др. (4 мая 2018 г.). «Плейстоценовые геномы Северной Африки связывают человеческие популяции Ближнего Востока и Африки к югу от Сахары» . Наука . 360 (6388): 548–552. Бибкод : 2018Sci...360..548V . дои : 10.1126/science.aar8380 . ISSN 0036-8075 . ПМИД 29545507 . S2CID 206666517 .
- ^ Лопес С., ван Дорп Л., Хелленталь Г. (21 апреля 2016 г.). «Расселение людей из Африки: длительные дебаты» . Эволюционная биоинформатика онлайн . 11 (Приложение 2): 57–68. дои : 10.4137/EBO.S33489 . ПМЦ 4844272 . ПМИД 27127403 .
- ^ Скерри Э.М., Томас М.Г., Маника А., Ганц П., Сток Дж.Т., Стрингер С. и др. (август 2018 г.). «Эволюционировал ли наш вид в разделенных популяциях по всей Африке, и почему это имеет значение?» . Тенденции в экологии и эволюции . 33 (8): 582–594. Бибкод : 2018TEcoE..33..582S . дои : 10.1016/j.tree.2018.05.005 . ПМК 6092560 . ПМИД 30007846 .
- ^ «Один вид, множество происхождений» . www.shh.mpg.de. Проверено 11 января 2022 г.
- ^ Валлини Л. и др. (7 апреля 2022 г.). «Генетика и материальная культура поддерживают неоднократные экспансии в палеолитическую Евразию из популяционного центра из Африки» . Academic.oup.com . Проверено 21 февраля 2023 г.
- ^ Халласт П., Агджоян А., Балановский О., Сюэ Ю., Тайлер-Смит С. (февраль 2021 г.). «Происхождение современных неафриканских Y-хромосом человека из Юго-Восточной Азии» . Генетика человека . 140 (2): 299–307. дои : 10.1007/s00439-020-02204-9 . ПМЦ 7864842 . ПМИД 32666166 .
- ^ Дурвасула А., Шанкарараман С. (февраль 2020 г.). «Восстановление сигналов призрачной архаической интрогрессии в африканском населении» . Достижения науки . 6 (7): eaax5097. Бибкод : 2020SciA....6.5097D . дои : 10.1126/sciadv.aax5097 . ПМК 7015685 . ПМИД 32095519 . «Неафриканские популяции (ханьцы в Пекине и жители Юты северного и западноевропейского происхождения) также демонстрируют аналогичные закономерности в CSFS, что позволяет предположить, что компонент архаического происхождения был общим до разделения африканского и неафриканского населения... Одна из интерпретаций недавнего времени интрогрессии, которую мы документируем, заключается в том, что архаические формы сохранялись в Африке до недавнего времени. С другой стороны, архаическая популяция могла раньше интрогрессировать в современную человеческую популяцию, которая затем впоследствии скрестилась с предками существующих у нас популяций. Анализируемые здесь модели не являются взаимоисключающими, и вполне вероятно, что история африканских популяций включает в себя генетический вклад множества дивергентных популяций, о чем свидетельствует большой эффективный размер популяции, связанный с интрогрессирующей архаической популяцией... Учитывая неопределенность в наших оценках времени интрогрессии, мы задались вопросом, может ли совместный анализ CSFS геномов CEU (жителей штата Юта с северным и западноевропейским происхождением) и YRI обеспечить дополнительное разрешение. В рамках модели C мы смоделировали интрогрессию до и после разделения между африканскими и неафриканскими популяциями и наблюдали качественные различия между двумя моделями в высокочастотных аллельных интервалах CSFS в африканских и неафриканских популяциях (рис. S40). ). Используя ABC для совместного сопоставления высокочастотных аллелей CSFS в CEU и YRI (определяемых как частота более 50%), мы обнаруживаем, что нижний предел 95% вероятного интервала времени интрогрессии старше, чем смоделированный раскол между CEU и YRI (2800 против 2155 поколений BP), указывающий на то, что по крайней мере часть архаических линий, наблюдаемых в YRI, также является общей с CEU...»
- ^ [1] Архивировано 7 декабря 2020 года в дополнительных материалах Wayback Machine для восстановления сигналов призрачной архаической интрогрессии в африканских популяциях», раздел «S8.2». «Мы смоделировали данные, используя те же априорные данные, что и в разделе S5.2, но вычислили спектр. как для YRI [западноафриканская йоруба], так и для CEU [население европейского происхождения]. Мы обнаружили, что наиболее подходящими параметрами были архаичное время разделения 27 000 поколений назад (95% HPD: 26 000–28 000), доля примеси 0,09 (95% HPD: 0,04-0,17), время примеси 3000 поколений назад (95% HPD) : 2800–3400), а эффективная численность популяции составляет 19 700 особей (95% HPD: 19 300–20 200). Мы обнаружили, что нижняя граница времени примеси находится дальше, чем смоделированный раскол между CEU и YRI (2155 поколений назад), что дает некоторые доказательства в пользу события, предшествовавшего выходу из Африки. Эта модель предполагает, что многие популяции за пределами Африки также должны содержать гаплотипы, возникшие в результате этого интрогрессионного события, хотя обнаружение затруднено, поскольку многие методы используют несмешанные внешние группы для обнаружения интрогрессивных гаплотипов [Browning et al., 2018, Skov et al., 2018, Durvasula and Sankararaman , 2019] (5, 53, 22). Также возможно, что некоторые из этих гаплотипов были потеряны во время «узкого места» за пределами Африки».
- ^ Дурвасула А., Шанкарараман С. (февраль 2020 г.). «Восстановление сигналов призрачной архаической интрогрессии в африканском населении» . Достижения науки . 6 (7): eaax5097. Бибкод : 2020SciA....6.5097D . дои : 10.1126/sciadv.aax5097 . ПМК 7015685 . ПМИД 32095519 .
- ^ Чен Л., Вольф А.Б., Фу В., Ли Л., Акей Дж.М. (февраль 2020 г.). «Выявление и интерпретация очевидного неандертальского происхождения у африканских людей» . Клетка . 180 (4): 677–687.e16. дои : 10.1016/j.cell.2020.01.012 . ПМИД 32004458 . S2CID 210955842 .
- ^ Пфенниг А., Петерсен Л.Н., Качамбва П., Лашанс Дж. (6 апреля 2023 г.). «Эволюционная генетика и примесь в африканских популяциях» . Геномная биология и эволюция . 15 (4): evad054. дои : 10.1093/gbe/evad054 . ISSN 1759-6653 . ПМЦ 10118306 . ПМИД 36987563 .
- ^ Басби ГБ, Band G, Си Ле Q, Джаллоу М, Бугама Э, Мангано В.Д. и др. (июнь 2016 г.). «Примесь в Африке к югу от Сахары и внутри нее» . электронная жизнь . 5 . дои : 10.7554/eLife.15266 . ПМЦ 4915815 . ПМИД 27324836 .
- ^ Серра-Видаль Дж., Лукас-Санчес М., Фадлауи-Зид К., Бекада А., Заллуа П., Комас Д. (18 ноября 2019 г.). «Неоднородность палеолитической непрерывности населения и неолитической экспансии в Северной Африке» . Современная биология . 29 (22): 3953–3959.e4. Бибкод : 2019CBio...29E3953S . дои : 10.1016/j.cub.2019.09.050 . ISSN 0960-9822 . ПМИД 31679935 . S2CID 204972040 .
- ^ Перейти обратно: а б Пикрелл Дж.К., Паттерсон Н., Лох П.Р., Липсон М., Бергер Б., Стоункинг М. и др. (февраль 2014 г.). «Древнее западно-евразийское происхождение в южной и восточной Африке» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (7): 2632–2637. arXiv : 1307.8014 . Бибкод : 2014PNAS..111.2632P . дои : 10.1073/pnas.1313787111 . ПМЦ 3932865 . ПМИД 24550290 .
- ^ Перейти обратно: а б с Чоудхури А., Арон С., Сенгупта Д., Хейзелхерст С., Рамзи М. (август 2018 г.). «Генетическое разнообразие Африки дает новое понимание истории эволюции и местных адаптаций» . Молекулярная генетика человека . 27 (С2): Р209–Р218. дои : 10.1093/hmg/ddy161 . ПМК 6061870 . ПМИД 29741686 .
- ^ Даймонд Дж., Беллвуд П. (апрель 2003 г.). «Фермеры и их языки: первые расширения». Наука . 300 (5619): 597–603. Бибкод : 2003Sci...300..597D . дои : 10.1126/science.1078208 . ПМИД 12714734 . S2CID 13350469 .
- ^ Кэмпбелл Л. (2021). Историческая лингвистика, четвертое издание . Массачусетский технологический институт Пресс. стр. 399–400. ISBN 978-0-262-54218-0 .
- ^ Джарви, Холл (2005). Переход к современности: очерки власти, богатства и веры . Издательство Кембриджского университета. п. 27. ISBN 978-0-521-02227-9 .
- ^ Шираи Н (2010). Археология первых земледельцев-скотоводов в Египте: новый взгляд на фаюмский эпипалеолит и неолит . Издательство Лейденского университета. ISBN 978-90-485-1269-0 . OCLC 852516752 .
- ^ Перейти обратно: а б с Ходжсон Дж.А., Маллиган С.Дж., Аль-Меери А., Раум Р.Л. (июнь 2014 г.). «Ранняя обратная миграция в Африку на Африканский Рог» . ПЛОС Генетика . 10 (6): e1004393. дои : 10.1371/journal.pgen.1004393 . ПМК 4055572 . ПМИД 24921250 .
- ^ Бленч Р. (2006). Археология, язык и африканское прошлое . АльтаМира Пресс. стр. 150–163. ISBN 978-0-7591-0466-2 .
- ^ Перейти обратно: а б Эрет С (1979). «О древности земледелия в Эфиопии» . Журнал африканской истории . 20 (2): 161–177. дои : 10.1017/S002185370001700X . JSTOR 181512 . S2CID 162986221 – через JSTOR.
- ^ Нёт В. (2011). Истоки семиозиса: эволюция знаков в природе и культуре . Де Грюйтер Мутон. п. 293. ИСБН 978-1-134-81623-1 .
- ^ Перейти обратно: а б Шрайнер Д. (2018). «Повторный анализ данных последовательности всего генома 279 древних евразийцев выявил существенную наследственную гетерогенность» . Границы генетики . 9 : 268. дои : 10.3389/fgene.2018.00268 . ISSN 1664-8021 . ПМК 6062619 . ПМИД 30079081 .
и компонент Африки к югу от Сахары у натуфийцев, который локализуется на территории современной южной Эфиопии.
- ^ Андерхилл П.А., Пассарино Г., Лин А.А., Шен П., Мирасон Лар М., Фоли Р.А. и др. (январь 2001 г.). «Филогеография бинарных гаплотипов Y-хромосомы и происхождение современных человеческих популяций» . Анналы генетики человека . 65 (Часть 1): 43–62. дои : 10.1046/j.1469-1809.2001.6510043.x . ISSN 0003-4800 . ПМИД 11415522 . S2CID 9441236 .
- ^ Ибрагим М.Е. (26 апреля 2021 г.). «Генетическое разнообразие суданцев: понимание происхождения и последствий для здоровья» . Молекулярная генетика человека . 30 (R1): R37–R41. дои : 10.1093/hmg/ddab028 . ISSN 1460-2083 . ПМЦ 8223596 . PMID 33864377 .
- ^ Эрет С., Кейта С.О., Ньюман П. (3 декабря 2004 г.). «Истоки афроазиатского языка». Наука . 306 (5702): 1680, ответ автора 1680. doi : 10.1126/science.306.5702.1680c . ISSN 1095-9203 . ПМИД 15576591 . S2CID 8057990 .
- ^ Эрет С (1979). «О древности земледелия в Эфиопии». Журнал африканской истории . 20 (2): 161–177. дои : 10.1017/S002185370001700X . ISSN 0021-8537 . JSTOR 181512 . S2CID 162986221 .
- ^ Кларк Дж.Д., Брандт С.А. (1984). От охотников к фермерам: причины и последствия производства продуктов питания в Африке . Издательство Калифорнийского университета. стр. 11, 180. ISBN. 978-0-520-04574-3 .
- ^ Бултоса Дж., Тейлор-младший (1 января 2004 г.), «TEFF» , в Wrigley C (ред.), Энциклопедия зерновых наук , Оксфорд: Elsevier, стр. 281–290, doi : 10.1016/b0-12-765490-9/ 00172-5 , ISBN 978-0-12-765490-4 , получено 29 марта 2022 г.
- ^ Шлебуш CM, Якобссон М (31 августа 2018 г.). «Сказки о миграции, смешении и отборе людей в Африке» . Ежегодный обзор геномики и генетики человека . 19 : 405–428. doi : 10.1146/annurev-genom-083117-021759 . ISSN 1545-293X . ПМИД 29727585 . S2CID 19155657 .
- ^ Перейти обратно: а б с д и ж г час Шунеманн В.Дж., Пельтцер А., Велте Б., ван Пелт В.П., Молак М., Ван CC и др. (30 мая 2017 г.). «Геномы древнеегипетских мумий предполагают увеличение африканского происхождения к югу от Сахары в постримские периоды» . Природные коммуникации . 8 (1): 15694. Бибкод : 2017NatCo...815694S . дои : 10.1038/ncomms15694 . ISSN 2041-1723 . ПМЦ 5459999 . ПМИД 28556824 .
- ^ Молинаро Л., Монтинаро Ф., Йелмен Б., Марнетто Д., Бехар Д.М., Кивисилд Т. и др. (11 декабря 2019 г.). «Западноазиатские источники евразийского компонента у эфиопов: переоценка» . Научные отчеты . 9 (1): 18811. Бибкод : 2019НатСР...918811М . дои : 10.1038/s41598-019-55344-y . ISSN 2045-2322 . ПМК 6906521 . ПМИД 31827175 .
- ^ Гандини Ф., Ахилли А., Пала М., Боднер М., Брандини С., Хубер Г. и др. (5 мая 2016 г.). «Картирование расселения людей на Африканском Роге из рефугиумов Аравийского ледникового периода с использованием митогеномов» . Научные отчеты . 6 (1): 25472. Бибкод : 2016NatSR...625472G . дои : 10.1038/srep25472 . ISSN 2045-2322 . ПМЦ 4857117 . ПМИД 27146119 .
- ^ «Ошибка в отчете «Древний эфиопский геном обнаруживает обширную евразийскую примесь в Восточной Африке» (ранее озаглавленный «Древний эфиопский геном обнаруживает обширную евразийскую примесь на всем африканском континенте») М. Гальего Льоренте, Э. Р. Джонс, А. Эрикссон, В. Сиска , К.В. Артур, Дж.В. Артур, М.К. Кертис, Дж.Т. Сток, М. Колторти, П. Пьеруччини, С. Стреттон, Ф. Брок, Т. Хайэм, Ю. Парк, М. Хофрейтер, Д. Г. Брэдли, Дж. Бхак, Р. Пинхаси, А. Маника» . Наука . 351 (6275): ааф3945. 19 февраля 2016 г. doi : 10.1126/science.aaf3945 . ISSN 1095-9203 . ПМИД 26912899 .
- ^ Лопес С., Тарекегн А., Группа G, ван Дорп Л., Берд Н., Моррис С. и др. (11 июня 2021 г.). «Свидетельства взаимодействия генетики и культуры в Эфиопии» . Природные коммуникации . 12 (1): 3581. Бибкод : 2021NatCo..12.3581L . doi : 10.1038/s41467-021-23712-w . ISSN 2041-1723 . ПМК 8196081 . ПМИД 34117245 .
- ^ Дьюар Р.Э., Райт Х.Т. (1 декабря 1993 г.). «История культуры Мадагаскара». Журнал мировой предыстории . 7 (4): 417–466. дои : 10.1007/BF00997802 . hdl : 2027.42/45256 . S2CID 21753825 .
- ^ Берни Д.А., Берни Л.П., Годфри Л.Р., Юнгерс В.Л., Гудман С.М., Райт Х.Т. и др. (1 июля 2004 г.). «Хронология позднего доисторического Мадагаскара». Журнал эволюции человека . 47 (1–2): 25–63. Бибкод : 2004JHumE..47...25B . дои : 10.1016/j.jhevol.2004.05.005 . ПМИД 15288523 .
- ^ Пьеррон Д., Разафиндразака Х., Пагани Л., Рико Ф.К., Антао Т., Капредон М. и др. (январь 2014 г.). «Общегеномные свидетельства примеси австронезийцев и банту и культурной реверсии в группе охотников-собирателей на Мадагаскаре» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (3): 936–941. Бибкод : 2014PNAS..111..936P . дои : 10.1073/pnas.1321860111 . ПМЦ 3903192 . ПМИД 24395773 .
- ^ Перейти обратно: а б Кусума П., Брукато Н., Кокс М.П., Пьеррон Д., Разафиндразака Х., Аделаар А. и др. (май 2016 г.). «Контраст лингвистического и генетического происхождения азиатских исходных популяций малагасийцев» . Научные отчеты . 6 (1): 26066. Бибкод : 2016НатСР...626066К . дои : 10.1038/srep26066 . ПМК 4870696 . ПМИД 27188237 .
- ^ Перейти обратно: а б Хейске М., Альва О., Переда-Лот В., Ван Шалквик М., Радимилахи С., Летелье Т. и др. (апрель 2021 г.). «Генетические данные и исторические теории азиатского и африканского происхождения нынешнего малагасийского населения» . Молекулярная генетика человека . 30 (R1): R72–R78. дои : 10.1093/hmg/ddab018 . ПМИД 33481023 .
- ^ Николас Б. и др. (4 февраля 2019 г.). «Свидетельства австронезийского генетического происхождения в Восточной Африке и Южной Аравии: сложное распространение с Мадагаскара и Юго-Восточной Азии» . Геномная биология и эволюция, том 11, выпуск 3 .
- ^ Добон Б., Хасан Х.Ю., Лаюни Х., Луис П., Риканьо-Понсе И., Жернакова А. и др. (май 2015 г.). «Генетика популяций Восточной Африки: нило-сахарский компонент в африканском генетическом ландшафте » Научные отчеты 5 : 9996. Бибкод : 2015NatSR... 5E9996D дои : 10.1038/srep09996 . ПМЦ 4446898 . ПМИД 26017457 .
- ^ Арауна Л.Р., Комас Д. (2017), «Генетическая гетерогенность между берберами и арабами», eLS , John Wiley & Sons, Ltd, стр. 1–7, doi : 10.1002/9780470015902.a0027485 , ISBN 978-0-470-01590-2
- ^ Перейти обратно: а б Чен Л., Вольф А.Б., Фу В., Ли Л., Акей Дж.М. (февраль 2020 г.). «Выявление и интерпретация очевидного неандертальского происхождения у африканских людей» . Клетка . 180 (4): 677–687.e16. дои : 10.1016/j.cell.2020.01.012 . ПМИД 32004458 . S2CID 210955842 .
- ^ Гонсалес-Фортес Г., Тасси Ф., Тручки Е., Хеннебергер К., Пайманс Х.Л., Диес-дель-Молино Д. и др. (30 января 2019 г.). «Западный маршрут доисторической миграции человека из Африки на Пиренейский полуостров» . Труды Королевского общества B: Биологические науки . 286 (1895): 20182288. doi : 10.1098/rspb.2018.2288 . ISSN 0962-8452 . ПМК 6364581 . ПМИД 30963949 .
- ^ Мурджани П., Паттерсон Н., Хиршхорн Дж.Н., Кейнан А., Хао Л., Ацмон Г. и др. (апрель 2011 г.). «История потока африканских генов в южных европейцев, левантийцев и евреев» . ПЛОС Генетика . 7 (4): e1001373. дои : 10.1371/journal.pgen.1001373 . ISSN 1553-7404 . ПМК 3080861 . ПМИД 21533020 .
- ^ Botigué LR, Henn BM, Gravel S, Maples BK, Gignoux CR, Corona E и др. (16 июля 2013 г.). «Поток генов из Северной Африки способствует различному генетическому разнообразию человека в южной Европе» . Труды Национальной академии наук Соединенных Штатов Америки . 110 (29): 11791–11796. Бибкод : 2013PNAS..11011791B . дои : 10.1073/pnas.1306223110 . ISSN 0027-8424 . ПМК 3718088 . ПМИД 23733930 .
- ^ Аутон А., Брик К., Бойко А.Р., Лохмюллер К.Э., Новембре Дж., Рейнольдс А. и др. (май 2009 г.). «Глобальное распространение геномного разнообразия подчеркивает богатую и сложную историю континентальных человеческих популяций» . Геномные исследования . 19 (5): 795–803. дои : 10.1101/gr.088898.108 . ISSN 1088-9051 . ПМК 2675968 . ПМИД 19218534 .
- ^ Райх Д., Прайс А.Л., Паттерсон Н. (май 2008 г.). «Главнокомпонентный анализ генетических данных» . Природная генетика . 40 (5): 491–492. дои : 10.1038/ng0508-491 . ISSN 1546-1718 . ПМИД 18443580 . S2CID 34837532 .
- ^ Черный В, Фортес-Лима С, Триска П (26 апреля 2021 г.). «Демографическая история и динамика примесей в популяциях африканского Сахеля» . Молекулярная генетика человека . 30 (R1): R29–R36. дои : 10.1093/hmg/ddaa239 . ISSN 1460-2083 . ПМИД 33105478 .
- ^ Перейти обратно: а б с Хабер М., Меззавилла М., Бергстрем А., Прадо-Мартинес Дж., Халласт П., Саиф-Али Р. и др. (декабрь 2016 г.). «Генетическое разнообразие Чада раскрывает африканскую историю, отмеченную многочисленными голоценовыми евразийскими миграциями» (PDF) . Американский журнал генетики человека . 99 (6): 1316–1324. дои : 10.1016/j.ajhg.2016.10.012 . ПМК 5142112 . ПМИД 27889059 . S2CID 38169172 .
- ^ Кэмпбелл MC, Хирбо Дж.Б., Таунсенд Дж.П., Тишкофф С.А. (1 декабря 2014 г.). «Заселение африканского континента и диаспоры в новый мир» . Текущее мнение в области генетики и развития . 29 : 120–132. дои : 10.1016/j.где.2014.09.003 . ISSN 0959-437X . ПМЦ 4308437 . ПМИД 25461616 .
- ^ Перейти обратно: а б с д и ж г час Бергстрем А., Стрингер С., Хайдиньяк М., Шерри Э.М., Скоглунд П. (февраль 2021 г.). «Происхождение современного человеческого происхождения». Природа . 590 (7845): 229–237. Бибкод : 2021Natur.590..229B . дои : 10.1038/s41586-021-03244-5 . PMID 33568824 . S2CID 231883210 .
- ^ Перейти обратно: а б с д Гад Й.З., Хасан Н.А., Муса Д.М., Фуад Ф.А., Эль-Сайед С.Г., Абдельазим М.А. и др. (апрель 2021 г.). «Результаты древнего анализа ДНК египетских человеческих мумий: ключ к разгадке болезней и родства» . Молекулярная генетика человека . 30 (R1): R24–R28. дои : 10.1093/hmg/ddaa223 . ПМИД 33059357 .
- ^ Лорей О., Ратнаяке С., Базинет А.Л., Стоквелл Т.Б., Соммер Д.Д., Роланд Н. и др. (март 2018 г.). «Биологическое определение пола головы египетской мумии возрастом 4000 лет для оценки возможности восстановления ядерной ДНК из наиболее поврежденных и ограниченных судебно-медицинских образцов» . Гены . 9 (3): 135. doi : 10.3390/genes9030135 . ПМЦ 5867856 . ПМИД 29494531 .
- ^ Хавасс З., Исмаил С., Селим А., Салим С.Н., Фаталла Д., Васеф С. и др. (декабрь 2012 г.). «Возвращение к гаремному заговору и смерти Рамсеса III: антропологическое, судебно-медицинское, радиологическое и генетическое исследование». БМЖ . 345 : е8268. дои : 10.1136/bmj.e8268 . hdl : 10072/62081 . ПМИД 23247979 . S2CID 206896841 .
- ^ Гурдин Дж. П., Кейта С., Гурдин Дж. Л., Анселин А. «Древнеегипетские геномы северного Египта: дальнейшее обсуждение» . Природные коммуникации .
- ^ «Раскрыта шокирующая правда о смерти Такабути» . Проверено 29 января 2020 г.
- ^ Перейти обратно: а б Орас Э., Андерсон Дж., Тырв М., Вахур С., Раммо Р., Реммер С. и др. (16 января 2020 г.). «Мультидисциплинарное исследование двух мумий египетских детей, хранящихся в Художественном музее Тартуского университета, Эстония (Поздний/греко-римский период)» . ПЛОС ОДИН . 15 (1): e0227446. Бибкод : 2020PLoSO..1527446O . дои : 10.1371/journal.pone.0227446 . ISSN 1932-6203 . ПМК 6964855 . ПМИД 31945091 .
- ^ Вай С., Сарно С., Лари М., Луизелли Д., Манци Г., Галлинаро М. и др. (март 2019 г.). «Предковая митохондриальная линия N из неолитической «зеленой» Сахары» . Научные отчеты . 9 (1): 3530. Бибкод : 2019НатСР...9.3530В . дои : 10.1038/s41598-019-39802-1 . ПМК 6401177 . ПМИД 30837540 .
- ^ Перейти обратно: а б ван де Лоосдрехт М., Бузуггар А., Хамфри Л., Пост С., Бартон Н., Аксиму-Петри А. и др. (май 2018 г.). «Плейстоценовые геномы Северной Африки связывают человеческие популяции Ближнего Востока и Африки к югу от Сахары» . Наука . 360 (6388): 548–552. Бибкод : 2018Sci...360..548V . дои : 10.1126/science.aar8380 . ПМИД 29545507 . S2CID 206666517 .
- ^ Перейти обратно: а б Чон С. (2020). «Современные тенденции в изучении древней ДНК: помимо миграции людей в Европе и вокруг нее». Справочник по изучению мумий . Спрингер. стр. 1–16. дои : 10.1007/978-981-15-1614-6_10-1 . ISBN 978-981-15-1614-6 . OCLC 1182512815 . S2CID 226555687 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т Са Л и др. (16 августа 2022 г.). «Филогеография митохондриальных линий к югу от Сахары за пределами Африки подчеркивает роль голоценовых изменений климата и атлантической работорговли» . Международный журнал молекулярных наук . 23 (16): 9219. doi : 10.3390/ijms23169219 . ISSN 1661-6596 . OCLC 9627558751 . ПМЦ 9408831 . ПМИД 36012483 . S2CID 251653686 .
- ^ Осман М.М., Хасан Х.И., Эльнур М.А., Маккан Х., Гебремескель Э.И., Гайс Т. и др. «Вариации последовательностей митохондриального HVRI и всего митогенома отражают схожие сценарии генетической структуры и происхождения жителей северо-восточной Африки» (PDF) . Мета Джин . Архивировано из оригинала (PDF) 25 июня 2021 года . Проверено 12 мая 2021 г.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и Пфенниг А и др. (29 марта 2023 г.). «Эволюционная генетика и примесь в африканских популяциях» . Геномная биология и эволюция . 15 (4): evad054. дои : 10.1093/gbe/evad054 . OCLC 9817135458 . ПМЦ 10118306 . ПМИД 36987563 . S2CID 257803764 .
- ^ Перейти обратно: а б с д Приходова Е, Аустерлиц Ф, Цижкова М, Новачкова Й, Рико FX, Хофманова З и др. (ноябрь 2020 г.). «Сахельское скотоводство с точки зрения вариантов, связанных с персистенцией лактазы» (PDF) . Американский журнал физической антропологии . 173 (3): 423–436. дои : 10.1002/ajpa.24116 . ПМИД 32812238 . S2CID 221179656 .
- ^ Шерри Э (26 октября 2017 г.). «Археология каменного века Западной Африки» . Оксфордская исследовательская энциклопедия истории Африки . Издательство Оксфордского университета. п. 1. дои : 10.1093/акр/9780190277734.013.137 . ISBN 978-0-19-027773-4 . OCLC 1013546425 . S2CID 133758803 .
- ^ Батини С. и др. (сентябрь 2011 г.). «Дополнительные данные: признаки доземледельческих процессов заселения в Африке к югу от Сахары, выявленные филогеографией ранних линий Y-хромосомы» . Молекулярная биология и эволюция . 28 (9): 2603–2613. дои : 10.1093/molbev/msr089 . hdl : 10400.13/4486 . ISSN 0737-4038 . OCLC 748733133 . ПМИД 21478374 . S2CID 11190055 .
- ^ Хабер М. и др. (август 2019 г.). «Редкая глубоко укоренившаяся африканская гаплогруппа Y-хромосомы D0 и ее значение для расселения современного человека за пределы Африки» . Генетика . 212 (4): 1421–1428. дои : 10.1534/genetics.119.302368 . ISSN 0016-6731 . OCLC 8291848146 . ПМК 6707464 . ПМИД 31196864 . S2CID 189817793 .
- ^ Перейти обратно: а б с д и ж г час Шрайнер Д., Ротими К.Н. (апрель 2018 г.). «Гаплотипы, основанные на полногеномных последовательностях, раскрывают единое происхождение аллели серпа во время влажной фазы голоцена» . Американский журнал генетики человека . 102 (4): 547–556. дои : 10.1016/j.ajhg.2018.02.003 . ISSN 0002-9297 . OCLC 7353789016 . ПМЦ 5985360 . ПМИД 29526279 .
- ^ Тромбетта Б и др. (июнь 2015 г.). «Филогеографическое уточнение и крупномасштабное генотипирование гаплогруппы E Y-хромосомы человека дают новое представление о расселении ранних скотоводов на африканском континенте» . Геномная биология и эволюция . 7 (7): 1940–1950. дои : 10.1093/gbe/evv118 . ISSN 0016-6731 . OCLC 8291848146 . ПМЦ 4524485 . ПМИД 26108492 .
- ^ Перейти обратно: а б Миллер Э.Ф. и др. (август 2018 г.). «Глобальная демографическая история человеческих популяций, выведенная на основе целых митохондриальных геномов» . Королевское общество открытой науки . 5 (8): 180543. Бибкод : 2018RSOS....580543M . дои : 10.1098/rsos.180543 . OCLC 8582185081 . ПМК 6124094 . ПМИД 30225046 . S2CID 52289534 .
- ^ Фриги С. и др. (август 2010 г.). «Древняя локальная эволюция африканских гаплогрупп мтДНК в тунисском берберском населении» . Биология человека . 82 (4): 367–384. дои : 10.3378/027.082.0402 . ISSN 0018-7143 . JSTOR 41466688 . OCLC 4668535228 . ПМИД 21082907 . S2CID 27594333 .
- ^ Подгорна Э и др. (ноябрь 2013 г.). «Генетическое влияние населения бассейна озера Чад в Северной Африке, подтвержденное митохондриальным разнообразием и внутренними вариациями гаплогруппы L3e5». Анналы генетики человека . 77 (6): 513–523. дои : 10.1111/ahg.12040 . ISSN 0003-4800 . OCLC 6998015647 . ПМИД 25069842 . S2CID 24672148 .
- ^ Басби ГБ и др. (21 июня 2016 г.). «Примесь в Африке к югу от Сахары и внутри нее» . электронная жизнь . 5 : е15266. дои : 10.7554/eLife.15266 . ПМЦ 4915815 . ПМИД 27324836 . S2CID 6885967 .
- ^ Гурдасани Д. и др. (3 декабря 2014 г.). «Проект изменения африканского генома формирует медицинскую генетику в Африке» . Природа . 517 (7534): 327–332. Бибкод : 2015Natur.517..327G . дои : 10.1038/nature13997 . ISSN 0028-0836 . OCLC 9018039409 . ПМК 4297536 . ПМИД 25470054 . S2CID 4463627 .
- ^ Чоудхури А. и др. (октябрь 2020 г.). «Большие глубины африканских геномов определяют миграцию и здоровье людей» . Природа . 586 (7831): 741–748. Бибкод : 2020Natur.586..741C . дои : 10.1038/s41586-020-2859-7 . ISSN 0028-0836 . OCLC 8691160009 . ПМЦ 7759466 . ПМИД 33116287 . S2CID 226039427 .
- ^ Берд Н. и др. (29 марта 2023 г.). «Плотная выборка этнических групп в африканских странах раскрывает мелкомасштабную генетическую структуру и обширную историческую примесь» . Достижения науки . 9 (13): eabq2616. Бибкод : 2023SciA....9.2616B . дои : 10.1126/sciadv.abq2616 . ISSN 2375-2548 . OCLC 9819554112 . ПМЦ 10058250 . ПМИД 36989356 . S2CID 257834341 .
- ^ Фан С., Келли Д.Э., Белтрам М.Х., Хансен М.Е., Маллик С., Ранчиаро А. и др. (26 апреля 2019 г.). «Эволюционная история Африки выведена на основе данных последовательности всего генома 44 коренных африканских популяций» . Геномная биология . 20 (1): 82. дои : 10.1186/s13059-019-1679-2 . ISSN 1474-760X . ПМК 6485071 . ПМИД 31023338 .
- ^ Фан С. и др. (2 марта 2023 г.). «Полногеномное секвенирование раскрывает сложную демографическую историю африканского населения и признаки местной адаптации» (PDF) . Клетка . 186 (5): 923–939. дои : 10.1016/j.cell.2023.01.042 . ISSN 0092-8674 . OCLC 9791150466 . ПМЦ 10568978 . ПМИД 36868214 . S2CID 257286226 .
- ^ Перейти обратно: а б с д и ж Вонкам А., Адеймо А (8 марта 2023 г.). «Использование нашего общего африканского происхождения для понимания эволюции и здоровья человека» (PDF) . Клеточная геномика . 3 (3): 100278. doi : 10.1016/j.xgen.2023.100278 . ПМЦ 10025516 . ПМИД 36950382 . S2CID 257458855 .
- ^ Аманзугаген Н. и др. (14 октября 2016 г.). «Высокое древнее генетическое разнообразие человеческих вшей Pediculus humanus из Израиля открывает новые сведения о происхождении вшей группы B» . ПЛОС ОДИН . 11 (10): e0164659. Бибкод : 2016PLoSO..1164659A . дои : 10.1371/journal.pone.0164659 . ПМК 5065229 . ПМИД 27741281 . S2CID 14033603 .
- ^ Перейти обратно: а б с Эсо К., Вонкам А. (апрель 2021 г.). «Эволюционная история серповидноклеточной мутации: значение для глобальной генетической медицины» . Молекулярная генетика человека . 30 (Р1): Р119–Р128. дои : 10.1093/hmg/ddab004 . ПМК 8117455 . ПМИД 33461216 .
- ^ Лаваль Г., Пейрень С., Зидан Н., Харман С., Рено Ф., Патен Е. и др. (март 2019 г.). «Недавнее адаптивное приобретение африканскими охотниками-собирателями тропических лесов мутации серповидных клеток позднего плейстоцена предполагает наличие прошлых различий в подверженности малярии» . Американский журнал генетики человека . 104 (3): 553–561. дои : 10.1016/j.ajhg.2019.02.007 . ПМК 6407493 . ПМИД 30827499 .
- ^ Перейти обратно: а б с д Ясин Н.Т., Аль-Мамури Х.С., Хасан М.К. (январь 2020 г.). «Серповидные гаплотипы β-глобина среди пациентов с серповидноклеточной анемией в Басре, Ирак: поперечное исследование» . Иракский журнал гематологии . 9 (1): 23. дои : 10.4103/ijh.ijh_20_19 . S2CID 216082225 .
- ^ Перейти обратно: а б с д Стевердинг Д. (декабрь 2020 г.). «Распространение паразитов в результате миграционной деятельности человека» . Вирулентность . 11 (1): 1177–1191. дои : 10.1080/21505594.2020.1809963 . ПМЦ 7549983 . ПМИД 32862777 .
- ^ Агути I и др. (декабрь 2008 г.). «Молекулярная основа β-талассемии в Марокко: возможные причины молекулярной гетерогенности» (PDF) . Генетическое тестирование . 12 (4): 563–568. дои : 10.1089/gte.2008.0058 . ПМИД 18976160 . S2CID 46000591 .
- ^ Мурунга П. и др. (2018). «Разнообразие D-петли митохондриальной ДНК шлемованных цесарок в Кении и его влияние на функциональный полиморфизм гена HSP70» . БиоМед Исследования Интернэшнл . 2018 : 1–12. дои : 10.1155/2018/7314038 . OCLC 8754386965 . ПМК 6258102 . ПМИД 30539018 . S2CID 54463512 .
- ^ Гонсалес-Фортес Дж. и др. (2019). «Западный маршрут доисторической миграции человека из Африки на Пиренейский полуостров» . Труды Королевского общества B: Биологические науки . 286 (1895): 20182288. doi : 10.1098/rspb.2018.2288 . ПМК 6364581 . ПМИД 30963949 . S2CID 104296971 .
- ^ Липсон М., Рибо И., Маллик С., Роланд Н., Олальде И., Адамски Н. и др. (январь 2020 г.). «Древние западноафриканские собиратели в контексте истории населения Африки» . Природа . 577 (7792): 665–670. Бибкод : 2020Natur.577..665L . дои : 10.1038/s41586-020-1929-1 . ПМЦ 8386425 . ПМИД 31969706 . S2CID 210862788 .
- ^ Перейти обратно: а б с д и ж г час я дж Ван К., Гольдштейн С., Блисдейл М., Клист Б., Бостоен К., Баква-Луфу П. и др. (июнь 2020 г.). «Древние геномы раскрывают сложные закономерности перемещения, взаимодействия и замещения населения в странах Африки к югу от Сахары» . Достижения науки . 6 (24): eaaz0183. Бибкод : 2020SciA....6..183W . дои : 10.1126/sciadv.aaz0183 . ПМЦ 7292641 . ПМИД 32582847 .
- ^ Перейти обратно: а б с д и ж г час я дж Ван К., Гольдштейн С., Блисдейл М., Клист Б., Бостоен К., Баква-Луфу П. и др. (июнь 2020 г.). «Древние геномы раскрывают сложные закономерности перемещения, взаимодействия и замещения населения в странах Африки к югу от Сахары» . Достижения науки . 6 (24): eaaz0183. Бибкод : 2020SciA....6..183W . дои : 10.1126/sciadv.aaz0183 . ПМЦ 7292641 . ПМИД 32582847 .
- ^ Д'Атанасио Э. и др. (2018). «Заселение последней Зеленой Сахары, выявленное путем широкомасштабного повторения последовательности транссахарских отцов» . Геномная биология . 19 (1): 20. дои : 10.1186/s13059-018-1393-5 . ПМК 5809971 . ПМИД 29433568 .
Топология R1b-V88 указывает на миграцию из Европы в Африку. Действительно, наши данные предполагают европейское происхождение R1b-V88 около 12,3 тыс. лет назад.
- ^ Маркус Дж. и др. (2020). «Генетическая история от среднего неолита до представления на средиземноморском острове Сардиния» . Природные коммуникации . 11 (939): Дополнительный материал с.25. Бибкод : 2020NatCo..11..939M . дои : 10.1038/s41467-020-14523-6 . ПМК 7039977 . ПМИД 32094358 .
Два очень базальных R1b-V88 (с несколькими маркерами, все еще находящимися в наследственном состоянии) появляются у сербских охотников-собирателей возрастом около 9000 г. до н.э., что подтверждает мезолитическое происхождение клады R1b-V88 в этом обширном регионе или рядом с ним. Этот гаплотип, по-видимому, стал ассоциироваться с экспансией средиземноморского неолита… он обнаружен у человека, похороненного на стоянке Эльс-Трокс в Пиренеях (современный Арагон, Испания), датированного 5178–5066 гг. до н.э., и у одиннадцати древних сардинцев из нашей выборки. Интересно, что маркеры субклада R1b-V88 R1b-V2197, который в настоящее время обнаруживается у большинства африканских носителей R1b-V88, происходят только у особи Els Trocs и двух древних сардинских особей. Эта конфигурация предполагает, что ветвь V88 впервые появилась в Восточной Европе, смешалась с особями ранних европейских фермеров (EEF) (после предположительно примеси по признаку пола), а затем распространилась вместе с EEF в западное Средиземноморье. … Западно-евразийское происхождение R1b-V88 дополнительно подтверждается недавним филогенетическим анализом, который ставит современные сардинские гаплотипы-носители базальными по отношению к африканским гаплотипам R1b-V88. Предполагаемое время слияния сардинской и африканской ветвей, предполагаемое там, приходится на субплювиальный неолит («Зеленая Сахара», около 7000–3000 лет до нашей эры). Предыдущие наблюдения аутосомных следов голоценовой примеси с евразийцами для нескольких чадских популяций (Haber et al. 2016) обеспечивают дальнейшее подтверждение гипотезы о том, что по крайней мере некоторые предки EEF пересекли Сахару на юг.
- ^ Груни В. и др. (2019). «Анализ Y-хромосомы и фамилий для реконструкции прошлых популяционных структур: население Сардинии как тестовый пример» . Международный журнал молекулярных наук . 20 (5763): 5763. doi : 10.3390/ijms20225763 . ПМК 6888588 . ПМИД 31744094 .
Недавняя детальная реконструкция филогении гаплогруппы R1b-V88 показала, что редкие европейские линии R1b-V88 (R1b-M18 и R1b-V35) произошли от корня филогении гораздо раньше (около 12,34 тыс. лет назад), чем разделение африканских линий (7,85 ± 0,90 тысяч лет назад), что подтверждает происхождение R1b-V88 за пределами Африки и последующее распространение в Африке к югу от Сахары в период Последней Зеленой Сахары в среднем голоцене. Интересно, что недавние исследования древней ДНК выявили самые древние образцы R1b-V88 (датированные 11 и 9 тысячелетием) в Восточной Европе (Сербия и Украина соответственно) и более поздние образцы R1b-V88 (датированные 7 и 6 тысячелетиями) в Испании и Германии. , тем самым поддерживая европейское происхождение
- ^ Аллентофт М (2024). «Популяционная геномика послеледниковой Западной Евразии» . Природа . 625 (7994): Дополнительная информация, стр. 48. Бибкод : 2024Natur.625..301A . дои : 10.1038/s41586-023-06865-0 . ПМЦ 10781627 . ПМИД 38200295 .
Недавно зарегистрированные образцы, принадлежащие к гаплогруппе R1b, были распределены между двумя отдельными группами в зависимости от того, входили ли они в состав основного европейского субклада R1b1a1b (R1b-M269). Особи, помещенные за пределы этого субклада, были преимущественно выходцами из восточноевропейского мезолита и неолита и составляли часть редких ранних расходящихся линий R1b. Два украинца принадлежали к субкладу R1b1b (R1b-V88), обнаруженному среди современных жителей Центральной и Северной Африки, что еще раз подтверждает древнее восточноевропейское происхождение этой клады.
- ^ Шрайнер Д. и др. (2018). «Генетическая история Чада» . Американский журнал биологической антропологии . 167 (4): 804–812. дои : 10.1002/ajpa.23711 . ПМК 6240361 . ПМИД 30259956 .
- ^ Куличова И и др. (2017). «Внутренняя диверсификация гаплогрупп, не принадлежащих к югу от Сахары, в сахельском населении и распространение скотоводства за пределы Сахары» . Американский журнал биологической антропологии . 164 (2): 424–434. дои : 10.1002/ajpa.23285 . ПМИД 28736914 .
- ^ Перейти обратно: а б с Кинтана-Мурси Л., Куах Х., Хармант С., Лука Ф., Массонне Б., Патин Е. и др. (февраль 2008 г.). «Материнские следы глубокого общего происхождения и асимметричного потока генов между пигмеями-охотниками-собирателями и фермерами, говорящими на языке банту» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (5): 1596–1601. Бибкод : 2008PNAS..105.1596Q . дои : 10.1073/pnas.0711467105 . ПМК 2234190 . ПМИД 18216239 .
- ^ Сара А. Тишкофф и др. 2007, История говорящих по щелчку популяций Африки, основанная на генетических вариациях мтДНК и Y-хромосомы. Молекулярная биология и эволюция 2007 24(10):2180-2195.
- ^ Луис Кинтана-Мурси и др. Разнообразие мтДНК в Центральной Африке: от охотников-собирателей до земледелия. CNRS-Институт Пастера, Париж
- ^ Сильва М., Альшамали Ф., Силва П., Каррильо С., Мандлате Ф., Хесус Тровоада М. и др. (июль 2015 г.). «60 000 лет взаимодействия между Центральной и Восточной Африкой, задокументированные основной африканской митохондриальной гаплогруппой L2» . Научные отчеты . 5 : 12526. Бибкод : 2015NatSR...512526S . дои : 10.1038/srep12526 . ПМЦ 4515592 . ПМИД 26211407 .
- ^ Джарвис Дж.П., Шейнфельдт Л.Б., Сой С., Ламберт С., Омберг Л., Ферверда Б. и др. (2012). «Модели происхождения, признаки естественного отбора и генетическая связь с ростом западноафриканских пигмеев» . ПЛОС Генетика . 8 (4): e1002641. дои : 10.1371/journal.pgen.1002641 . ПМК 3343053 . ПМИД 22570615 .
- ^ Лопес Эрраес Д., Боше М., Тан К., Теунерт С., Пугач И., Ли Дж. и др. (ноябрь 2009 г.). «Генетическая изменчивость и недавний положительный отбор в человеческих популяциях по всему миру: данные почти 1 миллиона SNP» . ПЛОС ОДИН . 4 (11): е7888. Бибкод : 2009PLoSO...4.7888L . дои : 10.1371/journal.pone.0007888 . ПМЦ 2775638 . ПМИД 19924308 .
- ^ Бозцола М., Траваглино П., Марзилиано Н., Меацца С., Пагани С., Грассо М. и др. (ноябрь 2009 г.). «Низкий рост пигмеев связан с серьезной недостаточной экспрессией рецептора гормона роста». Молекулярная генетика и обмен веществ . 98 (3): 310–313. дои : 10.1016/j.ymgme.2009.05.009 . ПМИД 19541519 .
- ^ Льоренте М.Г., Джонс Э.Р., Эрикссон А., Сиска В., Артур К.В., Артур Дж.В. и др. (ноябрь 2015 г.). «Древний эфиопский геном обнаруживает обширную евразийскую примесь на всем африканском континенте». Наука . 350 (6262): 820–822. Бибкод : 2015Sci...350..820L . дои : 10.1126/science.aad2879 . hdl : 2318/1661894 . ПМИД 26449472 . S2CID 25743789 .
- ^ Льоренте М.Г., Джонс Э.Р., Эрикссон А., Сиска В., Артур К.В., Артур Дж.В. и др. (13 ноября 2015 г.). «Дополнительные материалы для древнего эфиопского генома показывают обширную евразийскую примесь в Восточной Африке» . Наука . 350 (6262): 820–822. Бибкод : 2015Sci...350..820L . дои : 10.1126/science.aad2879 . hdl : 2318/1661894 . ПМИД 26449472 . S2CID 25743789 .
- ^ Хелленталь Дж., Берд Н., Моррис С. (апрель 2021 г.). «Структура и особенности происхождения эфиопов в полногеномной аутосомной ДНК» . Молекулярная генетика человека . 30 (Р1): Р42–Р48. дои : 10.1093/hmg/ddab019 . ПМЦ 8242491 . ПМИД 33547782 .
- ^ Пагани Л., Кивисилд Т., Тарекегн А., Эконг Р., Пластер С., Гальего Ромеро I и др. (13 июля 2012 г.). «Генетическое разнообразие Эфиопии свидетельствует о языковой стратификации и сложном влиянии на генофонд Эфиопии» . Американский журнал генетики человека . 91 (1): 83–96. дои : 10.1016/j.ajhg.2012.05.015 . ISSN 1537-6605 . ПМК 3397267 . ПМИД 22726845 .
- ^ Перейти обратно: а б с д и Прендергаст М.Е., Липсон М., Савчук Е.А., Олальде И., Огола К.А., Роланд Н. и др. (июль 2019 г.). «Древняя ДНК показывает многоступенчатое распространение первых пастухов в Африке к югу от Сахары» . Наука . 365 (6448): 6275. Бибкод : 2019Sci...365.6275P . дои : 10.1126/science.aaw6275 . ПМК 6827346 . ПМИД 31147405 .
- ^ Перейти обратно: а б с д и Прендергаст М.Е., Липсон М., Савчук Е.А., Олальде И., Огола К.А., Роланд Н. и др. (5 июля 2019 г.). «Дополнительные материалы для древней ДНК показывают многоступенчатое распространение первых скотоводов в Африке к югу от Сахары» . Наука . 365 (6448): eaaw6275. Бибкод : 2019Sci...365.6275P . дои : 10.1126/science.aaw6275 . ПМК 6827346 . ПМИД 31147405 .
- ^ Перейти обратно: а б с Скоглунд П., Томпсон Дж.К., Прендергаст М.Е., Миттник А., Сирак К., Хайдиньяк М. и др. (сентябрь 2017 г.). «Реконструкция структуры доисторического населения Африки» . Клетка . 171 (1): 59–71.e21. дои : 10.1016/j.cell.2017.08.049 . ПМЦ 5679310 . ПМИД 28938123 .
- ^ Перейти обратно: а б с д Бриэль ES и др. (29 марта 2023 г.). «Дополнительные файлы данных о переплетении африканских и азиатских генетических корней средневековых народов побережья Суахили» . Природа . 615 (7954): 866–873. Бибкод : 2023Natur.615..866B . doi : 10.1038/s41586-023-05754-w . ISSN 0028-0836 . OCLC 9819552636 . ПМЦ 10060156 . ПМИД 36991187 . S2CID 250534036 .
- ^ Перейти обратно: а б с д Липсон М. и др. (23 февраля 2022 г.). «Таблица расширенных данных 1. Древние люди, проанализированные в этом исследовании: древняя ДНК и глубокая структура населения африканских собирателей к югу от Сахары» . Природа . 603 (7900): 290–296. Бибкод : 2022Natur.603..290L . дои : 10.1038/s41586-022-04430-9 . ISSN 0028-0836 . OCLC 9437356581 . ПМК 8907066 . ПМИД 35197631 . S2CID 247083477 .
- ^ Невес да Нова Фернандес VC. «Характеристика генетических маркеров высокого разрешения на Аравийском полуострове и Ближнем Востоке» (PDF) . Электронные диссертации «Белая роза» онлайн . Университет Лидса.
- ^ Перейти обратно: а б с Чоудхури А., Сенгупта Д., Рамзи М., Шлебуш К. (апрель 2021 г.). «Миграция и примесь говорящих на языке банту на юге Африки» . Молекулярная генетика человека . 30 (R1): R56–R63. дои : 10.1093/hmg/ddaa274 . ПМЦ 8117461 . ПМИД 33367711 .
- ^ Перейти обратно: а б Шлебуш К.М., Мальмстрем Х., Гюнтер Т., Сьёдин П., Коутиньо А., Эдлунд Х. и др. (ноябрь 2017 г.). «Древние геномы Южной Африки оценивают расхождение современного человека от 350 000 до 260 000 лет назад» . Наука . 358 (6363): 652–655. Бибкод : 2017Sci...358..652S . дои : 10.1126/science.aao6266 . ПМИД 28971970 . S2CID 206663925 .
- ^ Перейти обратно: а б Шлебуш К.М., Мальмстрем Х., Гюнтер Т., Сьёдин П., Коутиньо А., Эдлунд Х. и др. (3 ноября 2017 г.). «Дополнительные материалы к древним геномам Южной Африки оценивают дивергенцию современного человека от 350 000 до 260 000 лет назад» . Наука . 358 (6363): 652–655. Бибкод : 2017Sci...358..652S . дои : 10.1126/science.aao6266 . ПМИД 28971970 . S2CID 206663925 .
- ^ Перейти обратно: а б Найт А., Андерхилл П.А., Мортенсен Х.М., Животовский Л.А., Лин А.А., Хенн Б.М. и др. (март 2003 г.). «Расхождение африканской Y-хромосомы и мтДНК дает представление об истории щелкающих языков» . Современная биология . 13 (6): 464–473. Бибкод : 2003CBio...13..464K . дои : 10.1016/S0960-9822(03)00130-1 . ПМИД 12646128 . S2CID 52862939 .
- ^ Хаммер М.Ф., Карафет Т.М., Редд А.Дж., Джарджанази Х., Сантакьяра-Бенерекетти С., Судьялл Х. и др. (июль 2001 г.). «Иерархические закономерности глобального разнообразия Y-хромосом человека» . Молекулярная биология и эволюция . 18 (7): 1189–1203. doi : 10.1093/oxfordjournals.molbev.a003906 . ПМИД 11420360 .
- ^ Найду Т., Шлебуш К.М., Маккан Х., Патель П., Махабир Р., Эразмус Дж.К. и др. (сентябрь 2010 г.). «Разработка метода расширения с единой базой для определения гаплогрупп Y-хромосомы в популяциях Африки к югу от Сахары» . Исследовательская генетика . 1 (1): 6. дои : 10.1186/2041-2223-1-6 . ПМЦ 2988483 . ПМИД 21092339 .
- ^ Чен Ю.С., Олкерс А., Шурр Т.Г., Когельник А.М., Хуопонен К., Уоллес, округ Колумбия (апрель 2000 г.). «Вариации мтДНК южноафриканских кунгов и хве - и их генетические связи с другими африканскими популяциями» . Американский журнал генетики человека . 66 (4): 1362–1383. дои : 10.1086/302848 . ПМК 1288201 . ПМИД 10739760 .
- ^ Тишкофф С.А., Гондер М.К., Хенн Б.М., Мортенсен Х., Найт А., Жиньу С. и др. (октябрь 2007 г.). «История говорящих по щелчку популяций Африки, выведенная на основе генетических вариаций мтДНК и Y-хромосомы» . Молекулярная биология и эволюция . 24 (10): 2180–2195. дои : 10.1093/molbev/msm155 . ПМИД 17656633 .
- ^ Шлебуш CM, Найду Т, Судьял Х (ноябрь 2009 г.). «Мини-секвенирование SNaPshot для определения митохондриальных макрогаплогрупп, обнаруженных в Африке». Электрофорез . 30 (21): 3657–3664. дои : 10.1002/elps.200900197 . ПМИД 19810027 . S2CID 19515426 .
- ^ Хенн Б.М., Жинью Ч.Р., Джобин М., Гранка Дж.М., Макферсон Дж.М., Кидд Дж.М. и др. (март 2011 г.). «Геномное разнообразие охотников-собирателей предполагает южноафриканское происхождение современных людей» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (13). Национальная академия наук : 5154–5162. дои : 10.1073/pnas.1017511108 . ПМК 3069156 . ПМИД 21383195 .
- ^ Каплан М (2011). «Изучение генов бросает вызов происхождению человека в Восточной Африке» . Научный американец . Издательская группа «Природа» . Проверено 22 июня 2012 г.
- ^ Перейти обратно: а б с д Пфайффер С (2020). «Болезнь как фактор африканских археологических данных» . Африканский археологический обзор . 37 (3): 487–490. дои : 10.1007/s10437-020-09405-7 . ПМЦ 7445818 . ПМИД 32863518 .
- ^ Перейра Л., Мутеса Л., Тиндана П., Рамзи М. (май 2021 г.). «Генетическое разнообразие и адаптация Африки определяют повестку дня точной медицины». Обзоры природы. Генетика . 22 (5). Обзоры природы: 284–306. дои : 10.1038/s41576-020-00306-8 . ПМИД 33432191 . S2CID 231587564 .
- ^ Перейти обратно: а б Бентон М.Л., Абрахам А., ЛаБелла А.Л., Эббот П., Рокас А., Капра Дж.А. (май 2021 г.). «Влияние эволюционной истории на здоровье и болезни человека» . Обзоры природы. Генетика . 22 (5): 269–283. дои : 10.1038/s41576-020-00305-9 . ПМЦ 7787134 . ПМИД 33408383 .
- ^ Перейти обратно: а б с Прендергаст М.Э., Савчук Е.А., Сирак К.А. (19 октября 2022 г.). «Генетика и африканское прошлое» . Оксфордская исследовательская энциклопедия истории Африки . Издательство Оксфордского университета. ISBN 978-0-19-027773-4 . OCLC 1013546425 .
| Подтемы | |
|---|---|
| Генетическая история по регионам | |
| Популяционная генетика по группе |
|